0022-538X/93/042317-10$02.00/0
CopyrightC 1993, AmericanSociety for Microbiology
Multiple Positive and Negative cis-Acting Elements
That Mediate
Transactivation by bell in the
Long
Terminal
Repeat of Human
Foamy
Virus
KIJEONGLEE, ANN HWEELEE,ANDYOUNGCHUL SUNG* Department of Life Science, Center for Biofunctional Molecules, Pohang Institute
of
ScienceandTechnology,
Pohang, Republic
of
KoreaReceived 28 September 1992/Accepted 10January 1993
The bell protein of human foamyvirus (HFV), aretrovirus, regulates expression ofthegenelinkedtothe HFVlongterminalrepeat(LTR)and is essential for viralgeneexpression. The mechanism of actionof thebell
protein isunknown, but its action is mediated through the U3 region of the LTR. To determine which U3
sequences arecritical for transactivationby bell,aseries of hybridvectorsconsistingofamutantHFV LTR andthechloramphenicolacetyltransferasegenewereconstructed and tested for their responsivenesstothebell protein by using transientassays after transfection. The target sequences for transactivation by bell were
mappedtofive regions in the U3 domain ofthe LTR: nucleotides -559to-506, -454to-418, -360to-342, -327 to -284, and -116 to -89 (+1 represents the transcription initiation site). No significant sequence
similaritywasidentifiedamongthe fivetargetsites.The observation that the multiple distinct elements in the
HFV LTRarethetargets for bell transactivation is different fromobservations withother human retroviral systems.The regulation mechanism of HFV bell protein-mediated transactivationappearstobeanalogousto that ofsomeDNAvirustransactivators that increase transcription from numerousdifferent viralpromoters with little sequencesimilarity shared amongthem.Wedemonstratedthat multiplebell-responsiveelements (BRE) canactasbell-dependent enhancerelements, whilea singlecopyofoneBRE,BREe, can serve as an
upstream activating element in both orientations. In addition, the region between -466 and -498 was identified as responsible for the downregulation ofgene expression directed by BREa, which requires its
upstream sequenceelementtoactas abel1-dependent enhancer elementinaheterologouspromoter.
Humanfoamy virus (HFV) isamember of the subfamily Spumavirinae of the family Retroviridae. HFV has been detected in specimens from patients with nasopharyngeal carcinoma(1), chronic myeloid leukemia(42), toxic enceph-alopathy (5), de Quervain's thyroiditis (37, 40), and non-A, non-B hepatitis after blood transfusions (29). However, the role of HFV in thepathogenesis of these diseases remainsto
be determined (39). The full genome of HFV has been
molecularly cloned and sequenced (8, 24). Its genomic organization shows striking similarity to those of other humanretroviruses, especially human T-cell leukemia virus type I (HTLV-I)and humanimmunodeficiencyvirustype1 (HIV-1).
The nucleotide sequence analysis shows that the HFV
genomecontainsatleast threeopenreading frames,termed bell,bel2,andbel3, inadditiontogag,pol, andenv,which
arewell conservedamongretroviralgenomes.The bellgene
encodes a protein functioning as a transactivator for tran-scription directed bythe HFV long terminal repeat (LTR) (31).bell has been showntobe essentialfor virusreplication invitro (22). HIV-1 and HTLV-I also encode an essential
transactivator whichrequiresatargetelement in the LTR(3, 4, 32, 34, 36). HIV-1 tat stimulates LTR-directed gene expression by directly interacting with an RNA target
se-quence termed TAR located downstream of the promoter, positions +19 to +42 (7, 13, 15). Efficient transcription depends on the specific sequence, secondary structure, location, and orientation of the target element (33). In HTLV-I, thetax protein transactivates LTR-directed
tran-* Correspondingauthor.
scriptionviaa21-bpdirectrepeatsequencelocated between -252 and -75 in theU3regionof the LTR(4,34). However, thetaxproteinhasnotbeen showntobind DNAdirectlyand is instead believedtoactivate LTR-directedtranscription via currentlyunidentifiedcellularDNAbinding proteins (9, 23, 27). Previous reports revealed that the bell gene of HFV
encodes a transcriptional transactivator which stimulates
HFVLTR-directed gene expression. Also, the target sites for bell were roughly mapped to two regions in the U3 domain of the LTR (17, 38).
To determine theprecisetargetsequencesfor the action of the bell protein, wehavesystematically analyzedthe HFV LTRby mutagenesis. Furthermore, hybrid promoters con-taining the whole HFV LTR or a part thereof and the enhancerless thymidinekinase(tk) promoter ofherpes sim-plexvirus (HSV)or the enhancerless c-fospromoter were
made. By using transient expression assays, the target elements forbell,termed bell-responsive elements(BRE), weremappedtofiveregionsinthe U3 domain of theLTR. In addition, we have shown that the region consisting of nucleotides -498 to -466 (the -498 to -466 region)
con-tains a negative regulatory element which downregulates BREa-directedgeneexpressioninaheterologous promoter.
MATERIALS AND METHODS
Plasmid construction.Nucleic acid restriction and modify-ing enzymes were used according to the suppliers'
(Pro-mega, Boehringer Mannheim Biochemicals, and KOSCO) recommendations. Oligonucleotides were synthesizedwith an Applied Biosystems DNA synthesizer and purified by denaturing polyacrylamide gel electrophoresis. Plasmid
2317
on November 9, 2019 by guest
http://jvi.asm.org/
HFV-CAT was constructed
by
inserting
theKpnI-NarI
DNAfragment (-777
to+351)
upstream of the bacterialchloramphenicol acetyltransferase
(CAT)
gene,followedby
the simian virus 40
poly(A)
sequence. HFV-CAT-M andHFV-CAT-MI
(see Fig. 3)
wereconstructedby generating
SmaI
and EcoRI sites at nucleotides -149 and -74 of HFV-CAT and aHincII
site at nucleotide -119 ofHFV-CAT-M, respectively,
withthe Muta-Genephagemid
in vitromutagenesis
system(primers
used: 5'-CACClTl
CCC CCG GGC AAC T-3' forSmaI,
5'-TTA CTA TAG AAT TCC TTA A-3' forEcoRI,
and 5'-TCT GCG TTA ACG AAA GAT TCG-3' forHinclI[Bio-Rad]) (20).
Underlined nucle-otidesweremutatedtogenerate therestrictionenzymesites. The 5' and 3' deletion mutants wereconstructedby
firstcleaving plasmids
HFV-CAT and HFV-CAT-M withXhoI andEcoRI,
respectively,
andthenincubating
the linearized DNA with BAL 31 exonuclease.Digested
DNAs were removed at varioustimes,
and theprotruding
ends were filledby using
the Klenowfragment
of DNApolymerase
I.The end-filled DNAs were
ligated
withKpnI
linker for 5'deletion mutants and with EcoRI linker for 3' deletion mutants.
Next,
the DNAswere doubledigested
withKpnI
andClaI
for 5' deletion mutants orXhoI and EcoRI for 3'deletionmutants.Deleted
fragments
ofthe HFV LTRweregel
purified andreplaced by
thecorresponding
fragments
of HFV-CAT and HFV-CAT-Mdigested with
the same en-zymes. Each deletion mutant wasanalyzed by
restrictiondigestions
and double-stranded DNAsequencing
with prim-erscomplementary
to either the Rregion
ofthe HFV LTR(+54
to+72;
5'-CAA TAT AAA ATACTT-3')
or the M13 forwardprimer (Promega).
Plasmid-11/+351
was con-structedby deleting
theXbaI-ClaI
fragment (nucleotides
-11 to
+351)
of HFV-CAT.Internal deletion mutants were constructed
by
treatmentwith
appropriate
restrictionand/or
modifying
enzymes andligation
with T4 DNAligase. HFV-IC(Ac)
andHFV-IICE(Ac,e)
were constructedby digesting plasmids
HFV-CAT andHFV-IE(Ae), respectively,
withHindIIIandtreat-ing
theresulting digests
with BAL31 exonuclease. Deletedplasmids
weremade flushwiththe Klenowfragment
of DNApolymerase
I andreligated
with T4 DNAligase
in the presence of SmaI linker.HFV-IA(Aa*)
andHFV-IE(Ae)
were obtained
by deleting
theBstEII-SalI(-570
to-506)
and SmaI-EcoRI
(-149
to-74)
DNAfragments
fromHFVAp-1MII
(see below)
andHFV-CAT-M, respectively.
HFV-IIBC(Ab,c)
was constructedby inserting
theKpnI-AvaII
(-777
to-434) fragment
ofHFV-CAT into theKpnI site ofplasmid
-341/+351.
To obtainplasmid
HFV-III(Aa,c,e),
aSall site wasgenerated
atposition
-512 with respect tothewild-type
LTRtranscription
initiation siteofHFV-IICE(Ac,e) by oligonucleotide-directed
invitromuta-genesis (5'-CTA
ATT TCATCCTGTLCGLACTC
TCTGTC AATG-3')
followedby
deletion of theBstEII-SalI
DNAfragment (-570
to-506).
HFV-IV(Aa,b,c,e)
andHFV-V(Aa,b,c,d,e)
were constructed from HFV-IICE(Ac,e) and HFV-CAT-Mby deleting
theBstEII-SmaI (-570 to -302) and BstEII-EcoRI(-570
to -74) fragments, respectively.HFV-ANRE was constructed by deleting the Sall-SmaI
(-508
to-454)
DNAfragment
ofHFVAp-1MII.The AP-1 mutants
HFVAp-1MI
andHFVAp-1MII
were constructedby oligonucleotide-directed
in vitro mutagene-sis. Theprimers
used were 5'-CTA ATT TCA TCCT(I
CGA
CTC TCT GTC AAT G-3'(HFVAp-lMII)
and5'-GGG TCC ATC TCG AGC ACT TCC CCG GGC AAT GAAGGG-3'
(HFVAp-lMI
andHFVAp-1MII).
Hybrids
of HFV LTRregions
with the truncated tkpromoterof HSV and with the c-fos promoter were gener-ated by placing the HFV LTR either upstream or down-stream of the enhancerless tk promoter or the c-fos pro-moter. The configuration of LTR along the tk and c-fos promoter in each hybrid plasmid is schematically drawn in Fig. 6A. LTR-tk-U, LTR-tk-U-R, and LTR-tk-D were con-structedbyinserting the entire HFV LTR (nucleotides -777
to +351)into theSall site in both orientations and the SmaI
site of -37tk CAT (6), respectively. fos-U and LTR-fos-Dwereconstructedby inserting the HFV LTR (-777to
+351)into theSalI site andXhoIsite of A-71c-fos CAT (11,
28), respectively.
BRE(a)-tk and BRE(a)-tk-R were constructed by subclon-ing the BstEII-EcoRI fragment (-570 to -506) ofplasmid
-777/-506into theXbaI site of -37tk CAT in the same and
oppositeorientations with respect tothe promoter,
respec-tively. BRE(a')-tkwasconstructedby placing the
KpnI-SalI
DNA fragment (-777 to -509) of HFVAp-1MII into the XbaI site of -37tk CAT in the same orientation. BRE(a)-tk-D andBRE(a2)-tk-Dwereobtainedby inserting thesame BstEII-EcoRIfragment into theSmaI site of -37tk CAT as a monomer and a dimer, respectively. BRE(b)-tkwas ob-tained by inserting the SmaI-HindIII fragment (-453 to
-360)ofplasmidHFVAp-1MIIinto theXbaI site of -37tk
CAT. To construct BRE(c)-tk and BRE(c3)-tk, synthetic
oligonucleotides (5'-AGC Tfl-T CAC ATA CTC AGT AGC
TGT TT-3' and its partially complementary sequence
5'-CTA GAA ACA GCT ACT GAG TAT GTGAA-3')
contain-ing a portion of the HFV LTR (-360 to -335) were
prepared.Thecomplementary oligonucleotidesweretreated
with T4polynucleotide kinase, hybridized,andinserted into the XbaI site of -37tk CAT. BRE(d)-tk, BRE(dl)-tk,
BRE(d2)-tk, and BRE(d3)-tk contain the
Kp4nI-Hinfl
DNAfragments of plasmids -342/+351 (-342 to -127), -284/
+351
(-284
to-127),
-249/+351 (-249 to -127), and-182/+351 (-182to -127), respectively, attheXbaI site of
-37tk CAT in the orientation opposite to that of the pro-moter. BRE(d')-tk was constructed by inserting the DNA
fragment containing the -327 to -276 region, which was
prepared by polymerasechainreaction,into theXbaI site of
-37tk CAT in the opposite orientation. BRE(e)-tk and
BRE(e)-tk-Rwereconstructedby insertingthe SmaI-EcoRI
fragment (-149to-74)of HFV-CAT-MintotheXbaI site of
-37tk CAT in the same and opposite orientations,
respec-tively,
with respect to the promoter. BRE(e'1)-tk andBRE(e'2)-tk
wereconstructed byinsertingtheSmaI-HincII(-149 to -117) and HincII-EcoRI (-116 to -74) DNA
fragmentsinto the XbaI site of-37tkCAT in the same and
opposite orientations, respectively. BRE(e)-tk-D,
BRE(e)-tk-DR,andBRE(e2)-tk-DcontaintheSmaI-EcoRIfragment
at the SmaI site of -37tk CAT in the same and opposite
orientations and as a dimer with respect to the promoter.
Theorientation of eachfragmentin allhybrid constructs was
confirmed by double-stranded DNA sequencing with the
CAT gene
primer
(5'-GGC CGT AAT ATC CAG G-3') or reverseprimer (Promega).
BREa/NRE-tkwas obtained byinserting
the BstEII-EcoRI fragment (-570 to -466) ofplasmid-777/-466 into the XbaI site of -37tk CAT. BREe/
NRE-tk-R, BREe/NRE-tk-R', and NRE/RSV-CAT were
constructed by inserting the SalI-SmaI fragment (-512 to
-435)
ofHFVAp-1MII
into the SalI site ofBRE(e)-tk-R inthesameandoppositeorientations and into theNdeIsite of
pRSV-CAT, respectively.
Cell culture and transfection assay. BHK-21 cells were grown in Dulbecco's modified Eagle's medium supple-mented with 10% fetal calf serum. Plasmid transfections
on November 9, 2019 by guest
http://jvi.asm.org/
x
U3 RU CAT Poly(A) |
~~b ~TATA
777 7 a
+11
m
IBFV-CAT -561/+351 -506/+351
i I
i I
CAT Activity % conversion bol-A bol-S <0.1 275 <0.1 182 <0.1 107 <0.1 165 <0.1 65.2 <0.1 87.5 <0.1 85.8 <0.1 10.7 <0.1 8.7 <0.1 7.5 <0.1 6.8 <0.1
<0.1 <0.1
I I <0.1
8.1 7.3 7.4
2.5
i <0.1 0.2
i -I <0.1 <0.1
I-I <0.1 <0.1
FIG. 1. Schematicdiagramof thewild-typeHFV LTRand the 5' deletionmutantsof theHFV LTRand theirresponsestobell.Sequential
5' deletionmutantsweregenerated by controlled BAL 31 exonuclease digestion from parental plasmid HFV-CAT, and the deleted fragments usedtoconstructCAT fusion plasmidsaredepicted with their 5' endpoints extending from nucleotides -561to-13 (+1 correspondstothe transcription initiation site) and theircommon 3' endpoint at +351. The effectorplasmids pSbell-S (bel-S) and pSBell-A (bel-A)were
constructedby insertinga1,048-bp SspI fragmentofpHSRV comprisingtheentire bellopenreading frame downstream of the simian virus
40earlypromoterin thesenseand antisenseorientations, respectively (31). Regulatory regions defined by 5' sequential deletion analysisare
indicatedby theopenboxes (BREc, -360to-342; BREe, -127to-89). The solid vertical bars and the solid ovalrepresentthe AP-1 binding
sites and TATAbox, respectively. Deletion mutantsweretransfected into BHK-21 cells and levels of CATactivitywere determinedas
described in Materials and Methods. Restrictionenzymesites:K,KpnI;X, XhoI; N,Narl; C, ClaI.
were carried out by the DEAE-dextran method (30). Cells (106) were plated in a 100-mm-diameter dish a day before
transfection and transfected with 2 p,geach of effector and reporterplasmidsin thepresenceof80,M chloroquine.The cellsweretreated with10%dimethylsulfoxide for 150safter
exposuretothetransfection cocktail.Forty-eighthours after transfection, the cells were harvested and assayed for the expression level of CAT as previouslydescribed (12). The cell extractswere prepared bythree cyclesoffreezingand thawing. The level ofacetylated [14C]chloramphenicolwas
measuredby liquidscintillationcountingof the spotscutout from the plate after the acetylated and unacetylated forms
were separated by thin-layer chromatography. A 30-min time point and 50 ,ug of protein lysates were used for all reactions. In cases in which the level of acetylation of chloramphenicol was more than 80%, the protein lysates were diluted for the reaction and the CAT values were
correctedbythe dilution factor.AllCATassaydatareported inthis articlearefrompointsin the linearrangeof theassay.
The results shown are the averages of a minimum of four separate experiments for each deletion mutant, with stan-dard deviations withinarangeof 20%.Theprotein
concen-trationwasdetermined withaBio-Radproteinassaykit with bovineserum albuminas a standard.
RESULTS
5' and 3' progressive deletion analysis. Previous reports revealed that the bellgeneof HFV encodesatranscriptional transactivator that acts on the U3 region of the LTR. To identify the functional regions responsible for bell action,
we constructed plasmid HFV-CAT, which contains the entire HFV LTR(nucleotides-777to+351)upstreamof the CATgene.A series of deletionsstartingat the 5' end of the LTRwere carried outby controlled digestion of the HFV-CAT DNA withBAL31exonuclease. The structure of each deletion is illustrated in Fig. 1. The activities of individual LTR deletion mutants were determined by measuring the levels of CATactivityafter cotransfectioninto BHK-21 cells either with the bell expression plasmid (pSbell-S) orwith
the controlplasmid (pSbell-A).Theparental plasmid, HFV-CAT,showedmorethan2,750-fold inductionbybell.
Dele-tion of the R andU5 regionsin -11/+351did not haveany effect on the bell response, as expected on the basis of a -443/+351
-412/+351 -380/+351 -360/+351 -342/+351 -284/+351 -249/+351 -182/+351 -174/+351 -169/+351 -133/+351 -130/+351 -127/+351 -106/+351 -89/+351 -53/+351 -13/+351
i i <0.i 11.0
0 s
<0.1 11.0
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.158.470.71.391.2]U3 RU5 CAT I Pol(A)
/.tI.~..4-b1J1IIIIIIIIIIIII2 zj
CAT
Activity
-777 %-i
;, i'W
1 t conversionI
| F
+1
bel-A be1-S
HWV-CAT I--11/+351 1--777/-1491 -777/-186
1--777/-270
--777/-276
--777/-304
--777/-320
--777/-328 -_-, I,IJ3 v.
-777/-357I i
-777/-366I i
-777/-4181 l
-4 <0.1 I <0.1 I I.-I <0.1 I a-I <0.1
---14 I <0.1
----I <0.1
- I-I <0.1
i I <0.1
a I <0.1
I I <0.1 i- <0.1
I.-.--I<0.1
I - <0.1 275 292 248 244 270 260 221 216 242 262 229 168 153
-777/-4401 1 ' <0.1 34.1
-777/-4541 1 1 <0.1 6.5
-777/-4661 -1 l <0.1 6.1
-777/-4981 1 l- <0.1 102
-777/-5061 1
1-1
<0.1 140-777/-5591
-
1 <0.1 <0.1-777/-587i- <0.1 <0.1
FIG. 2. Localization oftwoBREandoneNREby transient-expressionanalysisof 3' deletionmutants.Deletionmutants weregenerated
byBAL31exonucleasedigestionfromthe EcoRIsite(nucleotide-74)toward the 5'direction,with theircommon5' and3'endpointsat-777
to +351,respectively. Each deletionmutantcontainedadeletionwith 5'endpoint varyingfrom -587to-149 anda common3'endpointat -74(+1 denotesthetranscription initiationsite). -11/+351wasconstructedbydeletingtheXbaI-ClaIfragmentofHFV-CAT.Regulatory regions defined by3'serial deletionanalysisareindicatedbythe open boxesand the hatched box:BREa,-559to-506; BREb,-454to-418; NRE, -498to-466. The solidvertical bars and solid ovalaredescribedinthelegendtoFig.1.The deletionmutants weretransfectedinto BHK-21 cells, andlevels of CATactivityweredeterminedasdescribedinMaterials andMethods. Restrictionenzymesites: K,KpnI;X, XhoI;XI,XbaI;C,ClaI; SI,SmaI;E,EcoRI. bel-A andbel-S indicatepSbell-AandpSbell-S, respectively.
previousreport(Fig. 2)(17).Deletions from the 5' end of the
LTRtopositions -561, -506, -443, -412, -380,and -360
resulted in slightly progressive reductions of the bell re-sponsebyuptothreefold(Fig. 1). These reductions may be due to the removal of the target sites for bell or other
regulatory sequences such as AP-1 binding sites that are
required for basal promoter activity(14, 25).In contrast, an
eightfolddecrease in thebell responsewasevident after the
deletion of an additional 18 nucleotides (deletion -342/
+351), butaresidual bell response remained. The level of
CAT activity remained relatively constant from this point
until the deletionwas extended up to -127 from the tran-scription start site, indicating that the DNA sequence 3' downstream toposition -127 still contains sufficient infor-mationtomediatebell transactivation. However, deletion of a further 38 nucleotides, resulting in plasmid -89/+351, almostcompletely abolished the bell response. The results from the 5' deletion analysis demonstrate that the ability of the HFV LTRtorespond tobell is dependent on at least two
specific sequences located at positions -360 to -342 and
-127 to -89. We propose that these elements be termed BREc and BREe, respectively.
To confirm the two regions, BREc and BREe, identified by the 5' deletion analysis, we generated an EcoRI site at
position -74 by oligonucleotide-directed mutagenesis
(HFV-CAT-M) and introduced a series of deletions toward
the 5' side starting atthe EcoRI site (Fig. 2). Interestingly,
deletion of positions -74 to -418 (deletion -777/-418),
includingboth BREc andBREe, didnotshow anysignificant
effectonthebellresponse,suggestingthat neither BREcnor
BREeisabsolutelyrequiredfor thebellresponse. Afurther
36-bp deletion (-777/-454) resulted in a considerable de-crease in CAT activity, with some of the bell response retained. Surprisingly, deletion ofpositions -466 to -498 resulted in an increase in bell-mediated transactivation of about16-fold, suggestingthat thisregioncontainsanegative
regulatorysequencewhichdownregulates the bellresponse.
Successive deletions to -506 resulted in only a slight in-crease in the bell response. In contrast, a deletion of a further53 nucleotides to -559completely shut off
transac-tivation by bell. These unexpected findings suggest that
there are twomore targetsites forbell (positions -454 to -418 and -559to -506) as well as one negative regulatory elementatpositions -498to -466. We propose that the two additional targetsites forbell,positions -454 to -418 and
-559 to -506, be termed BREb and BREa, respectively,
and that thenegative regulatory element, positions -498 to -466, be termed NRE.
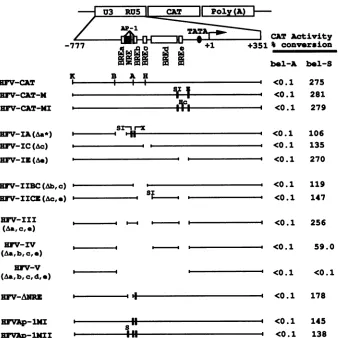
Internal deletion and point mutation
analysis.
Takento-gether, the results obtainedby sequential deletion analysis
identified four positive elements and one negative element
responsible for the regulation of bell transactivation.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.148.466.75.372.2]J
UJ3RUJ5
1I CAT 1IPOlY
(A) IA CAT Activity
-777 "
HZ
w +1 +351% conversionMM MM i
I
bel-A bel-SNFV-CAT NFV-CAT-K HNV-CAT-KI
Y-TIA(Aa*)
BV-IC(Ac)
NFV-IZ(A.) NFV-IIBC(Ab, c)
UvIICZ(Ac,.)
EV-III (a,c, ) WFv-Iv (Aa,b,c,a)
W3V-V
(A,b,c,d,
*)
BUV-ANRE
NFVAp-lMII
1 B A 1
i
III
,
~~~~~~~~~
T1
SI_~MC
i I - i
8I
I.-4I I I
I~~~~F- i . F
I8o
--<0.1 275 <0.1 281 <0.1 279 <0.1 106 <0.1 135 <0.1 270 <0.1 119 <0.1 147 <0.1 256 <0.1 59.0 <0.1 <0.1
<0.1 178 <0.1 145 <0.1 138
FIG. 3. Effects ofdeletionsof putative BRE and NRE on HFV LTR promoter activity. Internaldeletion mutants were constructed by
treatmentwithappropriate restrictionand/or modifying enzymes. The deleted regions of each deletion mutant are as follows:
HFV-IA(A&a*),
nucleotide-570 to -506; HFV-IC(Ac), -365 to -322;HFV-IE(Ae), -149 to -74; HFV-IIBC(Ab,c), -432 to -342; HFV-IICE(Ac,e), -403 to-302 and -149 to-74;HFV-III(Aa,c,e), -570 to -506, -403 to -302, and -149 to -74;HFV-IV(Aa,b,c,e),-570 to -302 and -149 to -74; HFV-V(Aa,b,c,d,e), -570 to -74; HFV-ANRE, -508 to -454. The point mutants containing the mutated AP-1 binding sites, HFVAp-1MI andHFVAp-IMII, were generated by oligonucleotide-directed in vitro mutagenesis. Point mutations are indicated as solid vertical bars in themapsof the mutants. Open boxes represent the putative BRE. All of the BRE except BREd are described in the legends toFig.1and2, and theBREd maps to the region from -302 to -149. These deletion mutants were transfected into BHK-21 cells, and levels of CATactivityweredeterminedasdescribed inMaterials and Methods. Restrictionenzymesites: K,
KpnI;
B, BstEII; A,AvaII;H,HindIII;Hc,HincII; SI,SmaI;E, EcoRI; X, XhoI;
S,
SalI.bel-A and bel-S refer to pSbell-A and pSbell-S, respectively.quential deletion mutagenesis could notidentify redundant
elements in whichasinglecopyis sufficient to activate gene
expression. In addition, 3' sequential deletion mutagenesis
mighthave resulted in thejuxtaposition of distal sequences,
resultingin artifactualactivationorinactivation of
transcrip-tion. To investigate whether there are unidentified target
sites forbell,wegeneratedinternaldeletion mutantswhich
havedeletions of one, two,three,orallof the BREidentified
by progressive deletion. Combinatorial deletions of one,
two,orthree of the BRE didnotshowanysignificant effect
on thebell response (Fig. 3). Interestingly, plasmid
HFV-IV(Aa,b,c,e)
from which all four BRE are absent, retainedbell-mediated transactivation, albeit at a lower level than
that of thewild-typeHFV-CATplasmid. Bycontrast,
addi-tional deletion of a region from positions -302 to -149,
resulting inplasmid
HFV-V(Aa,b,c,d,e),
resulted inacom-plete lossof transactivationbybell (Fig. 3).These
experi-ments indicate that the region between -302 and -149 contains thefifth target site forbelltransactivation (BREd).
To investigate the role of NRE in the HFV LTR as a
whole, we constructed HFV-ANRE by deleting the
SalI-SmaIDNAfragmentofHFVAp-lMIIandassayed it for the
bellresponse(Fig. 3).Deletion of NRE appears to have little influenceonthebell-mediatedtransactivation of the whole LTR.
Since the deletion from the 5' end to position -360,
including three AP-1 binding sites, showed a moderate
decrease in thebellresponse (Fig. 1),wetested the role of these AP-1binding sites in the bell response. Mutationsin
these AP-1 binding sites (HFVAp-lMI and HFVAp-lMII)
led to anapproximately twofold decrease in bell-mediated
transactivation, suggesting that three AP-1 binding sites
contribute to the optimal activity of the HFV promoter. These resultsareingoodagreementwith those ofa
previous
report(25).
Effect of BRE and NRE in a heterologous promoter. To determine whether the sequences
mapped
asthe targetsites forbell were themselves sufficient for the bell response in the context ofaheterologous
promoter, we subcloned theDNA fragments containing each BRE upstream of the
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.146.484.76.414.2]EIFV LTR 4 JA II
-777 Id IQ U w +1 +351
H
m
CAT activity(% conversion)
-37tk
CAT
ll-IFikjIIIATII---777 -509
/|mz CAT ,>
q/
ERE(a')-tk
A
S
A
S
-570 -506 A
HRE
(a) -tk/-
f j
iZ-Cff
S
-453 -360 A
BRE(b)-tk II t/ CT
-360 -335 A
BRE (c)-tk /
/-S
-360 -335 A
ERE(c3) -tk CAT
S
-127 -342 A
U~~~~~~~
BRE(d)-tk
//MSWtk
A-74-149 A
BRE(e) -tk-R k
-149 -117 A
BRE (e'1)-t k //
-74-116 A
BRE(e'2) -tk / /
-127 -342
BRE(d)-tk /CAT
2.1
* 3.5
* 2.0
*4S
25.2* 21.5
*
* 31.3*
4.5*
*
80.6* 2.2
5.6
* 2.1
*
*
42.3O 4.5
*
B85.8
.
5.2*
p
52.0* 2.3
*
4
4.5
* 3.6
* z
~45.7
CAT activity
(% conversion)
bell-A bell-S
-I1
4.5 85.8-127 -284
BRE(dl) -till CAT >
-127 -249
BRE(d2) -tk
/j
-tkJI7CII----127 -182
HRE(d3) -tk/ic CAT >--K
-276 -327
BRE(d') -tk / t/ /
2.1
1.8
1.7
4.5
4.2
3.9
4.1 75.9
FIG. 4. Transactivation of the enhancerless HSVtkpromotercontaining the putative BRE by bell. Hybridconstructswereconstructed by inserting the fragment containing the putativeBREupstreamofthe enhancerlesstkpromoterofHSV. Numbersrepresentthepositions of the
HFV LTR withrespect toitstranscription initiation site. Arrows indicate the orientation of theDNAfragment relativetothat of theenhancerless
tkpromoter.(A)Schematicdiagram of theHFVLTR-containingmultiple regulatory regionsandhybridconstructs, includingeachputative
BRE,and theirresponsetobell.Aand SrepresentpSbell-A and pSbell-S, respectively.The BREaredescribed in thelegendstoFig. 1, 2,
and 3. CATactivitywasmeasuredaspreviouslydescribed (12)and is indicated beside each lane of theautoradiogram. (B)Finemappingof
BREdbyserial deletionanalysis.The construction of eachplasmidand the transfectionassayaredescribed in Materials and Methods.
2322 B)
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.143.467.56.667.2]hancerless HSV tk promoter andassayed them for thebell response(Fig. 4A).Thetruncated tk promoter contains only 37bpupstreamfrom thetranscriptionstartsitewith only one
transcription element, aTATA box(6).BREainBRE(a)-tk
significantly elevated the basal promoter activity in the
absence of bell. However, no significant induction bybell
wasdetected, indicating that BREa acted as a bell-indepen-dent element in a heterologous promoter. This result sug-geststhat BREarequiresanyotherelement within the LTR in ordertoconfer thebellresponse.To testthispossibility,
we constructed BRE(a')-tk, which contains BREa and its
upstream sequence, and assayed it for the bell response. BREa in BRE(a')-tkfunctionsas a bell-dependentelement
(Fig. 4A), indicatingthat BREa requires itsupstream DNA
sequencein ordertoact as abell-dependentelementinthe context of the heterologous promoter as well as the HFV promoter. By contrast, the plasmids containing BREb,
BREd, and BREe upstream of the enhancerless promoter
showed significant increases in CAT activity bybell,
sug-gestingthattheyaresufficienttoconferthebell
responsive-ness tothetk promoter.
To determine the 5' border of BREd, we subcloned the
K,pnI-Hinfl
DNA fragment of the plasmids -342/+351,-284/+351, -249/+351,
and -182/+351 in the oppositeorientationupstreamof theenhancerlesstk promoter,
result-inginBRE(d)-tk, BRE(dl)-tk, BRE(d2)-tk, and BRE(d3)-tk,
respectively, and assayed them for the bell response.
Plas-mid BRE(d)-tk was transactivated by bell, but plasmids
BRE(dl)-tk,BRE(d2)-tk,andBRE(d3)-tk completely lacked
thebell response
(Fig. 4B).
Todetermine theprecisetarget sequence of BREd for bell transactivation, we placed the -327to -276 DNAfragmentupstreamof theenhancerless tk promoterto generate BRE(d')-tk and assayed it forthe bell response. Plasmid BRE(d')-tk was transactivated bybell,
and the level oftransactivationwascomparabletothat ofBRE(d)-tk.
By combining these observations with the results ofthe internal deletion analysis, we concluded that the fifth target site for bell (BREd)is located in the region between -327 and -284. Inaddition,we subdivided BREeinto twofragments andpositionedthem upstream of the tk
promoter.PlasmidBRE(e'2)-tk, containingthe -116to-74
fragment,wastransactivatedbybell,butplasmid
BRE(e'1)-tk, containing
the-149to-117fragment,was not.Thus,we couldultimatelynarrowBREetotheregion betweennucle-otides -116and -89.
Asinglecopyof BREc inBRE(c)-tkhad littleeffectonthe
level of the CATactivityin thepresenceof bell (Fig. 4A).
However, when three copies of BREc were inserted
up-stream ofthe promoter at the same position, the resulting
plasmid, BRE(c3)-tk, showed a marked increase in CAT
activityinthe presence ofbell (Fig. 4A).
To examine theeffect of NREon twootherBRE, BREa
and BREe, and a
heterologous
viral promoter, the Roussarcomavirus
(RSV) LTR,
weconstructedBREa/NRE-tk,
BREe/NRE-tk-R,
BREe/NRE-tk-R',
andNRE/RSV-CAT
and then
assayed
them for the bell response. NRE inBREa/NRE-tk
completely
abolished the increase in basalpromoter activity directed by BREa in the presence or absence of bell
(Fig. 5). However,
NRE inBREe/NRE-tk-R,
BREe/NRE-tk-R',
andNRE/RSV-CAT
did not showanyeffecton the
ability
oftransactivation of BREe andonthe promoter
activity
of the RSV LTR(Fig. 5).
These results suggestthat NRE canfunctiononly
onadjacent
BREa in aheterologouspromoter.
Multiple BREfunction as a
bell-dependent
enhancer.En-hancer elements have beendefinedas
regulatory
sequencesCAT activity (% oonvrsion)
bell-S
-37tk CALT
-570 -506
ER
(a)-tk/okc
7 RBRZa/NRX-tk
BRZ(o)-tk-R
BRZo/NRZ-tk-R
-512
Be/NR-tk-R
' //1IkI MRSV-CLT
2.1
21.5
2.4
3.5
31.3
3.5
45.8
//T
, 90.1 90.5-512-453
[image:7.612.326.565.80.358.2]NRN/RSV-CAT CA 88.7 91.5
FIG. 5. Effects of NREonBRE andonRSV LTR-directedgene
expression.Theregionbetween nucleotides-570and-466,
includ-ingBREa andNRE,wasplaced upstreamofanenhancerless HSV tk promoter. TheSalI-SmaI fragment (-512 to -453) containing
NREwasplaced upstreamof BREe and the RSV LTR. Numbers above the maps represent the positions of the HFV LTR with
respect to its transcription initiation site. Arrows indicate the orientation of the DNAfragmentrelative to that of the enhancerless
tk promoter. Hatched boxes, hatched ovals, and stippled ovals
representNRE, BREa,andBREe, respectively.Cellswere cotrans-fected with each hybrid construct and abell expression plasmid (pSbell-S) or the control plasmid (pSbell-A). CAT assays were
performed48 h laterasdescribed in Materials and Methods.
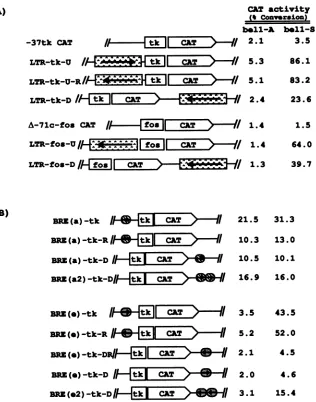
that activate transcription of a linked gene at a distance, regardless of their orientation. To determine whether an HFV LTRcontainingfive BRE andoneNREcould actas an enhancerelement,the entire LTRsequence(-777to+351) was inserted upstream or downstream of the promoter in both orientations into two testplasmids lackinganenhancer, -37tk CAT and A-71c-fos CAT (Fig. 6A). The resulting plasmidsweretransfected intoBHK-21cells withorwithout the bellexpression plasmid, and transientexpressionof the CAT enzymewas usedas a measure of thetranscriptional activity.Asexpected,two controltestplasmidsexhibiteda
basal level of CATactivityand did notrespondto bell(Fig.
6A). In the absence of the bell expression plasmid,all five
recombinantplasmidsgavebackgroundlevels of CAT activ-ity that were about the same as those observed for test plasmids.When cellswerecotransfected withabell expres-sionplasmid, pSbell-S,and the recombinant plasmids
con-tainingthe HFVLTR,CAT activitieswere increased 10- to
50-fold. It is noteworthythat the level of the CATactivity
mediatedbybellwas somewhathigherwhenthe LTR was
placed upstream than downstream of each promoter (Fig.
6A).These data indicate that the HFV LTRcontainingthe
-570-466
A'
itk
-74 -149
-512 -453
1
1=
tI k
I
Cwr-::..
on November 9, 2019 by guest
http://jvi.asm.org/
A) CAT activity (% Converslon) bell-A bell-S
-37tk CAT 2.1
LTR-tk-U | CAT 5.3
LTR-tk-U-R9^^T5.1
LTR-tk-D CAT/ 2.4
A-71c-fos CAT 1.4
LTR-foa-U CATfoQ 1.4
LTR-fos-D/
fo|
CAT/O
1.3B)
3.5 86.1 83.2 23.6 1.5 64.0 39.7
21.5 31.3
BRN
(a)
-tk
/
[
Z
BRZ(a)-tk-R
/
CALT/
BRZ(a)-tk-D
BRN(a2)-tk-D/I-
tklIQT /
BRN(0)-tk
/I|@CZIT>-ff
BRN(0)-tk-R
/
[/
URN(e)-tk-D
I-4Ei
LiQS>-BRN
()-tk-DIH
JL
QT-3 '-IR
BRN(e2)-tk-D10.3 10.5 16.9
3.5 5.2 2.1 2.0 3.1
13.0 10.1 16.0
43.5 52.0 4.5 4.6 15.4
FIG. 6. Characterization of the enhancer element of the HFVLTR.(A)Theentire HFVLTRwasplacedupstreamordownstreamofthe enhancerless HSVtk promoterorc-fospromoterfusedtothe CATgene.Arrowsdenote theorientation of the HFV LTRrelativetothat of theenhancerlesspromoter. (B)The sequencebetweennucleotides -570 and -506containingBREaand theregionbetween-149 and -74 containingBREewereplacedupstreamordownstreamofanenhancerless HSVtk promoterin both orientationswith eitherone ortwocopies
of therespectiveBRE.Cellswerecotransfected with eachhybridconstructandabellexpression plasmid(pSbell-S)orthecontrolplasmid (pSbell-A). CATassayswereperformed 48hlaterasdescribed in Materials and Methods.
five BRE and NRE does function as a bell-dependent enhancer.
Todetermine whether asingle BREcould independently
act as an enhancer element, two of the BRE, BREa and
BREe,wereplaced in both orientations either upstream or
downstreamofthe tk promoter(Fig. 6B). BRE(a)-tk,which
hasasinglecopyofBREa,showed elevated basal promoter
activity, not responding to bell. Also, BRE(a)-tk-R and
BRE(a)-tk-Dshowedelevated basal promoter activity, albeit
at alowerlevelthanBRE(a)-tk.
Interestingly,
twocopies ofBREainBRE(a2)-tk-D conferredmore elevated basal
pro-moter
activity
thanthesinglecopyinBRE(a)-tk-D,suggest-ing that twocopiesof BREaseem tohave acooperative or
additive effect. Plasmids containing one copy of BREe upstreamofthepromoterineither orientationshowed about a10-foldincreaseinCATactivitywhen cotransfectedwitha
bellexpression plasmid (Fig. 6B). Bycontrast,theplasmids
containing a single copy of BREe downstream of the
pro-moterineither orientation didnotshowany
significant
bell response(Fig. 6B).
The observation that both BREa and BREe downstream of the promoter showed a lower bell responseis reminiscent of theCATactivityfound in LTR-tk-D and LTR-fos-D (Fig. 6A). Whentwo copies of BREe were inserted downstream of the promoter at the sameposition, the resulting plasmid showed about a fivefold
increase in CATactivity by bell (Fig. 6B).These observa-tions suggest thattwocopiesofBREe haveacooperativeor
synergistic effect.
DISCUSSION
The results presented heredemonstrate that as manyas
fivedifferentcis-actingelements needed forbellfunctionare
presentin the U3regionof the LTR:positions -559to-506
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.148.464.73.475.2]for BREa, -454 to -418 for BREb, -360 to -342 for BREc, -327 to-284 for BREd, and -116 to -89 for BREe. One of the five BRE, BREa, required its upstream DNA sequence element for bell-dependent transcriptional activation, whereas each of the other four BRE functioned in a bell-dependent manner by itself in a heterologous promoter. Our results arepartially consistent with those of previous work showing two target regions for bell in the U3 domain of the LTR (17, 38). Furthermore, we showed that the -498 to -466region downregulates the gene expression directed by BREain aheterologous promoter.
The HFV LTRcontaining at least five BRE and one NRE as a whole appears to act as a bell-dependent enhancer element. A single copy of BREe activated bell-dependent
transcriptionwhen positionedupstream of the enhancerless
tk promoter but had little detectable function at 1.65-kb downstreamof thetranscription start site,suggesting that a single copy of BREe by itself is sufficient for action only as an upstream activating element. A dimer of BREe was
sufficient to activate the enhancerless tk promoter
down-streamof the CAT geneatthe same position. In addition, a singlesyntheticcopyofBREchad little effect on thelevel of
CATactivitymediatedbybell,whilethreesyntheticcopies
of BREc showed a markedincrease in CAT activity in the presence of bell. The increase in CAT activity was much greater than would be expected if the effects of the two copies of BREa and three copies of BREc were additive,
suggestingthat adjacentBRE sites cooperateto produce a
synergistic effectin the bell transactivation. Since a single
targetsite for bell appearstobesufficientforbell-mediated
transactivation, it will be interesting to determine why an
arrayofBREexists in the U3 regionofthe HFV LTR. The presence of multiple copies ofBRE could be expected to
giveanoptimal level ofLTR-directed geneexpressionandto
have a strong evolutionary advantage; mutations arising in
onesite would becompensatedforby other sitesinthe LTR. The fact that five distinct BRE which have no apparent sequence similarity are the targets forbell transactivation suggests that the mechanism of action of the HFV bell
proteinissimilartothat of some DNA virus transactivators
which increase transcription from numerous differentviral
promoters that havelittle sequencesimilarity. The
adenovi-rus ElA protein, for example, activates transcription from
manydifferent adenovirusearly promoters(16, 26), implying
thattransactivation isnotmediated via direct interactionof
the ElA protein with a specific ElA-responsive promoter
element. In some other promoters, ElA increases the
activ-ity ofafactor that interacts with the TATA box(21, 41).The
generality
ofElAprotein
canbefurther emphasized by itsabilitytostimulatetranscription byRNApolymeraseIII
(18,
19,35). Therefore,it is clear that further studiesare
required
in order to determine the mechanism by which HFVbell
regulates LTR-directed gene expression via at least five
different DNA target sites
(BRE)
and one negativeregula-torysequence
(NRE).
There are a number of
putative
DNA binding sites forvarious cellulartranscription factorsintheU3regionof the HFV LTR. Most notable is the established consensus se-quence for AP-1 binding, which occurs three times. The
AP-1 bindingsites arecritical for the action ofsome other
retroviraltransactivators, suchasthevisna-maedi virus Tat
protein (10, 14). In contrast, point mutations in the three
AP-1bindingsites of the HFV LTR resulted in
only
aslight
decrease (about
twofold)
in response to bell (Fig. 3),indi-catingthat themechanism ofbell-mediated transactivation
differs from that of visnavirus,inwhichanAP-1 site closeto
the TATA boxseems tobe one
major
targetfor thetrans-activator. In
addition,
aplasmid containing
fivecopies
ofsynthetic
AP-1binding
sites from thecollagenase
gene promoter(5'-AGC
TTG ATG AGTCAG CAG-3')
upstream of the HSV tk promoter(TRE-CAT
[2])
results inanincreaseof CAT
activity only by
twofold in the presence of bell(unpublished results), suggesting
that the roleof theputative
AP-1 site inbell-mediated
transactivation,
if any, is minor.ACKNOWLEDGMENTS
Wethank V.terMeulen for his kindgiftofplasmids pSbell-Sand
pSbell-A,
W. A. Haseltine forplasmid pRSV-CAT, and Yo Han Choifortechnical assistance. WeacknowledgeH.S.Shin and C.B. Chae forcritical reviews of themanuscript.Thisworkwassupported by Research Institute of Science and Technology grantR92061 and Center forBiofunctional Molecules grantCBM-92-06.
REFERENCES
1. Achong,B. G.,P. W.Mansell,M.A. Epstein,and P.Clifford. 1971.An unusual virus in cultures fromahuman nasopharyn-gealcarcinoma. J. Natl. Cancer Inst.46:299-307.
2. Angel, P., M.Imagawa, R. Chiu,B. Stein,R. J. Imbra,H. J. Rahmsdorf,C.Jonat,P.Herrlich,and M.Karin. 1987.Phorbol ester-induciblegenescontainacommoncis elementrecognized byaTPA-modulatedtrans-actingfactor.Cell49:729-739. 3.
Arya,
S. K., C. Guo, S. F. Josephs, and F. Wong-Staal. 1985.Trans-activatorgene of human T-lymphotropicvirus type III
(HTLV-III).Science 229:69-73.
4. Brady, J.,K.-T.Jeang, J.Duvall,andG.Khoury.1987. Identi-fication of
p4Ox-responsive
regulatory sequences within the human T-cell leukemia virus type I long terminal repeat. J. Virol.61:2175-2181.5. Cameron, K. R., S. M. Birchall, and M. A. Moses. 1978. Isolation offoamyvirusfrompatientwithdialysis
encephalop-athy.Lancetii:796.
6. Courey, A. J., and R. Tjian. 1988. Analysis ofSpl in vitro reveals multiple transcriptional domains, including a novel
glutamine-richactivation motif. Cell 55:887-898.
7. Cullen,B. R. 1990.TheHIV-1 tatprotein: anRNA
sequence-specific processivityfactor? Cell63:655-657.
8. Flugel, R. M., A. Rethwilm, B. Maurer, and G. Darai. 1987. Nucleotidesequenceanalysisoftheenvgene and itsflanking
regionof the humanspumaretrovirusrevealstwonovelgenes.
EMBOJ.6:2077-2084.
9. Fujisawa, J., M. Toita,T. Yoshimura, and M.Yoshida. 1991. The indirect association of human T-cell leukemia virus tax
proteinwithDNAresultsintranscriptionalactivation.J.Virol. 65:4525-4528.
10. Gdovin,S.L.,andJ.E.Clements. 1992. Molecularmechanism of visnavirustat:identificationofthetargets for
transcriptional
activation and evidence fora
post-transcriptional
effect. Virol-ogy188:438-450.11. Gilman, M. Z., R. N. Wilson, and R. A. Weinberg. 1986. Multipleprotein-bindingsites inthe5'-flankingregion
regulate
c-fos expression.Mol. Cell. Biol.6:4305-4316.
12. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express
chloramphenicol
acetyl-transferaseinmammalian cells. Mol.Cell. Biol.2:1044-1051.
13. Hauber, J.,and B. R.Cullen.1988. Mutational
analysis
of thetrans-activation-responsive region of the human
immunodefi-ciencyvirustypeIlongterminalrepeat.J.Virol. 62:673-679. 14. Hess,J.L.,J.A.Small,andJ.E.Clements.1989.
Sequences
inthevisna viruslongterminalrepeatthatcontrol
transcriptional
activity and respondto viraltrans-activation: involvement of AP-1 sites in basal activity and trans-activation. J. Virol. 63:3001-3015.15. Jakobovits,A.,D.H.Smith,E. B.Jakobovits,and D.J.
Capon.
1988. A discrete element 3' of human
immunodeficiency
virus 1(HIV-1) and HIV-2 mRNA initiation sitesmediates
transcrip-tional activation by an HIV trans activator. Mol. Cell. Biol.
on November 9, 2019 by guest
http://jvi.asm.org/
8:2555-2561.
16. Jones,N.C.,P. W. J. Rigby, andE.B.Ziff. 1988.Trans-acting protein factors and the regulation ofeukaryotic transcription: lessons from studies on DNA tumor viruses. Genes Dev. 2:267-281.
17. Keller, A.,K. M.Partin,M.Lochelt,H.Bannert,R. M.Flogel, and B. R. Cullen. 1991.Characterization of thetranscriptional trans activator of humanfoamy retrovirus. J. Virol. 65:2589-2594.
18. Kovesdi,I., R. Reichel, and J. R. Nevins. 1986.ElA transcrip-tioninduction involves enhanced binding ofafactor to upstream promoter sequences. Science 231:719-722.
19. Kovesdi, I.,R.Reichel,andJ.R. Nevins. 1986.Identification of acellular transcription factor involved inElA trans-activation. Cell 45:219-228.
20. Kunkel, T.A.1985.Rapid and efficientsite-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82-488-492.
21. Leang, K.,L.Brunet,and A. J.Berk.1988.Factorsresponsible for the highertranscriptional activity ofextractsof adenovirus-infected cells fractionate with the TATA box transcription factor.Mol. Cell. Biol. 8:1765-1774.
22. Lochelt, M., H.Zentgraf,and R. M.Flugel. 1991. Construction ofaninfectious DNA clone of thefull-length human spumaret-rovirus genome and mutagenesis of the bell gene. Virology 184:43-54.
23. Marriott, S. J.,L Boros, J. F. Duvall,andJ.N. Brady. 1989. Indirect binding of human T-cell leukemia virus type I tax1 toa responsivenesselement in the viral long terminal repeat. Mol. Cell. Biol. 9:4152-4160.
24. Maurer, B., H, Bannert, G. Darai, and R. M. Flfigel. 1988. Analysis of the primary structure of the long terminal repeat and the gag andpol genes of the humanspumaretrovirus. J. Virol. 62:1590-1597.
25. Maurer, B.,E.Serfling, V. terMeulen,andA.Rethwilm.1991. Transcription factorAP-1modulates theactivity of the human foamy virus long terminal repeat. J. Virol. 65:6353-6357. 26. McKnight, S.,and R.Tjian.1986.Transcriptional selectivity of
viralgenes inmammaliancells.Cell46:795-805.
27. Nyborg, J. K.,W.S.Dynan, I.S. Y.Chen,and W. Wachsman. 1988.Binding of host-cellfactors to DNA sequences in the long terminal repeat of human T-cell leukemia virus typeI: implica-tion for viralexpression. Proc. Natl. Acad. Sci. USA 85:1457-1461.
28. Pierce, J.W.,M.Lenardo,and D.Baltimore. 1988. Oligonucle-otide that binds nuclear factor NF-KB acts as a lymphoid-specificandinducible enhancer element.Proc.Natl.Acad. Sci. USA85:1482-1486.
29. Prince, A. M., B.A. A.Williams,T.Huima, L. Bardina, and B.
Brotman. 1984. Isolationofavirus fromchimpanzee liver cell cultures inoculated with sera containing the agent of non-A, non-Bhepatitis.Lancetii:1071-1075.
30. Queen, C., andD. Baltimore. 1983. Immunoglobin gene tran-scription is activated by downstream sequence element. Cell 33:741-748.
31. Rethwilm,A., 0. Erlwein, G. Baunach, B. Maurer,and V. ter Meulen. 1991. The transcriptional transactivator of human foamy virus mapstothe bellgenomic region. Proc. Natl. Acad. Sci. USA 88:941-945.
32. Rosen,C., J. G. Sodroski, andW. A.Haseltine. 1985. Location ofcis-actingregulatorysequences in the human T-cell lympho-tropicvirustype III(HTLV-III/LAV) long terminal repeat. Cell 41:813-823.
33. Selby,M.J.,E.S. Bain, P.A.Luciw, and B.M. Peterlin. 1989. Structure, sequence, and position of thestem-loop intar deter-mine transcriptional elongationby tat through the HIV-1 long terminal repeat. Genes Dev.3:547-558.
34. Shimotohno, K.,M.Takano,T.Teruuchi,and M. Miwa. 1986. Requirement ofmultiplecopies ofa21-nucleotide sequencein the U3regions of human T cell leukemia virus typeIandtypeII long terminal repeats for trans-acting activation of transcription. Proc. Natl. Acad. Sci. USA 83:8112-8116.
35. Sivaraman, L., and B. Thimmappaya. 1987. TIwo promoter-specific host factors interact with adjacent sequences in an ElA-inducible adenovirus promoter. Proc. Natl. Acad. Sci. USA 84:6112-6116.
36. Sodroski, J., C.Rosen, W.C. Goh, and W. Haseltine. 1985. A transcriptional activator proteinencodedby the x-lorregion of the humanT-cell leukemia virus. Science228:1430-1434. 37. Stancek, D., M. Stancekova, M. Janotka, P. Hnilica, and D.
Oravec. 1975.Isolation andsomeserological and epidemiolog-ical data on the viruses recovered from the patients with subacute thyroiditis de Quervain. Med. Microbiol. Immunol. 161:133-144.
38. Venkatesh,L.K.,P. A.Theodorakis,andG.Chinnadurai.1991. Distinctcis-acting regions in U3 regulate trans-activation of the human spumaretrovirus long terminal repeat by the viral bell geneproduct. Nucleic Acids Res. 19:3661-3666.
39. Weiss,R. A.1988. A virus insearch ofdisease.Nature(London) 333:497-498.
40. Werner, J., and H. Gelderblom. 1979. Isolationof foamy virus frompatients with de Quervain thyroiditis. Lancetii:258-259. 41. Wu, L., D.S. E. Rosser, M. C. Schmidt, and A.Berlk 1987. A
TATA box implicated in ElA transcriptional activation of a simple adenovirus2promoter. Nature(London) 326:512-515. 42. Young,D., J. Samuels,andJ.K.Clarke.1973.Afoamy virus of
possible human origin isolated in BHK-21 cells. Arch. Gesamte Virusforsch. 42:228-234.