Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Different Regions of Primase Subunit p48 Control Mouse
Polyomavirus and Simian Virus 40 DNA Replication In Vitro
ARMIN R. KAUTZ,1† ANNEROSE SCHNEIDER,1KLAUS WEISSHART,1‡ CHRISTIANE GEIGER,2
ANDHEINZ-PETER NASHEUER1*
Abteilung Biochemie, Institut fu¨r Molekulare Biotechnologie e.V., D-07745 Jena,1and Genzentrum,
Ludwig-Maximilians-Universita¨t Mu¨nchen, D-81377 Munich,2Germany
Received 20 July 2000/Accepted 17 November 2000
DNA polymerase␣-primase (pol-prim), a complex consisting of four subunits, is the major species-specific
factor for mouse polyomavirus (PyV) and simian virus 40 (SV40) DNA replication. Although p48 is the most conserved subunit of pol-prim, it is required for in vitro PyV DNA replication but can inhibit cell-free SV40 DNA replication. Production of chimeric human-mouse p48 revealed that different regions of p48 are involved in supporting PyV DNA replication and inhibiting SV40 DNA replication. The N and C-terminal parts of p48 do not have species-specific functions in cell-free PyV DNA replication, but the central part (amino acids [aa] 129 to 320) controls PyV DNA replication in vitro. However, PyV T antigen physically binds to mouse, human, and chimeric pol-prim complexes independently, whether they support PyV DNA replication or not. In contrast to the PyV system, the inhibitory effects of mouse p48 on SV40 DNA replication are mediated by N- and C-terminal regions of p48. Thus, a chimeric p48 containing human aa 1 to 128, mouse aa 129 to 320, and human aa 321 to 418 is active in both PyV and SV40 DNA replication in vitro.
Papovaviruses are small DNA tumor viruses (47). For their DNA replication these viruses contribute a viral origin of rep-lication (ori) and the large tumor antigens (Tag) of mouse polyomavirus (PyV) and simian virus 40 (SV40) or the E1 and E2 proteins of papillomaviruses; all other replication factors are supplied by the host (6, 42, 47, 49). Therefore, papovaviral DNA replication in vivo and in vitro has served as a model system to study virus-host interactions and the mechanisms of DNA replication in mammalian cells (37, 47, 49).
These studies allowed the development of a model for eu-karyotic DNA replication which relies on unwinding of double-stranded (ds) DNA and stepwise assembly of multiprotein complexes mediated by the viral initiator of DNA synthesis, large Tag. After Tag has recognized and formed a double hexamer at the replication origin, specific distortions of the dsDNA occur (10, 25). The following steps of DNA replication require the specific recruitment of host replication proteins, such as DNA polymerase ␣-primase (pol-prim), eukaryotic single-stranded (ss) DNA-binding protein, replication protein A (RPA), and topoisomerase I (topo I), to the origin of rep-lication (7, 11, 12, 14, 26, 27, 39, 40, 41). The interaction of Tag with pol-prim stimulatesoribinding by Tag (29). Melting and unwinding of ori dsDNA by Tag’s helicase activity requires stabilization of the ssDNA by RPA. The melting oforiDNA by Tag also needs the relaxation of supercoiled DNA by topo I. The first RNA primer is then synthesized by the primase ac-tivity of pol-prim. The preinitiation and initiation steps of viral
DNA synthesis at the origin of DNA replication require, in addition to the DNA-binding and enzymatic activities of the replication factors, specific physical contacts between these protein complexes (11, 48, 50, 51). The newly synthesized RNA is elongated by the DNA polymerase activity of pol-prim. After a replication factor C(RF-C)-catalyzed DNA polymerase switch to a leading strand synthesis complex consisting of pro-liferating cell nuclear antigen and DNA polymerase ␦, this enzyme complex carries out processive DNA synthesis (5, 19, 49). On the lagging strand initiation of the Okazaki fragment, DNA synthesis is also performed by pol-prim in cooperation with Tag and RPA (11, 26, 30, 50, 51). These RNA-DNA primers are then elongated by DNA polymerase␦orε(5, 19, 49). This model, which originated from work with cell-free systems of polyomaviruses and papillomaviruses, most likely also illustrates basic mechanisms of chromosomal DNA repli-cation (19).
Despite their similarities, PyV and SV40 show a significant difference in their DNA replication: PyV propagates only in mouse cells, whereas SV40 multiplies only in primate cells. Mouse pol-prim is the major species-specific factor for PyV DNA replication, whereas human pol-prim mediates host-spe-cific replication of SV40 DNA (4, 13, 28, 31, 44). Each of the four pol-prim subunits directly interacts with Tag (8, 12, 50), and these interactions play key roles in the permissiveness of the host cells: PyV Tag requires mouse p48 for its function (4, 13, 28), whereas SV40 Tag depends on the primate p180 sub-unit (21, 22, 31, 44).
Of the pol-prim subunits, the p48 polypeptide is the most highly conserved one between human and mouse (91% amino acid identity [43]), yet this polypeptide is the major species-specific factor for PyV DNA replication. Therefore, sequences of human and mouse p48 cDNA were exchanged, and chimeric human-mouse p48 cDNAs were expressed using baculovirus * Corresponding author. Mailing address: Institut fu¨r Molekulare
Biotechnologie e.V., Abt. Biochemie, Beutenbergstr. 11, D-07745 Jena, Germany. Phone: 49-3641-656290. Fax: 49-3641-656288. E-mail: [email protected].
† Present address: Institut fu¨r Biochemie, Universita¨t Erlangen-Nu¨rnberg, D-91054 Erlangen, Germany.
‡ Present address: Carl Zeiss Jena GmbH, D-07745 Jena, Germany.
1751
on November 9, 2019 by guest
http://jvi.asm.org/
vectors. These polypeptides were coexpressed with the p180, p68, and p58 subunits of pol-prim and purified as enzyme complexes. The chimeric pol-prim complexes were used to determine the parts of the mouse p48 subunit responsible for species-specific functions in PyV DNA replication.
MATERIALS AND METHODS
Production of the mutant p48 proteins.To obtain the chimeric human-mouse p48 subunits (see Fig. 1A), cDNAs were produced using the conserved restric-tion sites forBanII andNdeI to exchange the 5⬘and 3⬘ends, respectively. The constructs h(ml-320) and h(ml29-418) were transferred into the vectors pVL1392 and pVL1393, respectively. The cDNA containing the central part from mouse but human 5⬘and 3⬘ends was obtained by digesting pVL1392-h(m129-418) and pVL1393-h(m1-320) withPflMI and withBglII orBamHI. These DNAs were ligated to construct pVL1392-h(m129-320). After in vivo recombination baculo-viruses expressing recombinant proteins were created using Baculo-Gold DNA and standard procedures (Pharmingen, Hamburg, Germany) (35).
Proteins.PyV Tag, SV40 Tag, and the pol-prim complex (p180-p68-p58-p48) were purified from baculovirus-infected insect cells as described previously (4, 43, 44). RPA was bacterially expressed and purified as outlined before (18, 34, 45). Human topo I expressed in yeast and purified as described by Lisby et al. (24) was a generous gift of M. Lisby, University of Århus, Denmark. Monoclonal antibodies SJK237-71, SJK287-38 (46), F5 (36), and PAb101 (15, 16) specific for PyV and SV40 Tag were purified by affinity chromatography (17).
Protein manipulations.Protein concentration was determined according to the method of Bradford (3) using a commercial reagent with bovine immuno-globulin G as a standard (Bio-Rad, Munich, Germany). Sodium dodecyl sulfate (SDS) gel electrophoresis was carried out as described previously (20), with 10-kDa ladders (Life Technologies) as molecular mass markers. After polyacryl-amide gel electrophoresis, proteins were detected by silver staining according to the method of Nasheuer and Grosse (32).
DNA synthesis on⌽X174 ssDNA.The DNA replication of⌽X174 ssDNA was carried out in a reaction containing 66 ng of⌽X174 ssDNA (New England Biolabs) (33), 20 mM Tris-acetate (pH 7.3), 5 mM magnesium acetate, 20 mM potassium acetate, 1 mM dithiothreitol, 0.1 mg of bovine serum albumin (BSA) per ml, 1 mM ATP, 0.1 mM each of CTP, GTP, UTP, dATP, dCTP, dGTP, TTP, and 0.1 mM [␣-32P]dCTP (100 cpm/pmol) (Amersham Pharmacia Biotech,
Freiburg, Germany). Comparisons between different pol-prim preparations were made by adding 0.2 U of primase per assay. The incorporation of dCMP was determined by acid precipitation of DNA and scintillation counting.
Preparation of S100 extracts and replication of PyV in vitro.S100 extracts were prepared from logarithmically growing human 293S or mouse FM3A cells as previously described (43, 44). Cells were harvested by centrifugation and then washed twice with phosphate-buffered saline (PBS) and once with hypotonic buffer. The cells were resuspended in hypotonic buffer, incubated for 10 min on ice, and broken by 12 strokes in a Dounce homogenizer. The extracts were centrifuged at 4°C and 11,000⫻g. The supernatant was then adjusted to 100 mM NaCl and clarified by a second centrifugation at 100,000⫻g(S100 extract). Depletion of pol-prim from S100 extracts was performed essentially as previously described (43, 44).
The replication of PyV DNA in vitro was performed as previously described (4, 44). Briefly, the assay contained 1.2g of PyV Tag, 250 ng of pUC-Py1 DNA (PyV origin DNA [39]), and 200g of S100 or depleted S100 extract in 30 mM HEPES-NaOH (pH 7.8), 1 mM dithiothreitol, 7 mM magnesium acetate, 1 mM EGTA (pH 7.8), 4 mM ATP, 0.3 mM each of CTP, GTP, and UTP, 0.1 mM each of dATP and dGTP, 0.05 mM each of dCTP and dTTP, 40 mM creatine phos-phate, 80g of creatine kinase per ml, and 5Ci each of [␣32P]dCTP and
[␣32P]dTTP (3,000Ci/mmol) (Amersham Pharmacia Biotech). The cell-free
SV40 DNA replication contained 0.6g of SV40 Tag and 250 ng of pUC-HS (39). Pol-prim was added as indicated. The incorporation of radioactive de-oxynucleoside monophosphates (dNMPs) was measured by acid precipitation of DNA and scintillation counting. The total radioactivity was measured after spotting 5l of a 200-fold dilution of the replication assay onto GF52 filters (Schleicher & Schu¨ll, Dassel, Germany).
Initiation of replication on PyV and SV40 DNA.Initiation reactions were performed essentially as previously described (4, 39, 44). Briefly, the PyV initi-ation assay (40l) was assembled on ice and contained 0.25g of pUC-Pyl DNA (PyV origin DNA), 1.6g of PyV Tag, and 1g of RPA in 30 mM HEPES-KOH (pH 7.8), 7 mM magnesium acetate, 1 mM EGTA, 1 mM dithiothreitol, 0.2 mM UTP, 0.2 mM GTP, 0.01 mM CTP, 4 mM ATP, 40 mM creatine phosphate, 1g of creatine kinase, 0.3g of topo I, 0.25 mg of heat-treated BSA per ml, and 20
Ci of [␣-32P]CTP (3,000 Ci/mmol) (Amersham Pharmacia Biotech).
Recombi-nant pol-prim was added as indicated in the figure legends. SV40 initiation reactions (40l) were carried out as described above but contained 0.25g of pUC-HS DNA, 0.6g of SV40 Tag, and 0.5g of RPA. After incubation for 1 h at 37°C, one-eighth of the reaction mixture was used to estimate the amount of incorporated nucleotides by spotting it onto DE81 paper (38). The reaction products were precipitated with 0.8 M LiCl, 10 mM MgCl2, 10g of sonicated
salmon sperm DNA (Sigma), and 120l of ethanol for 1 h on dry ice, washed twice with 75% ethanol-water, dried, redissolved in 45% formamide–5 mM EDTA–0.05% xylene cyanol FF–0.05% bromphenol blue at 65°C for 30 min, heated for 3 min at 95°C, and electrophoresed in denaturing 20% polyacrylamide gels for 3 to 4 h at 600 V as described previously (4, 39). The reaction products were visualized by autoradiography.
ELISA.Modified enzyme-linked immunosorbent assays (ELISAs) were car-ried out as described previously (39). Briefly, pol-prim complexes or BSA (1g in 50l of PBS) was immobilized for 1 h at room temperature (RT). After washing three times with PBS, blocking solution (5% BSA [Fraction V; Sigma] in PBS) was incubated for 1 h at RT. The blocking solution was removed by washing three times with PBS. Then PyV Tag (1g in 50 l of PBS) was incubated for 1 h at RT in binding buffer (PBS supplemented with 8 mM MgCl2
and 4 mM adenylylimidodiphosphate [AMP-PNP]) and removed by washing three times with PBS. Bound Tag was recognized by monoclonal antibody F5 (5 g in 50l of PBS for 1 h at RT) and a secondary horseradish peroxidase-conjugated antibody (Dianova, Hamburg, Germany). The ELISA was developed with the horseradish peroxidase substrate kit according to the supplier’s instruc-tions (Bio-Rad).
RESULTS
Production of pol-prim containing chimeric human-murine
p48 primase.The murine p48 subunit controls the replication
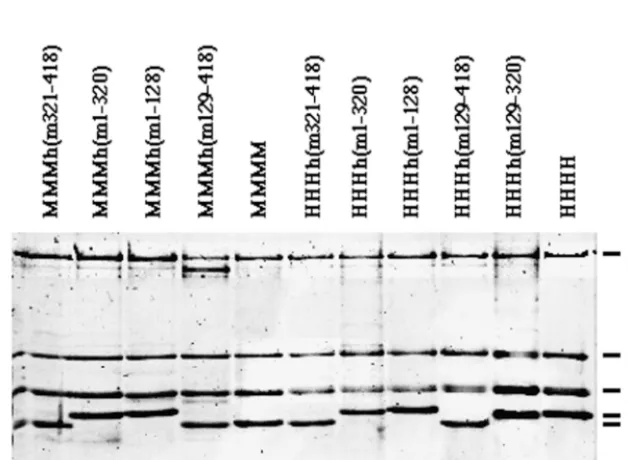
of PyV DNA in vitro. Human p180 determines the species specificity of SV40 DNA replication in vitro, whereas mouse p48 poisons the initiation of DNA replication of this primate virus in vitro (4, 44). These different activities raise the ques-tion of whether the same amino acids and activities of mouse p48 are involved in the regulation of DNA replication of both polyomaviruses. To address this question, regions of murine and human p48 were exchanged and chimeric proteins were produced using baculovirus vectors (Fig. 1). The chimeric p48 subunits of pol-prim are named according to their murine amino acids. Thus, h(ml-128) contains the murine amino acids (aa) 1 to 128, with the remaining amino acids being encoded by the human cDNA, whereas h(m321-418) consists of the C-terminal aa 321 to 418 from mouse and the human aa 1 to 320. These proteins were then coexpressed with human or mouse p180, p68, and p58 subunits (HHH and MMM, respectively, in complex designations). The recombinant proteins assembled into protein complexes which contained all four subunits and could be purified to near homogeneity by phosphocellulose and immunoaffinity chromatography (Fig. 2). All purified en-zyme complexes had DNA polymerase and primase activity. The specific DNA polymerase and primase activities of the protein complexes varied from 1,300 to 15,600 U/mg and from 170 to 2,280 U/mg, respectively (Table 1).
SDS gel electrophoresis of these immunoaffinity-purified chimeric complexes revealed that the pol-prim complex MMMh(m129-418) (MMMx refers to hybrid pol-prims con-taining chimeric p48 plus murine p180, p68, and p58) had a p180 subunit which was significantly more degraded than those of the other purified complexes (Fig. 2, lane 4). This proteo-lysis of p180 could be due to a high protease activity during purification. The SDS gel electrophoresis also showed that the p48 polypeptides containing the human aa 321 to 418 had a significantly higher apparent molecular mass than those
con-1752 KAUTZ ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
taining the murine C terminus (Fig. 2). The shifts of these polypeptides cannot be explained by an increase of molecular mass, since the predicted molecular masses of murine and human p48 have a difference of 0.6 kDa. Therefore, structural
features that cannot be resolved by SDS gel electrophoresis are most likely the cause of these shifts.
DNA synthesis by pol-prim with a chimeric human-murine
[image:3.612.116.486.73.322.2]p48 primase.To determine whether the subunits of chimeric
FIG. 1. Construction of chimeric human-murine p48 subunits. Chimeric p48s containing human and murine sequences were named according to their murine amino acids. The remaining amino acids are from human origin. The ability of these hybrid p48 polypeptides to support PyV and SV40 DNA replication is summarized on the right.
FIG. 2. Production of pol-prim complexes containing a chimeric p48 subunit. One microgram of each purified pol-prim complex (pol-prim with murine p180, p68, p58, and chimeric p48 [lanes 1 to 4], murine pol-prim [lane 5], pol-prim with human p180, p68, p58, and chimeric p48 [lanes 6 to 10], and human pol-prim [lane 11]) was analyzed by SDS gel electrophoresis and stained with silver.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.144.458.466.696.2]complexes cooperate, their ability to synthesize DNA on nat-ural ssDNA templates was studied. In the presence of the four deoxy- and ribonucleotides and Mg2⫹, all pol-prim complexes synthesized DNA (Fig. 3).The mouse pol-prim and the chi-meric protein complexes containing the three large murine subunits were in general more effective at DNA synthesis on ssDNA than the other enzyme complexes. However, the en-zyme complexes containing chimeric p48 synthesized DNA at least with an activity similar to that of the four-subunit human pol-prim (Fig. 3, compare columns 6 to 11).
The N and C termini of mouse p48 are not required for
species-specific replication of PyV DNA in vitro. The mouse
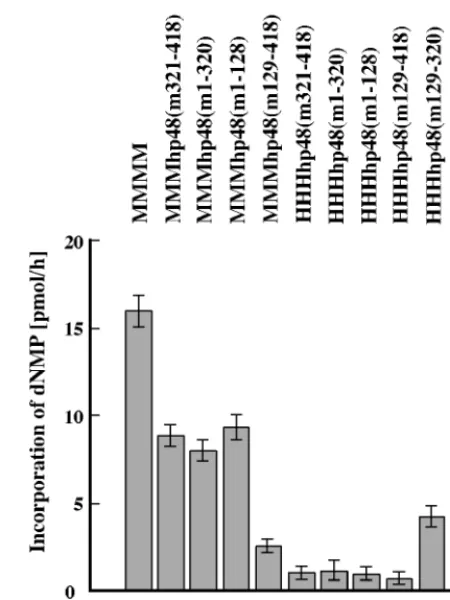
p48 subunit controls PyV DNA replication in vitro during the initiation of leading strand DNA synthesis through an un-known mechanism. The chimeric p48 subunits were tested in the replication of PyV DNA. To compensate for enzyme in-stabilities of recombinant proteins we used the same level of enzyme activity in each assay as indicated in the figures. As expected, mouse pol-prim supported in vitro PyV DNA repli-cation, whereas the human enzyme did not (Fig. 4). Low levels of active protein complexes which contained aa 1 to 320 or 129 to 418 of mouse p48 supported PyV DNA replication. The observed DNA synthesis products were resistant to DpnI, which degrades both the unmethylated input DNA from Esch-erichia coli and partially synthesized products (e.g., Fig. 4A, lanes 2 and 4), indicating that they were the result of semicon-servative DNA replication (Fig. 4A, lanes 7 to 10 and 15 to 22, and B, lanes 3 to 6, 13 to 16, and 19 to 22). However, hybrid pol-prim with p48 subunits containing only the N-terminal aa 1 to 128 or the C-terminal aa 321 to 418 of mouse p48 did not support cell-free PyV DNA replication. There were no
quali-tative differences between complexes with the three mouse subunits p180, p68, and p58 and those with the three human polypeptides (Fig. 4, compare panels A and B). In summary, all complexes with a p48 containing the central region of mouse p48 (aa 129 to 320) synthesized DNA that was resistant to DpnI digestion (Fig. 4A and B; summarized in Fig. 1), whereas complexes lacking this region of mouse p48 did not produce DpnI-resistant synthesis products (Fig. 4A and B).
Quantification of the cell-free PyV DNA replication re-vealed that pol-prim with three mouse proteins [MMMh(m1-FIG. 3. DNA synthesis on ssDNA by chimeric pol-prim complexes. The purified pol-prim complexes (0.3 U of primase) (murine pol-prim [column 1], pol-prim with murine p180, p68, p58, and chimeric p48 [columns 2 to 5], pol-prim with human p180, p68, p58, and chimeric p48 [columns 6 to 10], and human pol-prim [column 11]) were used to synthesize nucleic acids on ssDNA in the presence of nucleoside triphosphates, deoxynucleoside triphosphates, and Mg2⫹. The assay
was repeated twice, and the means and standard deviations are pre-sented.
FIG. 4. Replication of PyV DNA by chimeric pol-prim complexes. (A) Increasing amounts (0.25 and 0.5 U of DNA polymerase) of recombinant pol-prim containing murine p180, p68, p58, and chimeric p48 (lanes 3 to 18), four mouse subunits (lanes 19 to 22), or four human subunits (lanes 23 to 26) were added to depleted human 293S cell extracts (lanes 1 and 2) supplemented with PyV Tag and a PyV origin-containing plasmid. (B) The replication products of these extracts were also determined with recombinant enzymes containing three human subunits and hybrid p48. The assays were carried out in the absence of recombinant pol-prim (lanes 1 and 2) or in the presence of recombinant mouse (lanes 3 to 6), human (lanes 23 and 24), and hybrid pol-prim containing the three human subunits p180, p68, and p58, together with a chimeric subunit p48 (lanes 7 to 22). DNA synthesis products for panels A and B were analyzed for complete DNA replication by digestion with 10 U each ofEcoRI andDpnI (even-numbered lanes). In parallel, the products were linearized with 10 U ofEcoRI (odd-numbered lanes). The DNA synthesis products were visualized by autoradiography. The arrow at the right side of the figure indicates the linearized DNA synthesis products. (C) The incorporation of dNMPs into PyV DNA was determined by acid precipitation of DNA and scintillation counting (light and dark columns, showing means and standard deviations of a minimum of three replication assays with 0.25 and 0.5 primase units, respectively, of the indicated pol-prim). Columns 1, negative control of depleted extracts; columns 2, mouse pol-prim; columns 3 to 10, hybrid pol-prim as indicated; columns 11, human pol-prim.
TABLE 1. Enzyme activities of recombinant pol-prim complexes
Enzyme
complex Protein concn(mg/ml)
Sp act of DNA polymerase
(U/mg)
Sp act of primase (U/mg)
MMMM 0.23 3,560 260
MMMh(m321-418) 2.8 1,270 305
MMMh(ml-320) 1.3 2,340 492
MMMh(ml-128) 0.4 6,900 1,900
MMMh(ml29-418) 0.89 3,370 820
HHHh(m321-418) 0.23 10,800 1,080
HHHh(m1-128) 0.52 5,000 750
HHHh(m1-320) 0.14 15,600 2,280
HHHh(m129-418) 0.34 7,350 764
HHHh(m129-320) 0.45 3,130 170
HHHH 0.52 4,923 960
1754 KAUTZ ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.316.541.68.368.2] [image:4.612.53.294.84.220.2]on November 9, 2019 by guest
http://jvi.asm.org/
320) and MMMh(m129-418)] supported DNA replication more efficiently than those with three human subunits (Fig. 4C, compare columns 4, 6, 8, and 10). These data suggest that the mouse p48 mediates species-specific replication of PyV DNA, whereas the other subunits have stimulatory effects on the replication reactions.
Amino acids within the central region of mouse p48 control
species specificity of cell-free PyV DNA replication.Since all
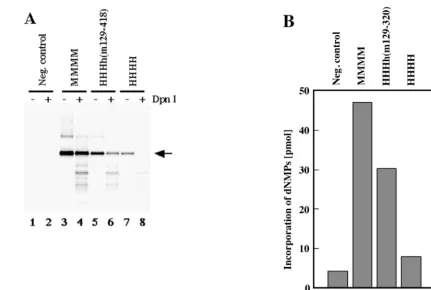
proteins containing the central part of mouse p48 were active in PyV DNA replication these findings suggest that aa 129 to 320 control PyV DNA replication. Therefore, a chimeric p48 containing this region from mouse, together with human N-and C-terminal amino acids, was produced. The enzyme com-plex HHHh(m129-320) was highly active in PyV DNA repli-cation in vitro, andDpnI-resistant DNA was synthesized (Fig. 5A, lane 6). Quantification of the reaction showed that the incorporation of radioactively labeled dNMPs by HH-Hh(m129-320) was about 65% of that by mouse pol-prim (Fig. 5B, compare columns 2 and 3).
Initiation of PyV DNA replication with hybrid pol-prim in
the presence of purified proteins.The initiation of PyV DNA
replication is the species-specific step. Therefore, we tested whether we could reproduce the above results by using the initiation reaction of PyV DNA replication exclusively with
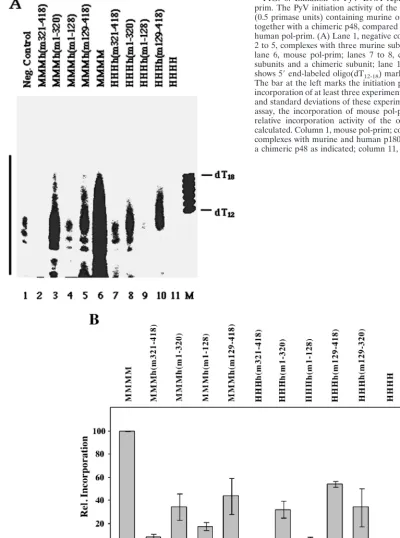
purified proteins. Various pol-prim complexes (0.5 primase units) were used to initiate PyV DNA replication. These low amounts of mouse enzyme complex were sufficient to support the reaction, whereas the same amounts of human pol-prim were not (Fig. 6).The enzyme complexes containing the central part (aa 129 to 320) of mouse p48 were active, whereas those missing this region from mouse were inactive (Fig. 1 and 6).The other pol-prim subunits did not interfere with the ini-tiation activity of the chimeric enzyme complexes, and the active chimeric complexes reached 35 to 50% of the activity of mouse pol-prim (Fig. 6B, compare columns 1, 3, 5, 7, 9, and 10). Thus, the initiation assay, which exclusively depends on purified proteins, underlines the fact that the central part of mouse p48 mediates the species specificity of PyV DNA rep-lication.
PyV, Tag physically interacts with mouse, human, and
chi-meric pol-prim complexes.These results raised the question of
[image:6.612.86.517.69.359.2]whether the interaction of PyV Tag with pol-prim is species specific. Equal amounts of each enzyme complex were immo-bilized and then incubated with PyV Tag. PyV Tag physically bound to mouse and human pol-prim, as well as to the chi-meric complexes (Fig. 7). The results suggest that the physical interaction of PyV Tag with the pol-prim complex is not the species-specific step, since Tag binds with a similar efficiency to FIG. 5. aa 129 to 320 of mouse p48 control species specificity of cell-free PyV DNA replication. The ability of the chimeric pol-prim complex (0.5 DNA polymerase units) containing human p180, p68, p58, and chimeric h(m129-320) to support PyV DNA replication in vitro was determined by using depleted extracts from human 293 cells (A, lanes 5 and 6). This activity was compared with the replication activity of murine (A, lanes 3 and 4) and human (A, lanes 7 and 8) (0.5 DNA polymerase units) pol-prim. DNA synthesis products for panel A were analyzed for complete DNA replication by digestion with 10 U each ofEcoRI andDpnI (even-numbered lanes). In parallel, the products were linearized with 10 U of
EcoRI (odd-numbered lanes). For lanes 1 and 2, the synthesis products of depleted 293S extracts are presented. To compare the replication activities in a quantitative way the incorporated dNMPs were precipitated with trichloroacetic acid and measured by scintillation counting (B). Columns 1 to 4, depleted 293S extracts and mouse, chimeric HHHh(m129-320), and human pol-prim, respectively.
1756 KAUTZ ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
both pol-prim complexes that are active and those which are inactive in cell-free PyV DNA replication (Fig. 4 to 7). This finding is consistent with data published earlier which show that SV40 Tag binds to human and mouse pol-prim (39).
Amino acids within the N and C termini of p48 inhibit SV40
DNA replication in vitro.The mouse p48 subunit is required
for PyV DNA replication, but in a complex with the three human subunits mouse p48 poisons SV40 DNA replication.
This phenotype can be rescued by mouse p58 (4, 44). This behavior raised the question of whether the same region of mouse p48 is involved in the negative regulation of SV40 DNA replication and in the positive control of PyV DNA replication (44). The four chimeric complexes containing the N or C terminus of mouse p48 did not support initiation of SV40 DNA replication (Fig. 8, lanes 3 to 6; summarized in Fig. 1), although these enzyme complexes initiated DNA synthesis and elon-FIG. 6. Initiation of PyV DNA replication by recombinant pol-prim. The PyV initiation activity of the chimeric pol-prim complexes (0.5 primase units) containing murine or human p180, p68, and p58, together with a chimeric p48, compared with recombinant mouse and human pol-prim. (A) Lane 1, negative control without pol-prim; lanes 2 to 5, complexes with three murine subunits and a chimeric subunit; lane 6, mouse pol-prim; lanes 7 to 8, complexes with three human subunits and a chimeric subunit; lane 11, human pol-prim. Lane M shows 5⬘end-labeled oligo(dT12-18) markers as indicated at the right. The bar at the left marks the initiation products. (B) The radioactive incorporation of at least three experiments was determined, and means and standard deviations of these experiments are presented. For each assay, the incorporation of mouse pol-prim was set to 100, and the relative incorporation activity of the other enzyme complexes was calculated. Column 1, mouse pol-prim; columns 2 to 10, hybrid enzyme complexes with murine and human p180, p68, and p58, together with a chimeric p48 as indicated; column 11, human pol-prim.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.70.471.80.618.2]gated these primers on natural ssDNA templates (Fig. 3). Interestingly, the pol-prim complex HHHh(m129-320) con-taining the central part from mouse p48 synthesized primers in the cell-free SV40 system (Fig. 8, lane 2). Its efficiency was about 53% of that of human pol-prim (Fig. 8, compare lanes 2 and 7). In addition, HHHh(m129-320) supported SV40 DNA replication, and the incorporation of dNMPs with HH-Hh(m129-320) reached about 62% of that with human pol-prim (data not shown). These results indicate that amino acids within the N or C terminus of mouse p48 can suppress initia-tion of SV40 DNA replicainitia-tion in vitro.
DISCUSSION
The cell-free polyomavirus DNA replication systems of PyV and SV40 have provided detailed insights into the mechanisms of eukaryotic DNA replication (for review, see reference 49 and references therein). The comparison of these DNA repli-cation assays allowed the discrimination of separate essential functions of pol-prim in the control of viral DNA replication and explained the host specificity of these two viruses at least in part (reviewed in reference 42).
Control of PyV DNA replication by mouse p48. The p48
subunit of pol-prim has essential functions in the initiation of leading and lagging strand DNA synthesis and in the control of PyV and SV40 DNA replication (4, 5, 13, 49, 50, 51). The initiation and DNA replication of PyV in vitro is supported by mouse p48 together with three human subunits of heterotet-rameric pol-prim (4). We show here that the three large mouse subunits of the enzyme complex influence functions of mouse p48 and chimeric p48 sequences during PyV DNA replication
and stimulate the replication efficiency of cell-free DNA rep-lication (Fig. 4). MMMh(m1-320) and MMMh(m129-418) sup-ported DNA replication more efficiently in the cell-free PyV system than the complexes HHHh(m1-320) and HHHh(m129-418) (Fig. 4). This increased activity seems to be dependent on a higher DNA synthesis rate of MMMh(m1-320) and MMMh(m129-418), since their initiation activity in the PyV system does not significantly differ from that of the other two chimeric enzyme complexes (Fig. 6).This interpretation is sup-ported by the finding that MMMh(m1-320) and MMMh(m129-418) synthesized DNA more efficiently on a natural⌽⫻174 ssDNA template than did the other two en-zyme complexes (Fig. 3).
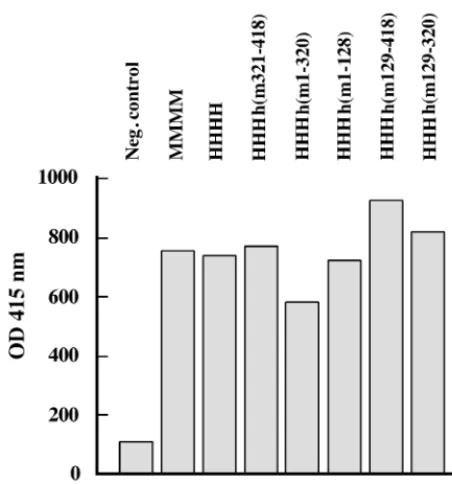
The replication of dsDNA containing a PyVorirevealed that mouse p48 controls species specificity of PyV DNA replication even in the presence of three other murine subunits. Especially the central residues of mouse p48 carry essential functions to FIG. 7. Interaction of PyV Tag with recombinant pol-prim. BSA (1
[image:8.612.61.287.69.311.2]g) (column 1) or purified, recombinant pol-prim complexes (1g) (columns 2 to 8, as indicated) were immobilized. After incubation with 1g of PyV Tag for 1 h at RT, Tag–pol-prim complexes were detected with Tag-specific antibody F5. The means of three experiments are presented. OD, optical density.
FIG. 8. Initiation of SV40 DNA replication by recombinant pol-prim. Pol-prims (0.8 U of primase activity) were tested in the SV40 initiation reaction. Lane 1, assay without prim; lanes 2 to 6, pol-prim containing three human subunits and chimeric p48; lane 7, hu-man pol-prim. Labeled oligo (dT12-18) marker is indicated at the right. The bar at the left marks the initiation products.
1758 KAUTZ ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
support PyV DNA replication in vitro. The shift in apparent molecular mass determined by SDS gel electrophoresis is not a parameter that affects the host specificity of PyV DNA repli-cation, since this shift is most likely caused by amino acids in the region aa 321 to 421 of human p48 which do not interfere with PyV DNA replication in human extracts (Fig. 2 and 4 to 6). However, the species specificity of PyV DNA replication only becomes manifest within the initiation complex composed of PyV Tag, pol-prim, and RPA at an origin of DNA replica-tion. Although PyV Tag physically binds to pol-prim, the direct physical contacts do not show species-specific effects (Fig. 7) (4). Therefore, the primase subunit p48 of the nonpermissive host might differ in its preferential initiation sequence at the viralori. Alternatively, conformational restrictions within the initiation complex consisting of PyV Tag, RPA, pol-prim, topo I, and the PyVorimay hinder recognition of the DNA tem-plate by the human primase. These restrictions may occur at any stage of the initiation process, as our assay does not dis-tinguish between them (2, 5, 6, 19, 23, 49). Especially, Tag serves as a mediator protein (1) to support RNA synthesis by pol-prim on an RPA-bound template by primase. PyV Tag is most likely able to carry out these activities at the PyVoriwith mouse p48 but not with the human p48.
Activity of mouse p48 in SV40 DNA replication.In
combi-nation with human p180, p68, and p58, the mouse p48 has been found to poison initiation of SV40 DNA replication (4, 44). The results presented here indicate that the inhibitory effect of mouse p48 on SV40 DNA replication and its supporting func-tion(s) in PyV DNA replication depend on different mecha-nisms and can be separated. In the presence of either the murine N or C terminus the initiation of SV40 DNA replica-tion is inhibited, whereas the central part of mouse p48 is required for the initiation of PyV DNA replication (Fig. 1, 6, and 8). However, the chimeric protein h(m129-320) supports the cell-free DNA replication of both PyV and SV40 (Fig. 1, 4, 5, 6, and 8). The mechanism for the inhibitory effect of mouse p48, and more precisely its N- and C-terminal amino acids, on SV40 DNA replication is still unclear, but this inhibition is relieved by substituting the human primase subunit p58 with the mouse polypeptide (4). These findings suggest that the N and C termini of p48 cooperate with p58 during the initiation of SV40 DNA replication. The human p58 might fail to func-tionally interact with these regions of mouse p48, whereas mouse p58 does interact (4). This interpretation is supported by earlier findings that the N and C termini of p48 are involved in its physical binding to p58 (9). However, this cooperation of the two mammalian subunits is not disturbed in the initiation of DNA synthesis on ssDNA, the initiation of leading strand DNA synthesis in the PyV origin, and the initiation of Okazaki fragments on the lagging strand (Fig. 3 to 6)(39, 44).
ACKNOWLEDGMENTS
We thank F. Grosse and R. Smith for critical reading of the manu-script and A. Willitzer for synthesizing oligonucleotides.
This work was financially supported by the Deutsche Forschungsge-meinschaft (Na 190/8, Na 190/10, and Na 190/12) and the EC (CT970125). The IMB is a Gottfried-Wilhelm-Leibniz Institut and is financially supported by the federal government and the Land Thu¨rin-gen.
REFERENCES
1.Beernink, H. T., and S. W. Morrical.1999. RMPs: recombination/replication mediator proteins. Trends Biochem. Sci.24:385–389.
2.Bhattacharyya, S., H. Lorimer, and C. Prives.1995. Murine polyomavirus and simian virus 40 large T antigens produce different structural alterations in viral origin DNA. J. Virol.69:7579–7585.
3.Bradford, M. M.1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem.72:248–254.
4.Bru¨ckner, A., F. Stadlbauer, L. A. Guarino, A. Brunahl, C. Schneider, C. Rehfuess, C. Prives, E. Fanning, and H.-P. Nasheuer.1995. The mouse DNA polymerase␣-primase subunit p48 mediates species-specific replica-tion of polyomavirus DNA in vitro. Mol. Cell. Biol.15:1716–1724. 5.Burgers, P. M.1998. Eukaryotic DNA polymerases in DNA replication and
DNA repair. Chromosoma107:218–227.
6.Challberg, M., and T. J. Kelly.1989. Animal virus DNA replication. Annu. Rev. Biochem.58:671–717.
7.Collins, K. L., and T. J. Kelly.1991. The effects of T antigen and replication protein A on the initiation of DNA synthesis by DNA polymerase␣-primase. Mol. Cell. Biol.11:2108– 2115.
8.Collins, K. L., A. A. R. Russo, B. Y. Tseng, and T. J. Kelly.1993. The role of the 70kDa subunit of human DNA polymerase ␣ in DNA replication. EMBO J.12:4555–4566.
9.Copeland, W. C., and X. Tan.1995. Active site mapping of the catalytic mouse primase subunit by alanine scanning mutagenesis. J. Biol. Chem.
270:3905–3913.
10. Dean, F. B., J. A. Borowiec, T. Eki, and J. Hurwitz.1992. The simian virus 40 T antigen double hexamer assembles around the DNA at the replication origin. J. Biol. Chem.267:14129–14137.
11. Dornreiter, I., L. F. Erdile, I. U. Gilbert, D. von Winkler, T. J. Kelly, and E. Fanning.1992. Interaction of DNA polymerase␣-primase with cellular rep-lication protein A and SV40 T antigen. EMBO J.11:769–776.
12. Dornreiter, I., A. Ho¨ss, A. K. Arthur, and E. Fanning.1990. SV40 T antigen binds directly to the catalytic subunit of DNA polymerase alpha. EMBO J.
9:3329–3336.
13. Eki, T., T. Enomoto, C. Masutani, A. Miyajima, R. Takada, Y. Murakami, T. Ohno, F. Hanaoka, and M. Ui.1991. Mouse DNA primase plays the prin-cipal role in determination of permissiveness for polyomavirus DNA repli-cation. J. Virol.65:4874–4881.
14. Gannon, J. V., and D. P. Lane.1990. Interactions between SV40 T antigen and DNA polymerase␣. New Biol.2:84–92.
15. Gurney, E. G., R. O. Harrison, and J. Fenno.1980. Monoclonal antibodies against simian virus 40 T antigens: evidence for distinct subclasses of large T antigen and for similarities among nonviral T antigens. J. Virol.34:752–763. 16. Gurney, E. G., S. Tamowski, and W. Deppert.1986. Antigenic binding sites of monoclonal antibodies specific for simian virus 40 large T antigen. J. Vi-rol.57:1168–1172.
17. Harlow, E., and D. P. Lane.1988. Antibodies: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
18. Henricksen, L. A., C. B. Umbricht, and M. S. Wold.1994. Recombinant replication protein A: expression, complex formation, and functional char-acterization. J. Biol. Chem.269:11121–11132.
19. Hu¨bscher, U., H.-P. Nasheuer, and J. Syva¨joja.2000. Eukaryotic DNA polymerases, a growing family. Trends Biochem. Sci.25:143–147. 20. Laemmli, U. K.1970. Cleavage of structural proteins during the assembly of
the head of bacteriophage T4. Nature227:680–685.
21. Li, J. J., and T. J. Kelly.1984. Simian virus 40 DNA replication in vitro. Proc. Natl. Acad. Sci. USA81:6973–6977.
22. Li, J. J., and T. J. Kelly.1985. Simian virus 40 DNA replication in vitro: specificity of initiation and evidence for bidirectional replication. Mol. Cell. Biol.5:1238–1246.
23. Li, L., B. L. Li, M. Hock, E. Wang, and W. R. Folk.1995. Sequences flanking the pentanucleotide T-antigen binding sites in the polyomavirus core origin help determine selectivity of DNA replication. J. Virol.69:7570–7578. 24. Lisby, M., B. O. Krogh, F. Boege, O. Westergaard, and B. R. Knudsen.1998.
Camptothecins inhibit the utilization of hydrogen peroxide in the ligation step of topoisomerase I catalysis. Biochemistry37:10815–10827.
25. Mastrangelo, I. A., P. V. C. Hough, J. S. Wall, M. Dodson, F. B. Dean, and J. Hurwitz.1989. ATP-dependent assembly of double hexamers of SV40 T antigen at the viral origin of DNA replication. Nature338:658–662. 26. Melendy, T., and B. Stillman.1993. An interaction between replication
protein A and SV40 T antigen appears essential for primosome assembly during SV40 DNA replication. J. Biol. Chem.268:3389–3395.
27. Moses, K., and C. Prives.1994. A unique subpopulation of murine DNA polymerase␣-primase specifically interacts with polyoma virus T antigen and stimulates DNA replication. Mol. Cell. Biol.14:2767–2776.
28. Murakami, Y., T. Eki, M. Yamada, C. Prives, and J. Hurwitz.1986. Species-specificin vitrosynthesis of DNA containing the polyoma virus origin of replication. Proc. Natl. Acad. Sci. USA83:6347–6351.
29. Murakami, Y., and J. Hurwitz.1993. Functional interactions between SV40
on November 9, 2019 by guest
http://jvi.asm.org/
T antigen and other replication proteins at the replication fork. J. Biol. Chem.268:11008–11017.
30. Murakami, Y., and J. Hurwitz.1993. DNA polymerase ␣stimulates the ATP-dependent binding of simian virus tumor T antigen to the SV40 origin of replication. J. Biol. Chem.268:11018–11027.
31. Murakami, Y., C. R. Wobbe, L. Weissbach, F. B. Dean, and J. Hurwitz.1986. Role of DNA polymerase␣and DNA primase in simian virus 40 DNA replicationin vitro. Proc. Natl. Acad. Sci. USA83:2869–2873.
32. Nasheuer, H.-P., and F. Grosse.1988. DNA polymerase␣-primase from calf thymus. Determination of the polypeptide responsible for primase activity. J. Biol. Chem.263:8981– 8988.
33. Nasheuer, H.-P., and F. Grosse.1987. Immunoaffinity-purified DNA poly-merase␣displays novel properties. Biochemistry26:8458–8466.
34. Nasheuer, H.-P., D. von Winkler, C. Schneider, I. Dornreiter, I. Gilbert, and E. Fanning.1992. Purification and functional characterization of bovine RP-A in an in vitro SV40 DNA replication system. Chromosoma102:S52– S59.
35. O’Reilly, D. R., L. K. Miller, and V. A. Luckow.1992. Baculovirus expression vectors—a laboratory manual. W. H. Freeman and Co., New York, N.Y. 36. Pallas, D. C., C. Schley, M. Mahoney, E. Harlow, B. S. Schaffhausen, and
T. M. Roberts1986. Polyomavirus small T antigen: overproduction in bac-teria, purification, and utilization for monoclonal and polyclonal antibody production. J. Virol.60:1075–1084.
37. Reynisdottir, I., S. Bhattacharyya, D. Zhang, and C. Prives. 1999. The retinoblastoma protein alters the phosphorylation state of polyomavirus large T antigen in murine cell extracts and inhibits polyomavirus origin DNA replication. J. Virol.73:3004–3013.
38. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
39. Schneider, C., K. Weisshart, L. A. Guarino, I. Dornreiter, and E. Fanning.
1994. Species-specific functional interactions of DNA polymerase␣-primase with SV40 T antigen require SV40 origin DNA. Mol. Cell. Biol.14:3176– 3185.
40. Simmons, D. T., T. Melendy, D. Usher, and B. Stillman.1996. Simian virus
40 large T antigen binds to topoisomerase I. Virology222:365–374. 41. Smale, S. T., and R. Tjian.1986. T antigen-DNA polymerase␣complex
implicated in simian virus 40 DNA replication. Mol. Cell. Biol.6:4077–4087. 42. Smith, R. W. P., and H.-P. Nasheuer.2000. Control of papovaviral DNA replication, p. 67–92.InPandalai (ed.), Recent research developments in virology, vol. 2. Transworld Research Network, Trivandrum, India. 43. Stadlbauer, F., A. Brueckner, C. Rehfuess, C. Eckerskorn, F. Lottspeich, V.
Fo¨rster, B. Y. Tseng, and H.-P. Nasheuer.1994. DNA replication in vitro by recombinant DNA-polymerase-␣-primase. Eur. J. Biochem.222:781–793. 44. Stadlbauer, F., C. Voitenleitner, A. Bru¨ckner, E. Fanning, and H.-P.
Nasheuer.1996. Species-specific replication of simian virus 40 DNA in vitro requires the p180 subunit of human DNA polymerase␣-primase. Mol. Cell. Biol.16:94–104.
45. Stigger, E., F. B. Dean, J. Hurwitz, and S. H. Lee.1994. Reconstitution of functional human single-stranded DNA-binding protein from individual sub-units expressed by recombinant baculoviruses. Proc. Natl. Acad. Sci. USA
91:579–583.
46. Tanaka, S., S.-Z. Hu, T. S.-F. Wang, and D. Korn.1982. Preparation and preliminary characterization of monoclonal antibodies against human DNA polymerase␣. J. Biol. Chem.257:8386–8390.
47. Tooze, J.1981. Molecular biology of tumor virus, part 2. DNA tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
48. Trowbridge, P. W., R. Roy, and D. T. Simmons.1999. Human topoisomerase I promotes initiation of simian virus 40 DNA replication in vitro. Mol. Cell. Biol.19:1686–1694.
49. Waga, S., and B. Stillman.1998. The DNA replication fork in eukaryotic cells. Annu. Rev. Biochem.67:721–751.
50. Weisshart, K., H. Fo¨rster, E. Kremmer, B. Schlott, F. Grosse, and H.-P. Nasheuer.2000. Protein-protein interactions of the primase subunits p58 and p48 with simian virus 40 T antigen are required for efficient primer synthesis in a cell-free system. J. Biol. Chem.275:17328–17337.
51. Weisshart, K., P. Taneja, and E. Fanning1998. The replication protein A binding site in simian virus 40 (SV40) T antigen and its role in the initial steps of SV40 DNA replication. J. Virol.72:9771–9781.
1760 KAUTZ ET AL. J. VIROL.