0022-538X/06/$08.00⫹0 doi:10.1128/JVI.00709-06
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Complementation of Human Immunodeficiency Virus Type 1
Replication by Intracellular Selection of
Escherichia coli
tRNA
3LysSupplied in
trans
Anna McCulley and Casey D. Morrow*
Department of Cell Biology, University of Alabama at Birmingham, Birmingham, Alabama 35294-0024
Received 7 April 2006/Accepted 12 July 2006
Human immunodeficiency virus type 1 (HIV-1) exclusively selectstRNA3
Lys
as the primer for the initiation of reverse transcription, even though bothtRNA3
Lys
andtRNA1,2
Lys
are found in HIV-1 virions. Alteration of the HIV-1 primer-binding site (PBS) to be complementary to alternate tRNAs results in the use of those tRNAs for replication, indicating that primer complementarity with the PBS is an important determinant of primer selection. In previous studies, we have exploited this fact to develop a system in which yeast (Saccharomyces cerevisiae) tRNAPheis provided intransto complement the replication of HIV-1 with a PBS complementary to yeast tRNAPhe. Recent studies have demonstrated that the presence of lysyl-tRNA synthetase in HIV-1 virions might account for the preference for the selection oftRNA3
Lys
in HIV-1 replication. To establish a complemen-tation system more reflective of HIV-1 primer selection, we have altered the HIV-1 PBS to be complementary to theEscherichia coli tRNA3
Lys
, which shares near identity with mammaliantRNA3
Lys
except in the 3ⴕ-terminal 18-nucleotide sequence that binds to the PBS.E. coli tRNA3
Lys
expressed from a plasmid was aminoacylated in mammalian cells. Cotransfection of cells with a plasmid that encodesE. coli tRNA3
Lys
and a plasmid encoding an HIV-1 provirus with a PBS complementary toE. coli tRNA3
Lys
resulted in the production of infectious virus. A comparison of the two complementation systems revealed that higher levels of intracellularE. coli tRNA3
Lys
than of yeast tRNAPhe were needed to achieve equal levels of infectious virus, indicating that there was no preferential selection ofE. coli tRNA3
Lys
. To examine the specificity of tRNALysselection,E. coli tRNA3
Lys
was modified totRNA1,2
Lys
. This tRNA was also aminoacylated when expressed in mammalian cells and comple-mented the infectivity of HIV-1 at levels similar to those seen forE. coli tRNA3
Lys
. Additional mutations in the anticodon ofE. coli tRNA3
Lys
were constructed; these mutations did not significantly correlate with the capacity of the tRNA primer to complement infectivity of HIV-1, even though they had a drastic effect on the amino-acylation of the tRNAs. The results of these studies demonstrate thatE. coli tRNA3
Lys
provided intranscan complement HIV-1 genomes with the PBS altered toE. coli tRNA3
Lys
. However, the capacity of tRNA3
Lys
to interact with lysyl-tRNA synthetase does not entirely explain the enhanced preference for selection oftRNA3
Lys
for the replication of HIV-1.
The conversion of the single-stranded RNA genome of ret-roviruses into a double-stranded DNA intermediate prior to integration in the host cell chromosome requires a virally en-coded enzyme, reverse transcriptase, and a host cell tRNA (31). The initiation of reverse transcription (minus-strand viral DNA synthesis) begins with the extension of a cellular tRNA that is bound to a specific sequence of viral RNA genome known as the primer-binding site (PBS) (31). The PBS is an 18-nucleotide sequence that is located at the 5⬘end of the viral genome and is complementary to the 3⬘-terminal 18-nucleotide sequence of the primer tRNA (24, 25, 31). The primer tRNA is selected from the host cell intracellular milieu. Even though different retroviruses select different tRNA primers for reverse transcription, primer selection is conserved within a group of retroviruses (20, 21). Human immunodeficiency virus type 1 (HIV-1), like most lentiviruses, selectstRNA3Lys as the primer
for reverse transcription (20, 21).
Previous studies from this laboratory and others have shown that substitution of the PBS to be complementary to
alterna-tive tRNAs results in a virus that can transiently utilize this tRNA for replication (4, 17, 34). Since it is difficult to manip-ulate endogenous levels of tRNA, we have developed a complementation system that required tRNA to be provided in
trans for HIV-1 infectivity (13, 14, 37, 38). As described in
previous reports, the alteration of the HIV-1 PBS to be com-plementary to yeast (Saccharomyces cerevisiae) tRNAPhe
re-sulted in a virus that was noninfectious in mammalian cells unless yeast tRNAPhe was provided in trans. Expression of
yeast tRNAPhe from a cDNA resulted in a tRNA that had
undergone aminoacylation, nuclear transport, and inclusion into the cycle of host cell protein synthesis (15). The ability of the tRNA to be transported from the nucleus to the cytoplasm was critical for the selection of the tRNA as a primer (13).
While the system utilizing yeast tRNAPhehas revealed some
of the basic elements of the tRNA molecule required for primer selection, recent studies have suggested that HIV-1 has evolved several different mechanisms by whichtRNA3Lyscan be
preferentially selected for encapsidation (3, 9, 10, 16, 19). Early studies demonstrated that Gag-Pol was needed for the enrich-ment of HIV-1 virions with tRNA1,2,3Lys , since pseudovirions
which did not contain Gag-Pol incorporated a variety of tRNAs but showed no preference fortRNA1,2,3
Lys (16, 19).
Re-* Corresponding author. Mailing address: University of Alabama at Birmingham, Department of Cell Biology, 802 Kaul Building, 720 20th Street South, Birmingham, AL 35294-0024. Phone: (205) 934-5705. Fax: (205) 934-5733. E-mail: [email protected].
9641
on November 8, 2019 by guest
http://jvi.asm.org/
cently, lysyl-tRNA synthetase has been found within the HIV-1 virion (3, 9). The finding of lysyl-tRNA synthetase in the virion led to the postulation that a selective incorporation oftRNA1,2,3Lys
in the virions is facilitated through interaction with lysyl-tRNA synthetase. However, the chaperoning oftRNA1,2,3Lys into HIV-1
virions by lysyl-tRNA synthetase does not explain whytRNA3
Lys,
as opposed to tRNA1,2Lys, is preferentially selected for HIV-1
replication, since both tRNAs are present at relatively equal amounts in the HIV-1 virion (11). Previous studies from this laboratory and others have revealed that HIV-1 with a PBS altered totRNA1,2Lys had severely reduced capacity for
replica-tion and reverted back to utilizingtRNA3
Lysfollowing limited in
vitro culture (1, 12). In order fortRNA1,2Lys to be stably utilized
during replication, these viruses require supplementary muta-tions within the U5 with the altered PBS (1, 12). Even with the alterations to allow use oftRNA1,2
Lys, the virus does not replicate
with kinetics the same as those of the wild type. Collectively, the results of these studies suggest that tRNA3
Lys might have
unique properties that would facilitate the preferential selec-tion and use of this tRNA as a primer for HIV-1 replicaselec-tion. To further understand the preferential selection mechanism oftRNA3
Lys, it would be advantageous to have a
complementa-tion system that requires the seleccomplementa-tion of exogenously added
tRNA3
Lys. Since the levels of endogenoustRNA
3
Lysare difficult to
manipulate in mammalian cells, we have approached this prob-lem by developing a compprob-lementation system which requires the addition ofEscherichia coli tRNA3Lysintransto complement
the HIV-1 provirus with the PBS ofE. coli tRNA3
Lys.
Modifica-tions totRNA3Lys have been shown to be important for HIV-1
replication (6, 7).E. coli tRNA3
Lyshas many base modifications
that are either identical or similar to the corresponding mam-maliantRNA3
Lys base modifications, as well as a high level of
sequence identity to mammaliantRNA3Lys (28). In the current
study, we have found that the HIV-1 provirus with theE. coli tRNA3Lys PBS requires cotransfection of the plasmid encoding
theE. coli tRNA3
Lys to generate infectious virus. We have
dem-onstrated that E. coli tRNA3Lys undergoes aminoacylation.
Greater amounts of E. coli tRNA3
Lys than of yeast tRNAPhe
were required to achieve similar levels of complementation, indicating that no selective preference exists for E. coli tRNA3Lys, even though the tRNA can interact with lysyl-tRNA
synthetase. Furthermore, alteration of theE. coli tRNA3
Lys
an-ticodon totRNA1,2Lysresulted in complementation levels similar
to those found withE. coli tRNA3
Lys, suggesting that additional
features of primer selection, other than tRNA interaction with lysyl-tRNA synthetase, are probably important for the prefer-ential use oftRNA3Lys as a primer.
MATERIALS AND METHODS
Tissue culture.293HEK and JC53L cells were cultured in Dulbecco’s mod-ified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic (Gibco/BRL, Gaithersburg, MD). All cell cultures were maintained in a 37°C incubator supplemented with 5% CO2.
Proviral plasmids.The plasmid HXB2gpt, which encodes the HIV-1 provirus, was used for the substitution of the PBS in order to create proviral HIV-1 mutants containing a PBS complementary to the first 3⬘-terminal 18 nucleotides of either yeast tRNAPhe
orE. colitRNALys
(15, 26, 37). A previously constructed pUC119 PBS shuttle vector that contains an HIV-1 DNA fragment of the 5⬘long terminal repeat (LTR), PBS, and thegagleader region was used as a template for PBS mutagenesis (39). Mutagenesis of the PBS in the pUC119 PBS shuttle vector to yeast tRNAPhe
PBS was performed using the GeneEditor in vitro site-directed mutagenesis system (Promega, Madison, WI) with the following
mutagenic primer: 5⬘TCTCTAGCAGTGGTGCGAATTCTGTGGATGGAAA GCGAAAGGGAAACCAGAGGAGC3⬘. Mutagenesis of the PBS in the pUC119 PBS shuttle vector to PBS complementary toE. coli tRNALys
was performed using a QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA) with the following primers: 5⬘CTCTAGCAGTGGTGGGTCGTGC AGGACTTGAAAGCGAAAGGGAAACCAGA3⬘ (forward) and 5⬘TCTG GTTTCCCTTTCGCTTTCAAGTCCTGCACGACCCACCACTGCTAGAG3⬘ (reverse). The manufacturer’s instructions were followed for all mutagenic re-actions. The pUC119 PBS shuttle vector with the substituted PBS was digested with BssHII and HpaI enzymes in order to release an 868-bp fragment. The fragment was ligated back into pHXB2gpt, which was digested with BssHII and HpaI restriction enzymes. Resulting HIV-1 proviral mutants were labeled pHXB2(yPBSPhe) and pHXB2(EcPBSLys). All mutations were screened by
re-striction digests. Final mutants were verified by DNA sequencing.
tRNA plasmids.The yeast tRNAPhe
gene was constructed previously (14, 15). TheE. colitRNALysgene was constructed using PCR extension with the
follow-ing primers: 5⬘GCAGGGCTCGAGGTCCGGGTCGTTAGCTCAGTTGGTA GAGCAGTTGACTTTTAATC AATTGGTCGCAGG3⬘(forward) and 5⬘GCG GACGAAGCTTCCAAAAAATGGGTCGTGCAGGACTTGAACCTGCGA CCAATTGATTAAAAGTCAA3⬘(reverse). The PCR product was TA cloned into pGEM-T Easy vector (Promega, Madison, WI), and the resultant plasmid was digested with XhoI and HindIII in order to release theE. colitRNALys
gene (approximately 100 bp). TheE. colitRNALys
gene was then ligated into an LS9 plasmid, downstream of the human U6snRNA promoter, by use of the XhoI and HindIII restriction sites (14, 15, 18). The end product was a plasmid labeled pU6EcLys
. The anticodon bases ofE. colitRNALys
were substituted to CUU (corresponding to the anticodon oftRNA1,2
Lys
), CUA, UUA, and UCA by use of QuikChange site-directed mutagenesis (Stratagene, La Jolla, CA), pU6EcLysas
the template, and the following primers: 5⬘GGTAGAGCAGTTGACTCTTAA TCAATTGGTCGCAGGTT3⬘(CUU forward) and 5⬘AACCTGCGACCAATT GATTAAGAGTCAACTGCTCTACC3⬘(CUU reverse); 5⬘GGTAGAGCAGT TGACTCTAAATCAATTGGTCGCAGGTT3⬘(CUA forward) and 5⬘AACCT GCGACCAATTGATTTAGAGTCAACTGCTCTACC3⬘(CUA reverse); 5⬘GG TAGAGCAGTTGACTTTAAATCAATTGGTCGCAGGTT3⬘ (UUA forward) and 5⬘AACCTGCGACCAATTGATTTAAAGTCAACTGCTCTACC3⬘ (UUA reverse); and 5⬘GGTAGAGCAGTTGACTTCAAATCAATTGGTCGCAGG TT3⬘(UCA forward) and 5⬘AACCTGCGACCAATTGATTTGAAGTCAACTG CTCTACC3⬘(UCA reverse). The resultant plasmids were labeled pU6EcLys1,2
, pU6EcCUA, pU6EcUUA, and pU6EcUCA. All plasmids were screened with
restric-tion digest reacrestric-tions and verified by DNA sequencing.
Transfections.Complementation of HIV-1 proviral mutants was accomplished by cotransfection of tRNA-carrying plasmids with HIV-1 proviral mutants into 293HEK cells. Complementation of HIV-1 proviral genomes was described previously (13, 14, 37, 38). Cotransfection was achieved by using a calcium phosphate-based mammalian transfection kit (Stratagene, San Diego, CA) with the instructions scaled down for six-well plates. Briefly, 293HEK cells were seeded at a concentration of 2⫻105cells per well. The cells were cotransfected
with 500 ng of proviral plasmid and 50 ng, 100 ng, 500 ng, and 1,000 ng of tRNA-carrying plasmid 24 h later. At approximately 7 h posttransfection, the cells were washed once with 1⫻phosphate-buffered saline and supplied with fresh media. Supernatants were collected approximately 48 h posttransfection, centrifuged at 3,000⫻g, and used in a JC53L assay to determine luciferase activity, which has been determined to correlate to the units of the infectious virus that is being tested.
Viral infection.Supernatants collected from cotransfections were used in a JC53L reporter assay in order to determine infectious viral units (36). JC53L cells were seeded 24 h preinfection. Supernatants were diluted 1:3 in DMEM supplemented with 2% FBS and subsequently with two sequential 1:5 dilutions. The cells were incubated with the virus for 2 h in a 37°C incubator supplemented with 5% CO2. After 2 h, DMEM with 10% FBS was added to each well, and the
cells were incubated for an additional 48 h. To determine luciferase activity, cells were lysed using M-PER mammalian protein extraction reagent (Pierce, Rock-ford, IL), and approximately 20l of each lysed sample was transferred to a microplate. Reporter lysis buffer (Promega, Madison, WI) was added to each sample in the microplate, and the light intensity was measured using a LUMIstar luminometer (BMG Labtech, Durham, NC). Uninfected cells in wells repre-sented the background luciferase activity, which was subtracted from all other samples. Luciferase activity for pHXB2(yPBSPhe) and pHXB2(EcPBSLys),
with-out complementing tRNA, was set as the background activity and was subtracted from complementation samples. The luciferase values for two dilutions per sample were averaged. Relative light units (rLU) per ml were calculated by dividing the luciferase values by their corresponding dilution values.
9642 MCCULLEY AND MORROW J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
RNA isolation and tRNA analysis. 293HEK cells were transfected with pU6EcLys
, pU6EcLys1,2
, pU6EcCUA
, pU6EcUUA
, or pU6EcUCA
with the use of the mammalian transfection kit (Stratagene, San Diego, CA). The first set of transfected cells was used for the collection of total RNA, while the second set was used for the collection of aminoacyl tRNAs at 48 h posttransfection. The collection and isolation of total RNA and aminoacylated tRNAs was performed as previously described (13–15). Total RNA and aminoacyl tRNA were also isolated from mock-transfected 293HEK cells. Previously constructed oligonu-cleotide probes that are complementary to yeast tRNAPheand mammalian
tRNA3
Lys
were used for Northern analysis (14).E. colitRNALys
was detected using a [␥-32P]ATP-kinased oligo labeled with the use of Ready-to-go T4
polynucle-otide kinase (Amersham Pharmacia Biotech, Piscataway, N.J.) with the following probe: 5⬘-GGTCGTGCAGGATTCGAACCTGCGACCAATTGATTAAAAG TCAACTGCTCTACCAACTGAGCTAACGAC3⬘. Analyses of total RNA and aminoacyl tRNA were performed using acidic polyacrylamide gels and Northern blotting (13–15). The membranes were exposed to X-ray film, which was devel-oped using an SRX-101A developer (Konica, Wayne, NJ). Areas of the mem-brane corresponding to the bands on the X-ray film were excised and counted for radioactivity with an LS 5000TA scintillation counter (Beckman Coulter, Fuller-ton, CA).
In vitro transcription.In vitro transcripts were designed, and reactions were carried out as indicated in a MEGAshortscript T7 kit (Ambion, Austin, TX). In vitro transcripts for yeast tRNAPhewere prepared using the MEGAshortscript
T7 kit with previously obtained oligonucleotides (14). The following oligonucle-otides were used with a plasmid template pU6EcLysin order to produceE. coli
tRNA3
Lys
in vitro transcripts: 5⬘CTGCAGTAATACGACTCACTATAGGGTCG TTAGCTCAGTTGGT3⬘(T7EcLysforward) and 5⬘TGGTGGGTCGTGCAG
GACTTGAACCT3⬘ (T7EcLysreverse). The transcripts were diluted to yield
final concentrations of 5 ng, 10 ng, 20 ng, 40 ng, and 80 ng, which were used as standards in the Northern blots.
RESULTS
Construction of HIV-1 proviral genome with PBS comple-mentary toE. coli tRNA3
Lys
.tRNA3Lys contains several modified
nucleotides as a result of posttranscriptional processing. Pre-vious studies have shown that nucleotides within the anticodon loop impart a unique structure to this tRNA which could account for its preferential selection by HIV-1 as the primer (2, 32). The E. coli tRNA3Lys has near identity with mammalian
tRNA3Lysin the anticodon, T⌿C, and D-loop regions (Fig. 1A).
Differences between these two tRNAs exist mainly in the ac-ceptor stem region (3⬘-terminal 18 nucleotides), which inter-acts with the PBS of HIV-1. To expressE. coli tRNA3Lys, we have
utilized a plasmid similar to that previously reported for the expression of yeast tRNAPhe(14, 15). This plasmid contains a
[image:3.585.142.440.72.385.2]U6 snRNA polymerase III promoter and nucleotides at the 3⬘ terminus necessary for polymerase III termination. The tRNA gene was cloned in to the transcription cassette between the
FIG. 1. MammaliantRNA3Lys,Escherichia coli tRNA3Lys, and HIV-1 proviral PBS sequences. (A) Cloverleaf depictions of mammaliantRNA3Lysand E. coli tRNA3
Lys. The boldface nucleotides in the mammaliantRNA 3
Lysrepresent the 3⬘-terminal 18 nucleotides that are complementary to the HIV-1 PBS; the anticodon of the tRNA is boxed. The boldface nucleotides in theE. coli tRNA3Lysrepresent base differences from mammaliantRNA3Lys. Posttranscriptionally modified bases are shown for both tRNAs. (B) Schematic representation of the U5 region with proviral sequences from wild-type HXB2 and mutant HXB2(EcPBSLys). The underlined sequences correspond to the A-rich regions and the PBS, which are complementary to mammaliantRNA3Lysfor the wild-type HXB2 andE. coli tRNA3Lysfor the mutant HXB2(EcPBSLys).
on November 8, 2019 by guest
http://jvi.asm.org/
promoter and the termination signal. The PBS of the HIV-1 proviral genome (HXB2) was modified to be complementary to the 3⬘-terminal 18 nucleotides ofE. coli tRNA3Lys. The
nu-cleotides of the wild-type HIV-1 PBS and the altered HIV-1E. coli PBS have a degree of sequence variation sufficient to preclude the native tRNA primer from binding to the altered HIV-1 PBS (Fig. 1B).
Complementation of HIV-1 genomes by tRNA supplied in trans.Previous studies from our laboratory have shown that a mutation within the first 9 nucleotides of the PBS can have a drastic impact on the infectivity of wild-type HIV (33). Thus, the nucleotide variation that exists between the wild type HIV-1 PBS and the altered HIV-1 PBS, as well as between the 3⬘-terminal 18 nucleotides ofE. coli tRNA3
Lys and mammalian
tRNA3Lys, should preclude the use of the mammaliantRNA3Lysby
the altered HIV-1. To determine whether theE. coli tRNA3
Lys
would complement the replication of the altered HIV-1 pro-viral genome (PBS toE. coli tRNA3
Lys), cotransfection
experi-ments were done with the proviral plasmid (pHXB2EcPBSLys)
containing the altered PBS and with different amounts of the plasmid (pU6EcLys) encodingE. coli tRNA
3
Lys. For comparison,
we utilized the HIV-1 provirus in which the PBS was altered to be complementary to yeast tRNAPhe and plasmid (pU6Phe)
encoding the cDNA of yeast tRNAPhe. For these studies, the
production of infectious virus was measured by using a JC53L assay, in which a HeLa indicator cell line was infected with viruses recovered from cotransfections. The indicator cell line contains a luciferase gene under the control of the HIV-1 LTR. The expression of luciferase is dependent upon infection, reverse transcription, and expression of Tat (5, 36). Consistent
with our previously reported results with yeast tRNAPhe, we
found that no infectious virus was produced unless the plasmid encoding yeast tRNAPhe was provided in the cotransfection
and that increasing the amounts of plasmid encoding yeast tRNAPheresulted in an increase in infectious virus, as
deter-mined by the luciferase activity induced in JC53L cells, to a level of approximately 106over background (proviral plasmid
transfected without tRNAPhe) (Fig. 2A and B) (14, 15). We
next tested the complementation system, which requires co-transfection of HIV-1 proviral plasmid containing a PBS toE.
coli tRNA3Lys in conjunction with the plasmid encodingE. coli
tRNA3
Lys. We obtained a low basal level of luciferase activity
(approximately 1,000 light units) after transfection of the pro-viral plasmid alone, in the absence of the plasmid encodingE.
coli tRNA3Lys. Cotransfection of theE. coli tRNA3Lys plasmid in
conjunction with the altered HIV-1 proviral plasmid (PBS to
E. coli tRNA3Lys) resulted in a level of production of infectious
virus that was approximately fivefold lower than that of wild-type HIV-1 (Fig. 2A and B). Increasing the amounts of theE.
coli tRNA3
Lysplasmid in cotransfections resulted in an increase
of infectious virus that reached a plateau at a level approxi-mately 2⫻105greater than that of the background control (no
E. coli tRNA3Lys) (Fig. 2B). Surprisingly, the overall levels of the
infectious virus generated in theE. coli tRNA3
Lys
complemen-tation system under these conditions were approximately five-fold less than those for the same system with yeast tRNAPhe
(Fig. 2B).
We next wanted to investigate the reason for the lower complementation in theE. coli tRNA3Lys system. In previous
[image:4.585.109.482.70.244.2]studies, we have shown that the aminoacylation of yeast
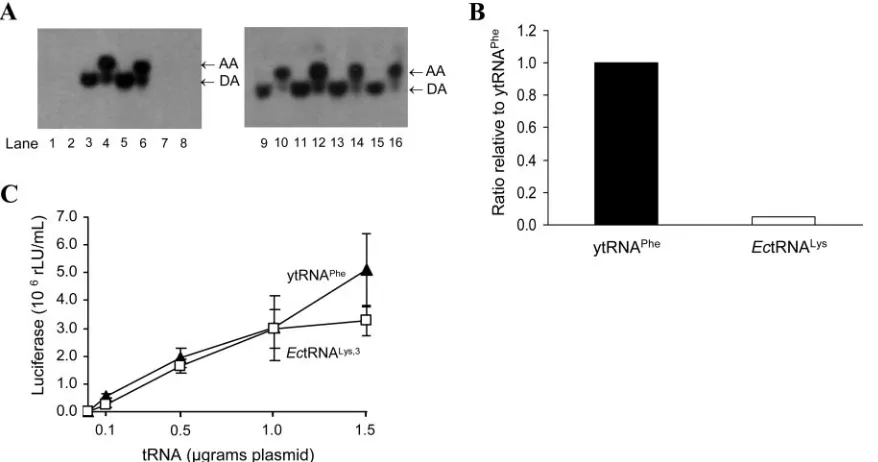
FIG. 2. Complementation of HXB2(EcPBSLys) with the plasmid that encodesE. coli tRNA 3
Lys and of HXB2(yPBSPhe) with the plasmid that encodes yeast tRNAPhe. (A) Representation of luciferase activity obtained from JC53L cells after infection with viruses that were collected from cotransfections of 293HEK cells with 500 ng of HIV-1 proviral plasmids in the presence or the absence of 500 ng of plasmid encoding the specified tRNA, relative to that for wild-type HIV-1 transfected at 500 ng. Dilutions of collected supernatants that were acquired from cotransfections were used to infect the JC53L cell line which contains a luciferase gene under the transcriptional control of the HIV-1 LTR (5, 36). (B) Luciferase activity obtained from JC53L cells after infection with viruses that were collected from cotransfections of 293HEK cells with 500 ng of proviral plasmids and tRNA plasmids that were titrated in at the indicated quantities. Luciferase activity, in rLU/ml, for the complementation of plasmid HXB2(yPBSPhe) with pU6Pheis represented by closed triangles, and that of plasmid HXB2(EcPBSLys) with pU6EcLysis represented by open squares. Wild-type HXB2 (500 ng and no tRNA) is represented by a closed square. Background luciferase activity obtained from mock-transfected cultures was subtracted from each sample. Background luciferase activity obtained from HXB2(yPBSPhe) alone was subtracted from all comple-mentation samples of HXB2(yPBSPhe) with yeast tRNAPhe, while background luciferase activity obtained from HXB2(EcPBSLys) alone was subtracted from all complementation samples of HXB2(EcPBSLys) withE. coli tRNA
3
Lys. The data denote means⫾standard deviations derived from three independent experiments.
9644 MCCULLEY AND MORROW J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
tRNAPhewas an important element in facilitating the selection
of this tRNA as a primer for HIV-1 replication (14, 15). One explanation for the lower complementation activity ofE. coli
tRNA3
Lys could be that it does not undergo aminoacylation in
mammalian cells. However, Schimmel’s group previously re-ported that E. coli tRNA3
Lys is aminoacylated by mammalian
lysyl-tRNA synthetase (27). To confirm this result, we analyzed the aminoacylation status of E. coli tRNA3
Lys in mammalian
cells. Following the transfection of the pU6EcLys plasmid
en-coding the cDNA forE. coli tRNA3
Lys, we found that majority of
the E. coli tRNA3Lys was aminoacylated (Fig. 3A); the minor
levels of deacylatedE. coli tRNA3
Lys noted in this experiment
were also found by analysis of yeast tRNAPhein cells
trans-fected with the pU6Pheplasmid encoding the cDNA for yeast
tRNAPheand were possibly due to the hydrolysis of the amino
acid-tRNA bond during the processing of the RNA sample.
Next, we compared the total amounts of yeast tRNAPheandE.
coli tRNA3
Lys generated in cells following transfection of their
respective cDNAs. In this case, we titrated in various amounts of each plasmid DNA encoding the tRNAs and determined the quantity of tRNA molecules compared to known standards generated through in vitro transcription. Surprisingly, we found that the levels ofE. coli tRNA3Lys were approximately 20
times less than those for yeast tRNAPhe(Fig. 3B). The reason
for this difference in tRNA amounts following transfection of plasmids with essentially the same promoter elements for the tRNAs was not clear, although this phenomenon could be due to differential regulation of tRNA pools in the cell for tRNALys
versus tRNAPhe. To follow up this result, we then adjusted the
levels of the plasmids encoding E. coli tRNA3
Lys and yeast
tRNAPheto give equal levels of production of infectious virus
[image:5.585.76.515.69.300.2](Fig. 3C). Under these conditions, we found that from the four
FIG. 3. Expression ofE. coli tRNA3Lysin mammalian cells. (A) Analysis of aminoacylation forE. coli tRNA3LysandE. coli tRNA1,2Lys. The migration of the aminoacylated (AA) and deacylated (DA) samples is shown. Cytoplasmic tRNAs were collected from 293HEK cells that were transfected with pU6Phe, pU6EcLys, and pU6EcLys1,2. All cytoplasmic RNA was isolated under acidic conditions, and 3.7g of the RNA was loaded per well. Lanes 1, 2, 9, and 10 were loaded with cytoplasmic RNA from mock transfection; lanes 3, 4, 11, and 12 were loaded with cytoplasmic RNA from pU6EcLystransfection; lanes 5, 6, 13, and 14 were loaded with cytoplasmic RNA from pU6EcLys1,2transfection; and lanes 7, 8, 15, and 16 were loaded with cytoplasmic RNA from pU6Phetransfection. Lanes 1 to 8 were probed forE. coli tRNA
3
Lys, while lanes 9 to 16 were probed for mammaliantRNA3Lys. Deacylated controls were prepared by adjustment of pH to basic conditions and incubation for 1 h at 42°C. Deacylated samples are shown in lanes 1, 3, 5, 7, 9, 11, 13, and 15. The exposure times for the blots varied. (B) Relative ratio ofE. coli tRNA3Lysmolecules to yeast tRNAPhe. 293HEK cells were transfected with 500 ng of pU6EcLysand pU6Phe. Total RNA was collected, and 15g was loaded per lane (Northern blot). In vitro-transcribed standards ofE. colitRNALysand yeast tRNAPhewere loaded at 5 ng, 10 ng, 20 ng, 40 ng, and 80 ng per lane, respectively. The blots were probed forE. colitRNALysand yeast tRNAPheand exposed to X-ray film. Areas of the membrane corresponding to the bands on film were excised and counted for radioactivity with a scintillation counter. Known amounts of in vitro-transcribed tRNA were used to generate a standard curve (R2⫽0.99). Using this curve, we found that the amount of tRNA molecules per sample forE. coli tRNA
3
Lyswas 0.42 ng per 15g total RNA and that for yeast tRNAPhewas 8.91 ng per 15g total RNA. (C) Luciferase activity obtained from JC53L cells after infection with viruses that were collected from cotransfections of 293HEK cells with 500 ng of proviral plasmids and tRNA plasmids that were titrated in at the indicated quantities. Note that yeast tRNAPhehad 20 times less plasmid DNA thanE. coli tRNA
3
Lys to normalize amounts of intracellularE. coli tRNA3Lysand yeast tRNAPhe. Subsequent analysis of intracellular levels of each tRNA expressed at each concentration revealed four times moreE. coli tRNA3
Lysthan yeast tRNAPhe. Carrier DNA (pUC19) was included with yeast tRNAPheplasmid to account for lower DNA concentrations during calcium phosphate cotransfections. Luciferase activity, in rLU/ml, for complementation of plasmid HXB2(yPBSPhe) with pU6Pheis represented by closed triangles, and that of plasmid HXB2(EcPBSLys) with pU6EcLysis represented by open squares. Background luciferase activity obtained from mock-transfected cultures was subtracted from each sample. Background luciferase activity obtained from HXB2(yPBSPhe) alone was subtracted from all complementation samples of HXB2(yPBSPhe) with yeast tRNAPhe, while background luciferase activity obtained from HXB2(EcPBSLys) alone was subtracted from all complementation samples of HXB2(EcPBSLys) withE. coli tRNA
3 Lys. The data denote means⫾standard deviations derived from three independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
amounts of tRNA plasmids transfected, an average of four times more intracellularE. coli tRNA3Lys than yeast tRNAPhe
was needed for the production of equal amounts of virus. Thus, we conclude thatE. coli tRNA3Lys can function as a primer for
HIV-1 but exhibits no enhanced selection/complementation compared to that for yeast tRNAPheor that for mammalian
tRNALys.
Complementation of HIV-1 withE. coli tRNA1,2
Lys
provided in trans. We next wanted to resolve the question of whether HIV-1 with a PBS complementary toE. coli tRNA3Lys would
show a preferential selection forE. coli tRNA3
LysversusE. coli
tRNA1,2Lys. Earlier studies have shown that although bothtRNA3Lys
and tRNA1,2
Lys are found in HIV-1 virions, HIV-1 has a clear
preference fortRNA3Lys, since the alteration of the PBS to be
complementary totRNA1,2
Lys did not result in a virus that could
utilize tRNA1,2Lys (1, 12). Mutations within U5, or the primer
activation site, are required for this virus to maintain a PBS complementary totRNA1,2Lys following in vitro culturing;
how-ever, these viruses grow more slowly than the wild type (1, 12, 23). As a result, HIV-1 has evolved a clear preference for the selection oftRNA3
Lys overtRNA
1,2
Lys. To determine if this is also
the case for the HIV-1 provirus designed to useE. coli tRNA3Lys,
we modified the anticodon region ofE. coli tRNA3
Lys so that it
corresponded to that fortRNA1,2Lys. The anticodon fortRNA1,2Lysis
CUU, whereas the anticodon fortRNA3
Lysis UUU (Fig. 4). We
first determined if this anticodon base mutation would affect the capacity of thisE. coli tRNA1,2
Lysto be aminoacylated, given
that the anticodon of tRNALys is also an important identity
element for synthetase recognition. We analyzed the amino-acylation status ofE. coli tRNA1,2Lysgenerated from transfection.
No clear differences were observed between the level of ami-noacylation of E. coli tRNA3Lys and that for E. coli tRNA1,2Lys,
indicating that both tRNAs are competent to interact with mammalian lysyl-tRNA synthetase (Fig. 3A). Next, we tested the ability of theE. coli tRNA1,2
Lysto complement the replication
of HIV-1 with the PBS complementary to E. coli tRNA3Lys.
Titration of increasing amounts of plasmid pU6EcLys1,2and
pU6EcLysencodingE. coli tRNA 1,2
LysandE. coli tRNA
3
Lysresulted
in the complementation of HIV-1. Thus, the substitution of the
E. coli tRNA3
Lys anticodon to that fortRNA
1,2
Lys did not have an
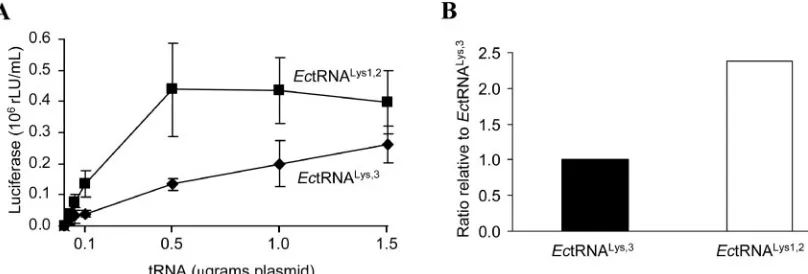
impact on the capacity of this tRNA to complement the rep-lication. In fact, analysis of the complementation levels for all concentrations of plasmid analyzed revealed that the amount of infectious virus recovered was somewhat greater with the plasmid encodingE. coli tRNA1,2Lysthan with the plasmid
encod-ingE. coli tRNA3
Lys (Fig. 5A). Finally, we compared the total
amounts ofE. coli tRNA1,2Lys andE. coli tRNA3Lys found in
trans-fected cells. Using identical amounts of plasmid, we found that intracellular E. coli tRNA3Lys levels were approximately two
times lower than those forE. coli tRNA1,2
Lys (Fig. 5B). If higher
amounts ofE. coli tRNA1,2Lys in the cell are taken into account,
thenE. coli tRNA1,2
Lys and E. coli tRNA
3
Lys complement the
al-tered HIV-1 genome at similar levels, indicating that there is no preference by the HIV-1 provirus fortRNA3
Lys.
Complementation of HIV-1 with E. coli tRNALys
mutants provided intrans.Alteration of nucleotide U35 intRNA3
Lysto A
or G leads to a severe loss of aminoacylation due to poor recognition of the tRNA by the lysyl-tRNA synthetase, while alterations of U34 and U36 have a less severe effect on ami-noacylation (22, 29, 30). Because our earlier studies had found that aminoacylation of the tRNA is important for primer se-lection, we decided to determine whether or not this was also the case forE. coli tRNA3Lys. To address this point, we generated
mutations within the anticodon region ofE. coli tRNA3
Lys that
altered the anticodon from UUU to CUA, to UUA, and to UCA (Fig. 6A). We then compared the levels of complemen-tation of these mutant tRNAs with those for wild-typeE. coli
tRNA3
Lys. Analysis of the complementation for each of the
mu-tant tRNALysrevealed that the level of infectious virus for the
mutant with the anticodon CUA was approximately equal to that of the wild-typeE. coli tRNA3Lys(Fig. 6B). Mutation of the
anticodon UUU to UCA or UUA somewhat compromised the capacities of these mutant tRNAs to complement HIV-1 rep-lication to the level found withE. coli tRNA3
[image:6.585.94.494.67.262.2]Lys (Fig. 6B). We
FIG. 4. Cloverleaf diagrams of lysine tRNA molecules. The anticodon ofE. coli tRNA3
Lys was substituted from UUU to CUU in order to representtRNA1,2Lys. The U34C base change is indicated by an arrow. MammaliantRNA1,2Lysis shown for comparison. Boldface nucleotides correspond to the 3⬘-terminal 18 nucleotides that interact with the PBS.
9646 MCCULLEY AND MORROW J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
next determined whether the levels of complementation were consistent with the levels of aminoacylation of the mutant tRNAs. We found that tRNA mutants with CUA and UUA anticodon alteration were aminoacylated, albeit at low levels, while the mutant with UCA showed no detectable aminoacyl-ation (Fig. 6C). Interestingly, the mutant with the CUA anti-codon had complementation, but not aminoacylation, compa-rable to that of E. coli tRNA3
Lys. All three altered tRNAs
demonstrated considerably less aminoacylation than theE. coli
tRNA3
Lys. The observed results suggest that primer selection is
not entirely dependent on tRNA aminoacylation.
DISCUSSION
In the current study, we have further investigated the mech-anism for the preferential selection oftRNA3Lys as the primer
for HIV-1 reverse transcription. A complementation system which utilizesE. coli tRNA3Lys as the primer for HIV-1 reverse
transcription was developed. The PBS of the HIV-1 proviral genome was modified to be complementary to the 3⬘-terminal 18 nucleotides ofE. coli tRNA3
Lys. The production of infectious
virus was dependent upon the expression of E. coli tRNA3Lys.
However, no preference was found for tRNA3
Lys, yeast
tRNAPhe, ortRNA 1,2
Lys with respect to complementation levels.
Finally, the lack of aminoacylation fortRNA3
Lysanticodon
mu-tants did not correlate to the complementation levels produced by cotransfection of those mutant tRNA plasmids with the HIV-1 proviral plasmid containing the PBS complementary to
E. coli tRNA3
Lys, indicating that interaction with the lysyl-tRNA
synthetase does not entirely explain HIV-1 primer preference. Previous studies from this laboratory and others have ad-dressed the issues of primer preference by using HIV-1 provi-ruses in which the PBS was altered to be complementary to
tRNAs other thantRNA3Lys (4, 17, 34). In each case, it was
found that the resulting virus was unstable and reverted back to utilize tRNA3Lys as the primer, highlighting the fact that
HIV-1 prefers to selecttRNA3
Lys as the primer for replication.
Previous studies have suggested that viral (HIV-1 Gag-Pol) and cellular (lysyl-tRNA synthetase) proteins are important for the preferential selection and use oftRNA3Lys(3, 9, 10). Since it
is difficult to manipulate the endogenous levels oftRNA3
Lys, our
earlier studies used a complementation system by supplying yeast tRNAPhe in trans (14, 15, 37, 38). A limitation of this
yeast tRNAPhe complementation system, though, was the
in-ability to access the viral proteins or lysyl-tRNA synthetase which might be needed for preferential selection oftRNA3Lys.
The use ofE. coli tRNA3Lys circumvents some of these issues
because, as shown in our studies,E. coli tRNA3Lysis
aminoacyl-ated following expression in mammalian cells, indicating the interaction with the mammalian synthetase. Since the antico-don loop ofE. coli tRNA3Lys contains transcriptional
modifica-tions analogous to mammaliantRNA3Lysmodifications and the
tRNAs are alike in sequence, we expected that complementa-tion using this system would be enhanced compared to the
yeast tRNAPhe complementation (2, 28, 32). However, we
found that the absolute complementation levels observed for
E. coli tRNA3Lyswere lower than those for yeast tRNAPhe. The
absolute differences in complementation levels were most likely due to the smaller amounts ofE. coli tRNA3Lys expressed
in the transfected cells, even though both tRNAs were ex-pressed from identical plasmids. To achieve similar levels of complementation, we still needed approximately four times more intracellulartRNA3Lysthan tRNAPhe. Thus, there was no
preferential selection of E. coli tRNA3
Lys, even though this
tRNA could interact with lysyl-tRNA synthetase.
[image:7.585.91.495.69.206.2]One of the unique features of the HIV-1 primer selection is
FIG. 5. Complementation of HXB2(EcPBSLys) with plasmids that encode E. coli tRNA 3
Lys and E. coli tRNA 1,2
Lys. (A) 293HEK cells were cotransfected with 500 ng of proviral plasmids and with tRNA plasmids that were titrated in at the indicated quantities. Dilutions of collected supernatants that were acquired from cotransfections were used to infect the JC53L cell line which contains a luciferase gene under the transcriptional control of the HIV-1 LTR (5, 36). Luciferase activity, in rLU/ml, for complementation of plasmid HXB2(EcPBSLys) with pU6EcLys1,2is represented by closed squares, and that of plasmid HXB2(EcPBSLys) with pU6EcLysis represented by closed diamonds. Background luciferase activity obtained from mock-transfected cultures was subtracted from each sample. Background luciferase activity obtained from HXB2(EcPBSLys) alone was subtracted from all complementation samples of HXB2(EcPBSLys) withE. colitRNALys. The data denote means⫾ standard deviations derived from three independent experiments. (B) Relative ratio ofE. coli tRNA3Lys toE. coli tRNA1,2Lys. 293HEK cells were transfected with 500 ng of pU6EcLysand pU6EcLys1,2. Total RNA was collected, and 15g was loaded per lane (Northern blot). In vitro-transcribed standards ofE. colitRNALyswere loaded at 5 ng, 10 ng, 20 ng, 40 ng, and 80 ng per lane, respectively. The blots were probed forE. colitRNALys and exposed to X-ray film. Areas of the membrane corresponding to the bands on film were excised and counted for radioactivity with a scintillation counter. Known amounts of in vitro-transcribed tRNA were used to generate a standard curve (R2⫽0.99). Using this curve, we found that the amount of tRNA molecules per sample ofE. coli tRNA3Lyswas 0.42 ng per 15g total RNA and that forE. coli tRNA1,2Lyswas 1.00 ng per 15 g total RNA.
on November 8, 2019 by guest
http://jvi.asm.org/
the preference fortRNA3LysovertRNA1,2Lys. This is not due to the
inability of tRNA1,2
Lys to be incorporated into HIV-1 virions,
since previous studies have shown that tRNA1,2Lys is generally
present at levels equal to and sometimes greater than those for
tRNA3Lys (11). If incorporation into the virion was the sole
determinant for primer selection, then one would suspect that an HIV-1 provirus which might utilize tRNA1,2Lys rather than
tRNA3
Lys as the primer for reverse transcription could be
gen-erated. Previous studies have shown that alteration of the pro-viral PBS to be complementary totRNA1,2
Lysdoes not result in a
virus that stably utilizestRNA1,2Lys as a primer for reverse
tran-scription (1, 12, 23). It is only through additional mutations in the U5 region (A-loop or primer activation signal) that the virus can stably utilize tRNA1,2
Lys. However, even under these
conditions, the virus has a replication capacity that is reduced compared to that of the wild-type virus. To further explore this
FIG. 6. Complementation of HIV-1 infectivity withE. coli tRNA3Lysmutants. (A) Cloverleaf structures of mutant tRNALys. The anticodon of E. coli tRNA3
Lyswas mutated from UUU to CUA, to UUA, and to UCA. Boldface nucleotides indicate the anticodon of each tRNA. Base changes are indicated by arrowheads. (B) Complementation of HXB2(EcPBSLys) infectivity with plasmids that encodeE. colitRNALysanticodon mutants. 293HEK cells were cotransfected with 500 ng of proviral plasmids and tRNA plasmids that were titrated in at the indicated quantities. Dilutions of supernatants that were collected from cotransfections were used to infect the JC53L cell line (5, 36). Luciferase activity, in rLU/ml, for complementation of plasmid HXB2(EcPBSLys) with pU6EcLysis represented by closed diamonds, that with pU6EcLysCUAis represented by closed squares, that with pU6EcLysUCAis represented by closed triangles, and that with pU6EcLysUUAis represented by⫻. Background luciferase activity obtained from mock-transfected cultures was subtracted from each sample. Background luciferase activity obtained from HXB2(EcPBSLys) alone was subtracted from all complementation samples of HXB2(EcPBSLys) withE. colitRNALysanticodon mutants. The data denote means ⫾ standard deviations derived from three independent experiments. Note that the standard deviation at 1g for pU6EcLysUUAis slightly shifted for clarity in viewing. (C) Aminoacylation forE. colitRNALysanticodon mutants. The migration of the aminoacylated (AA) and deacylated (DA) samples is shown. Cytoplasmic tRNAs were collected from 293HEK cells that were transfected with pU6EcLys, pU6EcLysCUA, pU6EcLysUUA, and pU6EcLysUCA. All cytoplasmic RNA was isolated under acidic conditions. Lanes 1 and 2 were loaded with cytoplasmic RNA from pU6EcLysUCA transfection; lanes 3 and 4 were loaded with cytoplasmic RNA from pU6EcLysUUAtransfection; lanes 5 and 6 were loaded with cytoplasmic RNA from pU6EcLysCUAtransfection; lanes 7 to 10 were loaded with cytoplasmic RNA from pU6EcLystransfection; and lanes 11 and 12 were loaded with cytoplasmic RNA from mock transfection. All samples were probed forE. colitRNALys. Deacylated controls were prepared by adjustment of samples to basic conditions and incubation for 1 h at 42°C. Deacylated samples are shown in lanes 2, 4, 6, 8, 10, and 12.
9648 MCCULLEY AND MORROW J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
selectivity fortRNA3Lys, we substituted theE. coli tRNA3Lys
anti-codon to correspond to that fortRNA1,2
Lysand then analyzed the
capacity of this tRNA to complement the HIV-1 proviral ge-nome in which the PBS was complementary toE. coli tRNA3
Lys.
While the E. coli tRNA1,2Lys did complement this genome, we
were surprised to find that the levels of complementation fol-lowing normalization for intracellular tRNA levels were simi-lar to those fortRNA3
Lys, indicating that there was no
prefer-ential selection and use ofE. coli tRNA3LysoverE. coli tRNA1,2Lys.
The facts that bothE. coli tRNA1,2
LysandE. coli tRNA
3
Lysinteract
with the lysyl-tRNA synthetase (as confirmed by the analysis of the aminoacylation status of these tRNAs following transfec-tion) and that the virus shows no preference for the E. coli
tRNA3
Lys over E. coli tRNA
1,2
Lys imply that the preference for
mammaliantRNA3Lys overtRNA1,2Lys may be more complex than
the capacity to interact with lysyl-tRNA synthetase.
A further insight into the complexity of primer selection came from our analysis of additionalE. coli tRNA3
Lys mutants
containing substitutions of the anticodon nucleotides. The an-ticodon of the tRNALys is a critical identity element for
syn-thetase recognition. The substitution of nucleotide U35 in
tRNA3
Lys to an A or a G leads to the loss of binding and
aminoacylation by the lysyl-tRNA synthetase (22, 29, 30). Upon testing our mutants in the complementation system, we found that certain mutants had complementation levels close to that forE. coli tRNA3
Lys. The mutant with an anticodon CUA
had complementation levels comparable to those for wild-type
E. coli tRNA3
Lys, while tRNAs with anticodon UCA or UUA
had slightly lower levels of complementation, suggesting that the selective preference fortRNA3
Lysdoes not fully reside in the
unique features of the tRNA molecule. That is, structural features oftRNA3
Lys, such as greater flexibility in the anticodon
region, are probably not entirely responsible for the preferen-tial use oftRNA3
Lysas the primer for HIV-1 reverse
transcrip-tion, although it is possible that structural features oftRNA3Lys
are more important in the processivity of the reverse transcrip-tase (2, 6, 21). A previous study suggests that the tRNA3Lys
anticodon is a key determinant for the incorporation of the primer by HIV-1 and that packaging correlates with amino-acylation (8). However, ourtRNA3
LysUCA mutant, which is not
aminoacylated by the synthetase due to the U35C mutation, complements infectivity of the mutant HIV-1 provirus. These results highlight the possibility that primer selection and pack-aging may be two independent mechanisms. This idea is also supported by our previous studies using a virus that is engi-neered to use tRNAHis. Analysis of the tRNA content of this
virus revealed that it contained amounts/ratios of tRNA1,2,3Lys
similar to those for the wild-type virus (39). Recent studies have confirmed these results by use of viruses which stably use tRNAHis or tRNAMet(35). Collectively, these results suggest
that the selection of the primer used for reverse transcription and the inclusion of the primer in HIV-1 virions might not be linked. Previous studies from our laboratory have suggested that primer selection might be linked with viral translation (14, 15). If this is the case, the availability of certain tRNAs for use in translation could impact their selection as primers for re-verse transcription. The preferential selection oftRNA3Lys as
the primer for HIV-1 reverse transcription might be due to a coordinated process between primer selection and viral trans-lation. How this occurs is unknown, but the use of theE. coli
tRNA3Lys system will facilitate studies to explore this
relation-ship.
ACKNOWLEDGMENTS
We thank the members of the Morrow laboratory for helpful sug-gestions. We thank Adrienne Ellis for preparation of the manuscript. The DNA sequencing was carried out by the UAB CFAR DNA Se-quencing Core (AI 27767). C.D.M. acknowledges helpful suggestions from M.A.R.
A.M. was supported by training grant T32 AI 07493. This research was supported by a grant from the NIH (AI34749).
REFERENCES
1.Abbink, T. E. M., N. Beerens, and B. Berkhout.2004. Forced selection of a human immunodeficiency virus type 1 variant that uses a non-self tRNA primer for reverse transcription: involvement of viral RNA sequences and the reverse transcriptase enzyme. J. Virol.78:10706–10714.
2.Agris, P. F., R. Guenther, P. C. Ingram, M. M. Basti, J. W. Stuart, E. Sochacka, and A. Malkiewicz. 1997. Unconventional structure of tRNA(Lys)SUU anticodon explains tRNA’s role in bacterial and mamma-lian ribosomal frameshifting and primer selection by HIV-1. RNA3:420– 428.
3.Cen, S., H. Javanbakht, S. Kim, K. Shiba, R. C. Craven, A. Rein, K. L. Ewalt, P. Schimmel, K. Musier-Forsyth, and L. Kleiman.2002. Retrovirus-specific packaging of aminoacyl-tRNA synthetases with cognate primer tRNAs. J. Virol.76:13111–13115.
4.Das, A. T., B. Klaver, and B. Berkhout.1995. Reduced replication of human immunodeficiency virus type 1 mutants that use reverse transcription primers other than the naturaltRNA3Lys. J. Virol.69:3090–3097.
5.Derdeyn, C. A., J. M. Decker, J. N. Sfakianos, X. Wu, W. A. O’Brien, L. Ratner, J. C. Kappes, G. M. Shaw, and E. Hunter.2000. Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 modulated by coreceptor specificity defined by the V3 loop of gp120. J. Virol.74:8358– 8367.
6.Isel, C., J. M. Lanchy, S. F. Le Grice, C. Ehresmann, B. Ehresmann, and R. Marquet.1996. Specific initiation and switch to elongation of human immu-nodeficiency virus type 1 reverse transcription require the post-transcrip-tional modifications of primer tRNA3Lys. EMBO J.15:917–924. 7.Isel, C., R. Marquet, G. Keith, C. Ehresmann, and B. Ehresmann.1993.
Modified nucleotides of tRNA(3Lys) modulate primer/template loop-loop interaction in the initiation complex of HIV-1 reverse transcription. J. Biol. Chem.268:25269–25272.
8.Javanbakht, H., S. Cen, K. Musier-Forsyth, and L. Kleiman.2002. Corre-lation between tRNALys3aminoacylation and its incorporation into HIV-1.
J. Biol. Chem.277:17389–17396.
9.Javanbakht, H., R. Halwani, S. Cen, J. Saadatmand, K. Musier-Forsyth, H. Gottlinger, and L. Kleiman.2003. The interaction between HIV-1Gagand human lysyl-tRNA synthetase during viral assembly. J. Biol. Chem.278:
27644–27651.
10.Jiang, M., J. Mak, Y. Huang, and L. Kleiman.1994. Reverse transcriptase is an important factor for the primer tRNA selection in HIV-1. Leukemia
8:S149–S151.
11.Jiang, M., J. Mak, A. Ladha, E. Cohen, M. Klein, B. Rovinski, and L. Kleiman.1993. Identification of tRNAs incorporated into wild-type and mutant human immunodeficiency virus type 1. J. Virol.67:3246–3253. 12.Kang, S.-M., Z. Zhang, and C. D. Morrow.1999. Identification of a human
immunodeficiency virus type 1 that stably uses tRNALys1,2 rather than
tRNALys,3for initiation of reverse transcription. Virology257:95–105.
13.Kelly, N. J., and C. D. Morrow.2005. Structural elements of the tRNA TC loop critical for nucleocytoplasmic transport are important for human im-munodeficiency virus type 1 primer selection. J. Virol.79:6532–6539. 14.Kelly, N. J., and C. D. Morrow.2003. Yeast tRNAPheexpressed in human
cells can be selected by HIV-1 for use as a reverse transcription primer. Virology313:354–363.
15.Kelly, N. J., M. T. Palmer, and C. D. Morrow.2003. Selection of retroviral reverse transcription primer is coordinated with tRNA biogenesis. J. Virol.
77:8695–8701.
16.Kohorchid, A., H. Javannbakht, S. Wise, R. Halwani, M. A. Parniak, M. A. Wainberg, and L. Kleiman.2000. Sequences within Pr160gag-pol affecting the selective packaging of primer tRNALys,3into HIV-1. J. Mol. Biol.299:
17–26.
17.Li, X., J. Mak, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A. Parniak.1994. Effects of alterations of primer-binding site sequences on human immunodeficiency virus type 1 replication. J. Virol.68:6198–6206. 18.Lobo, S. M., and N. Hernandez.1989. A 7 bp mutation converts a human
RNA polymerase II snRNA promoter into an RNA polymerase III pro-moter. Cell58:55–67.
19.Mak, J., M. Jiang, M. A. Wainberg, M.-L. Hammarskjold, D. Rekosh, and L. Kleiman.1994. Role of Pr160gag-polin mediating the selective incorporation
on November 8, 2019 by guest
http://jvi.asm.org/
of tRNALysinto human immunodeficiency virus type 1 particles. J. Virol.
68:2065–2072.
20.Mak, J., and L. Kleiman.1997. Primer tRNAs for reverse transcription. J. Virol.71:8087–8095.
21.Marquet, R., C. Isel, C. Ehresmann, and B. Ehresmann.1995. tRNAs as primer of reverse transcriptases. Biochimie77:113–124.
22.McClain, W. H., K. Foss, R. A. Jenkins, and J. Schneider.1990. Nucleotides that determineEscherichia colitRNAArgand tRNALysacceptor identities
revealed by analyses of mutant opal and amber suppressor tRNAs. Proc. Natl. Acad. Sci. USA87:9260–9264.
23.Moore, K. L., B. R. Kosloff, N. J. Kelly, R. L. Kirkman, L. C. Dupuy, S. McPherson, and C. D. Morrow.2004. HIV type 1 that select tRNAHisor
tRNALys1,2as primers for reverse transcription exhibit different infectivities
in peripheral blood mononuclear cells. AIDS Res. Hum. Retrovir.20:373– 381.
24.Panet, A., and H. Berliner.1978. Binding of tRNA to reverse transcriptase of RNA tumor viruses. J. Virol.26:214–220.
25.Peters, G., and J. E. Dahlberg.1979. RNA-directed DNA synthesis in Moloney murine leukemia virus: interaction between the primer tRNA and the genome RNA. J. Virol.31:398–407.
26.Ratner, L., A. Fisher, L. L. Jagodzinski, H. Mitsuya, R.-S. Liou, R. C. Gallo, and F. Wong-Staal. 1987. Complete nucleotide sequences of functional clones of the AIDS virus. AIDS Res. Hum. Retrovir.3:57–69.
27.Shiba, K., T. Stello, H. Motegi, T. Noda, K. Musier-Forsyth, and P. Schimmel.1997. Human lysyl-tRNA synthetase accepts nucleotide 73 vari-ants and rescuesEscherichia colidouble-defective mutant. J. Biol. Chem.
272:22809–22816.
28.Sprinzl, M., N. Dank, S. Nock, and A. Schon.1991. Compilation of tRNA sequences and sequences of tRNA genes. Nucleic Acids Res.19:2127–2171. 29.Stello, T., M. Hong, and K. Musier-Forsyth.1999. Efficient aminoacylation of tRNALys,3by human lysyl-tRNA synthetase is dependent on covalent
continuity between the acceptor stem and the anticodon domain. Nucleic Acids Res.27:4823–4829.
30.Tamura, K., H. Himeno, H. Asahara, T. Hasegawa, and M. Shimizu.1992.
In vitrostudy ofE. colitRNAArgand tRNALysidentity elements. Nucleic
Acids Res.20:2335–2339.
31.Temin, H. M.1981. Structure, variation and synthesis of retrovirus long terminal repeat. Cell27:1–3.
32.Tisne, C., M. Rigourd, R. Marquet, C. Ehresmann, and F. Dardel.2000. NMR and biochemical characterization of recombinant human tRNALys,3
expressed inEscherichia coli: identification of posttranscriptional nucleotide modifications required for efficient initiation of HIV-1 reverse transcription. RNA6:1403–1412.
33.Wakefield, J. K., H. Rhim, and C. D. Morrow.1994. Minimal sequence requirements of a functional human immunodeficiency virus type 1 primer binding site. J. Virol.68:1605–1614.
34.Wakefield, J. K., A. G. Wolf, and C. D. Morrow.1995. Human immunode-ficiency virus type 1 can use different tRNAs as primers for reverse tran-scription but selectively maintains a primer binding site complementary to
tRNA3Lys. J. Virol.69:6021–6029.
35.Wei, M., S. Cen, M. Niu, F. Guo, and L. Kleiman.2005. Defective replication in human immunodeficiency virus type 1 when non-tRNA3Lysprimers are used
for reverse transcription. J. Virol.79:9081–9087.
36.Wei, X., J. M. Decker, H. M. Liu, Z. Zhang, R. B. Arani, J. M. Kilby, M. S. Saag, X. Wu, G. M. Shaw, and J. C. Kappes.2002. Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob. Agents Chemother.46:1896–1905. 37.Yu, Q., and C. D. Morrow.2000. Essential regions of the tRNA primer
required for HIV-1 infectivity. Nucleic Acids Res.28:4783–4789. 38.Yu, Q., and C. D. Morrow.2001. Identification of critical elements in the
tRNA acceptor stem and T⌿C loop necessary for human immunodeficiency virus type 1 infectivity. J. Virol.75:4902–4906.
39.Zhang, Z., S. M. Kang, A. LeBlanc, S. L. Hajduk, and C. D. Morrow.1996. Nucleotide sequences within the U5 region of the viral RNA genome are the major determinants for an human immunodeficiency virus type 1 to maintain a primer binding site complementary to tRNAHis. Virology226:306–317.
9650 MCCULLEY AND MORROW J. VIROL.