JOURNALOFVIROLOGY,Aug. 1992,P.5119-5126
0022-538X/92/085119-08$02.00/0
Copyright ) 1992,American Society for Microbiology
Envelope Glycoprotein and
CD4
Independence
of
vpu-Facilitated
Human
Immunodeficiency
Virus
Type 1 Capsid Export
XIAO JIANYAO,1HEINRICH GOTILINGER,2WILLIAM A. HASELTINE,2ANDERIC A. COHEN1*
Departement de MicrobiologieetImmunologie, Faculte deMedecine, Universite de Montreal, Montreal
Quebec, Canada H3C
3J7,1
and Division of HumanRetrovirology, Dana-Farber Cancer Institute, HarvardMedical School, Boston, Massachusetts 021159Received 21 March 1992/Accepted 28 April 1992
The effectofvpuon the release ofhuman immunodeficiency type 1capsid proteins was examined inthe presence orabsence of virus-encoded envelopeglycoproteins aswellasin cells which constitutively express
either theCD4orCD8protein. The results showthat vpu-mediated facilitatedexportof capsid proteins from HeLa cells doesnotrequireexpressionoftheenvelope glycoprotein.Theexperimentsalso showthatexportof virus capsidproteins from HeLacellsfacilitatedbyvpuisnotaffected bycoexpression of either the CD4orCD8 protein. Thevpuproteinactsintranstofacilitateexportof viruscapsid proteins from HeLa cells.
Thevpu protein is encodedby human immunodeficiency
virustype 1 (HIV-1) but notby closely related human and
simian retroviruses, including HIV-2and thesimian
immu-nodeficiencyviruses (4). The vpu protein is specified by a
short open reading frame that overlaps the 5' end of the
envelope glycoproteingene (env) (4, 15, 21).Previous stud-ies haveshown thatvpusignificantly increasesthenumber of
virus particles released into the supernatant of infected CD4+T-cell lines(11,20-22). Theincrease in the release of virusparticlesofVpu+viruses fromCD4+Tcellscompared with the release of otherwise isogenic Vpu- viruses is accompanied by a decrease in the accumulation of
cell-associated protein rather than by an increase in the total
amount of viral proteinsmade (11, 20, 22). These
observa-tionssuggest thatvpu increasesthe rateof virusexportby facilitatingeither therateofassemblyof the virusparticleor
the rate of release of the assembled particles from the infected cell surface.
The observation that the vpu protein and the envelope glycoproteinaremade fromasinglebicistronic mRNA raises the possibility that coordinate expression of vpu and the
envelope glycoproteins maybe required foran increase in
the efficiency of virus export (1, 18). It has also been
reported that vpu may affect maturation of the envelope glycoprotein by alteringtheprocessingof thenascent poly-peptideeitherdirectlyorbythedisruptionofanintracellular
complexformed between theenvelopeglycoprotein
precur-sorandthe CD4nascentprotein (23).
To study the role ofthe envelope glycoprotein and the HIV-1 receptor, the CD4 protein, on vpu export of virus
capsids, an experimental system that does not depend on
virus replication is needed. The feasibility ofusing HeLa cells as recipients in transfection assays for these studies
was investigated. In the initial experiment, 106 HeLa cells
weretransfected with 10or40,gof DNA oftheVpu-strain HXBc2(5), and theVpu+ strainHXBH10-vpu+/env+ (22), using the calcium phosphate coprecipitation technique (7) (Fig. 1A). Although all sequences of the HXBH10-vpu+/
env+ strain arederived from the IIIB isolate ofHIV-1, the
Correspondingauthor.
region of viralDNA between theSalI siteatnucleotide 5372 (+1 is the transcriptioninitiation site) andthe
KIpnI
site at nucleotide 5934 are derived from the BH10 provirus thatcontainsan intactvpu openreadingframe rather thanfrom
theVpu- HXBc2DNA(16)(Fig. 1A).
Virus protein export from HeLa cells was followed by
labeling the cells transfected with HXBc2 and
HXBH10-vpu+/env+ DNA with [35S]cysteine (50 ,uCi/ml) for 12 h
beginning at48 h posttransfection. The cellswerewashed and lysedin RIPAbuffer(10 mMTris-HCl [pH 7.4], 1 mM
EDTA, 100 mM NaCl, 1% Triton X-100, 0.1% sodium
dodecyl sulfate
[SDS],
0.25% deoxycholate, 0.2% phenyl-methylsulfonyl fluoride), and the virusproteinwasprecip-itated with an HIV-1-positive human serum. The virus proteinswerevisualizedbySDS-polyacrylamide gel
electro-phoresis (PAGE) (11.5% polyacrylamide gel) and
autoradi-ography. Thevirus-specificproteinspresentin the cell-free filtered supernatant fractionweredetectedby precipitation
of these proteins with an HIV-1-positive human serum.
Additionally, virus particles from the supernatant were
collectedbyhigh-speed centrifugationontoadensesucrose cushion. The virusproteins inthe pelletwereanalyzedon SDS-polyacrylamidegelsdirectlywithout
immunoprecipita-tion.
The data in Fig. 2A shows that the amounts of
virus-specific protein inHeLa cells transfected with HXBc2 and
HXB10-vpu+/env+provirusesaresimilar. The relative
abun-dance of the capsid precursor p55 and of the processed capsid proteins p17andp24/25aresimilar,as arethe relative amounts of the envelope glycoproteins gpl60 and gpl20 (lanes 1to4).
In contrast,theamountofcapsid proteinreleased into the cell supernatant as detected by using an HIV-1-positive
human serum is markedlydifferent. It is evident that
con-siderably more virus capsid proteins are present in the supernatant fluids of cells transfectedwith HXBH10-vpu+/
env+ (Fig. 2A, lanes 7 and8)thanin cells transfected with HXBc2-vpu- (lanes 5 and 6). There is no reproducible
change in the amount of the envelope glycoprotein gp120 presentin the supematant fluidsof cells transfected with the two DNAs.
The increased export of virusparticles bythe HXBH10-Vol. 66, No. 8
on November 9, 2019 by guest
http://jvi.asm.org/
A:
EviJ
I
tatEa vpu vpr rev
J
00
0 1 2 3 4 5 6
SaIl KpnlI
I r1
I
ro;7;-HXBH1O
r'
TAT
|
n i.&Ighnvw
I
EV1
I TAT
I
I
0 VPUI
I
I
ENV
k
II
LIV
IP
:
I*EZW
1TAT
I
* VIZ
ra
EE
tatD
Ea
rev
6nv
7 8 9 10
2
. .
VPU ENV
_ +
SVCMVvpu+
(ATG)
=~~~~~~~~
X
SVCMVvpu-(ACG)
vpu+|lenv+
HXBH10
vpu-/env.
HXBH1O vpu
+/env-HXBHIO
vpu-/env-B:
It
I I I I I
I
K
K X
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 5121
vpu+/env+strain is also reflected in the dramatic increase in
theamountofvirus particles pelleted from these supernatant fluids. Densitometric analysis of the autoradiogram shown in Fig. 2B indicates that at least 10 times more virus particles
arereleased from HeLa cells transfected with the
HXBH10-vpu+Ienv+DNA(lane 1) than from cells transfected with the Vpu- HXBc2 DNA(lane 2).
Role of envelope glycoprotein and CD4 in vpu-mediated capsid protein export. Thepreliminary experiments
demon-strate that the effect of vpu on export of HIV-1 virion
proteinscanbe assessedin HeLa cells. To examine the role of theenvelope glycoproteininvpu-mediated export of virus capsid proteins, a set of four isogenic strains of viral DNAs was made. These four strains are all derived from the HXBH10-vpu+/env+ DNA clone and are designated the HXBH10-vpu+/env+,HXBH10-vpu+/env-, HXBH10-vpu-/ env+, andHXBH10-vpu-/env- strains (Fig. 1A). The initi-ation codons of the open readingframes that specify the vpu andenvelope glycoproteinswerealteredfromATG to ACG
by site-directed mutagenesis (24).
To assess the role of the CD4 protein in vpu-mediated export of viral proteins, HeLacells that expresseither the CD4 protein (HeLa-T4) (3) or the CD8 protein (HeLa-T8) (14)wereused as recipients in these experiments. The CD4
protein present on the HeLa-T4 cell line has been demon-strated to serveas a functionalreceptorfor HIV-1 (3, 14).
For the initial experiments, the role of env proteins and CD4on vpu-mediatedcapsid export was assessed by trans-fection of HeLa-T4 and HeLa-T8 cells (106)with the set of four viral DNAs (10
pg).
Release of capsid antigens was determined by detection of p24 in the culture supernatantfluids, using a commercial radioimmune competition assay kit(NEK-040;NewEnglandNuclear).The amount of capsid
antigenin thefilteredsupernatantswasmeasuredinintervals
for 60 hposttransfection.
Thedata in Fig. 2C show that substantiallymore p24 is presentin culture supernatantfluids ofHeLa-T4cells
trans-fected with the
HXBH10-vpu+/env+
viral DNAthan in thesupernatant of cells which received the Vpu-/Env+ DNA. The amount ofp24 releasedby CD4 cells transfected with theVpu+/Env-virus is also greater than that released from CD4+ cells which received the Vpu-/Env- DNA. The
amount ofp24 protein released into the cell supematant is
similarfor HeLa-T4cells transfected with
Env'
and Env-viral DNAs.The data in Fig. 2C also show that the amount of p24
released into the cell supernatant of CD8 cells transfected with the Vpu+/Env+ DNA is significantly greaterthan the
amountreleasedfrom cells transfected with theVpu-/Env+
DNA. Similarly, the amount of capsid antigens released into the culture fluid ofHeLa-T8 cells is greater for cells trans-fectedwith the Vpu+/Env- DNA than it is for cells
trans-fected with the Vpu-/Env- DNA. As before, the total amount of p24 released into the supernatant fluids of the HeLa-T8 cells depends on the genotype of vpu but not the genotype of env. For all virus strains used, the amount ofp24 releasedfrom HeLa-T8 cells is about twice the comparable
amountreleased from HeLa-T4cells. This difference prob-ably reflects a consistent increase in the transfection effi-ciency of the HeLa-T8cells compared with that of HeLa-T4 cells.
Results similar to those described above were obtained whenrelease ofvirus-specified RNA-dependent polymerase wasmeasured (data not shown).
The distribution ofvirus-specific proteins between the cell and the culture supematant fluid was also determined. For theseexperiments, HeLa-T4 and HeLa-T8 cellswere trans-fected with 10 ,g ofHXBH10-vpu+/env+, HXBH10-vpu-/
env+,
HXBH10-vpu+/env-,
or HXBH10-vpu-/env- viral DNA. The HeLa-T4cells were labeled with 100RCi/ml
of [35S]methionine for either 2.5 or5 h beginning 60 hpost-transfection. Both cell lysates and supernatant fluidswere
immunoprecipitated by using a mix of HIV-1-positive human serum and monospecific anti-vpu peptide serum (4). The
immunoprecipitates were analyzed by SDS-PAGE (12.5 polyacrylamide gel) and autoradiography. The
autoradio-graphswere scanned toprovide quantitativecomparisons.
The data in Fig. 2D show that the amount of capsid proteins detected in HeLa-T4 cell lysates is independentof the viral DNA strainused. Densitometrictracingof thegels showsno
significant
difference in theamountofp24present inthe cells transfected with any of the four viral DNAs. Thedata alsoshow that the vpu proteinis present in lysatesof
cellstransfected with the
HXBH10-vpu+/env+
(lanes 1 and2)and
HXBH10-vpu+/env-
(lanes5 and6)DNAs.The vpuprotein is not detected in lysates of cells transfected with either theHXBH10-vpu-/env+(lanes 3 and 4)or
HXBH10-vpu-lenv- (lanes 7 and 8) DNA. The amounts of the
envelope glycoproteins gp160andgp120presentin HeLa-T4 cell lysates transfected with
HXBH10-vpu+/env+
andHXBH10-vpu-/env+
DNAs are similar in lysates of cells labeled both for 2.5 and for 5 h(lanes1 to4).There is a marked difference in the amount of capsid proteinspresent in the supematant fluids of HeLa-T4 cells transfected withVpu+asopposedtoVpu-DNA. Abundant
p24 antigen is detected by radioimmunoprecipitation in
supernatants of HeLa-T4 cells transfected with either
HXBH10-vpu+/env+
orHXBH10-vpu+/env-
viral DNA.ByFIG. 1. (A) Oligonucleotide-directed mutagenesisof the initiationcodon oftheenvgeneof HIV-1. Thegeneticmapof HIV-1indicating thepositionofthevpu genein the viral genome isshown.Theconstruction ofHXBH10-vpu+/env+ andHXBH10-vpu-/env+waspreviously
described (22). Segmentsof the HXBH10-vpu+/env+ andHXBH10-vpu-/env+ proviruseswereclonedintothepTZ19U Genescribevector by using the Sall and
KpnI
sites indicated.Alignment of the viralopenreading frameswithin thefragmentis shown.Oligonucleotide-directedmutagenesis was carried out to remove the env initiation codon, mutating ATG to ACG. These env mutants were cloned back into
HXBH10-vpu /env+ andHXBH10-vpu-/env+ bytaking advantage of the same restriction sitesto yield replication-defective proviruses
HXBH10-vpu+/env-andHXBH10-vpu-/env-,respectively.Arrowheads indicate intactinitiationcodons.The scale is in kilobases.(B)vpu expressorplasmids. Plasmid SVCMVvpu+wasconstructedbyinsertingaDNAfragmentencodingvpufrom theBH10molecular clone 3'to the cytomegalovirus(CMV)immediate-earlygene promoter(-990 to +66) (10)and 5' tothe simian virus 40 (SV40)16S RNAspliceand poly(A)site(6).SVCMVvpu+also containsasimian virus40origin ofreplication.The297-bpXbaI-SacIDNAfragment (nucleotides6085
to 6382) encoding vpu and the N-terminal portion (42 aminoacids) ofenv was generatedbypolymerase chain reaction (17), using the
HXBH10-vpu+/env+ molecular clone asthe template. SVCMVvpu- wasderived from SVCMVvpu+except that apoint mutation in the initiation codon of vpuwas introduced byoligonucleotide-directed mutagenesis, replacing ATG with ACG. The open boxes represent VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
5122 NOTES
A
1 2 3 4 M 5 6 7 8
B
a'
i
-F _-0
gP:20
D
1 2 CELL LYSATE
+ + m,+ +
>?; ... > >
CC c
CD J0 Is I0I~
gp160* # * *
-CDm; m5 0 Wa CD C:
C. X C. >x a E X X
2.5 5 2.5 5 2.5 5 2.5 5 5 2.5 5 2.5 5 2.5 5 2.5 5 5
gplBO0 OM,e
gpi2O
p55 j7 .
t
X
*s
;3$-3
jwMd - p24 - O
p17 -
I4V
- * - - p17 - SNC
-p24 W w-_ _
vpu* I'
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
ela-T8O HXBH 0
vpu+/env+
0
HXBH 0 vpu-/env+2 24 36 48 60
ila-T8
I.El
_HX3HIO
vpu+/env-[7_ ii HXBHI10
11
. ~~vpu-/env-2 24 36 48 60 1 2 24 36 48 60
Times after transfection (hours ) gpl60 ,
gpl2O /
p55
-p41
-p24
-SUPERNATANT
a
C
6
Vn
N
r
J. VIROL.
Imp,mm-, lqwl
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 5123 contrast, only a very small amount of p24 is detected by
radioimmunoprecipitation in supernatant fluids of HeLa-T4 cells transfected with either the HXBH10-vpu-/env+ or
HXBH10-vpu-/env- viral DNA (Fig. 2D; compare lanes 1
and2with lanes 3 and 4 and lanes 5 and 6 with lanes 7 and
8).
There is no reproducible difference in the amount of
envelope glycoprotein gp120released into the media from
cells transfected with the Vpu+/Env+ and Vpu-/Env+
DNAs. The release ofgpl20 into the supernatant fluid by
viruses derived from the IIIB isolate has previously been shown to beindependentof thesynthesisof capsid proteins
(9, 12). No vpu protein can be detected in thesupernatant
fluids of HeLa-T4 cells transfected with HXBH10-vpu+/
env'
(Fig. 2D, lanes1and2) andHXBH10-vpu+/env- (lanes5 and6) viral DNAs.
Distribution ofvirus-specific proteins between cells and
the culture fluidof HeLa-T8 cellstransfected withthe setof
four viral DNAs showed a pattern similar to the one
ob-servedfor HeLa-T4 and HeLacells (datanotshown). Theseresultsindicate thatthe effectof vpu on the export ofvirus capsid proteins from HeLa cells does not require expression oftheenvelopeglycoproteins and isnot affected bycoexpressionof the CD4protein.
Effect of vpu on export ofinfectiousvirus.Theinfectivity of
supernatant fluids of HeLa-T8 cells transfected with
HXBH10-vpu+/env+ and
HXBH10-vpu-/env+
viral DNAs for the humanCD4+T-celllineMT4(8)wasdetermined.Forthis experiment, 200
pl
ofcellular supernatant fluids washarvested fromHeLa-T8 cellstransfected with10
p,g
of viral DNA 60 h posttransfection. The supernatant fluids were used toinfect 5 x 105MT4 cells.Theinfected cultureswereexaminedforevidenceofvirus replication 3 to 6 days postinfection. Infection was moni-tored by determination of the percentage of cells in the culture that arepositive forvirus-specific antigens,asjudged by immunofluorescence using anti-HIV-1 p24 monoclonal
antibodies (2). The amount ofvirus-specified RNA-depen-dentDNApolymeraseactivity released intothesupernatant fluid of the MT4 cultureswas also measured (13), and the
cultures were scored forsyncytium formation.
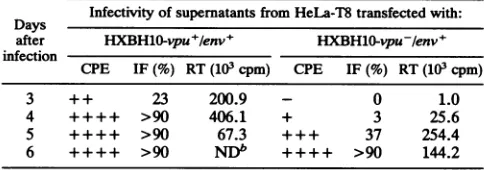
The datainTable 1 show that virus in thesupernatantsof
cultures transfected with the HXBH10-vpu+/env+ DNA initiatesinfection muchmorerapidlythandoes virus inthe supernatant fluids of cells which receive the Vpu-/Env+
DNA. Previous experiments haveshown that thevpu gene
itselfhaslittleeffect on the infectivityor rateofreplication
ofthevirus in T-celllines,asjudged byallthreeassays used
(22). Theseexperimentsindicate that the level ofinfectious
[image:5.612.315.557.93.178.2]viruspresent in the culture supematants ofcellstransfected
TABLE 1. Effect of vpuonexport ofinfectiousparticlesa
Days Infectivityof
supernatants
from HeLa-T8 transfected with: after HXBH10-vpu+/env+ HXBHO-vpu-/env+ infectionCPE IF(%) RT(103 cpm) CPE IF(%) RT(103 cpm)
3 + + 23 200.9 - 0 1.0
4 ++++ >90 406.1 + 3 25.6
5 ++++ >90 67.3 +++ 37 254.4
6 ++++ >90 NDb ++++ >90 144.2 a HeLa-T8cellsweretransfected with 10pgofHXBH10-vpu+/env+ or
HXBH10-vpu-/env+.Forty-eight hours posttransfection, 200pl ofcell-free
supernatantfrom both cultures was usedtoinfect5x105MT4cells. Effects of virusinfection within each culture were thenmonitored at regular intervals. Cytopathic effects (CPE) were measured by daily countof syncytia. Spread of infection and virus production were determined by measuringp24-specific immunofluorescence (IF) and reverse transcriptase activity (RT), respec-tively. For the latter data, background values have been subtracted.
bND,notdetermined.
with
HXBH10-vpu+/env+
DNA isgreater than the amount presentin cells transfected withHXBH10-vpu-/env+
DNA. Effect of vpu in ahumanCD4+ T-cellline.The effect of vpu oncapsidrelease wasalso examined inthe humanCD4+ cell line MT4. Forthis experiment, 107 MT4 cellsweretrans-fected with 12 pLg of either HXBH10-vpu+/env- or
HXBH10-vpu-/env- DNA by the DEAE-dextran method
(19).The amount ofcapsid proteins transientlyreleased into
the supernatantof eachtransfectedculturewas determined
by measuring the virus-specified RNA-dependent DNA polymerase activity at intervals for 60 h posttransfection
(Fig. 3A). Bythismeasure,particleproductionwashigherat all times posttransfection in the MT4 culture transfected with theVpu+/Env- DNAthan in the MT4 culture
trans-fected with theVpu-/Env-DNA. At36 hposttransfection,
the amountof enzymeactivityreleased into thesupernatant
of MT4 cells transfected with the Vpu+/Env- DNA was fourfold greater than the amountreleased from cells trans-fected with theVpu-/Env-DNA.These results indicatethat vpu increases the release of virus capsid proteins in MT4
supernatantfluidindependentlyof the env genotype. The distribution of viral proteins between the cells and
supernatant fluidswasalso examined.Thetransfected cells were labeled for 5 h with
[35S]methionine
(100p,Ci/ml)
beginning60 hposttransfection. The labeledcells and
super-natant-containing virus were immunoprecipitated with an
HIV-1-positive humanserum.Theimmunoprecipitateswere
analyzedonSDS-12.5% polyacrylamide gels.
The data in Fig. 3B show that the level ofp24
immuno-precipitated is threefold higher in the supernatant ofMT4
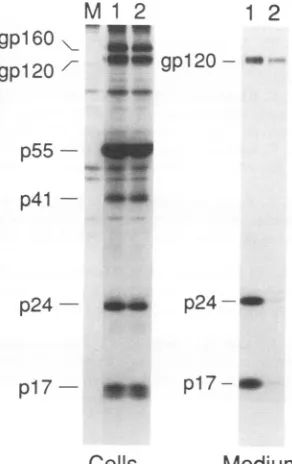
FIG. 2. Role ofenvelope glycoprotein and CD4 invpu-mediated capsid protein export. (A) Immunoprecipitation of viruscapsid and envelope proteins from HeLa cell lysates (lanes 1 to 4) and supernatant fluids (lanes 5 to 8). The cells were transfected with 10
pLg
(odd-numberedlanes)or40p,g(even-numbered lanes)of HXBc2(lanes 1, 2, 5,and6)orHXBH10-vpu+/env+ (lanes 3, 4, 7,and8).LaneM, mock-transfected cells. (B) Sedimentation of viral particles from culture supernatants. HeLa cells were transfected with 40 pg of HBXH10-vpu+/env+ (lane 1)orHXBc2(lane2),andparticulatematerial released into thesupernatantswaspelleted througha20%sucrose cushionat27,000rpmfor 2.5 hat4°CinaBeckmanSW41 Tirotor.ThepelletsweredisruptedinLaemmlisamplebufferanddirectlyanalyzed
bySDS-PAGE. NC, nucleocapsidprotein.(C)Virusexport. HeLa-T4andHeLa-T8weretransfected with10 pgofHXBH10-vpu+/env+ or HXBH10-vpu-/env+ provirus(top).Inparallel, HeLa-T4 andHeLa-T8weretransfected with 10 pg ofHXBH10-vpu+/env-or HXBH10-vpu-/env- (bottom). At the indicated times, virus export was determined by measurement of virion-associated p24 in the cell-free supematants.Data represent theresultsfromoneofthree identicalexperiments. (D)Immunoprecipitation. HeLa-T4cellstransfected with the indicated proviruses were pulse-labeled for 2.5 (odd-numbered lanes) and 5 (even-numbered lanes) h with
[35S]methionine
60 h posttransfection. Bothcells andclarifiedsupernatantswerelysedandimmunoprecipitated withamixofHIV-1-positivehumanserumand VOL.66,1992on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 2. Effect ofvpu on exportofviralparticlesintransa RTactivity (103cpm)
vpuexpressor
HXBH10-vpuY/env+
HXBH10-vpu-/env-SVCMVvpu+ 14.7 60.1
SVCMVvpu- 4.8 14.3
ISVCMVvpu+ orSVCMVvpu-expression plasmid (7.5jig)was
cotrans-fectedwith 10pgofHXBH10-vpu+/env- or10 ,gof
HXBHl1-vpuf/env-(molar ratio of2:1)in HeLa-T8 cells; 48 h posttransfection, viral particle
exportwasdeterminedby measuringreversetranscriptase(RT) activity in the cell-freesupernatant.Background values have been subtracted.
cells transfected with the Vpu+/Env- DNA than in the supernatant of MT4cells transfected withthe
Vpu-/Env-DNA (compare lanes 1 and 2). As described in previous
studies (11, 20, 22), the increase in p24 released in the supernatantisaccompanied byareduction in theamountof cell-associated p24 (Fig. 3B; compare lanes 1 and 2). As
expected, the envelopeglycoproteinsarenotdetected inthe
cell lysate or in the cell-free supernatant of MT4 cells transfected with the Vpu+/Env- and Vpu-/Env-
con-structs.
vpu acts intrans. To determinewhether the vpu protein
actsintranstoincreaseexportof virion proteins, the ability ofaplasmid thatexpresses vputoincrease the level ofvirion
protein present in the supernatant of cells transfectedwith
theVpu-/Env+and Vpu-/Env- DNAswas examined. For
thisexperiment, the openreadingframeexpressingthe vpu
10 20 30 40 50 6C0 proteinwas placed3' to a cytomegalovirus promoteron a
separateplasmid. As acontrol, thesamevpu openreading
framein which the initiation codonATGis alteredtoACG
H ou rs afte r transfecti on was used (Fig. 1B). The vpu expression plasmids and the CELL LYSATE SUPERNATANT HIV-1 proviruses were introduced by cotransfection into
HeLa-T8cells.
B Thedata in Table 2show that the vpu
protein
increases theCo co E I CD @ EQamountofvirusexportedfrom cells,asmeasuredbyrelease
I X> > of theviralDNApolymerase activity into the cell
superna-tant, three- to fourfold. TheVpu- expressorplasmiddoes
notaltertheamountof viral DNApolymerasepresentin the
supematant oftheexpressorcells. This trans effect ofvpu
can be observed with both the HXBH10-vpu-/env+ and
-,s_ HXBH10-vpu-/env-constructs.
Determination of the amount of viral protein released in
''5 'K'JFA' .*.the supernatant of the transfectedcells by
immunoprecipi-tation confirms the results obtained by measurement of
reversetranscriptase activity(Fig. 4). Cotransfection of the
Vpu+expressorplasmid results in a significantincrease of
viralproteinexportin the cellsupernatant(lane 1)compared
with the level seen after cotransfection with the control
plasmid (lane 2). Incontrast,the expressionofvpudoesnot affect significantly theamount of capsid and envelope
gly-coproteinspresentinthe cell lysates (lanes 1 and 2). The experiments reported here demonstrate that vpu-p24 4vm_
supematants were collected and virion-associated reverse
tran-scriptase (RT) activitywasmeasured.(B)Cells from the MT4 cell
culturestransfected withHXBH10-vpu+/env- (lane1)or
HXBH10-vwu-/env- (lane 2) described above were labeled for 5 h with
[25S]methionine
60 h posttransfection. Both cells(left panel) and the12 3 1 2 3 correspondingcell-free supernatants (right panel)werelysed,
im-ole ofenvelope glycoproteinsinvpu-mediated capsid munoprecipitated with anHIV-1-positive human serum, and
ana-rtin CD4+T-cell line MT4. (A) MT4cells(107)were lyzedon anSDS-12.5%polyacrylamide gel. Themock-transfected
with 12 Lg of HXBH10-vpu+/env- (0) or HXBH10- MT4cellcontrol is shown in lane 3 of eachpanel. The position of
*). At each posttransfection time indicated, cell-free HIV-1viral protein is indicated.
I
A
18
1 5
0 0
v- 1 2
09
t
9
I-%._.
rr
cc
3
0
FIG. 3. R
protein
expo transfectedNvpu-/env-
(4
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.322.561.88.142.2]NOTES 5125
gpl60
gpl20
M1 2 1
gpl20--2
p55
-
p41-p24 - abw
p24-r
pl17
I*
pi17- _w [image:7.612.99.245.78.310.2]Cells
Medium
FIG. 4. transeffectofvpu.Twentymicrograms ofSVCMVvpu+
(lane 1) orSVCMVvpu- (lane 2)was cotransfected with 30 pg of HXBH10-vpu-/env+ inHeLa cells. Forty-eight hours posttransfec-tion, cells were metabolicallylabeled for 12 h with [sS]cysteine. Bothcells and the corresponding clarified supematantswere then
lysed, immunoprecipitated with an HIV-1-positive human serum,
and analyzed on an SDS-11.5% polyacrylamide gel. The mock-transfected HeLa cellsareshownin lane M. The positions of HIV-1
viral proteinsareindicated.
mediatedexportof the capsid proteins isindependentofthe expression of the envelope glycoproteins. The amount of
capsidproteins released intothe supernatant fluids of cells
transfected with viral DNA is dramatically increased in the presence of a functional vpu regardless of coexpression of the envelope glycoprotein from the same genome. The
dramatic effect ofvpuoncapsidexport,independent ofenv,
wasdetermined bymeasurementof theamountof p24inthe culturesupematantbyuseofradioimmune competition and
precipitationassays.Theincrease inexportofvirusparticles wasdetermined bymeasurementof theamountofassembled
as well as infectious virus particles in culture supernatant fluids.
The experiments reported here also show that
coexpres-sion of CD4 in the target cell has no measurable effect on
vpu-directed increase inexportofcapsid proteins,atleastin this experimental context. Expression of the vpu protein
significantly increases the level ofcapsid proteinspresentin the supernatantof HeLacells which constitutively express
either the CD4 or CD8 protein. An effect of vpu on the
biosynthesis and export of envelope glycoprotein in cells whichexpress CD4 isnotfound in these experiments. The levels of cell-associated gp160 and gpl20 are similar in
HeLa-T4 cellstransfected withVpu+/Env+andVpu-/Env+ viralDNAs.
There is a significant difference between the effect on
capsid proteins ofvpuinHeLacells compared withits effect in CD4+ T-cell lines. In HeLa cells, the amount of capsid proteininthecell lysateisnotmeasurably decreasedbyvpu.
detected in the cell lysates. This difference is probably more apparentthan real. In HeLa cells, the ratio of cell-associated to supernatant capsid protein in the presence of vpu is only about 20:1. In the absence of vpu, almost no capsid protein is released from the cell. In contrast, in T lymphocytes, the
fraction ofcapsid proteins released from the cell increases from about 20% to almost 80% in the presence of vpu (22). The experiments also demonstrate that vpu acts intransto facilitate export of capsid proteins made from viral DNA defectiveinvpu.The amountof capsid proteinreleasedinto the cell supernatant increases by at least fivefold in the presence of vpu. This observation should permit character-ization of sequences in the vpu protein as well as sequences within the capsid proteins and their precursors that are required for vpu-facilitated
export.
WethankJacques Friborg and TanyaDorfmanfor helpful discus-sions. The following reagents were obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NLAID: HeLa-T4 (HT-6C) from Bruce Chesebro and HeLa-T8 from RichardAxel.
E.A.C. isarecipientof a career award from the National Health ResearchandDevelopment Program (NHRDP) of Canada. H.G. is aSheldon Andelson AmFAR Scholar. This work was supported by a NHRDP/MRC grant to E.A.C. and by National Institutes of HealthgrantAI29873 to H.G. W.A.H. is supported by NCDDGAI
24845 and Cancer Center grant P30 CA06516 and by Center for AIDS Researchgrant P30 AI28691. W.A.H. and E.A.C. are recip-ientsofaNATOcollaborative research grant.
REFERENCES
1. Arrigo, S. J., and I. S. Y. Chen. 1991. Rev is necessary for translation but not cytoplasmic accumulation of HIV-1 vif, vpr, andenv/vpu 2RNAs. Genes Dev. 5:808-819.
2. Boulerice, F.,and M. A. Wainberg. 1990. Inhibition by AZT of HIV-1 replication in acutely infected U-937 cells. J. Leukocyte Biol. 47:498-505.
3. Chesebro, B.,andK.Wehrly. 1988. Development of a sensitive quantitative focal assay for human immunodeficiency virus infectivity. J.Virol. 62:3779-3788.
4. Cohen, E. A., E. F. Terwilliger, J. G. Sodroski, and W. A. Haseltine. 1989. Identification of a protein encoded by the vpu geneofHIV-1. Nature (London) 334:532-534.
5. Fisher, A. G., E. Colleti, L. Ratner, R. C. Gallo, and F. Wong-Staal. 1985. AmolecularcloneofHTLV-3 with biological activity. Nature (London) 316:262-265.
6. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomeswhich express chloramphenicol acetyl-transferaseinmammalian cells. Mol. Cell. Biol. 2:1044-1051. 7. Graham, F. L., andA. J.vanderEb.1973. A new technique for
theassay ofinfectivity ofhuman adenovirus 5 DNA. Virology 52:456-457.
8. Harada,S., Y.Koyanagi, and N. Yamamoto. 1985. Infection of HTLV-III/LAV in HTLV-1-carryingcells MT-2 and MT-4 and application in aplaque assay. Science229:563-566.
9. Helseth, E.,M.Kowalski, D.Gabuzda,U.
Olshevsky,
W. Hasel-tine, and J. Sodroski. 1990. Rapid complementation assays measuringreplicativepotentialofHIV-1 envelope glycoprotein mutants. J. Virol.64:2415-2420.10. Jeang, K.T.,D.R.Rawlins, P. J. Rosenfeld, J. H. Shero, T. J.
Kelly, and G. S. Hayward. 1987. Multiple tandemly repeated binding sites forcellularnuclearfactor 1 that surround the major
immediate-early
promotersof Simian and human cytomegalovi-rus. J.Virol.61:1559-1570.11. Klimkait,T., K.Strebel,M.D.Hoggan, M. A. Martin, and J. M. Orenstein. 1990.Thehumanimmunodeficiency virus type 1-spe-cificprotein vpu isrequired for efficient virus maturation and release.J. Virol. 64:621-629.
12. Kowalski,M.,J.Potz, L.Basiripour, T. Dorfman, W. C. Goh, E. VOL.66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
ciency virus envelopeglycoprotein. Science 237:1351-1355. 13. Lee, M. H., K. Sano, F. E. Morales, and D. T. Imagawa.1987.
Sensitive reverse transcriptaseassay to detect and quantitate human immunodeficiency virus. J. Clin. Microbiol. 25:1717-1721.
14. Maddon, P. J., A. G.Daigleish, J.S.McDougal,P. R.Clapham, R.A. Weiss, and R.Axel.1986.The T4geneencodes theAIDS virusreceptorand isexpressed in the immunesystemand the brain. Cell 47:333-348.
15. Matsuda, Z., M.-J. Chou, M. Matsuda, J.-H. Huang, Y.-M. Chen, R. Redfield,K.Mayer, M. Essex, and T.-H. Lee. 1988. Humanimmunodeficiencyvirustype1hasanadditionalcoding
sequencein the centralregionofthegenome.Proc. Natl.Acad. Sci. USA 35:6968-6972.
16. Myers, G., S. F. Josephs, A. B. Rabson, T. F. Smith,and F. Wong-Staal (ed.). 1988. Human retroviruses and AIDS. Los Alamos NationalLaboratory,LosAlamos,N. Mex.
17. Scharf, S. J., G. T. Horn, and H. A. Erlich. 1986. Direct cloning andsequence analysisofenzymatically amplified genomic
se-quences.Science 233:1076-1078.
18. Schwartz, S., B. K. Felber, E.-M. Fenyo,andG. N. Paviakis. 1990.Env andvpuproteinsof human immunodeficiencyvirus
type1areproducedfrommultiplebicistronic mRNAs. J. Virol.
64:5448-5456.
19. Sodroski, J., W.C. Goh, C. Rosen, A. Tartar, D. Portelie, A. Burny, and W.A. Haseltine.1986. Replicative and cytopathic potential of HTLV-3/LAV with sor gene deletions. Science 231:1549-1553.
20. Strebel, K., T. Klimkait, F. Maldarelli, and M. A. Martin.1989. Molecular and biochemical analyses of human immunodefi-ciencyvirustype1vpuprotein.J.Virol. 63:3784-3791.
21. Strebel, K., T. Klimkait, and M. A. Martin. 1988. A novelgene
ofHIV-1,vpu,and its 16-kilodaltonproduct. Science 241:1221-1223.
22. Terwilliger, E. F., E. A. Cohen, Y.-C. Lu,J.G. Sodroski,and
W. A. Haseltine. 1989. Functional role of human immunodefi-ciencyvirustype 1vpu. Proc. Natl.Acad. Sci. USA
86:5163-5167.
23. Willey, R. L., F. Maldarelli, M. A. Martin, and K. Strebel.1992. Humanimmunodeficiencyvirustype1vpuprotein regulatesthe
formationof intracellular gpl6O-CD4 complexes. J. Virol. 66: 226-234.
24. Zoller, M. J., and M. Smith. 1982. Oligonucleotide-directed mutagenesis using M13-derivedvectors:anefficient and general
procedurefor theproduction of pointmutations inanyfragment
of DNA. Nucleic AcidsRes. 10:6487-6500.