Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Second-Site Reversion of a Human Immunodeficiency Virus
Type 1 Reverse Transcriptase Mutant That Restores
Enzyme Function and Replication Capacity
ISABEL OLIVARES,
1VI´CTOR SA
´ NCHEZ-MERINO,
1MIGUEL A. MARTI´NEZ,
2ESTEBAN DOMINGO,
3CECILIO LO
´ PEZ-GALI´NDEZ,
1ANDLUIS MENE´NDEZ-ARIAS
3*
Centro Nacional de Biologı´a Fundamental, Instituto de Salud Carlos III, 28220 Majadahonda (Madrid),
1Fundacio´n
Irsi-Caixa, Hospital Universitario Germans Trias i Pujol, Badalona (Barcelona),
2and Centro de Biologı´a Molecular
“Severo Ochoa,” Consejo Superior de Investigaciones Cientı´ficas-Universidad Auto´noma de Madrid,
Cantoblanco, 28049 Madrid,
3Spain
Received 17 March 1999/Accepted 10 May 1999
Nonconservative substitutions for Tyr-115 in the reverse transcriptase (RT) of human immunodeficiency
virus type 1 (HIV-1) lead to enzymes displaying lower affinity for deoxynucleoside triphosphates (dNTPs)
(A. M. Martı´n-Herna´ndez, E. Domingo, and L. Mene´ndez-Arias, EMBO J. 15:4434–4442, 1996). Several
mutations at this position (Y115W, Y115L, Y115A, and Y115D) were introduced in an infectious HIV-1 clone,
and the replicative capacity of the mutant viruses was monitored. Y115W was the only mutant able to replicate
in MT-4 cells, albeit very poorly. Nucleotide sequence analysis of the progeny virus recovered from
superna-tants of four independent transfection experiments showed that the Y115W mutation was maintained.
How-ever, in all cases an additional substitution in the primer grip of the RT (M230I) emerged when the virus
increased its replication capacity. Using recombinant HIV-1 RT, we demonstrate that M230I mitigates the
polymerase activity defect of the Y115W mutant, by increasing the dNTP binding affinity of the enzyme. The
second-site suppressor effects observed were mediated by mutations in the 66-kDa subunit of the RT, as
demonstrated with chimeric heterodimers. Examination of available crystal structures of HIV-1 RT suggests
a possible mechanism for restoration of enzyme activity by the second-site revertant.
Human immunodeficiency virus type 1 (HIV-1) reverse
tran-scriptase (RT) plays an essential role in the replication of the
single-stranded genomic RNA of the virus. Reverse
transcrip-tion involves the synthesis of a double-stranded proviral DNA
which integrates in the host genome, in a process mediated by
the RNA-dependent and DNA-dependent polymerase and the
endonuclease (RNase H) activities of the enzyme (for recent
reviews, see references 1, 10, and 31). HIV-1 RT is also a target
for chemotherapeutic intervention in the control of AIDS. The
crystallographic structure of HIV-1 RT has been determined in
the absence of ligands (28), complexed with nonnucleoside RT
inhibitors (15), and complexed with double-stranded DNA
(13). In addition, a crystal structure of a covalently trapped
catalytic complex of HIV-1 RT containing a DNA
template-primer and a deoxynucleoside triphosphate (dNTP) has been
recently reported (11). HIV-1 RT is a heterodimer composed
of two subunits of 66 and 51 kDa, with subdomains termed
fingers, thumb, palm, and connection in both subunits and an
RNase H domain in the larger subunit only. The polymerase
active site resides within the palm subdomain of the 66-kDa
subunit, which contains the catalytic aspartic acid residues 110,
185, and 186. Other residues in their vicinity, such as Lys-65,
Arg-72, Asp-113, Ala-114, Tyr-115, and Gln-151, are involved
in the interaction with the incoming dNTP (11). The role of
these amino acids in polymerase function has been investigated
by site-directed mutagenesis (3, 33, 36). Nonconservative
sub-stitutions at several of these positions often lead to the loss of
RT function and virus viability (16).
In previous studies, we used site-directed mutagenesis to
produce HIV-1 RT variants with substitutions at Tyr-115. This
amino acid was systematically replaced by Phe, Trp, Val, Ile,
Met, Leu, Ala, Ser, Cys, Asn, His, Gly, Asp, Lys, and Pro.
While the substitution of Phe for Tyr-115 rendered RT fully
active, other replacements affected the polymerase activity of
the enzyme, by increasing the
K
mvalues for the incorporation
of dNTPs (17, 18), suggesting an altered dNTP binding
func-tion. This effect was more pronounced in those variant RTs
with smaller and less hydrophobic side chains at position 115.
To study the viability of virus harboring mutations at position
115, we introduced the substitutions Y115W, Y115L, Y115A,
and Y115D in the RT of an infectious HIV-1 clone by
site-directed mutagenesis. In transfection experiments with one of
the variant RTs (Y115W mutant), viable virus was recovered,
but in all cases, the genotypic analysis of the progeny revealed
that it contained an additional substitution at position 230 (Ile
instead of Met). This second-site reversion appears to
com-pensate for the dNTP binding defect shown by the Y115W
mutant.
MATERIALS AND METHODS
Cell lines and molecular clones.COS-1 cells and MT-4 cells were maintained
as monolayer and suspension cultures, respectively, in RPMI 1640 medium supplemented with 10% fetal bovine serum and 2 mM glutamine. Virus used in this study was an infectious molecular clone of HIV-1 isolate 89ES061 previously described (21, 22). The 5⬘ end of the proviral DNA, including the 5⬘ long terminal repeat region and the viral genesgag,pol, andvif, was cloned in theXbaI andEcoRI sites of plasmid pBSK (Stratagene) to obtain plasmid p61F A. The 3⬘ end of the proviral DNA, including theenvgene and the 3⬘long terminal repeat, was cloned in pBSK at theEcoRI andXbaI sites to generate plasmid p61F B. Cotransfection with plasmids p61F A and p61F B renders virus infectious after in vivo ligation (21). MT-4 cells were a gift of D. D. Richman (University of
* Corresponding author. Mailing address: Centro de Biologı´a
Molecular “Severo Ochoa,” Consejo Superior de Investigaciones
Ci-entı´ficas-Universidad Auto´noma de Madrid, Cantoblanco, 28049
Madrid, Spain. Phone: 34-91-3978477. Fax: 34-91-3974799. E-mail:
6293
on November 9, 2019 by guest
http://jvi.asm.org/
California at San Diego). Mutant HIV clones were obtained by site-directed mutagenesis. A 4,385-bpPstI-SalI fragment derived from p61F A, which contains the entirepolgene, was cloned in the mutagenesis vector pALTER-1 (Promega). In vitro mutagenesis reactions were carried out by following the manufacturer’s instructions, and mutations Y115W, Y115L, Y115A, and Y115D were intro-duced with the mutagenic oligonucleotides previously described (17, 18). The mutagenizedPstI-SalI insert was then excised and cloned back into p61F A for its use in the transfection experiments.
DNA transfection experiments.Transfections were performed as previously
reported (21). Briefly, 3⫻106COS-1 cells were electroporated with 10g of
subgenomic clones p61F A and p61F B. Forty-eight hours after transfection, 4⫻ 106MT-4 cells were added to the culture. Viral replication was monitored by RT
activity and p24 antigen detection in culture supernatants. Virion-associated RT activity was determined as described by Willey et al. (34), after 10l of trans-fection supernatant was mixed with 50l of an RT reaction mixture which contained a template-primer of poly(rA) (5 g/ml) and oligo(dT)12–18(1.57
g/ml) in 50 mM Tris (pH 7.8)–75 mM KCl–2 mM dithiothreitol–5 mM MgCl2–
0.05% Nonidet P-40–0.5Ci of [32P]dTTP (800 Ci/mmol). In some assays, cold
dTTP was added to the reaction mixture to achieve a final nucleotide concen-tration of 10M. Production of p24 antigen in cell-free supernatants was mea-sured with an antigen capture kit (SAIC Frederick, AIDS Vaccine Program).
Infections. Supernatants from transfection experiments collected on days
when maximum levels of p24 antigen were detected were used to infect 5⫻106
fresh MT-4 cells. Viruses were grown to obtain a stock, which was then titrated in an MT-4 plaque assay (9, 29). Infections were carried out at a multiplicity of infection of 0.01 PFU per cell, and viral replication was monitored by measuring RT activity in culture supernatants and determination of cell viability by the trypan blue staining method.
RNA isolation, amplification, and nucleotide sequence analysis.Culture
su-pernatants were passed through 0.45-m-pore-size filters and treated with DNase A for 30 min to eliminate input DNA. Total RNA was obtained from 20 l of treated transfection supernatants (2). RNA amplification was done with the one-tube RT-PCR PCR system (Titan; Boehringer Mannheim) with primers 47RU (5⬘GTATTAGTAGGACCTACACCT 3⬘, positions 2055 to 2075) and 59RD (5⬘ATGATTCCTAATGCATATTGTGAGT 3⬘, complementary to posi-tions 3622 to 3646). Primers are numbered according to the sequence of Ratner et al. (27). These primers amplify a 1,591-bp fragment. After reverse transcrip-tion at 50°C for 30 min and a 5-min incubatranscrip-tion at 94°C, samples were subjected to 35 rounds of amplification. Each cycle comprised a 1-min denaturation step at 94°C, a 1-min annealing step at 55°C, and a 2-min extension step at 72°C. Nucleotide sequence analysis of the amplified product was performed with the fmol DNA sequencing system (Promega, Madison, Wis.) with the following primers: 14RD (5⬘GCACGATATCTAATCCTGGTGTCTCA 3⬘, complemen-tary to positions 2540 to 2561), 3⬘RU (5⬘GCGGGATCCTGAAAATCCATAC AATACTC 3⬘, positions 2278 to 2304), 15⬘RU (5⬘TAGATATCAGTACAAT GTGCTTCCAC 3⬘, positions 2555 to 2580), 58RU (5⬘GCCAGAAAAAGAC AGCTGGACTGT 3⬘, positions 2867 to 2890), and 59RD (5⬘ATGATTCCTA ATGCATATTGTGAGT 3⬘, complementary to positions 3622 to 3646). When sequences revealed a heterogeneity at a given position, the relative proportion of each mutant was estimated by densitometry of the specific bands in the sequenc-ing gel, with a PhosphorImager apparatus and the PCBAS program.
Expression and purification of recombinant HIV-1 RTs.Recombinant HIV-1
RT mutants used in this study were previously described (17, 18), except for those having Ile-230 instead of Met. In this case, the mutation was introduced with the Altered Sites in vitro mutagenesis system kit from Promega, following the manufacturer’s instructions. The mutagenic oligonucleotide used was 5⬘ GGAGTTCATAACCGATCCAAAGGAATGG 3⬘, and the introduced muta-tion was confirmed by DNA sequencing. Purificamuta-tion of mutant and wild-type (WT) HIV-1 RTs was carried out after independent expression of their subunits (p66 and p51), by following a previously described procedure (17). The 51-kDa subunit was obtained with an extension of 14 amino acid residues at its N-terminal end, including six consecutive histidines to facilitate its purification by metal chelate affinity chromatography. All RTs were purified as p66-p51 het-erodimers. Briefly, RTs were prepared by mixing the cell pellets of p66 and p51 clones at a 10:1-to-15:1 ratio. The bacteria were lysed, sonicated, and centri-fuged, and supernatants were applied to a nickel-nitriloacetic acid-agarose col-umn (Invitrogen Corporation, Carlsbad, Calif.). The colcol-umn was washed, and histidine-tagged RT was eluted with an imidazole gradient. RT-containing frac-tions were pooled; dialyzed against 50 mM Tris-HCl (pH 7.0) containing 25 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, and 10% glycerol (buffer A); and then passed through Q Sepharose (Amersham Pharmacia, Uppsala, Sweden). The RT was recovered in the nonbinding fraction and loaded onto Bio-Rex 70 (Bio-Rad Laboratories, Hercules, Calif.). RT was eluted with an NaCl gradient, and the RT-containing fractions were dialyzed against buffer A, concentrated in Cen-triprep-30 and Centricon-30 (Amicon, Beverly, Mass.) to less than 0.5 ml, and stored at⫺20°C. RT preparations were up to 90 to 95% pure, as judged by examination of Coomassie blue-stained gels.
Single nucleotide extension assays.Oligonucleotides PG5 (5⬘TGGTAGGG
CTATACAT 3⬘) and pT (5⬘GGATTTTAGACAGGAACGGT 3⬘) were labeled at their 5⬘termini with [␥-32P]ATP with T4 polynucleotide kinase, as previously
described (18). The phosphorylated primers were then annealed to templates. In the case of PG5, the template used was D2 (5⬘GGGATTAAATAAAATAGT
AAGAATGTATAGCCCTACCA 3⬘), a 38-mer synthetic oligonucleotide rep-resenting the HIV-1gagsequence that extends from nucleotides 1137 (5⬘end) to 1174 (3⬘ end) according to the sequence numbering of Ratner et al. (27). M13mp2 single-stranded DNA was the template used with oligonucleotide pT. The templates and their corresponding primers were annealed in 150 mM NaCl– 150 mM magnesium aspartate as described previously (18). Nucleotide incorpo-ration assays were performed in 20 l of 50 mM HEPES (pH 7.0)–15 mM NaCl–15 mM magnesium aspartate–130 mM KCH3COO–1 mM
dithiothrei-tol–5% polyethylene glycol 6000 (18). The template-primer concentration was 30 nM for D2/PG5 and 20 nM for M13mp2 single-stranded DNA/pT. The active enzyme concentration in these assays was around 6 nM. Reactions were initiated by incubating the enzyme with the corresponding annealed template-primer in the absence of dNTP (10 min at 37°C), followed by the addition of appropriate dNTPs at various concentrations. The reaction mixtures were incubated for 30 s at 37°C, and then the reactions were stopped by adding 8l of 10 mM EDTA in 90% formamide containing 3 mg of xylene cyanol FF per ml and 3 mg of bromophenol blue per ml. The extension products were resolved by electro-phoresis in 20% polyacrylamide–urea gels and quantitated with a BAS 1500 scanner. Elongation measurements were fitted to the Michaelis-Menten equa-tion, and thekcatandKmvalues were determined as previously described (17).
RESULTS
In vitro replication of HIV-1 with mutations affecting
Tyr-115 of RT.
To examine the effect on virus replication of several
mutations affecting Tyr-115 of HIV-1 RT, we introduced the
mutations Y115W, Y115L, Y115A, and Y115D into the
pro-viral genome. COS-1 cells were transfected with WT or
mu-tated proviral DNA, and after addition of MT-4 cells,
cocul-ture supernatants were monitored at various times for RT
activity and p24 antigen level. When WT DNA was used, RT
activity was detected around 12 days after transfection,
reach-ing a maximum level between days 14 and 18 (Fig. 1A), and
p24 was detected in the culture supernatants from day 9 (Fig.
FIG. 1. Replication kinetics of WT HIV-1 and Tyr-115 mutant virus in MT-4 cells. COS-1 cells were transfected by electroporation with 10g of each proviral DNA, as indicated in Materials and Methods. Forty-eight hours after transfec-tion, MT-4 cells were added. Samples were withdrawn from cultures transfected with WT provirus (F), mutant Y115W (E), and mock (no DNA) (Œ) at variousdays after transfection and then assessed for RT activity (A) and for the presence of p24 antigen (B).
on November 9, 2019 by guest
http://jvi.asm.org/
1B). In contrast, cultures transfected with mutants Y115L,
Y115A, and Y115D failed to produce any detectable RT
ac-tivity even after 52 days of culture and were found to be
negative for the presence of p24 (values indistinguishable from
those of the mock-transfection assays in Fig. 1). Mutant
Y115W displayed significant levels of RT activity from day 30
after transfection (Fig. 1A) and significant amounts of p24 in
culture supernatants from day 19 after transfection (Fig. 1B).
Supernatants of these cultures were able to infect MT-4 cells.
The data shown in Fig. 1 correspond to a single transfection
experiment, but similar results were obtained in three
addi-tional transfection experiments. The WT virus and the virus
recovered from transfections with Y115W were the only ones
that produced infectious progeny, although the detection of
RT activity in the supernatant was significantly delayed in the
case of Y115W, emerging more than 30 days after transfection
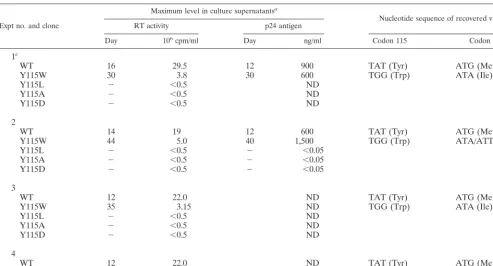
in the three other experiments (Table 1).
Mutation M230I restores the replication capacity of the
Y115W mutant.
The complete RT coding region of viruses
recovered from transfections with the Y115W mutant was
de-termined, after amplification of viral RNA from culture
super-natants obtained at the peak of RT activity. In all four
trans-fection experiments, only one mutation was observed in the RT
coding region, which resulted in the substitution of Ile for
Met-230 (Table 1). This amino acid change was produced by a
G-to-A transition at the third base of codon 230. In two
trans-fections, a mixture of A and T was detected at this position
(Table 1). In all cases, Trp-115 was maintained. To monitor the
kinetics of appearance of the Ile-230 variant, we carried out
RT-PCR and nucleotide sequence analysis with transfection
supernatants withdrawn on various days after transfection (Fig.
2). A weak band of amplification was obtained from
superna-tants collected on days 16 and 19 after transfection, indicating
a very low level of viral replication. By day 23, a significant
increase in the amplified band was observed. Nucleotide
se-quencing of the amplified products and a quantitation of the
relative proportions of Met and Ile at position 230 were also
carried out. In the supernatants of day 23, Ile-230 appeared
instead of Met, and this was coincident with a sharp increase in
the amount of RT-PCR-amplified material. The appearance of
substitution M230I correlated with an increase in p24 antigen
production (compare Fig. 1B and 2). In the peak of RT activity
(day 30 posttransfection of transfection experiment 1), more
than 95% of the viral population contained Ile at position 230.
These data reveal the rapid imposition of a mutant HIV-1 with
Ile at position 230. Mutations Y115W and M230I were the only
ones found in the RT-encoding region of viruses recovered in
any of the four experiments in which cells were transfected
with mutant Y115W. Thus, substitution M230I appears to
re-store the replication capacity of mutant Y115W, as a
compen-satory mutation for the presence of Trp at RT position 115.
Enzymatic characterization of RT mutants.
To study
whether M230I affected RT activity, steady-state kinetic
pa-rameters for the incorporation of nucleotides at the 3
⬘
end of
the primer were obtained for the WT RT, the single mutants
Y115W and M230I, and the double mutant Y115W/M230I
(Table 2). All of them displayed roughly similar
k
catvalues.
However, the
K
mvalues for the incorporation of dNTP were
[image:3.612.57.551.84.350.2]75- to 130-fold higher for the single mutant Y115W than for
the WT enzyme, resulting in a reduced catalytic efficiency,
TABLE 1. Cell transfections with HIV-1 cDNA clones
Expt no. and clone
Maximum level in culture supernatantsa
Nucleotide sequence of recovered virusb
RT activity p24 antigen
Day 106cpm/ml Day ng/ml Codon 115 Codon 230
1
cWT
16
29.5
12
900
TAT (Tyr)
ATG (Met)
Y115W
30
3.8
30
600
TGG (Trp)
ATA (Ile)
Y115L
⫺
⬍0.5
ND
Y115A
⫺
⬍0.5
ND
Y115D
⫺
⬍0.5
ND
2
WT
14
19
12
600
TAT (Tyr)
ATG (Met)
Y115W
44
5.0
40
1,500
TGG (Trp)
ATA/ATT (Ile)
dY115L
⫺
⬍0.5
⫺
⬍0.05
Y115A
⫺
⬍0.5
⫺
⬍0.05
Y115D
⫺
⬍0.5
⫺
⬍0.05
3
WT
12
22.0
ND
TAT (Tyr)
ATG (Met)
Y115W
35
3.15
ND
TGG (Trp)
ATA (Ile)
Y115L
⫺
⬍0.5
ND
Y115A
⫺
⬍0.5
ND
Y115D
⫺
⬍0.5
ND
4
WT
12
22.0
ND
TAT (Tyr)
ATG (Met)
Y115W
45
2.0
ND
TGG (Trp)
ATA/ATT (Ile)
daRT activity and p24 antigen level were determined periodically in the cell culture medium of all transfections. Days and values reported were those obtained at the
peak of RT activity or p24 antigen. The minus sign indicates that levels were not distinguishable from background (mock-infected cells), at all times tested (up to 30 days after transfection in experiments 1 and 3 and up to 45 days in experiments 2 and 4). ND, not determined.
bThe sequences reported were obtained from viral RNA amplified from supernatants collected when maximum RT activity was detected. Procedures are described
in Materials and Methods.
cThe variation in RT activity and p24 antigen level following transfection and infection by WT and mutant Y115W in this experiment is depicted in Fig. 1. dATA/ATT means that a mixture of the two nucleotides at the third base of the codon was present, according to the band pattern observed in sequencing gels.
on November 9, 2019 by guest
http://jvi.asm.org/
which is the likely cause of the replication defect observed in
viruses harboring this mutation. On the other hand, WT RT
and single mutant M230I had similar affinities for the incoming
nucleotide. Interestingly, the presence of M230I together with
Y115W resulted in an enzyme with an increased catalytic
ef-ficiency, which displayed a 9- to 65-fold-higher affinity for
dNTP, relative to the single mutant Y115W. This effect can be
attributed to the presence of both amino acid substitutions in
the 66-kDa subunit of HIV-1 RT, as demonstrated by using
chimeric heterodimers where substitutions were introduced in
p66, p51, or both subunits (Table 2).
The lower levels of RT activity obtained from transfection
supernatants with viruses having both mutations, Y115W and
M230I (Fig. 1A), could be explained by the lower affinity for
dNTP of the double mutant than of the WT enzyme, since
those RT activity assays were carried out in the presence of
very low concentrations of dTTP. However, the high levels of
p24 antigen detected in transfection supernatants of WT and
Y115W (Fig. 1) suggested that viruses recovered from both
transfection experiments were viable and replicated normally.
The growth curves obtained upon infection of MT-4 cells with
WT HIV-1 and the double mutant Y115W/M230I show that
the two viruses had similar replication kinetics (Fig. 3).
Inter-estingly, the double-mutant virus was more sensitive to the
concentration of dTTP in the RT activity assay. In agreement
with the enzymological data, the differences in RT activity
between the WT HIV-1 and the double-mutant virus were
minimized when the concentration of dTTP in the assay was
increased.
DISCUSSION
Several lines of evidence support the role of Tyr-115 and
Met-230 in RT function. We have found that viruses harboring
the mutation Y115L, Y115A, or Y115D showed no signs of
virus replication (Table 1). Similar results were previously
re-ported for mutant Y115N (16). In the present study, we have
also shown that the Y115W variant replicates very poorly until
the emergence of mutation M230I. Tyr-115 is part of a
con-served motif in many polymerases (25). Tyr and Phe are the
only amino acids found at this position in retroviral RTs, and
Phe is the only residue which can replace Tyr-115 of HIV-1 RT
without causing a detrimental effect in its DNA polymerase
activity (17, 18). The Y115F appears in vitro after passage of
the virus in the presence of abacavir and confers low-level
resistance to this carbocyclic nucleoside inhibitor (32). In
Moloney murine leukemia virus RT, an aromatic amino acid
residue at this position is required for infectivity, since only the
WT Phe-155 or Tyr can support virus replication (7). Met-230
is part of a conserved motif found in many RNA-dependent
DNA polymerases, including RTs and telomerases (20, 25, 37).
All retroviral RTs have Met or Leu at this position. In the case
of HIV-1, the substitution of Leu for Met-230 has been
de-tected in virus that was passaged in cell culture, in the presence
of the nonnucleoside RT inhibitor delavirdine (23). Ile-230 has
been found in one nonproductive HIV-1 clone (5).
[image:4.612.76.267.71.244.2]In this report, we demonstrate that the substitution of Ile for
Met-230 restores the replication capacity of viruses having the
Y115W mutation in the RT to WT levels. To our knowledge,
this is the most dramatic compensatory effect described so far
for HIV-1 clones with a defect in polymerase function.
Com-pensatory mutations play a role in the acquisition of drug
resistance, by increasing viral fitness. However, they usually
appear in viruses which already display a significant replicative
capacity. Examples for RT are the substitution of Ile for Val-75
that improves the replication capacity of multidrug-resistant
viruses having the mutations Q151M and F77L (12) and the
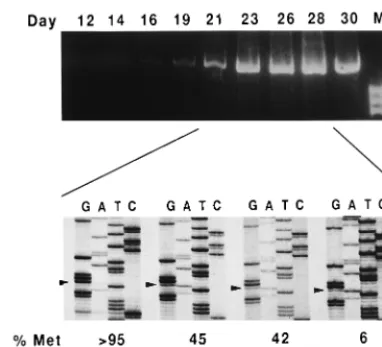
FIG. 2. Kinetics of imposition of Ile over Met-230 in viruses evolving aftertransfection with a clone harboring replacement Y115W. RT-PCR was carried out with transfection supernatants obtained at various days after transfection. Bands shown in lane M derive from digestion of⌽X174 DNA withHaeIII and correspond to fragments of 1,353, 1,078, and 872 bp, top to bottom, respectively. The relative amounts of Met and Ile at position 230 are shown below and were estimated by densitometry of the specific bands in the sequencing gels of the PCR products corresponding to days 21, 23, 26, and 28. Arrowheads indicate the position of the third base of codon 230 (G for Met-230 and A for Ile-230).
TABLE 2. Kinetic parameters for dNTP incorporation of WT and mutant RTs
aEnzyme
Value for template-primer:
D2/PG5 M13mp2 ssDNA/pT
Km(M) kcat(min⫺1) Km(M) kcat(min⫺1)
WT
0.173
⫾
0.027
b3.07
⫾
0.54
b0.082
⫾
0.032
2.54
⫾
0.11
Y115W
13.4
⫾
1.6
2.92
⫾
0.12
10.6
⫾
1.6
4.12
⫾
0.19
M230I
0.266
⫾
0.031
1.97
⫾
0.53
0.045
⫾
0.009
6.73
⫾
0.48
Y115W/M230I
1.49
⫾
0.09
3.87
⫾
0.35
0.162
⫾
0.021
3.06
⫾
0.86
p66
Y115W/p51
WT11.5
⫾
1.1
3.76
⫾
0.11
7.5
⫾
0.6
3.14
⫾
0.06
p66
Y115W/p51
M230I13.6
⫾
1.1
3.60
⫾
0.09
12.5
⫾
1.2
4.65
⫾
0.51
p66
Y115W/M230I/p51
WT3.09
⫾
0.45
7.52
⫾
3.50
0.226
⫾
0.013
7.48
⫾
0.12
aThe incorporated nucleotides at the 3⬘end of the primer were dTTP for template-primer D2/PG5 and dATP for M13mp2 ssDNA/pT. Data shown are the mean
values⫾standard deviations, obtained from a nonlinear least-squares fit of the kinetics data to the Michaelis-Menten equation. Each of the experiments was performed independently at least twice. The substitutions present in each subunit of the heterodimer are shown by a superscript.
bReported data for this enzyme with template-primer D2/PG5 were taken from reference 19.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.56.551.581.694.2]substitution L74V, which, besides V75I, can stimulate viral
growth in quinoxaline-resistant viruses having substitution
G190E (4, 14). In both cases, the compensatory mutation
ap-pears in the FINGERS subdomain and could affect the
posi-tioning of the template in the binding cleft of HIV-1 RT. Our
results show that the substitution Y115W impairs RT activity
by decreasing the dNTP binding affinity of the polymerase, but
the acquisition of the M230I mutation compensates for the
dNTP binding defect. The crystallographic structure of HIV-1
RT reveals that Tyr-115 is located close to the polymerase
active site, while Met-230 is part of the primer grip which forms
the

12-

13 hairpin, including residues 227 to 235. The ribose
moiety of the incoming dNTP projects into a small pocket lined
by the side chains of Asp-113, Tyr-115, Phe-116, and Gln-151
(11). Several amino acid substitutions associated with
resis-tance to nucleoside analogue inhibitors lie within the dNTP
binding site (e.g., K65R, L74V, M184V, F116Y, and Q151M)
(1, 6, 30). Met-230 interacts with the 3
⬘
-terminal phosphate of
the primer, making extensive contacts with nucleotides of the
3
⬘
primer terminus and with the side chain of Tyr-183 (6, 11,
13). It has been suggested that binding of nonnucleoside RT
inhibitors could induce a conformational change leading to
repositioning of the primer grip (6), but none of the amino
acids surrounding Met-230 appears to be involved in major
drug resistance mutations. Met-230 influences the proper
po-sitioning of the RT on the template-primer. Its replacement by
Ala led to alterations of both polymerase and RNase H
activ-ities (8, 24) and affected RNA primer selection, a step which is
important at the initiation of minus-strand DNA synthesis
(26). Not surprisingly, the mutation M230A caused severe
de-fects in proviral DNA synthesis and rendered virus
noninfec-tious (38). Crystallographic data have implicated the primer
grip in orienting the primer terminus for nucleophilic attack on
an incoming dNTP. In agreement with this proposal, it has
been shown that substitution of Ala for Met-230 produces a
significant decrease in the dNTP binding affinity of the enzyme
(35), an effect which is also observed with mutant HIV-1 RTs
having nonconservative substitutions at position 115 (17, 18).
Therefore, it is likely that Y115W leads to a less favorable
positioning of the
␣
-phosphate dNTP for attack by the 3
⬘
OH
of the primer terminus, and mutation M230I restores, at least
in part, the proper alignment required for catalysis.
Restoration of RT function by direct reversion of Trp-115 to
Tyr was probably limited by the two mutations needed to
convert the triplet for Trp (UGG) into a triplet for Tyr (UAU
or UAC). The two required mutations had to occur
simulta-neously in the same genomic RNA molecule since G
3
A alone
would lead to the termination codon UAG, and G
3
U (or
G
3
C) alone would produce an RT with Cys at position 115, an
enzyme with a very high
K
mfor dNTP binding (18). Therefore,
the genetic makeup of the virus (its initial position in sequence
space) may have prompted replacement M230I, thereby
re-vealing the implication of this residue in RT catalysis. The
results described in this paper illustrate the enormous
adapta-tive potential of HIV-1 and should guide future mutagenesis
experiments with the aim of gaining a better knowledge of the
DNA polymerization mechanism catalyzed by RTs, by
com-bining a biochemical with an evolutionary approach.
ACKNOWLEDGMENTS
We thank Gema Go´mez Mariano for help with DNA sequencing.
This work was supported by Fondo de Investigacio´n Sanitaria grants
98/0054-01, -02, and -03 and by an institutional grant of Fundacio´n
Ramo´n Areces to Centro de Biologı´a Molecular “Severo Ochoa.”
REFERENCES
1.Arts, E. J., and M. A. Wainberg.1996. Human immunodeficiency virus type
1 reverse transcriptase and early events in reverse transcription. Adv. Virus Res.46:97–163.
2.Boom, R., C. J. A. Sol, M. M. M. Salimans, C. L. Jansen, P. M. E.
Wertheim-van Dillen, and J. Wertheim-van der Noordaa.1990. Rapid and simple method for
purification of nucleic acids. J. Clin. Microbiol.28:495–503.
3.Boyer, P. L., A. L. Ferris, P. Clark, J. Whitmer, P. Frank, C. Tantillo, E.
Arnold, and S. H. Hughes.1994. Mutational analysis of the fingers and palm
subdomains of human immunodeficiency virus type-1 (HIV-1) reverse tran-scriptase. J. Mol. Biol.243:472–483.
4.Boyer, P. L., H.-Q. Gao, and S. H. Hughes.1998. A mutation at position 190
of human immunodeficiency virus type 1 reverse transcriptase interacts with mutations at positions 74 and 75 via the template-primer. Antimicrob. Agents Chemother.42:447–452.
5.Carlini, F., M. Federic, M. Equestre, S. Ricci, G. Ratti, Q. Zibai, P. Verani,
and G. B. Rossi.1992. Sequence analysis of HIV-1 proviral DNA from a non
producer chronically infected HUT-78 cellular clone. J. Viral Dis.1:40–55.
6.Ding, J., K. Das, Y. Hsiou, S. G. Sarafianos, A. D. Clark, Jr., A.
Jacobo-Molina, C. Tantillo, S. H. Hughes, and E. Arnold.1998. Structural and
functional implications of the polymerase active site region in a complex of HIV-1 RT with a double-stranded DNA template-primer and an antibody Fab fragment at 2.8 Å resolution. J. Mol. Biol.284:1095–1111.
7.Gao, G., and S. P. Goff.1998. Replication defect of Moloney murine
leuke-mia virus with a mutant reverse transcriptase that can incorporate ribonucle-otides and deoxyribonucleribonucle-otides. J. Virol.72:5905–5911.
FIG. 3. Replication kinetics of WT HIV-1 and the double mutant Y115W/ M230I. MT-4 cells (106) were infected at a multiplicity of infection of 0.01 PFU per cell. Infections were monitored by measuring RT activity (A and B) and cell viability (C). RT activity was determined either in the presence of 12.5 nM dTTP (A) or in the presence of 10M dTTP (B).F, WT HIV-1;E, double mutant
Y115W/M230I;Œ, mock.
on November 9, 2019 by guest
http://jvi.asm.org/
8.Ghosh, M., P. S. Jacques, D. W. Rodgers, M. Ottmann, J.-L. Darlix, and
S. F. J. Le Grice.1996. Alterations to the primer grip of p66 HIV-1 reverse
transcriptase and their consequences for template-primer utilization. Bio-chemistry35:8553–8562.
9.Harada, S., Y. Koyanagi, and N. Yamamoto.1985. Infection of HTLV-III/
LAV in HTLV-I-carrying cells MT-2 and MT-4 and application in a plaque assay. Science229:563–566.
10.Hottiger, M., and U. Hu¨bscher.1996. Human immunodeficiency virus type 1
reverse transcriptase. Biol. Chem. Hoppe-Seyler377:97–120.
11.Huang, H., R. Chopra, G. L. Verdine, and S. C. Harrison.1998. Structure of
a covalently trapped catalytic complex of HIV-1 reverse transcriptase: im-plications for drug resistance. Science282:1669–1675.
12. Iversen, A. K. N., R. W. Shafer, K. Wehrly, M. A. Winters, J. I. Mullins, B.
Chesebro, and T. C. Merigan.1996. Multidrug-resistant human
immunode-ficiency virus type 1 strains resulting from combination antiretroviral ther-apy. J. Virol.70:1086–1090.
13. Jacobo-Molina, A., J. Ding, R. G. Nanni, A. D. Clark, Jr., X. Lu, C. Tantillo,
R. L. Williams, G. Kamer, A. L. Ferris, P. Clark, A. Hizi, S. H. Hughes, and
E. Arnold.1993. Crystal structure of human immunodeficiency virus type 1
reverse transcriptase complexed with double-stranded DNA at 3.0 Å reso-lution shows bent DNA. Proc. Natl. Acad. Sci. USA90:6320–6324.
14. Kleim, J.-P., M. Ro¨sner, I. Winkler, A. Paessens, R. Kirsch, Y. Hsiou, E.
Arnold, and G. Riess.1996. Selective pressure of a quinoxaline
nonnucleo-side inhibitor of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (RT) on HIV-1 replication results in the emergence of nucle-oside RT-inhibitor-specific (RT Leu-743Val or Ile and Val-753Leu or Ile) HIV-1 mutants. Proc. Natl. Acad. Sci. USA93:34–38.
15. Kohlstaedt, L. A., J. Wang, J. M. Friedman, P. A. Rice, and T. A. Steitz.1992.
Crystal structure at 3.5 Å resolution of HIV-1 reverse transcriptase com-plexed with an inhibitor. Science256:1783–1790.
16. Larder, B. A., S. D. Kemp, and D. J. M. Purifoy.1989. Infectious potential
of human immunodeficiency virus type 1 reverse transcriptase mutants with altered inhibitor sensitivity. Proc. Natl. Acad. Sci. USA86:4803–4807.
17. Martı´n-Herna´ndez, A. M., E. Domingo, and L. Mene´ndez-Arias.1996.
Hu-man immunodeficiency virus type 1 reverse transcriptase: role of Tyr115 in deoxynucleotide binding and misinsertion fidelity of DNA synthesis. EMBO J.15:4434–4442.
18. Martı´n-Herna´ndez, A. M., M. Gutie´rrez-Rivas, E. Domingo, and L.
Mene´n-dez-Arias.1997. Mispair extension fidelity of human immunodeficiency virus
type 1 reverse transcriptases with amino acid substitutions affecting Tyr115. Nucleic Acids Res.25:1383–1389.
19. Mene´ndez-Arias, L.1998. Studies on the effects of truncating␣-helix E⬘of
p66 human immunodeficiency virus type 1 reverse transcriptase on template-primer binding and fidelity of DNA synthesis. Biochemistry37:16636–16644.
20. Nakamura, T. M., G. B. Morin, K. B. Chapman, S. L. Weinrich, W. H.
Andrews, J. Lingner, C. B. Harley, and T. R. Cech.1997. Telomerase
cata-lytic subunit homologs from fission yeast and human. Science277:955–959.
21. Olivares, I., G. Shaw, and C. Lo´pez-Galı´ndez.1997. Phenotypic switch in a
Spanish HIV type 1 isolate on serial passage on MT-4 cells. AIDS Res. Hum. Retroviruses13:979–984.
22. Olivares, I., C. Casado Herrero, M. D. Iglesias-Ussel, U. Dietrich, and C.
Lo´pez-Galı´ndez.1998. Complete sequence of an infectious molecular clone
derived from a Spanish HIV type I isolate. AIDS Res. Hum. Retroviruses
14:1649–1651.
23. Olmsted, R. A., D. E. Slade, L. A. Kopta, S. M. Poppe, T. J. Poel, S. W.
Newport, K. B. Rank, C. Biles, R. A. Morge, T. J. Dueweke, Y. Yagi, D. L.
Romero, R. C. Thomas, S. K. Sharma, and W. G. Tarpley.1996.
(Alkylamin-o)piperidine bis(heteroaryl)piperizine analogs are potent, broad-spectrum nonnucleoside reverse transcriptase inhibitors of drug-resistant isolates of human immunodeficiency virus type 1 (HIV-1) and select for drug-resistant
variants of HIV-1IIIBwith reduced replication phenotypes. J. Virol.70:3698–
3705.
24.Palaniappan, C., M. Wisniewski, P. S. Jacques, S. F. J. Le Grice, P. J. Fay,
and R. A. Bambara.1997. Mutations within the primer grip region of HIV-1
reverse transcriptase result in loss of RNase H function. J. Biol. Chem.
272:11157–11164.
25.Poch, O., I. Sauvaget, M. Delarue, and N. Tordo.1989. Identification of four
conserved motifs among the RNA-dependent polymerase encoding ele-ments. EMBO J.8:3867–3874.
26.Powell, M. D., M. Ghosh, P. S. Jacques, K. J. Howard, S. F. J. Le Grice, and
J. G. Levin.1997. Alanine-scanning mutations in the “primer grip” of p66
HIV-1 reverse transcriptase result in selective loss of RNA priming activity. J. Biol. Chem.272:13262–13269.
27.Ratner, L., W. Haseltine, R. Patarca, K. J. Livak, B. Starcich, S. F. Josephs,
E. R. Doran, J. A. Rafalski, E. Whitehorn, K. Baumeister, L. Ivanoff, S. R. Petteway, Jr., M. L. Pearson, J. A. Lautenberger, T. S. Papas, J. Ghrayeb,
N. T. Chang, R. C. Gallo, and F. Wong-Staal.1985. Complete nucleotide
sequence of the AIDS virus, HTLV-III. Nature313:277–284.
28.Rodgers, D. W., S. J. Gamblin, B. A. Harris, S. Ray, J. S. Culp, B. Hellmig,
D. J. Woolf, C. Debouck, and S. C. Harrison.1995. The structure of
unli-ganded reverse transcriptase from the human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA92:1222–1226.
29. Sa´nchez-Palomino, S., J. M. Rojas, M. A. Martı´nez, E. M. Fenyo¨, R. Na´jera,
E. Domingo, and C. Lo´pez-Galı´ndez.1993. Dilute passage promotes
expres-sion of genetic and phenotypic variants of human immunodeficiency virus type 1 in cell culture. J. Virol.67:2938–2943.
30. Tantillo, C., J. Ding, A. Jacobo-Molina, R. G. Nanni, P. L. Boyer, S. H.
Hughes, R. Pauwels, K. Andries, P. A. J. Janssen, and E. Arnold.1994.
Location of anti-AIDS drug binding sites and resistance mutations in the three-dimensional structure of HIV-1 reverse transcriptase. Implications for mechanisms of drug inhibition and resistance. J. Mol. Biol.243:369–387.
31. Telesnitsky, A., and S. P. Goff.1997. Reverse transcriptase and the
genera-tion of retroviral DNA, p. 121–160.InJ. W. Coffin, S. H. Hughes, and H. E. Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, Plain-view, N.Y.
32. Tisdale, M., T. Alnadaf, and D. Cousens.1997. Combination of mutations in
human immunodeficiency virus type 1 reverse transcriptase required for resistance to the carbocyclic nucleoside 1592U89. Antimicrob. Agents Che-mother.41:1094–1098.
33. Wakefield, J. K., S. A. Jablonski, and C. D. Morrow.1992. In vitro enzymatic
activity of human immunodeficiency virus type 1 reverse transcriptase mu-tants in the highly conserved YMDD amino acid motif correlates with the infectious potential of the proviral genome. J. Virol.66:6806–6812.
34. Willey, R. L., D. H. Smith, L. A. Lasky, T. S. Theodore, P. L. Earl, D. Moss,
D. J. Capon, and M. A. Martin.1988. In vitro mutagenesis identifies a region
within the envelope gene of the human immunodeficiency virus that is critical for infectivity. J. Virol.62:139–147.
35. Wo¨hrl, B. M., R. Krebs, S. H. Thrall, S. F. J. Le Grice, A. J. Scheidig, and
R. S. Goody.1997. Kinetic analysis of four HIV-1 reverse transcriptase
enzymes mutated in the primer grip region of p66. Implications for DNA synthesis and dimerization. J. Biol. Chem.272:17581–17587.
36. Wrobel, J. A., S.-F. Chao, M. J. Conrad, J. D. Merker, R. Swanstrom, G. J.
Pielak, and C. A. Hutchison III.1998. A genetic approach for identifying
critical residues in the fingers and palm subdomains of HIV-1 reverse tran-scriptase. Proc. Natl. Acad. Sci. USA95:638–645.
37. Xiong, Y., and T. H. Eickbush.1990. Origin and evolution of retroelements
based upon their reverse transcriptase sequences. EMBO J.9:3353–3362.
38. Yu, Q., M. Ottmann, C. Pechoux, S. Le Grice, and J.-L. Darlix.1998.
Mutations in the primer grip of human immunodeficiency virus type 1 re-verse transcriptase impair proviral DNA synthesis and virion maturation. J. Virol.72:7676–7680.
on November 9, 2019 by guest
http://jvi.asm.org/