0022-538X/09/$08.00⫹0 doi:10.1128/JVI.00715-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Respiratory Syncytial Virus Nonstructural Proteins Decrease Levels of
Multiple Members of the Cellular Interferon Pathways
䌤
Samer Swedan, Alla Musiyenko, and Sailen Barik*
Department of Biochemistry and Molecular Biology, University of South Alabama, College of Medicine, 307 University Blvd., Mobile, Alabama 36688-0002
Received 6 April 2009/Accepted 17 July 2009
Viruses of theParamyxoviridaefamily, such as the respiratory syncytial virus (RSV), suppress cellular innate immunity represented by type I interferon (IFN) for optimal growth in their hosts. The two uniquenons truc-tural (NS) proteins, NS1 and NS2, of RSV suppress IFN synthesis, as well as IFN function, but their exact targets are still uncharacterized. Here, we investigate if either or both of the NS proteins affect the steady-state levels of key members of the IFN pathway. We found that both NS1 and NS2 decreased the levels of TRAF3, a strategic integrator of multiple IFN-inducing signals, although NS1 was more efficient. Only NS1 reduced IKK, a key protein kinase that specifically phosphorylates and activates IFN regulatory factor 3. Loss of the TRAF3 and IKKproteins appeared to involve a nonproteasomal mechanism. Interestingly, NS2 modestly increased IKK levels. In the IFN response pathway, NS2 decreased the levels of STAT2, the essential transcription factor for IFN-inducible antiviral genes. Preliminary mapping revealed that the C-terminal 10 residues of NS1 were essential for reducing IKKlevels and the C-terminal 10 residues of NS2 were essential for increasing and reducing IKKand STAT2, respectively. In contrast, deletion of up to 20 residues of the C termini of NS1 and NS2 did not diminish their TRAF3-reducing activity. Coimmunoprecipitation studies revealed that NS1 and NS2 form a heterodimer. Clearly, the NS proteins of RSV, working individually and together, regulate key signaling molecules of both the IFN activation and response pathways.
Respiratory syncytial virus (RSV) is by far the most signifi-cant agent of pediatric respiratory disease for which no reliable antiviral or vaccine yet exists (45, 59). The lukewarm success of traditional active-immunization-based strategies has drawn fo-cus to cellular innate immunity, which acts as a broad antiviral defense. A major arm of innate antiviral immunity is the type I IFN family, represented by alpha interferon (IFN-␣) and IFN-(6, 46, 58). In counterdefense, however, members of the
Paramyxoviridaefamily have evolved accessory gene products
that neutralize or inhibit various steps of the IFN pathway, thus permitting robust virus growth (18, 27, 57, 71). It is now ap-preciated that a better understanding of the viral IFN suppres-sion mechanism(s) is essential for the prudent design of atten-uated vaccine strains and better overall therapy.
RSV encodes unique anti-IFN genes not found in any other member virus of this family. While other viruses generate IFN-suppressive proteins, mainly the V protein (1, 12, 18, 19, 21, 22, 27, 33, 50, 51, 57), from alternative translational reading frames in thePgene through “RNA editing,” the Pgene of RSV codes for the P protein only. Instead, the RSV genome contains two promoter-proximal genes that code for nonstruc-tural (NS) proteins, NS1 and NS2, which are so named because they are synthesized in RSV-infected cells but are not pack-aged in the mature virion structure. The predicted primary structures of the NS proteins do not share any significant homology with any other protein. Nonetheless, various lines of experimental evidence have clearly established a role for the
NS proteins in IFN suppression (7, 8, 15, 30, 40, 52, 56, 60, 61, 65, 66, 70, 72). Recombinant RSV strains were created in which either or both NS genes were deleted (⌬NS1, ⌬NS2,
⌬NS1/2) (30, 61, 65, 66, 70, 72). All RSV ⌬NS strains were found to be viable but replicated extremely poorly in IFN-proficient cells such as lung epithelial A549 cells; however, replication improved in IFN-negative Vero cells (8, 16, 29, 65, 66). The RSV⌬NS mutants, as well as wild-type RSV in which just NS1 expression was silenced by RNA interference, were highly attenuated in animals (8, 72, 76), consistent with ele-vated sensitivity to host IFN. In direct assays, RSV⌬NS acti-vated high levels of IFN and IFN-stimulated genes (ISG) (56, 60, 61).
In the generally accepted pathway of type I IFN induction by cytoplasmic RNA viruses such as RSV, a viral RNA species is recognized by the cell as a pathogen-associated molecular pat-tern. It binds to and activates cytoplasmic RNA sensors of the RIG-I (retinoic acid-inducible gene I) “RNA helicase” family (9, 24, 25, 34, 47, 57, 64, 73, 75). Collective evidence, including ours, has established that members of this family may display virus specificity and that RIG-I is essential for IFN induction in the RSV-infected cell (3, 20, 31, 33, 39, 41, 54, 74). Activation of the helicase domain of RIG-I allows it to interact with mitochondrial antiviral signaling (also called VISA, CARDIF, or IPS-1) through the CARD domains of the two proteins, which then leads to the activation of TRAF3 (9, 10, 20, 24, 47). In a parallel signaling pathway, the RSV fusion (F) protein or unidentified viral RNA can activate specific Toll-like receptors (TLR), such as TLR4 and TLR3, respectively, which also ul-timately activate TRAF3 (3, 36, 39). The TLR pathway ap-pears to be more relevant in dendritic cells, whereas the RIG-I pathway plays a greater role in lung epithelial cells, where RSV
* Corresponding author. Mailing address: Department of Biochem-istry and Molecular Biology, University of South Alabama, College of Medicine, 307 University Blvd., Mobile, AL 36688-0002. Phone: (251) 460-6860. Fax: (251) 460-6850. E-mail: [email protected].
䌤Published ahead of print on 22 July 2009.
9682
on November 8, 2019 by guest
http://jvi.asm.org/
infection occurs (8, 54, 72). Nonetheless, TRAF3 serves as the integration point of signals from both the RIG-I and TLR pathways, playing a strategically key role in IFN gene induc-tion. TRAF3-downstream Ser/Thr kinases of the IKK (inhibi-tor ofB kinase) family (13, 29, 67), namely, IKKε/TBK1, can assemble on different scaffolding proteins (viz., NAP1, TANK, and SINTBAD), leading to a kinase complex that phosphoryl-ates the C-terminal domain of IFN regulatory factor 3 (IRF3), leading to its activation and translocation from the cytoplasm to the nucleus (11, 13, 24, 26, 42). Phospho-IRF3, in cooper-ation with phospho-IRF7 and other transcription factors, such as NF-B and AP-1, then activates the type I IFN gene pro-moters (13, 26, 43).
In the IFN response pathway, IFN-␣and IFN-, synthesized as described above, bind to a common receptor (IFNAR), the cytoplasmic tail of which recruits kinases that phosphorylate the STAT (signal transducers and activators of transcription) proteins STAT1 and STAT2 (14, 55, 62). The active STAT proteins promote the assembly of the ISGF3 complex that translocates to the nucleus and binds to the IFN-stimulated response element found at the promoters of antiviral genes such as those for PKR, OAS, and Mx, leading to their tran-scriptional activation and the resultant virus-resistant state of the cell (58).
In theory, the RSV NS proteins can suppress IFN by inhib-iting one or multiple signaling factors of the IFN induction and/or response pathway described above. Regarding inhibi-tion of IFN-␣ and - gene induction, both NS1 and NS2 exerted their inhibitory activities individually, as well as coop-eratively (8, 60, 61). Both NS proteins prevented the phosphor-ylation of IRF-3, and the cooperative effect of the two proteins (NS1/2) in inhibiting IRF-3 activation was much greater than those of the individual proteins (60). The mechanism of this synergism remained unknown. As already said, NS proteins also inhibit the cell’s response to IFN, and STAT2 was shown to be a major target. While infection with wild-type RSV caused degradation of STAT2, ⌬NS1/2 RSV no longer de-creased STAT2 levels or IFN responsiveness (40, 52). Studies of NS proteins have been hindered due to the difficulties of recombinant expression; recently, this problem has been solved through the use of codon-optimized synthetic NS genes (40). Recombinant expression of the human RSV NS proteins caused a marked decrease in STAT2 levels and the consequent downstream IFN-␣/ response. The relative contributions of NS1 and NS2 to this process, however, are still an area of active research. Lo et al. (40) showed that STAT2 was reduced by recombinant NS2 alone and that recombinant NS1 had no effect. In a complementary study, Ramaswamy et al. (52) reached similar conclusions from virus-infected cells, demon-strating that infection with ⌬NS1 mutant or wild-type RSV degraded STAT2 to similar extents, whereas the degradation was significantly reduced when cells were infected with⌬NS2 RSV or when NS2 in the wild-type RSV-infected cell was silenced by a specific anti-NS2 small interfering RNA. The degradation of STAT2 by NS2 likely occurred via a proteaso-mal mechanism, as it could be inhibited by MG132, a classic proteasome inhibitor. In apparent contrast, Elliott et al. (15) recently used a combination of recombinant NS proteins, over-expressed recombinant proteins of the ubiquitin proteasome pathway, and small interfering RNA-mediated knockdown to
show that NS1 alone could degrade STAT2 by using elongin C and cullin 2, apparently acting as an E3 ligase. Interestingly, they also found that both NS1 and NS2 promoted ubiquitina-tion of STAT2, suggesting that NS2 must somehow contribute to this E3 ligase activity (15).
In this communication, we have investigated whether the NS1 and NS2 proteins lead to the reduction of other key members of the IFN pathway. Indeed, we show that NS1 and NS2 variously affect the levels of TRAF3 and IKKεby using potentially different but nonproteasomal mechanisms. We also find that NS1 and NS2 form a complex, thus providing a frame-work for their individual, as well as joint, roles in suppressing the various steps of IFN induction and IFN response.
MATERIALS AND METHODS
Cell culture and treatments.The human alveolar epithelial A549 cell line (ATCC CCL-185; American Type Culture Collection, Manassas, VA) and the African green monkey Vero cell line (ATCC CCL-81; American Type Culture Collection, Manassas, VA) were cultured in monolayers by using standard Ea-gle’s minimal essential medium containingL-glutamine, 10% heat-inactivated fetal bovine serum, 100 U/ml penicillin, and 100g/ml streptomycin. In exper-iments requiring activation of IFN signaling, the cells were treated with recom-binant IFN-␣(R&D Systems) at 500 U/ml (final concentration) 30 min prior to cell harvesting. Where mentioned, the cell-permeating proteasomal inhibitors MG132 (Z-Leu-Leu-Leu-CHO) and lactacystin (Sigma) were used at 10M (final concentration) 6 h prior to cell harvesting.
Virus growth.The wild-type RSV Long strain of the A serotype was grown on HEp-2 cell monolayers and the NS deletion mutants were grown on Vero cells by using standard cell culture conditions as described previously (4, 5). The virus was purified on a sucrose density gradient as previously described (68) and stored frozen in small portions at⫺80°C. For experiments requiring infection by RSV strains, a multiplicity of infection of 3 was used.
Plasmid constructs.Codon-optimized versions of the RSV genes for NS1 and NS2 have been described before (40) and were synthesized by using overlapping primers and PCR technology. To generate NS-specific antibodies, the NS genes were first cloned into the pET-15b prokaryotic expression vector (Novagen) to add a six-His tag at the N terminus of each protein. The recombinants were induced by isopropyl--D-thiogalactopyranoside (IPTG) in Escherichia coli BL21(DE3), purified by standard Ni2⫹-agarose affinity chromatography, and used as the antigen to raise polyclonal antisera in rats (Cocalico Biologicals, Reamstown, PA). For eukaryotic expression, the NS genes were cloned into the pCAGGS expression vector (44) such that a FLAG tag (DYKDDDDKP) or a hemagglutinin (HA) tag (YPYDVPD) was added to the N terminus of each protein. C-terminal mutant forms of NS1 and NS2 lacking 10, 20, or 30 amino acids were generated by simply introducing stop codons at the appropriate places by PCR. All clones, including the mutant forms, were verified by DNA sequenc-ing (Laragen, CA). The Myc-IKKε(in pcDNA3.1-myc) and FLAG-TRAF3 (pSG5-FLAG-TRAF3) expression clones were kind gifts from Ganes C. Sen (Lerner Research Institute, Cleveland Clinic, Cleveland, OH) and Paula Norris and Carl Ware (La Jolla Institute for Allergy and Immunology), respectively. Reporter plasmids in which Luc expression is under the control of the IFN- promoter or the IFN-responsive gene ISG54 promoter have been described before (53) and were kindly provided by Chris Basler (Mount Sinai School of Medicine, NY). The pCMV-Renilla Luc plasmid (48), constitutively expressing Renilla luciferase, was a kind gift from Ramesh Pillai (EMBL, Grenoble, France).
Transfections.Lipofectamine 2000 transfection reagent (Invitrogen) was used to transfect A549 cells according to the manufacturer’s protocol. All transfec-tions were carried out at about 90% confluence of the monolayer in a six-well plate configuration (10 cm2per well), and DNA amounts were optimized to
allow maximal gene expression and the least cytopathic effect. Before transfec-tion, the antibiotic-containing medium was removed, cells were washed twice with phosphate-buffered saline (PBS), and then antibiotic-free medium was added. All transfections were transient (i.e., for 24 h).
Immunoblotting (IB or Western blotting) and immunoprecipitation (IP). Upon completion of the experiments (i.e., 24 h posttransfection), the cell culture medium was removed and the cells were washed twice with PBS. For IB analysis, cells were scraped and lysed directly in 1⫻Laemmli sample buffer, aided by sonication. All steps were done on ice. Samples were boiled for 5 min and used
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9683
on November 8, 2019 by guest
http://jvi.asm.org/
for sodium dodecyl sulfate-polyacrylamide gel electrophoresis as described pre-viously (5). Mouse anti-FLAG antibody (MAB3118; Millipore) was used to detect FLAG-NS proteins or FLAG-TRAF3. Mouse anti-myc (R950-25; Invitro-gen), rabbit anti-actin (A2066; Sigma), rabbit anti-STAT2 (sc476; Santa Cruz), and goat anti-RSV (AB1128; Chemicon) antibodies were used to detect myc-IKKε, actin, STAT2, and RSV proteins, respectively. The corresponding second-ary antibodies used, conjugated to horseradish peroxidase, were sheep anti-mouse (931V; ECL), donkey anti-rabbit (934V; ECL), and rabbit anti-goat (A5420; Sigma) antibodies.
The Western blotting bands were developed by ECL using Pierce reagents (Pierce, Rockford, IL), and densitometric analysis was performed with Fuji Image Gauge V4.0 software. Band intensities of the experimental proteins were normalized to that of actin from the same membrane.
IP was done by standard procedures as described before (35).
Immunofluorescence (IF).A549 cells were grown on 18-mm round coverslips placed in a 12-well plate. While still on the plate, cells were rinsed twice with cold PBS, fixed with ice-cold 10% trichloroacetic acid for 15 min, and then fixed for 3 min each in ice-cold 70%, 90%, and 100% ethanol, in that order. The cells were next rinsed twice with cold PBS for 5 min each time, permeabilized with cold 0.2% Triton X-100 in PBS for 10 min, rinsed three times with PBS for 5 min each time, and incubated for 2 h with a primary antibody solution made in PBS. The unbound antibody was removed by washing three times with PBS, for 5 min each time, and the cells were incubated with an Alexa Fluor-conjugated secondary antibody in PBS for 1.5 h in the dark. The cells were finally rinsed four times with PBS and mounted by using the Gold Reagent (Invitrogen) according to manu-facturer’s instructions. Mouse anti-FLAG (MAB3118; Millipore) and Alexa Fluor-conjugated 488 donkey anti-mouse (A21202; Invitrogen) antibodies were used. Images were acquired in a Nikon TE2000E2 Perfect Focus fluorescence imaging microscope equipped with NIS-Elements 2.3 software.
RESULTS
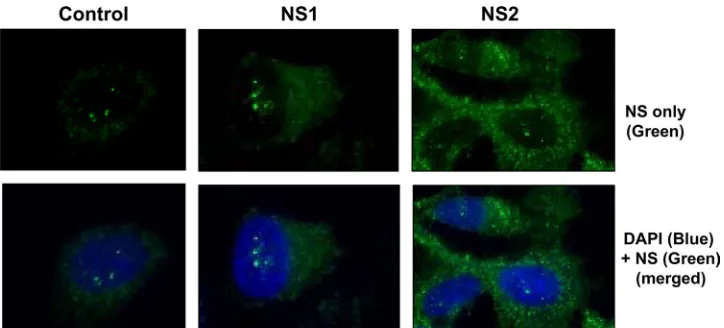
Recombinant expression of RSV NS proteins. We cloned the codon-optimized versions of both NS genes (40) under the control of the constitutive chicken actin promoter in the pCAGGS vector with an N-terminal FLAG tag. Transient transfection of A549 cells and IF staining with anti-FLAG antibody revealed the expression of both proteins (Fig. 1), which were primarily cytoplasmic, consistent with previous findings (60) and with their ability to suppress members of the IFN family that are also located in the cytoplasm. By IF (Fig. 1), as well as IB (for example, Fig. 2), NS2 expression from the recombinant clone was found to be more robust than that of NS1. We also tested our polyclonal rat NS antibodies, raised against bacterial NS proteins (see Materials and Methods), in
three kinds of applications, which revealed a profile of success and failure: whereas both antibodies reacted in IF, only the anti-NS1 antibody was reactive for IB (the anti-NS2 antibody failed) and only the anti-NS2 antibody was reactive for IP (the anti-NS1 antibody failed). Based on these results, we used these antibodies in specific applications only.
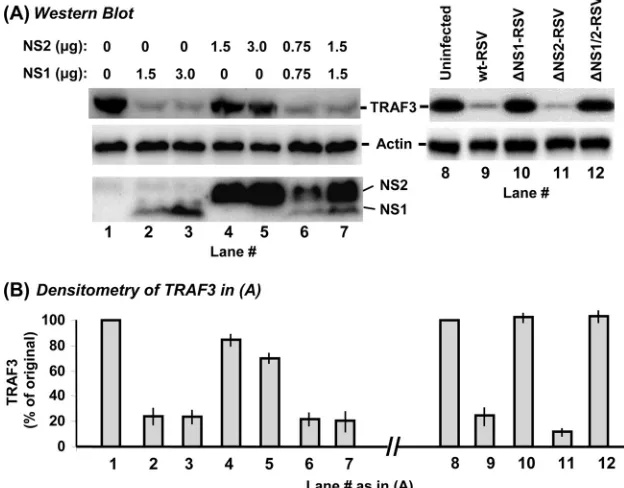
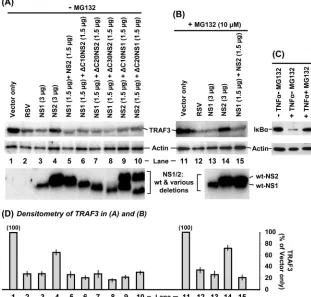
NS1 efficiently and NS2 weakly decreases TRAF3.We first tested if NS proteins affect TRAF3 levels. A549 cells were transiently transfected with FLAG-TRAF3 and FLAG-NS plasmids, and the levels of all recombinant proteins were de-termined by IB. Two different amounts of each NS plasmid and their combinations were tested. Note that, as it appeared in IF detection (Fig. 1), IB (Fig. 2A, left panel) also revealed that recombinant NS1 is expressed poorly compared to NS2, even when 3g of the NS1 plasmid (lane 3) is compared with 1.5g of the NS2 plasmid (lane 4). The reason for this dissimilar expression is unknown, but it could not be due to unequal detection efficiency because the same FLAG antibody was used for both. In either case, we could not enhance NS1 ex-pression further due to the general cytopathic effect of larger amounts of transfected DNA. Nonetheless, even the smaller amounts of NS1 caused a much greater reduction of TRAF3, compared to the relatively minor effect of much larger amounts of NS2 (Fig. 2). When NS1 and NS2 were expressed together, the NS1 effect was clearly dominant, as judged by the strong reduction of TRAF3, similar to what was achieved by NS1 alone.
These results were then verified in RSV infection; TRAF3 levels were determined in A549 cells infected with wild-type RSV or recombinant RSV lacking one or both NS genes. The results (Fig. 2A, right panel), indeed, showed extensive reduc-tion of TRAF3 in wild-type (lane 9) and⌬NS2 (lane 11) RSV infections, where the NS1 gene was intact. In contrast, normal levels of TRAF3 prevailed upon infection with⌬NS1 (lane 10) and the double-deletion⌬NS1/2 virus (lane 12) when the NS1 gene was lost.
[image:3.585.112.474.67.231.2]As one of our future goals is to use the recombinant NS clones to map the functional domains, we wanted to ensure that the functions of the recombinants are discernible and
FIG. 1. Cytoplasmic location of recombinantly expressed NS proteins. A549 cells were transiently transfected with pCAGGS-FLAG-NS1 and -NS2 plasmids or with the vector alone (control) as indicated at the top. The proteins were immunostained with FLAG antibody (green) and the nuclei were stained with 4⬘,6-diamidino-2-phenylindole (DAPI; blue) as described in Materials and Methods.
on November 8, 2019 by guest
http://jvi.asm.org/
reproducible in the RSV-infected cellular environment. To this end, we retested the TRAF3-reducing activities of the recom-binant NS1 and NS2 proteins in cells infected with ⌬NS1/2 RSV, which should provide all viral gene functions except NS1 and NS2. The results (Fig. 3) not only reconfirmed the relative TRAF3-reducing activities of recombinant NS1 and NS2 in the uninfected cell (lanes 2, 4, and 6) but also showed that they were reproducible in RSV-infected cytoplasm as well (corre-sponding lanes 3, 5, and 7, respectively).
Together, these results clearly document a striking loss of TRAF3 in the RSV-infected cell, caused primarily by the viral NS1 protein.
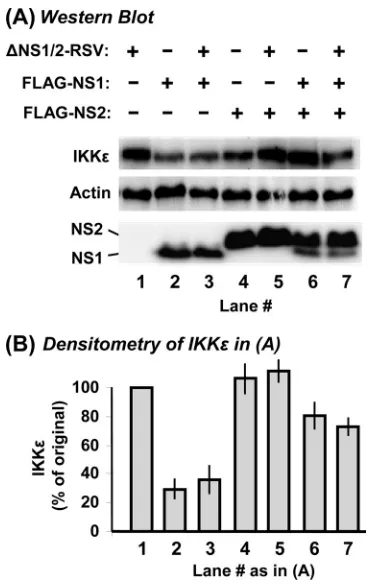
NS1, but not NS2, decreases IKK. We then conducted similar studies with the downstream kinase IKKε, which is normally responsible for activation of the IRFs. A549 cells were transiently transfected with Myc-IKKε and FLAG-NS plasmids, and the levels of all recombinant proteins were de-termined by IB. The results (Fig. 4) revealed a pattern that is generally similar to that of TRAF3 described above. Recom-binant NS1 protein (lanes 2 and 3), in spite of its poorer expression than that of NS2, significantly lowered IKKεlevels. Expression of NS2 by itself (lanes 4 and 5) failed to decrease IKKε, even when it is expressed two- to fourfold more abun-dantly than NS1. In excessively large amounts (lane 5), NS2 seemed to elevate IKKεlevels (an average of 22% increase by 3g NS2 plasmid), whose physiological relevance is question-able. Nonetheless, since both proteins are expressed in RSV infection, we examined the effect of joint expression of the two proteins on IKKε levels. Even with a skewed expression of
more NS2 than NS1 (lanes 6 and 7), there was a net reduction of IKKεcompared to that in untransfected A549 cells.
To compare the recombinant NS results with RSV infection, we measured IKKεlevels in A549 cells infected with⌬NS RSV strains. As shown in Fig. 4A (right side), IKKε levels were strongly reduced whenever the NS1 gene was intact (RSV-wt and RSV-⌬NS2, lanes 9 and 11) but remained unchanged when the NS1 gene was missing (the⌬NS1 and double mu-tants, lanes 10 and 12).
As for TRAF3, we wanted to make certain that the IKKε regulatory functions of the recombinants are reproducible in the RSV-infected cellular environment provided by ⌬NS1/2 RSV infection. These results (Fig. 5) reconfirmed the contrast-ing effects of recombinant NS1 and NS2 on IKKεand showed that the results were identical without and with⌬NS1/2 RSV infection (compare lanes 2, 4, and 6 with lanes 3, 5, and 7, respectively).
We conclude that NS1, but not NS2, plays a cardinal role in the reduction of IKKεlevels in RSV-infected cells.
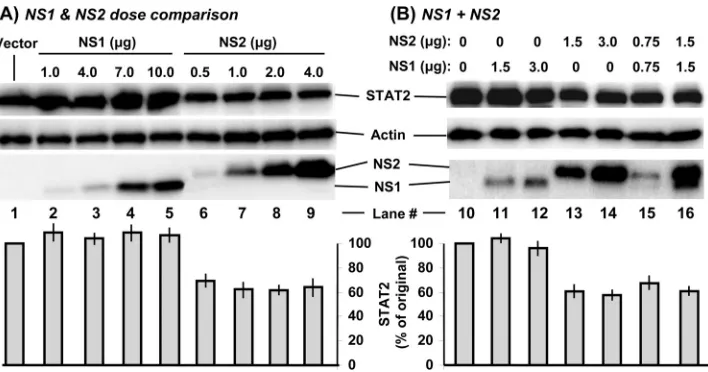
[image:4.585.134.446.72.316.2]Recombinant NS2 decreases STAT2 levels. As mentioned, the relative contributions of NS1 and NS2 to the loss of STAT2 remain equivocal, as either protein has been shown to promote STAT2 degradation by diverse approaches (15, 40, 52). We wanted to revisit this issue as the availability of recombinant NS1 and NS2 allowed us to perform a head-on comparison. First, in recognition of the unequal expression of the two NS proteins, we transfected each plasmid in a range of amounts, followed by quantification of the endogenous STAT2 levels. The results (Fig. 6A) unequivocally show that even the smallest
FIG. 2. Reduction of TRAF3 levels by NS1. A549 cells in 10-cm2wells were transfected with 4g FLAG-TRAF3 plasmid. In addition, they
were transfected with the indicated amounts (in micrograms) of pCAGGS-FLAG-NS plasmids (left) or infected with wild-type (wt) or NS deletion-containing RSV (right) as indicated. In transfection experiments (left), the empty pCAGGS vector was used wherever needed, to keep the amount of DNA constant at 7g in every well. Cells were processed 24 h later for IB to detect the proteins as shown in panel A, with specific antibodies described in Materials and Methods. The TRAF3 band intensities were normalized against actin and plotted (B) as a percentage of the respective control lanes (lane 1 for the left side of the panel, lane 8 for the right side of the panel). Average values of three experiments and standard error bars are presented.
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9685
on November 8, 2019 by guest
http://jvi.asm.org/
amounts of expressed NS2 (e.g., 0.5 and 1 g of the NS2 plasmid, respectively, in lanes 6 and 7) were quite effective in decreasing STAT2, whereas much larger amounts of the NS1 plasmid, expressing comparable amounts of protein (lanes 4 and 5), were ineffective.
We then tested if coexpression of NS1 would have any ad-ditive or negative effect over NS2’s ability to degrade STAT2. Since recombinant protein expression may vary from experi-ment to experiexperi-ment, we used two different amounts of the NS1 and NS2 plasmids. The results (Fig. 6B) revealed no regulation of NS2 activity by NS1. For example, the extent of the decrease in STAT2 levels (Fig. 6B) caused by 1.5g NS2 plasmid was essentially similar with (lane 16) or without (lane 13) 1.5g of the NS1 plasmid. We conclude that recombinant NS2 itself possesses STAT2-decreasing activity, confirming previous con-clusions (40, 52).
Deletion mutagenesis of NS proteins.Although NS proteins are important and unique IFN antagonists, there has been no attempt to map their functional domains. Interestingly,
not-withstanding their overall dissimilarity, both NS proteins con-tain the same DLNP tetramer at the very C-terminal end that is preceded by an aromatic residue (F/Y) (see Fig. 13). In fact, a rabbit antiserum directed against the 10-amino-acid C-ter-minal peptide of the NS2 protein that included the common tetrapeptide sequence efficiently recognized both NS1 and NS2 in Western blotting (2).
The availability of recombinant NS clones allowed us to attempt to map the functional domain(s) in NS proteins. In this preliminary effort, we created incremental deletions of 10-amino-acid segments from the C-terminal end. Thus, three mutant forms of each protein, designated ⌬C10, ⌬C20, and
⌬C30, were created by deleting 10, 20, and 30 amino acid residues, respectively. When tested in transient transfection with the FLAG antibody, all of the clones were expressed well (Fig. 7), the sole exception being⌬C30NS1, which was there-fore not included in our subsequent studies. The others were tested for loss of the cognate activities discovered as described above.
The C termini of the NS proteins play no role in decreasing TRAF3.Since both NS proteins decreased TRAF3 levels, we presumed that their relatively conserved C termini may be important in this process. To test this, we asked if any of our C-terminal deletion NS mutant proteins are defective in TRAF3 reduction. To approximate virus infection whereby both NS proteins are produced, we expressed each deletion mutant form of one NS protein in combination with the wild-type protein of the other NS and measured the TRAF3 levels as before. Unexpectedly, the results (Fig. 8) showed that all of the deletion-containing proteins, including those that lost as much as 30 residues of NS2 (lane 8) and 20 residues of NS1 (lane 10), retained full TRAF3-reducing activity.
As mentioned before, both NS1 and NS2 have been re-ported to degrade STAT2 by a proteasomal mechanism. We therefore investigated whether the loss of TRAF3 was also occurring through proteasomal degradation by testing if MG132 and lactacystin, two well-known proteasome inhibitors, could restore TRAF3 levels in NS-expressing cells. As a posi-tive control, we used IB␣, which is promptly degraded by cellular proteasome upon treatment of the cells with tumor necrosis factor alpha (TNF-␣). Unexpectedly, MG132 did not inhibit either NS protein’s ability to reduce TRAF3 (Fig. 8B); compare, for example, lane 15 (with MG132) and lane 5 (with-out MG132), having essentially the same levels of TRAF3, which is obvious in the densitometric scan (Fig. 8D). The results obtained with lactacystin were identical to those ob-tained with MG132 and hence are not shown. In the contrast-ing control, the degradation of IB␣ in response to TNF-␣ treatment was indeed inhibited by the same concentration of MG132 in the same type of cell (A549) in a parallel experiment (Fig. 8C). We conclude that NS1 and NS2 reduce TRAF3 levels through a novel nonproteasomal mechanism that also does not involve their common C termini.
The C termini of the NS proteins are important for regu-lating IKK.Recall that NS1 promotes loss of IKKε; in con-trast, NS2 actually promoted a modest increase (Fig. 9). In an attempt to map the domains of the two NS proteins important for these respective roles, we tested the C-terminal deletion mutant proteins. These experiments were performed as for TRAF3; i.e., deletion mutant forms of NS1 were combined
FIG. 3. Reduction of TRAF3 levels by recombinant NS1 in the RSV-infected cell environment. (A) A549 cells in 10-cm2wells were
first infected with dual NS deletion-containing RSV. The medium was removed 3 h later, and the cell monolayers were rinsed with PBS. Fresh medium was then added, and the cells were transfected with 4g FLAG-TRAF3 plasmid and the indicated pCAGGS-FLAG-NS plas-mids (3g for the single-NS plasmid, 1.5g each for dual transfec-tion). Cells were processed 24 h later for IB to detect the proteins as shown, with the specific antibodies described in Materials and Meth-ods. (B) The TRAF3 band intensities were normalized against actin and plotted as percentages of the no-NS control (lane 1). The mean⫾ the standard error of the mean of three experiments are presented.
on November 8, 2019 by guest
http://jvi.asm.org/
with wild-type NS2 and vice versa. The results (Fig. 9A and B) revealed that loss of as little as 10 amino acids from the C termini nearly completely abrogated both the IKKε-reducing activity of NS1 and the IKKε-increasing activity of NS2. For example, lane 4 represents the joint expression of two full-length NS proteins, showing about 70% of the basal levels of IKKε(Fig. 9C, lane 1), which appears to be an integration of the effects of NS1 (lane 2, 30%) and NS2 (lane 3, 120%). However, when the wild-type NS2 protein in the mixture was replaced with⌬C10NS2 (lane 5), the IKKεlevel dropped from 70% to about 30%, equal to the level seen with NS1 alone (lane 2), indicating loss of the stimulatory effect of NS2. Sim-ilarly, when⌬C10NS1 replaced full-length NS1 (lane 8), the IKKεlevels increased to 110%, approaching the NS2-only lev-els (lane 3, 120%). For both NS1 and NS2, larger deletions (⌬C20,⌬C30) showed no further decrease in IKKεregulatory activity; e.g., lanes 5 to 9 (NS2,⌬C10 to ⌬30) had identical values and so did lanes 8 and 9 (NS1,⌬C10 to⌬20). Finally, NS1-mediated loss of IKKε levels could not be inhibited by MG132 (Fig. 9B, compare lanes 2 and 12 or lanes 4 and 14). Use of just the deletion mutant protein⌬C10NS1 in the ab-sence of NS2 led to the same conclusion when appropriately normalized against actin (Fig. 9C, lanes 10 and 15). Proof that MG132 was functional came from the demonstration that it did inhibit TNF-␣-induced proteasomal degradation of IB␣, as shown previously in Fig. 8C. Moreover, in both Fig. 8 and 9, we noticed higher levels of NS proteins in the MG132-treated samples than in otherwise identical untreated ones (e.g., com-pare lanes 13 and 3 or lanes 14 and 4 in the bottom of Fig. 9A and B), suggesting protection of these highly unstable proteins (5, 15, 17, 39), likely from proteasomes.
FIG. 4. Reduction of IKKεlevels by NS1. Experiments were performed essentially as described in the legend to Fig. 2, by substituting 2g of the myc-IKKεplasmid for the FLAG-TRAF3 plasmid. The total amount of transfected DNA was kept constant at 5g by using the empty pCAGGS vector wherever needed. wt, wild type.
FIG. 5. Reduction of IKKεlevels by recombinant NS1 in the RSV-infected cell environment. These experiments were performed essen-tially as described in the legend to Fig. 3, by substituting 2g of the myc-IKKεplasmid for the FLAG-TRAF3 plasmid. The total amount of transfected DNA was kept constant at 5g by using the empty pCAGGS vector wherever needed.
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9687
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.70.255.378.670.2]Together, these results lead to the interesting conclusions that the C terminus of NS1 is involved in strongly lowering IKKεlevels by a nonproteasomal mechanism and that the C terminus of NS2 is also involved in regulating IKKε but its effect is weak and opposite, actually causing a small increase in IKKεlevels. As mentioned before, infection with RSV with NS deleted leads to much greater activation of IRF3 than does infection with wild-type RSV (60), and subsequent studies showed that IRF3 activation by wild-type RSV in fact requires IKKε(28). Thus, reduction of IKKεby NS1, as shown here, is particularly remarkable since it plays the crucial role of atten-uating the signal flowing to IRF3 via the IKKεaxis.
The C terminus of NS2 is important for decreasing STAT2.
Having confirmed that recombinant NS2 indeed decreases STAT2 (Fig. 6), we proceeded to determine where this func-tion maps by using the C-terminal delefunc-tions. The results (Fig. 10) first confirmed the degradation of STAT2 by NS2 (lane 3) but no effect of NS1 either alone (lane 2) or when coexpressed with NS2 (lane 4). Analysis of the deletions showed that loss of the last 10 C-terminal residues of NS2 resulted in total loss of STAT2-decreasing activity (lane 7). Further deletion of NS2 or any deletion of NS1 showed no greater loss or gain of
activity. Clearly, the C-terminal 10 residues of NS2 play an essential role in STAT2 reduction.
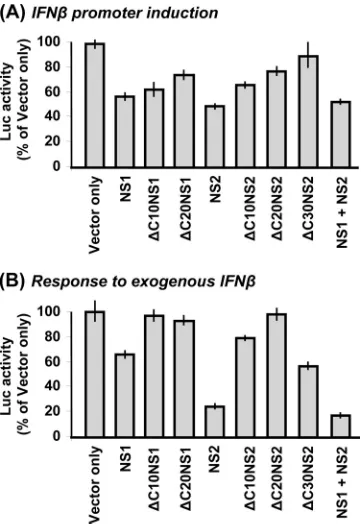
Correlation of NS domain analysis with IFN induction and IFN response phenotypes.Since our NS deletion analysis pre-sented above focused on the signaling intermediates, it became important to determine to what extent these properties are re-flected in the actual modulation of IFN synthesis and the IFN response. To monitor IFN gene induction and the response to exogenous IFN, we used IFN-promoter-Luc and ISG54 pro-moter-Luc plasmids, respectively, in transient transfections. In the induction assay, we used RSV⌬NS1/2 as the inducer; in the response assay, purified commercial IFN- was added to the medium. The ability of the NS1 and NS2 deletion mutant proteins to suppress these pathways was then tested by transfection with the corresponding NS expression plasmids. These results (Fig. 11), discussed below, were in general agreement with the previous deletion analysis but also implicated potentially novel directions. First, both NS1 and NS2 were able to suppress IFN- pro-moter activation (Fig. 11A), in agreement with downregulation of TRAF3 by both (Fig. 3) and that of IKKεby NS1 (Fig. 5). Interestingly, while NS1 and NS2 with C-terminal deletions retained their TRAF3-lowering activity (Fig. 8), they suffered incremental loss of IFN- promoter silencing activity (Fig. 11A). The IKKε-reducing activity of NS1 was more in tune with the IFN- promoter silencing activity, as NS1 with C-terminal deletions significantly lost both.
[image:7.585.115.469.67.253.2]Second, when the response to exogenous IFN was measured (Fig. 11B), NS2 showed strong suppression, consistent with its STAT2-reducing activity (Fig. 10). The loss of the IFN-silenc-ing activity in NS2 with C-terminal deletions (Fig. 11B) is also consistent with the loss of the STAT2-reducing activity of the same mutant proteins. However, NS1, which was unable to reduce STAT2 (Fig. 10), nonetheless had some IFN response suppression activity, and this can be attributed to the C-termi-nal 10 residues (Fig. 11B).
FIG. 6. Reduction of endogenous STAT2 levels by recombinant NS2. A549 cells in 10-cm2wells were transfected with the indicated amounts
of pCAGGS-FLAG-NS plasmids. The empty pCAGGS vector was also transfected wherever needed to keep the amount of DNA constant at 7 g in every well. Cells were processed 24 h later for IB to detect the proteins (top of each panel) with the specific antibodies described in Materials and Methods. The STAT2 band intensities were normalized against that of actin and plotted (bottom of each panel) as percentages of that of the control cells transfected with the empty pCAGGS vector (lanes 1 and 11 for the respective sets). The mean⫾the standard error of the mean of three experiments are presented.
FIG. 7. Expression of recombinant NS deletion mutant proteins. A549 cells in 10-cm2wells were transfected with 3g of wild-type or
C-terminal deletion-containing pCAGGS-FLAG-NS plasmid. Cells were processed 24 h later for IB with FLAG antibody. Note that the NS1 mutant protein with 30 amino acids deleted from the C terminus could not be detected.
on November 8, 2019 by guest
http://jvi.asm.org/
We conclude that both NS1 and NS2 suppress IFN through multiple mechanisms in addition to the ones involving TRAF3, IKKε, and STAT2, and some of them are also mediated by the C terminus of the NS proteins.
NS1 and NS2 form a complex.As it appeared that NS1 and NS2 both regulate various members of the IFN pathway, we wondered whether they associate with each other. To test this, we performed coimmunoprecipitation experiments with RSV-infected cells, which indeed showed that precipitation of NS2 also pulls down NS1 (Fig. 12A), suggestive of a heteromeric complex. We were unable to perform the reciprocal experi-ment because our NS1 antibody did not work for IP. However, we then performed these experiments with cells transfected with N-terminally FLAG- and HA-tagged NS clones. The re-sults (Fig. 12B) clearly showed that IP of tagged NS1 also pulled down tagged NS2 and vice versa. Since the C-terminal 10 residues of NS1 and NS2 were essential for regulating IKKε and STAT2 levels, we tested whether this region was also important for NS1-NS2 association. As shown,⌬C10 deletion of either NS1 or NS2 did not abrogate the association. We conclude that regions beyond the C-terminal 10 amino acids are involved in the formation and function of the NS1-NS2 complex.
DISCUSSION
The general conclusion of this study is that the two NS proteins of RSV target at least three key signaling molecules of the cellular type I IFN induction and response pathways, namely, TRAF3, IKKε, and STAT2, and perhaps others. Loss of TRAF3, where parallel IFN-inducing signals from the RIG-I and TLR pathways converge, should allow RSV to evade both viral RNA-induced and viral F protein-induced signals. Loss of IKKε would further reduce the signal down-stream, and loss of STAT2 would abrogate the cell’s response to IFN. Clearly, RSV has evolved the two unique NS proteins to simultaneously antagonize multiple members of the IFN pathway to guarantee optimal virus growth.
[image:8.585.134.445.72.369.2]A fundamental difficulty in NS studies is that the relative quantities of NS1 and NS2 in the RSV-infected cell are cur-rently uncertain because it is not clear if the available antibod-ies (2, 17; this study) react with them with equal efficiency. Recombinant expression of FLAG-tagged NS proteins elimi-nates this problem, as the same anti-FLAG antibody is used to detect both NS1 and NS2. Nonetheless, in light of this inherent obstacle, we have used various amounts of NS1 and NS2 plas-mids for optimal expression and then validated the results in
FIG. 8. Lack of a role for the C terminus of NS in TRAF3 reduction. (A, B) A549 cells in 10-cm2wells were transfected with 4g of
FLAG-TRAF3 plasmid. In addition, they were transfected with the indicated amounts of pCAGGS-FLAG-NS plasmids (or 3g of the empty vector in lane 1) or infected with RSV (lanes 2 and 12). Where indicated (B), MG132 was added 18 h later and the cells were incubated for another 6 h. (C) Degradation of IB␣by TNF-␣. Where indicated, A549 cells were treated with TNF-␣in the presence or absence of 10M MG132 for 20 min. All of the cells were then processed for IB with appropriate antibodies to detect actin, IB␣, or FLAG (for TRAF3 and NS). Note that ⌬C10NS2 (lane 6) and⌬C20NS2 (lane 7) migrated either to the same spot as NS1 or too close to NS1 to be resolved clearly. (D) The TRAF3 band intensities were normalized against actin and plotted as percentages of the respective control lanes (lanes 1 and 11). The mean⫾the standard error of the mean of three experiments are presented. wt, wild type.
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9689
on November 8, 2019 by guest
http://jvi.asm.org/
RSV-infected cells wherever possible for additional relevance. Gross misfolding of mutant proteins resulting in nonspecific loss of activity is always a possibility in deletion analysis; how-ever, our NS deletions retained TRAF3-reducing activity, eliminating this concern.
[image:9.585.133.448.73.391.2]It is clear from our results that the NS proteins use diverse mechanisms and domains. Although NS1 promoted the reduc-tion of both TRAF3 and IKKε, the same amounts of NS1 failed to reduce STAT2. The C terminus of NS1 was important for reducing IKKεonly, not TRAF3. NS2 was relatively benign for these two substrates; it reduced TRAF3 weakly and pro-moted only a modest increase of IKKε. In contrast, NS2 re-duced STAT2, while NS1 did not. These results pointed to the unique functions of NS1 and NS2, which were further under-scored by the differential roles of their C termini in these activities. In short, the C-terminal 10 residues of NS1 and NS2 were essential for reducing IKKε and STAT2 levels, respec-tively. In contrast, the C terminus of neither protein was needed for its TRAF3-reducing activity. One possibility is that the exact stoichiometry of the NS1/2 heterodimer is deter-mined by the relative amounts of NS1 and NS2, which may
FIG. 9. Restoration of IKKεlevels upon deletion of the C termini of the NS proteins. These experiments were designed essentially as described for TRAF3 in the legend to Fig. 8. (A, B) A549 cells in 10-cm2wells were transfected with 2g of myc-IKKεplasmid. In addition, they were
transfected with the indicated amounts of pCAGGS-FLAG-NS plasmids (or 3g of the empty vector only in lanes 1 and 11). Where indicated (B), MG132 was added 18 h later and the cells were incubated for another 6 h. All of the cells were then processed for IB with appropriate antibodies to detect the myc tag (IKKε), actin, IB␣(not shown), or the FLAG tag (NS1/2). Note that the mobility of⌬C10NS2 was identical to that of NS1, and therefore, the two bands could not be resolved (lane 5). Lane 10 was taken from a different experiment, as indicated by the dotted line. (C) The band intensities were normalized against that of actin and plotted as percentages of the respective control lanes (lane 1 and 11). The mean⫾the standard error of the mean of three experiments are presented. wt, wild type.
FIG. 10. Destruction of STAT2 requires the C terminus of NS2. A549 cells in 10-cm2wells were transfected with 3g of single plasmids
(pCAGGS-FLAG-NS plasmids or the empty vector) or 1.5g of each plasmid when transfected as a mixture (lane 4). All cells were pro-cessed 24 h posttransfection for IB with STAT2 or actin antibody. The STAT2 band intensities were normalized against actin and plotted as percentages of the control (lane 1). Average values of three experi-ments and standard error bars are presented.
on November 8, 2019 by guest
http://jvi.asm.org/
account for some of the differences reported in the literature, obtained by using overexpressed recombinant NS versus RSV infection. It is also possible that the heterodimer recruits other proteins to form an IFN-suppressive multisubunit complex. There is precedence in the IFN-suppressive V proteins of a number of paramyxoviruses, such as mumps virus, simian virus 5, and human parainfluenza virus type 2, which assemble a multiprotein complex containing E3 ligase activity that de-grades the STAT proteins (18, 27, 37, 52, 57, 69).
Among viral proteins, the RSV matrix protein (M) and the phosphoprotein (P) were found to be associated with NS1 in coimmunoprecipitation studies and in two-hybrid screening, respectively (17, 23). Interestingly, P is a viral transcription factor and M has multiple roles, including viral nucleocapsid packaging and possibly viral transcriptional shutoff at the late stage. It is possible that a subset of the NS1/2 complex recruits these proteins through interaction with NS1 and thus regulates viral gene expression. It is to be noted that although we have only focused on its IFN-suppressive activities, the NS1 protein has multiple roles, including regulation of viral RNA synthesis (2). It remains to be seen whether the interactions with P and M regulate the IFN-silencing function of NS1 in the actual RSV-infected cell.
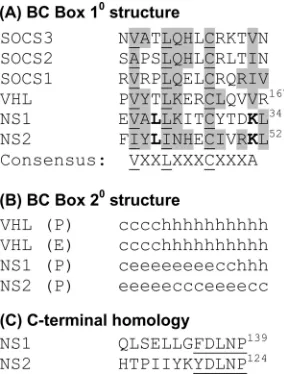
[image:10.585.73.254.68.330.2]As mentioned, Elliott et al. (15) demonstrated E3 ligase activity of the NS1 complex that degraded STAT2. These au-thors also used a CLUSTALW homology search to find a motif in NS1 that showed partial homology to the consensus se-quence for elongin C and cullin 2 binding (called BC box), namely, VXXLXXXCXXX (Fig. 13A), which occurs in arche-typal E3 ligases such as SOCS1 to -3 and VHL (32, 63). It was conjectured that this motif might be responsible for the E3 ligase activity of NS1 (15). As NS2 was shown to decrease STAT2 (40, 52) (Fig. 6 and 10), apparently by using a protea-somal mechanism, we searched for a similar consensus in NS2 and found one at residues 39 to 52 (Fig. 13A), located at a heterologous position from that of NS1. Both sequences showed distant homology with BC Box, as well as significant differences in primary structure and predicted secondary struc-ture (Fig. 13B). It will be interesting to determine whether this motif in the NS proteins is functionally important in the ubiq-uitination and degradation of STAT2. However, even if it is, the C termini may play important roles in this process as well,
FIG. 11. IFN suppression phenotypes of NS1 and NS2 deletion mutants. Infection and transfection were carried out as detailed in the legend to Fig. 3. In brief, A549 cells were first infected with dual NS deletion-containing RSV and 4 h later transfected with the indicated pCAGGS-FLAG-NS plasmids and either (A) an IFN promoter re-porter or (B) an IFN response rere-porter firefly Luc plasmid. All of the cells were also cotransfected with the pCMV-Renilla Luc plasmid for a transfection control. Cells were processed 24 h later for a dual-luciferase assay (Promega), and firefly Luc activity was normalized against that ofRenillaLuc. In each panel, the normalized activity with pCAGGS (vector only, no NS gene) was taken as 100 and all other activities are expressed as percentages thereof. The mean⫾the stan-dard error of the mean of three experiments are presented.
FIG. 12. Coprecipitation of NS1 and NS2. (A) A549 cells were infected with wild-type RSV, and cell lysates made at 18 h p.i. were subjected to IP with our polyclonal rat antibody against NS2 or a no-antibody control (protein A-Sepharose beads only). Precipitated samples were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and IB was performed with the rat NS1 antibody. Note that our rat anti-NS2 antibody works only in IP, not in IB, and vice versa for the anti-NS1 antibody. (B) A549 cells were cotransfected with equal amounts of a mixture of FLAG-NS and pCAGGS-HA-NS plasmids as follows: lane 1, FLAG-NS2 and pCAGGS-HA-NS1; lane 2, FLAG-⌬C10NS2 and HA-NS1; lane 3, FLAG-NS1 and HA-NS2; lane 4, FLAG-⌬C10NS1 and HA-NS2. Cell lysates were made 24 h later, and portions were directly subjected to IB with (i) FLAG antibody or (ii) HA antibody. The remaining portions of the lysates were processed for IP with FLAG antibody, and the precipitate was subjected to IB with either (iii) FLAG antibody or (iv) HA antibody.
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9691
on November 8, 2019 by guest
http://jvi.asm.org/
[image:10.585.319.521.69.335.2]since⌬C10NS2 was defective in this degradative activity. In alternative, nonproteolytic mechanisms, NS proteins may in-hibit the transcription, translation, or mRNA stability of some of its target genes, which remains to be established.
While the manuscript was in preparation, Ling et al. (38) published interesting results focusing on NS2, showing that its expression inhibited IFN gene transcription induced by both RIG-I and TLR3 pathways. They found that NS2 inhibited RIG-I by binding to the CARD domain of the latter and blocking its interaction with mitochondrial antiviral signaling, which is clearly a proteasome-independent mechanism. This is in agreement with our assertion that NS proteins indeed target multiple pathways. Evidently, the subunit composition of the various cellular complexes of NS1 and NS2 should shed light on the multiple functions and mechanisms of these unique RSV proteins. These studies will be facilitated by the recom-binant NS proteins and their mutant forms described here.
ACKNOWLEDGMENTS
This research was supported in part by NIH grant AI059267 and an Alabama EPSCoR-GRSP fellowship.
We thank Hong Jin (MedImmune LLC, Mountain View, CA) for the recombinant⌬NS RSV strains and Ganes C. Sen, Paula Norris, Carl Ware, Chris Basler, and Ramesh Pillai for various plasmids. S.B. is a member of the Lions-USA Eye Research group and thanks the Lions Club International Fund (LCIF) and the local Lions Clubs for the gift of the Nikon TE2000E2 imaging station.
REFERENCES
1.Andrejeva, J., K. S. Childs, D. F. Young, T. S. Carlos, N. Stock, S. Good-bourn, and R. E. Randall.2004. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN- promoter. Proc. Natl. Acad. Sci. USA101:17264–17269.
2.Atreya, P. L., M. E. Peeples, and P. L. Collins.1998. The NS1 protein of human respiratory syncytial virus is a potent inhibitor of minigenome tran-scription and RNA replication. J. Virol.72:1452–1461.
3.Bitko, V., A. Musiyenko, M. A. Bayfield, R. J. Maraia, and S. Barik.2008. Cellular La protein shields nonsegmented negative-strand RNA viral leader RNA from RIG-I and enhances virus growth by diverse mechanisms. J. Vi-rol.82:7977–7987.
4.Bitko, V., and S. Barik.2001. Phenotypic silencing of cytoplasmic genes using sequence-specific double-stranded short interfering RNA and its application in the reverse genetics of wild type negative-strand RNA viruses. BMC Microbiol.1:34.
5.Bitko, V., O. Shulyayeva, B. Mazumder, A. Musiyenko, M. Ramaswamy, D. C. Look, and S. Barik.2007. Nonstructural proteins of respiratory syn-cytial virus suppress premature apoptosis by an NF-B-dependent, interfer-on-independent mechanism and facilitate virus growth. J. Virol.81:1786– 1795.
6.Borden, E. C., G. C. Sen, G. Uze, R. H. Silverman, R. M. Ransohoff, G. R. Foster, and G. R. Stark.2007. Interferons at age 50: past, current and future impact on biomedicine. Nat. Rev. Drug Discov.6:975–990.
7.Bossert, B., S. Marozin, and K. K. Conzelmann.2003. Nonstructural pro-teins NS1 and NS2 of bovine respiratory syncytial virus block activation of interferon regulatory factor 3. J. Virol.77:8661–8668.
8.Bossert, B., and K. K. Conzelmann.2002. Respiratory syncytial virus (RSV) nonstructural (NS) proteins as host range determinants: a chimeric bovine RSV with NS genes from human RSV is attenuated in interferon-competent bovine cells. J. Virol.76:4287–4293.
9.Bowie, A. G., and K. A. Fitzgerald. 2007. RIG-I: tri-ing to discriminate between self and non-self RNA. Trends Immunol.28:147–150.
10.Bowie, A. G., and L. Unterholzner.2008. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol.8:911–922. 11.Chau, T.-L., R. Gioia, J.-S. Gatot, F. Patrascu, I. Carpentier, J.-P. Chapelle,
L. O’Neill, R. Beyaert, J. Piette, and A. Chariot.2008. Are the IKKs and IKK-related kinases TBK1 and IKK-εsimilarly activated? Trends Biochem. Sci.33:171–180.
12.Childs, K., N. Stock, C. Ross, J. Andrejeva, L. Hilton, M. Skinner, R. Randall, and S. Goodbourn.2007. mda-5, but not RIG-I, is a common target for paramyxovirus V proteins. Virology359:190–200.
13.Cle´ment, J. F., S. Meloche, and M. J. Servant. 2008. The IKK-related kinases: from innate immunity to oncogenesis. Cell Res.18:889–899. 14.Darnell, J. E., Jr.2007. Interferon research: impact on understanding
tran-scriptional control. Curr. Top. Microbiol. Immunol.316:155–163. 15.Elliott, J., O. T. Lynch, Y. Suessmuth, P. Qian, C. R. Boyd, J. F. Burrows, R.
Buick, N. J. Stevenson, O. Touzelet, M. Gadina, U. F. Power, and J. A. Johnston.2007. Respiratory syncytial virus NS1 protein degrades STAT2 by using the elongin-cullin E3 ligase. J. Virol.81:3428–3436.
16.Emeny, J. M., and M. J. Morgan.1979. Regulation of the interferon system: evidence that Vero cells have a genetic defect in interferon production. J. Gen. Virol.43:247–252.
17.Evans, J. E., P. A. Cane, and C. R. Pringle.1996. Expression and charac-terisation of the NS1 and NS2 proteins of respiratory syncytial virus. Virus Res.43:155–161.
18.Fontana, J. M., B. Bankamp, and P. A. Rota.2008. Inhibition of interferon induction and signaling by paramyxoviruses. Immunol. Rev.225:46–67. 19.Galinski, M. S., R. M. Troy, and A. K. Banerjee.1992. RNA editing in the
phosphoprotein gene of the human parainfluenza virus type 3. Virology 186:543–550.
20.Gee, P., P. K. Chua, J. Gevorkyan, K. Klumpp, I. Najera, D. C. Swinney, and J. Deval.2008. Essential role of the N-terminal domain in the regulation of RIG-I ATPase activity. J. Biol. Chem.283:9488–9496.
21.Hausmann, S., D. Garcin, C. Delenda, and D. Kolakofsky.1999. The versa-tility of paramyxovirus RNA polymerase stuttering. J. Virol.73:5568–5576. 22.He, B., R. G. Paterson, N. Stock, J. E. Durbin, R. K. Durbin, S. Goodbourn, R. E. Randall, and R. A. Lamb.2002. Recovery of paramyxovirus simian virus 5 with a V protein lacking the conserved cysteine-rich domain: the multifunctional V protein blocks both interferon-beta induction and inter-feron signaling. Virology303:15–32.
23.Hengst, U., and P. Kiefer.2000. Domains of human respiratory syncytial virus P protein essential for homodimerization and for binding to N and NS1 protein. Virus Genes20:221–225.
24.Hiscott, J. 2007. Triggering the innate antiviral response through IRF-3 activation. J. Biol. Chem.282:15325–15329.
25.Hiscott, J.2007. Convergence of the NF-B and IRF pathways in the reg-ulation of the innate antiviral response. Cytokine Growth Factor Rev.18: 483–490.
[image:11.585.91.233.69.256.2]26.Honda, K., and T. Taniguchi.2006. IRFs: master regulators of signaling by
FIG. 13. Sequence features of NS1 and NS2. (A) The BC Box homology domain, modeled after that reported by Elliot et al. (15), showing conservation among known members of this family (SOCS1 to -3, VHL). Conserved amino acids, present in at least one NS sequence and in another protein, are highlighted. The consensus 12-residue motif is shown at the bottom, with the invariant Val (or other aliphatic/ hydrophobic residues), Leu/Ile, and Cys residues underlined and the intervening variable residues denoted as X. The nonconsensus Lys (K) residue, found in the NS proteins only, is in bold. 10, primary.
(B) Local secondary (20) structures of the same 14-residue segment as
in panel A are shown for the VHL and NS proteins: P, predicted by using the Porter algorithm (49); E, experimental (32, 63); c, coil (loop region); h, alpha helix; e, extended sheet (sheet). (C) The homology at the C termini of NS1 and NS2 includes the underlined conserved terminal (F/Y)DLNP sequence. Residue numbers, corresponding to
the full-length protein, are noted at the ends of selected sequences.
on November 8, 2019 by guest
http://jvi.asm.org/
Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol.6:644–658.
27.Horvath, C. M.2004. Weapons of STAT destruction. Interferon evasion by paramyxovirus V protein. Eur. J. Biochem.271:4621–4628.
28.Indukuri, H., S. M. Castro, S.-M. Liao, L. A. Feeney, M. Dorsch, A. J. Coyle, and R. P. Garofalo, A. R. Brasier, and A. Casola.2006. Ikkepsilon regulates viral-induced interferon regulatory factor-3 activation via a redox-sensitive pathway. Virology353:155–165. (Erratum,357:114, 2007.)
29.Israe¨l, A.2000. The IKK complex: an integrator of all signals that activate NF-B? Trends Cell Biol.10:129–133.
30.Jin, H., H. Zhou, X. Cheng, R. Tang, M. Munoz, and N. Nguyen.2000. Recombinant respiratory syncytial viruses with deletions in the NS1, NS2, SH, and M2-2 genes are attenuated in vitro and in vivo. Virology273:210– 218.
31.Kato, H., O. Takeuchi, S. Sato, M. Yoneyama, M. Yamamoto, K. Matsui, S. Uematsu, A. Jung, T. Kawai, K. J. Ishii, O. Yamaguchi, K. Otsu, T. Tsu-jimura, C. S. Koh, C. Reis e Sousa, Y. Matsuura, T. Fujita, and S. Akira. 2006. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature441:101–105.
32.Kibel, A., O. Iliopoulos, J. A. DeCaprio, W. G. Kaelin, Jr.1995. Binding of the von Hippel-Lindau tumor suppressor protein to elongin B and C. Science 269:1444–1446.
33.Komatsu, T., K. Takeuchi, and B. Gotoh.2007. Bovine parainfluenza virus type 3 accessory proteins that suppress beta interferon production. Microbes Infect.9:954–962.
34.Komuro, A., D. Bamming, and C. M. Horvath.2008. Negative regulation of cytoplasmic RNA-mediated antiviral signaling. Cytokine43:350–358. 35.Kumar, R., B. Adams, A. Musiyenko, O. Shulyayeva, and S. Barik.2005. The
FK506-binding protein of the malaria parasite, Plasmodium falciparum, is a FK506-sensitive chaperone with FK506-independent calcineurin-inhibitory activity. Mol. Biochem. Parasitol.141:163–173.
36.Kurt-Jones, E. A., L. Popova, L. Kwinn, L. M. Haynes, L. P. Jones, R. A. Tripp, E. E. Walsh, M. W. Freeman, D. T. Golenbock, L. J. Anderson, and R. W. Finberg.2000. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol.1:398–401. 37.Li, T., X. Chen, K. C. Garbutt, P. Zhou, and N. Zheng.2006. Structure of
DDB1 in complex with a paramyxovirus V protein: viral hijack of a propeller cluster in ubiquitin ligase. Cell124:105–117.
38.Ling, Z., K. C. Tran, and M. N. Teng.2009. The human respiratory syncytial virus nonstructural NS2 protein antagonizes the activation of interferon- transcription by interacting with RIG-I. J. Virol.83:3734–3742.
39.Liu, P., M. Jamaluddin, K. Li, R. P. Garofalo, A. Casola, and A. R. Brasier. 2007. Retinoic acid-inducible gene I mediates early antiviral response and Toll-like receptor 3 expression in respiratory syncytial virus-infected airway epithelial cells. J. Virol.81:1401–1411.
40.Lo, M. S., R. M. Brazas, and M. J. Holtzman.2005. Respiratory syncytial virus nonstructural proteins NS1 and NS2 mediate inhibition of Stat2 ex-pression and alpha/beta interferon responsiveness. J. Virol.79:9315–9319. 41.Loo, Y. M., J. Fornek, N. Crochet, G. Bajwa, O. Perwitasari, L.
Martinez-Sobrido, S. Akira, M. A. Gill, A. García-Sastre, M. G. Katze, and M. Gale, Jr.2008. Distinct RIG-I and MDA5 signaling by RNA viruses in innate immunity. J. Virol.82:335–345.
42.Mori, M., M. Yoneyama, T. Ito, K. Takahashi, F. Inagaki, and T. Fujita. 2004. Identification of Ser-386 of interferon regulatory factor 3 as critical target for inducible phosphorylation that determines activation. J. Biol. Chem.279:9698–9702.
43.Nguyen, H., J. Hiscott, and P. M. Pitha.1997. The growing family of inter-feron regulatory factors. Cytokine Growth Factor Rev.8:293–312. 44.Niwa, H., K. Yamamura, and J. Miyazaki. 1991. Efficient selection for
high-expression transfectants with a novel eukaryotic vector. Gene108:193– 199.
45.Nokes, J. D., and P. A. Cane.2008. New strategies for control of respiratory syncytial virus infection. Curr. Opin. Infect. Dis.21:639–643.
46.Pestka, S.2007. The interferons: 50 years after their discovery, there is much more to learn. J. Biol. Chem.282:20047–20051.
47.Pichlmair, A., and C. Reis e Sousa.2007. Innate recognition of viruses. Immunity27:370–383.
48.Pillai, R. S., S. N. Bhattacharyya, C. G. Artus, T. Zoller, N. Cougot, E. Basyuk, E. Bertrand, and W. Filipowicz.2005. Inhibition of translational initiation by Let-7 MicroRNA in human cells. Science309:1573–1576. 49.Pollastri, G., and A. McLysaght.2005. Porter: a new, accurate server for
protein secondary structure prediction. Bioinformatics21:1719–1720. 50.Poole, E., B. He, R. A. Lamb, R. E. Randall, and S. Goodbourn.2002. The
V proteins of simian virus 5 and other paramyxoviruses inhibit induction of interferon-. Virology303:33–46.
51.Precious, B., D. F. Young, L. Andrejeva, S. Goodbourn, and R. E. Randall. 2005. In vitro and in vivo specificity of ubiquitination and degradation of STAT1 and STAT2 by the V proteins of the paramyxoviruses simian virus 5 and human parainfluenza virus type 2. J. Gen. Virol.86:151–158. 52.Ramaswamy, M., L. Shi, S. M. Varga, S. Barik, S., M. A. Behlke, and D. C.
Look.2006. Respiratory syncytial virus nonstructural protein 2 specifically
inhibits type I interferon signal transduction. Virology344:328–339. (Erra-tum,366:453, 2007.)
53.Reid, S. P., C. Valmas, O. Martinez, F. M. Sanchez, and C. F. Basler.2007. Ebola virus VP24 proteins inhibit the interaction of NPI-1 subfamily karyo-pherin alpha proteins with activated STAT1. J. Virol.81:13469–13477. 54.Sasai, M., M. Shingai, K. Funami, M. Yoneyama, T. Fujita, M. Matsumoto,
and T. Seya.2006. NAK-associated protein 1 participates in both the TLR3 and the cytoplasmic pathways in type I IFN induction. J. Immunol.177:8676– 8683.
55.Schindler, C., and C. Plumlee.2008. Interferons pen the JAK-STAT path-way. Semin. Cell Dev. Biol.19:311–318.
56.Schlender, J., B. Bossert, U. Buchholz, and K. K. Conzelmann.2000. Bovine respiratory syncytial virus nonstructural proteins NS1 and NS2 cooperatively antagonize alpha/beta interferon-induced antiviral response. J. Virol. 74: 8234–8242.
57.Schro¨der, M., and A. G. Bowie.2007. An arms race: innate antiviral re-sponses and counteracting viral strategies. Biochem. Soc. Trans.35:1512– 1514.
58.Sen, G. C., and S. N. Sarkar.2007. The interferon-stimulated genes: targets of direct signaling by interferons, double-stranded RNA, and viruses. Curr. Top. Microbiol. Immunol.316:233–250.
59.Simo˜es, E. A.2008. RSV disease in the pediatric population: epidemiology, seasonal variability, and long-term outcomes. Manag. Care17:3–6. 60.Spann, K. M., K. C. Tran, and P. L. Collins.2005. Effects of nonstructural
proteins NS1 and NS2 of human respiratory syncytial virus on interferon regulatory factor 3, NF-B, and proinflammatory cytokines. J. Virol.79: 5353–5362.
61.Spann, K. M., K. C. Tran, B. Chi, R. L. Rabin, and P. L. Collins.2004. Suppression of the induction of alpha, beta, and lambda interferons by the NS1 and NS2 proteins of human respiratory syncytial virus in human epi-thelial cells and macrophages. J. Virol.78:4363–4369.
62.Stark, G. R.2007. How cells respond to interferons revisited: from early history to current complexity. Cytokine Growth Factor Rev.18:419–423. 63.Stebbins, C. E., W. G. Kaelin, Jr., and N. P. Pavletich.1999. Structure of the
VHL-ElonginC-ElonginB complex: implications for VHL tumor suppressor function. Science284:455–461.
64.Takeuchi, O., and S. Akira.2008. MDA5/RIG-I and virus recognition. Curr. Opin. Immunol.20:17–22.
65.Teng, M. N., and P. L. Collins.1999. Altered growth characteristics of recombinant respiratory syncytial viruses which do not produce NS2 protein. J. Virol.73:466–473.
66.Teng, M. N., S. S. Whitehead, A. Bermingham, M. St. Claire, W. R. Elkins, B. R. Murphy, and P. L. Collins.2000. Recombinant respiratory syncytial virus that does not express the NS1 or M2-2 protein is highly attenuated and immunogenic in chimpanzees. J. Virol.74:9317–9321.
67.Tenoever, B. R., S.-L. Ng, M. A. Chua, S. M. McWhirter, A. García-Sastre, and T. Maniatis.2007. Multiple functions of the IKK-related kinase IKKεin interferon-mediated antiviral immunity. Science315:1274–1278.
68.Ueba, O.1978. Respiratory syncytial virus. I. Concentration and purification of the infectious virus. Acta Med. Okayama32:265–272.
69.Ulane, C. M., A. Kentsis, C. D. Cruz, J. P. Parisien, K. L. Schneider, and C. M. Horvath.2005. Composition and assembly of STAT-targeting ubiq-uitin ligase complexes: paramyxovirus V protein carboxyl terminus is an oligomerization domain. J. Virol.79:10180–10189.
70.Valarcher, J. F., J. Furze, S. Wyld, R. Cook, K. K. Conzelmann, and G. Taylor.2003. Role of alpha/beta interferons in the attenuation and immu-nogenicity of recombinant bovine respiratory syncytial viruses lacking NS proteins. J. Virol.77:8426–8439.
71.Weber, F., and O. Haller.2007. Viral suppression of the interferon system. Biochimie89:836–842.
72.Whitehead, S. S., A. Bukreyev, M. N. Teng, C. Y. Firestone, M. St Claire, W. R. Elkins, P. L. Collins, and B. R. Murphy.1999. Recombinant respira-tory syncytial virus bearing a deletion of either the NS2 or SH gene is attenuated in chimpanzees. J. Virol.73:3438–3442.
73.Yoneyama, M., and T. Fujita.2007. Function of RIG-I-like receptors in antiviral innate immunity. J. Biol. Chem.282:15315–15318.
74.Yoneyama, M., M. Kikuchi, K. Matsumoto, T. Imaizumi, M. Miyagishi, K. Taira, E. Foy, Y. M. Loo, M. Gale, Jr., S. Akira, S. Yonehara, A. Kato, and T. Fujita.2005. Shared and unique functions of the DExD/H-box helicases RIG-I, MDA5, and LGP2 in antiviral innate immunity. J. Immunol.175: 2851–2858.
75.Yoneyama, M., M. Kikuchi, T. Natsukawa, N. Shinobu, T. Imaizumi, M. Miyagishi, K. Taira, S. Akira, and T. Fujita.2004. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol.5:730–737.
76.Zhang, W., H. Yang, X. Kong, S. Mohapatra, H. San Juan-Vergara, G. Hellermann, S. Behera, R. Singam, R. F. Lockey, and S. S. Mohapatra.2005. Inhibition of respiratory syncytial virus infection with intranasal siRNA nanoparticles targeting the viral NS1 gene. Nat. Med.11:56–62. (Erratum, 11:233.)
VOL. 83, 2009 MULTIPLE CELLULAR TARGETS OF RSV NS PROTEINS 9693