0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.4.2631–2636.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Detection and Partial Characterization of Simian Immunodeficiency
Virus SIVsm Strains from Bush Meat Samples from Rural
Sierra Leone
Cristian Apetrei,
1,2* Michael J. Metzger,
1David Richardson,
3Binhua Ling,
1Paul T. Telfer,
1Patricia Reed,
1David L. Robertson,
3and Preston A. Marx
1,2Microbiology Division, Tulane National Primate Research Center, Covington,1and Tulane Health Sciences Center,
Tulane School of Public Health and Tropical Medicine, Department of Tropical Medicine, New Orleans,2Louisiana, and School of Biological Sciences,
University of Manchester, Manchester, United Kingdom3
Received 16 August 2004/Accepted 22 September 2004
Human immunodeficiency virus type 2 (HIV-2) originated from simian immunodeficiency viruses (SIVs) that
naturally infect sooty mangabeys (SMs; Cercocebus atys). In order to further investigate the relationship
between HIV-2 and SIVsm, the SIV specific to the SM, we characterized seven new SIVsm strains from SMs
sold in Sierra Leone markets as bush meat. Thegag,pol, andenvsequences showed that, while the viruses of
all seven SMs belonged to the SIVsm–HIV-2 lineage, they were highly divergent viruses, in spite of the fact that most of the samples originated from the same geographical region. They clustered in three lineages, two of
which have been previously reported. Two of the new SIVsm strains clustered differently in gag and env
phylogenetic trees, suggesting SIVsm recombination that had occurred in the past. In spite of the fact that our study doubles the number of known SIVsm strains from wild SMs, none of the simian strains were close to the groups in which HIV-2 was epidemic (groups A and B).
Despite an excellent understanding of the ancestry of human immunodeficiency virus type 1 (HIV-1) and HIV-2 in African nonhuman primates (1, 2), nothing is known about the circum-stances that led to the initial emergence and subsequent evo-lution of HIV to become significant human pathogen. The issue of pathogen emergence must be addressed if the poten-tial for emergence of new infectious diseases is going to be understood. The simian counterparts of HIV constitute a highly diverse group of viruses that have been reported to infect more than 35 species of African nonhuman primates throughout sub-Saharan Africa (1). These viruses have a high prevalence in free-living monkeys, with infection rates of up to 60 to 80% in some primate species (10, 28, 35). Some simian immunodeficiency viruses (SIVs) have been reported to grow in vitro on human peripheral blood cells (7, 23), suggesting that they may represent a potential threat of infection for humans. Lentiviruses have very probably been associated with non-human primates for thousands of years, whether or not their most recent common ancestor is ancient (6, 25) or recent (8). Recombination has contributed to the impressive picture of SIV diversity, with several simian species being infected by host-specific SIV types with diverse recombinant-strain ances-try (3, 5, 14, 22, 26, 27, 35). This history, involving host-depen-dent evolution, cross-species transmission events, and frequent recombination, explains the complex evolutionary history of the primate lentiviruses and accounts for the difficulties in analyzing SIV molecular data. The most notable cross-species transmission involved SIVcpz, which represents the ancestors
of HIV-1 (12, 19, 34). Coincidently, in western Africa, a similar event occurred about the same time (within 30 years) and involved transmission of the SIV specific to the sooty manga-bey (SIVsm) to humans, which reflects the ancestry of HIV-2 (10, 20, 21). Therefore, the ancestry is known, but the circum-stances under which epidemic HIV-1 and HIV-2 viruses emerged are completely unknown (2).
The geographic distribution of SIVsm-infected sooty manga-beys (SMs;Cercocebus atys) overlaps the area of HIV-2 ende-micity and includes Liberia, Sierra Leone, Coˆte d’Ivoire, Guin-ea-Bissau, and Guinea (38). SIVsm was apparently transmitted to humans in at least eight instances, forming groups A to H (9, 18, 21). These eight HIV-2 groups (previously called subtypes) are unevenly prevalent: groups A and B are endemic (17), while the other six correspond to single-person infections only (10, 18, 21, 39). Although SIVsm represents the ancestors of all HIV-2 viruses, no SIV counterpart has been found to cluster closely to groups A and B; in marked contrast, putative groups C to H cluster relatively closely to SIVsm strains in the SIVsm and HIV-2 phylogenies (10, 18, 21, 39), suggesting recent crossover infections. In this respect, their extremely low prev-alence and lack of any known association with disease, apart from that of putative group H, may signify that these viruses are less well adapted to their human hosts and less pathogenic (21).
Our group has previously reported the characterization of six SIVsm strains from household pets and feral SMs in Sierra Leone (10). Although four of these viruses (SIVsmSL92a, -d, -e, and -f) originated from animals in the same troop, they were highly divergent and clustered into two different lineages within the HIV-2–SIVsm–SIVmac tree (10). The most signif-icant phylogenetic relationship of these SIVsm isolates was
* Corresponding author. Mailing address: Division of Microbiology and Immunology, Tulane National Primate Research Center, 18703 Three Rivers Rd., Covington, LA 70433. Phone: (985) 871-6518. Fax: (985) 871-6248. E-mail: [email protected].
2631
on November 8, 2019 by guest
http://jvi.asm.org/
that of SIVsmSL92b, from a household pet, which was closely related to HIV-2 subtype E strain PA (10). Although this patient was living in Los Angeles, Calif., he had emigrated from the same geographical region where SIVsmSL92b-in-fected SMs were found. Therefore, these results (10) have offered the most compelling evidence thus far for the simian origin of HIV-2.
In order to evaluate the risk of SIVsm exposure to humans in rural regions of Sierra Leone, we have investigated the prevalence of SIVsm infection in bush meat from monkeys sold in outdoor markets and collected from 1992 to 1993. We have also characterized the phylogenetic relationships of newly discovered SIVsm strains to further investigate the relationship between SIVsm and HIV-2 in this country of HIV-2 emer-gence. No previous bush meat studies had been conducted in Sierra Leone. This is the first study of SIV prevalence and diversity to use bush meat tissues because previous studies relied on blood collected through intracardiac punction (30).
Samples.During a survey carried out in rural Sierra Leone
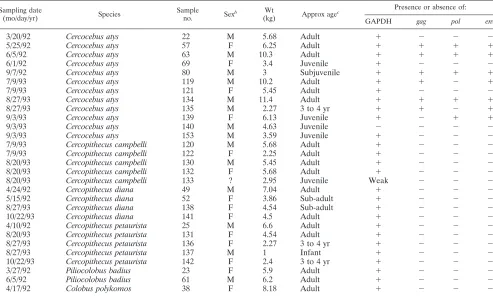
from 1992 to 1993, 29 spleen samples from indigenous monkey species were collected from carcasses in the market. Twelve samples were from SMs, five were from lesser spot-nosed mon-keys (Cercopithecus petaurista), five were from Campbell’s monkeys (Cercopithecus campbelli), four were from diana mon-keys (Cercopithecus diana), two were from red colobus mon-keys (Piliocolobus badius), and one was from a western red colobus monkey (Colobus polykomos) (Table 1).
DNA was isolated from 30 mg of tissue by using a DNeasy tissue kit (QIAGEN, Valencia, Calif.) according to the manu-facturer’s instructions for use in PCR amplification of proviral DNA.
PCRs.Control PCR with primers 5⬘-CCA TCA CCA TCT
TCC AGG AGC GAG-3⬘and 5⬘-CAC AGT CTT CTG GGT GGC AGT GAT-3⬘was done to show that the DNA extraction yielded amplifiable GAPDH (glyceraldehy3-phosphate de-hydrogenase). All but one of the extracted samples gave a positive result, confirming the availability of DNA in the sam-ples used subsequently for SIVsm PCR amplification.
[image:2.585.45.538.81.377.2]Nested PCR was performed to obtain amplified fragments from thegag,env, long terminal repeat (LTR), andpolregions. A 793-bpgagfragment was obtained by a nested-PCR protocol using primers GagA-GagB and GagC-GagF, as described pre-viously (21). Alternatively, primers GF1-GR1 and GF2A-GR3 (10) were used in a nested PCR to generate a 909-bp fragment in thegagregion. These two fragments completely overlapped. Nested primers were used for sequencing. A 438-bp fragment in the gp36envregion was obtained by a previously described nested-PCR protocol with primers EF4-ER1 and EF5A-ER2A (9). A 130-bp LTR fragment was obtained by a nested-PCR protocol using primers LTRA-LTRB and LTRC-LTRD, as described previously (21). A 602-bppolintegrase fragment was obtained by using a slight variation of previously described primers used to amplify divergent SIVs (4, 13–16, 30). Polis4B (5⬘-CCA GCH CAY AAA GGW ATA GGW GGA AA-3⬘)
TABLE 1. Wild nonhuman primates screened for SIVs in bush meat samples by species, age, location, and SIV statusa
Sampling date
(mo/day/yr) Species
Sample
no. Sex
b Wt
(kg) Approx age
c Presence or absence of:
GAPDH gag pol env
3/20/92 Cercocebus atys 22 M 5.68 Adult ⫹ ⫺ ⫺ ⫺
5/25/92 Cercocebus atys 57 F 6.25 Adult ⫹ ⫹ ⫹ ⫹
6/5/92 Cercocebus atys 63 M 10.3 Adult ⫹ ⫹ ⫹ ⫹
6/1/92 Cercocebus atys 69 F 3.4 Juvenile ⫹ ⫺ ⫺ ⫺
9/7/92 Cercocebus atys 80 M 3 Subjuvenile ⫹ ⫹ ⫹ ⫹
7/9/93 Cercocebus atys 119 M 10.2 Adult ⫹ ⫹ ⫺ ⫹
7/9/93 Cercocebus atys 121 F 5.45 Adult ⫹ ⫺ ⫺ ⫺
8/27/93 Cercocebus atys 134 M 11.4 Adult ⫹ ⫹ ⫹ ⫹
8/27/93 Cercocebus atys 135 M 2.27 3 to 4 yr ⫹ ⫹ ⫺ ⫹
9/3/93 Cercocebus atys 139 F 6.13 Juvenile ⫹ ⫺ ⫹ ⫹
9/3/93 Cercocebus atys 140 M 4.63 Juvenile ⫺ ⫺ ⫺ ⫺
9/3/93 Cercocebus atys 153 M 3.59 Juvenile ⫹ ⫺ ⫺ ⫺
7/9/93 Cercopithecus campbelli 120 M 5.68 Adult ⫹ ⫺ ⫺ ⫺
7/9/93 Cercopithecus campbelli 122 F 2.25 Adult ⫹ ⫺ ⫺ ⫺
8/20/93 Cercopithecus campbelli 130 M 5.45 Adult ⫹ ⫺ ⫺ ⫺
8/20/93 Cercopithecus campbelli 132 F 5.68 Adult ⫹ ⫺ ⫺ ⫺
8/20/93 Cercopithecus campbelli 133 ? 2.95 Juvenile Weak ⫺ ⫺ ⫺
4/24/92 Cercopithecus diana 49 M 7.04 Adult ⫹ ⫺ ⫺ ⫺
5/15/92 Cercopithecus diana 52 F 3.86 Sub-adult ⫹ ⫺ ⫺ ⫺
8/27/93 Cercopithecus diana 138 F 4.54 Sub-adult ⫹ ⫺ ⫺ ⫺
10/22/93 Cercopithecus diana 141 F 4.5 Adult ⫹ ⫺ ⫺ ⫺
4/10/92 Cercopithecus petaurista 25 M 6.6 Adult ⫹ ⫺ ⫺ ⫺
8/20/93 Cercopithecus petaurista 131 F 4.54 Adult ⫹ ⫺ ⫺ ⫺
8/27/93 Cercopithecus petaurista 136 F 2.27 3 to 4 yr ⫹ ⫺ ⫺ ⫺
8/27/93 Cercopithecus petaurista 137 M 1 Infant ⫹ ⫺ ⫺ ⫺
10/22/93 Cercopithecus petaurista 142 F 2.4 3 to 4 yr ⫹ ⫺ ⫺ ⫺
3/27/92 Piliocolobus badius 23 F 5.9 Adult ⫹ ⫺ ⫺ ⫺
6/5/92 Piliocolobus badius 61 M 6.2 Adult ⫹ ⫺ ⫺ ⫺
4/17/92 Colobus polykomos 38 F 8.18 Adult ⫹ ⫺ ⫺ ⫺
aAll samples were taken from spleens. The specimen sampled on 9/7/92 was from Gombu, Sierra Leone; the rest were from Bombowo, Sierra Leone. bM, male; F, female.
cAdult,⬎4 years; juvenile, 2 to 4 years; subjuvenile, 1 to 2 years; infant, 0 to 1 year.
on November 8, 2019 by guest
http://jvi.asm.org/
and PolORB (5⬘-ACT GCH CCT TCH CCT TTC CA-3⬘) were used in the first round of amplification, and Polis4B was used again in a seminested reaction with Unipol2B (CCC CTA TTC CTC CCY TTC TTT TAA).
PCR products were purified using a QIAquick gel extraction kit (QIAGEN) and sequenced by direct sequencing and dye terminator methodologies (ABI PRISM BigDye terminator cycle sequencing ready reaction kit with AmpliTaq FS DNA polymerase [PE Biosystems, Warrington, United Kingdom] on an automated sequencer [ABI 373, stretch model; Applied Biosystems]).
Phylogenetic analysis.Thegag,pol, andenvnucleotide
se-quence alignments were obtained from the Los Alamos Na-tional Laboratory HIV sequence database (http://hiv-web.lanl .gov). Newly derived SIVsm sequences were aligned using the CLUSTALW (37) profile alignment option. The resulting alignments were adjusted manually where necessary. Regions of ambiguous alignment and all gap-containing sites were excluded.
Phylogenetic trees were inferred from the nucleotide se-quence alignments by the neighbor-joining method (33) using the HKY85 model of nucleotide substitution (24) imple-mented with PAUP* (36). The reliability of branching order was assessed by performing 1,000 bootstrap replicates, again by using neighbor joining and the HKY85 model. Phylogenetic trees were also inferred by maximum likelihood by using PAUP* with models inferred from the alignment created by use of Modeltest (32). The neighbor-joining tree topology was used as the starting tree in a heuristic search using tree bisec-tion-reconnection branch swapping.
SIV prevalence in bush meat samples from indigenous
mon-keys from Sierra Leone.The nature of the bush meat samples
used in this study precluded a serological screening. Therefore, we screened the samples by PCR. Although this technique is less sensitive than serology in detecting divergent viruses, we used different sets of primers that have been previously shown to be very effective in detecting the SIVs that have been de-scribed to date (4, 10, 13–16, 21, 30). All samples but one were amplified by GAPDH primers, showing the existence of intact DNA in the extracted samples (Table 1).
Nested-PCR amplifications using different sets of SIV-spe-cific primers were positive for 7 out of the 12 SM samples included in this study. In four cases, PCR yieldedgag,pol, and
envsequences; in two cases (SL93-119 and SL93-135), onlygag
and env sequences were obtained; for the remaining one (SL93-139), onlypolandenvsequences were obtained (Table 1). These results show a high prevalence of SIVsm infection in free-living SMs, 63.6% of the tested SMs being positive for SIV. Interestingly, SIVsm prevalence was not significant-ly different between adult and juvenile SMs. Moreover, the youngest SM included in this study was SIVsm infected, pointing to a potential vertical transmission of SIVsm in our study group. However, there is no known connection between SIVsm93SL080 and a potential mother. Therefore, it cannot be concluded that the infection resulted from vertical trans-mission. However, one should note that an SIVsm prevalence of up to 4% was reported in pet SMs in Sierra Leone (10). Since in most cases pet monkeys are captured when they are infants, after their mothers are killed, vertical transmission of
SIV must be occurring and thus is inferred to be a significant mechanism of SIV transmission in the wild.
All other species were negative by PCR using all primer pairs. This result is surprising, since the prevalence of SIV was reported to be high in Central AfricanCercopithecusmonkeys. The animals included in our study group were mainly adults; therefore, one should expect a relatively high prevalence of SIVs. The lack of positive results is probably not due to high divergence, since at least the pol sets of primers were fre-quently used repeatedly and were extremely effective in detect-ing new and divergent SIVs in past studies (13–16, 30). There-fore, these results suggest a lower prevalence inCercopithecus
monkeys in West Africa, at least for the species tested and for a limited number of samples. Analyses of larger groups of monkeys are necessary before conclusions concerning SIV prevalence in these species can be drawn.
Phylogenetic analyses.Analysis of thegag,pol, andenv
se-quences showed that the newly characterized SIVsm strains clustered in the SIVsm–HIV-2 lineage (Fig. 1). Most of the bush meat SIVsm strains clustered with SIVsm strains from wild SMs from Sierra Leone. One strain from an infant SM (SIVsmSL93-080) did not cluster in the previously reported SIVsm SL clusters (10) but was more closely related to SIVsmLib1, a virus originating from the neighboring country of Liberia (Fig. 1). However, this relationship, although ob-served in all phylogenetic trees, was not supported by a high bootstrap value. Forgag, SIVsmSL93-063 and SIVsmSL93-119 clustered relatively closely to the HIV-2 putative group E strain PA, a human virus from a patient who emigrated from the region where these two SIVs were found. To date, SIVsmSL93-063 and SIVsmSL93-119 are the most closely re-lated simian counterparts of an HIV-2 lineage to be identified (Fig. 1A). SIVsmSL93-057 clusters in different phylogenetic positions ingag and pol trees (in which this strain is in the SIVsmSL92b cluster) from those in the envtree, where this virus clustered in the SIVsmSL92a cluster, indicating a possi-ble recombinant-strain history. Note that SIVsmSL93-139 also exhibits the same discordant clustering that SIVsmSL93-057 does in thepolandenvtrees. Another strain, SIVsmSL93-135, also showed different phylogenetic relationships ingagandenv
trees, being grouped with SIVsmSL93a in thegag trees and with SIVsmSL92b in theenvtree (Fig. 1A and C). This differ-ent clustering pattern in differdiffer-ent genomic fragmdiffer-ents is highly suggestive of recombination events that occurred in the wild.
Our study confirms the high prevalence of SIVsm in wild-living SMs. This prevalence is in the same range as those reported for other species of African nonhuman primates which are natural hosts of SIVs: African green monkeys (28) and mandrills (35). Our SIVsm infection rates in Sierra Leone bush meat samples from SMs is in the same range as that reported by our group previously when we used SM plasma samples (10) and more recently feces from SMs from the Tai Forest in Coˆte d’Ivoire (M. L. Santiago, F. Range, F. Bibollet-Ruche, C. Fruteau, R. Peho, J. F. Y. Brookfield, R. Noe, P. M. Sharp, G. M. Shaw, and B. H. Hahn, Abstr. 11th Conf. Ret-rovir. Oppor. Infect. 2004, abstr. 380, 2004). Therefore, our results may accurately reflect the real prevalence of SIVsm in the wild. These prevalence levels are significantly higher than that recently reported following the testing of blood from mon-keys sold in bush meat markets in Cameroon, which was only
on November 8, 2019 by guest
http://jvi.asm.org/
16% (30). This difference may be due to a greater stability of the virus in tissues such as spleen than in clotted blood sampled from dead monkeys in bush meat markets. An alternative explanation is that SIV prevalence varies with nonhuman pri-mate species and geographical location, as already reported in Cameroon (30).
Previous studies have shown that HIV-2 prevalence in Sierra Leone is very low (0.1%) (9) in spite of apparently massive exposure to SIVsm in the markets. Also, cross-species trans-mission to humans is rare in West Africa, as reported to date (9), with only 1 person out of 9,314 tested having SIVsm-like infection (9). This finding raises the question of whether AIDS is really a straightforward zoonosis (2), i.e., a human disease in all cases resulting directly from cross-species SIV transmission events, or is more accurately deemed a transient infection of zoonotic origin which is only occasionally transmissible in the human population. This does not negate the overwhelming evidence that HIV originates from natural cross-species SIV transmission from African primates. Cross-species transmis-sion of SIVsm (and SIVcpz) should be considered the proxi-mate cause of the HIV–AIDS epidemics plaguing the world, while the ultimate cause of HIV and AIDS is related to both viral properties and the evolution of an HIV form readily transmissible in the human population. Cross-species
transmis-sion is fostered by different factors, such as human behavior, environmental changes through deforestation, and industrial-ization during the last century. Viral adaptation to the new host is necessary for the emergence of the new virus: of the eight groups of HIV-2, only the epidemic groups A and B were shown to have pathogenic potential, whereas groups C to G comprise nonepidemic strains that are weakly pathogenic, rep-licate poorly in infected humans, and are found only within the range of SMs and persons who emigrated from western Africa (9, 21). However, the finding of a minor HIV-2 viral lineage (putative group H) that is pathogenic (18) indicates that there is not necessarily a link between pathogenic and epidemic potential. To date, there is no evidence that SIV infection of other African primates can result in HIV or AIDS. However, SIV-infected monkeys may represent a source for new zoo-notic events, given the propensity of SIV strains to recombine. Whereas the body of evidence seems to suggest that direct cross-species transmission may not result in the emergence of a successful virus, the outcome of a cross-species transmission may be successful if the infected individual is already infected with HIV. Such divergent recombinant events have been fre-quent in the evolutionary history of the primate lentiviruses (1, 3, 5, 13, 14, 26, 27, 35).
In spite of the fact that this study doubles the number of
FIG. 1. Phylogenetic relationships, inferred by maximum likelihood, of the newly derived SIVsm strains from Sierra Leone to other SIVsm, SIVmac, and HIV-2 viruses ingag(A),pol(B), andenv(C). After sites that included a gap were removed from the alignments, 348, 582, and 380 nucleotide sites, respectively, remained. Only significant (⬎75%) bootstrap values are shown. The phylogenies are midpoint rooted. New SIVsm strains from Sierra Leone are boxed. Letters on the right sides of the trees mark the HIV-2 groups.
on November 8, 2019 by guest
http://jvi.asm.org/
known SIVsm strains from wild SMs, there is still no evidence of simian counterparts to HIV-2 groups A and B. It is likely that the emergence of epidemic strains of HIV-2 did not occur in Sierra Leone but rather in a different region of the SMs’ natural range. Epidemiologic data point to Coˆte d’Ivoire and Guinea-Bissau. Note that the oldest known strains of HIV-2 groups A and B were found in Coˆte d’Ivoire (11). Until re-cently, PCR fragments from only two SIVsm strains were avail-able from Coˆte d’Ivoire, and they clustered closer to HIV-2 strains than to SIVsm strains from Sierra Leone (31). It was recently reported that SIVsm isolates from the Tai Forest show closer phylogenetic relationships with the epidemic groups of HIV-2, pointing to Coˆte d’Ivoire as their emergence area (San-tiago et al., 11th Conf. Retrovir. Oppor. Infect.). Alternatively, as HIV-2 groups A and B have been estimated to have emerged about 70 years ago (29), it is possible that the SIVsm sources of the HIV-2 lineages in areas of endemicity may have become extinct. More virological surveys of SIVsm diversity in the area of HIV-2 endemicity are needed to investigate this further.
Nucleotide sequence accession numbers.The GenBank
ac-cession numbers for the sequences in this study are AY864786 to AY864798.
This work was supported by grants RO1 AI-44596 and P51 RR000164 from the National Institute of Health.
We thank Tessa Williams, Meredith Hunter, and Nora Dillon for technical assistance and Theresa Secrist for administrative support.
REFERENCES
1.Apetrei, C., D. L. Robertson, and P. A. Marx.2004. The history of SIVs and AIDS: epidemiology, phylogeny and biology of isolates from naturally in-fected non-human primates (NHP) in Africa. Front. Biosci.9:225–254. 2.Apetrei, C., and P. A. Marx.2004. Simian retroviral infections in human
beings. Lancet364:137–138.
3.Bailes, E., F. Gao, F. Bibollet-Ruche, M. Peeters, P. A. Marx, B. H. Hahn, and P. M. Sharp. 2003. Hybrid origin of SIV in chimpanzees. Science 300:1713.
4.Barlow, K. L., A. O. Ajao, and J. P. Clewley.2003. Characterization of a novel simian immunodeficiency virus (SIVmonNG1) genome sequence from a mona monkey (Cercopithecus mona). J. Virol.77:6879–6888.
5.Beer, B. E., B. T. Foley, C. L. Kuiken, Z. Tooze, R. M. Goeken, C. R. Brown, J. Hu, M. St. Claire, B. T. Korber, and V. M. Hirsch.2001. Characterization of novel simian immunodeficiency viruses from red-capped mangabeys from Nigeria (SIVrcmNG409 and -NG411). J. Virol.75:12014–12027. 6.Beer, B. E., E. Bailes, R. Goeken, G. Dapolito, C. Coulibaly, S. G. Norley, R.
Kurth, J.-P. Gautier, A. Gautier-Hion, D. Vallet, P. M. Sharp, and V. M. Hirsch.1999. Simian immunodeficiency virus (SIV) from sun-tailed monkeys (Cercopithecus solatus): evidence for host-dependent evolution of SIV within theC. lhoestisuperspecies. J. Virol.73:7734–7744.
7.Beer, B. E., E. Bailes, P. M. Sharp, and V. M. Hirsch.1999. Diversity and evolution of primate lentiviruses, p. 460–474.InC. L. Kuiken, B. Foley, B. Hahn, B. Korber, F. McCutchan, P. A. Marx, J. W. Mellors, J. I. Mullins, J. Sodroski, and S. Wolinksy (ed.), Human retroviruses and AIDS 1999: a compilation and analysis of nucleic acid and amino acid sequences. Theo-retical Biology and Biophysics Group, Los Alamos National Laboratory, Los Alamos, N. Mex.
8.Charleston, M. A., and D. L. Robertson.2002. Preferential host switching by primate lentiviruses can account for phylogenetic similarity with the primate phylogeny. Syst. Biol.51:528–535.
9.Chen, Z., A. Luckay, D. L. Sodora, P. Telfer, P. Reed, A. Gettie, J. M. Kanu, R. F. Shadek, J. Yee, D. D. Ho, L. Zhang, and P. A. Marx.1997. Human immunodeficiency virus type 2 (HIV-2) seroprevalence and characterization of a distinct HIV-2 genetic subtype from the natural range of simian immu-nodeficiency virus-infected sooty mangabeys. J. Virol.71:3953–3960. 10.Chen, Z., P. Telfer, A. Gettie, P. Reed, L. Zhang, D. D. Ho, and P. A. Marx.
1996. Genetic characterization of new West African simian immunodefi-ciency virus SIVsm: geographic clustering of household-derived SIV strains with human immunodeficiency virus type 2 subtypes and genetically diverse viruses from a single feral sooty mangabey troop. J. Virol.70:3617–3627. 11.Clavel, F., D. Guetard, F. Brun-Vezinet, S. Chamaret, M. A. Rey, M. O.
Santos-Ferreira, A. G. Laurent, C. Dauguet, C. Katlama, C. Rouzioux, D.
Klatzmann, J. L. Champalimaud, and L. Montagnier.1986. Isolation of a new human retrovirus from West African patients with AIDS. Science233: 343–346.
12.Corbet, S., M. C. Mu¨ller-Trutwin, P. Versmisse, S. Delarue, A. Ayouba, J. Lewis, S. Brunak, P. Martin, F. Brun-Vezinet, F. Simon, F. Barre-Sinoussi, and P. Mauclere.2000.envsequences of simian immunodeficiency viruses from chimpanzees in Cameroon are strongly related to those of human immunodeficiency virus group N from the same geographic area. J. Virol. 74:529–534.
13.Courgnaud, V., B. Abela, X. Pourrut, E. Mpoudi-Ngole, S. Loul, E. Dela-porte, and M. Peeters.2003. Identification of a new simian immunodefi-ciency virus lineage with avpugene present among differentCercopithecus
monkeys (C. mona,C. cephus, andC. nictitans) from Cameroon. J. Virol. 77:12523–12534.
14.Courgnaud, V., M. Salemi, X. Pourrut, E. Mpoudi-Ngole, B. Abela, P. Auzel, F. Bibollet-Ruche, B. Hahn, A. M. Vandamme, E. Delaporte, and M. Peeters. 2002. Characterization of a novel simian immunodeficiency virus with avpu
gene from greater spot-nosed monkeys (Cercopithecus nictitans) provides new insights into simian/human immunodeficiency virus phylogeny. J. Virol. 76:8298–8309.
15.Courgnaud, V., P. Formenty, C. Akoua-Koffi, R. Noe, C. Boesch, E. Dela-porte, and M. Peeters.2003. Partial molecular characterization of two simian immunodeficiency viruses (SIV) from African colobids: SIVwrc from West-ern red colobus (Piliocolobus badius) and SIVolc from olive colobus ( Pro-colobus verus). J. Virol.77:744–748.
16.Courgnaud, V., X. Pourrut, F. Bibollet-Ruche, E. Mpoudi-Ngole, A. Bour-geois, E. Delaporte, and M. Peeters.2001. Characterization of a novel simian immunodeficiency virus from guereza colobus monkeys (Colobus guereza) in Cameroon: a new lineage in the nonhuman primate lentivirus family. J. Vi-rol.75:857–866.
17.Damond, F., C. Apetrei, D. L. Robertson, S. Souquiere, A. Lepretre, S. Matheron, J. C. Plantier, F. Brun-Vezinet, and F. Simon.2001. Variability of human immunodeficiency virus type 2 (HIV-2) infection in patients living in France. Virology280:19–30.
18.Damond, F., M. Worobey, P. Campa, I. Farfara, G. Colin, S. Matheron, F. Brun-Ve´zinet, D. L. Robertson, and F. Simon.2004. Identification of a highly divergent HIV-2 and proposal for a change in HIV-2 classification. AIDS Res. Hum. Retrovir.20:666–672.
19.Gao, F., E. Bailes, D. L. Robertson, Y. Chen, C. M. Rodenburg, S. F. Michael, L. B. Cummins, L. O. Arthur, M. Peeters, G. M. Shaw, P. M. Sharp, and B. H. Hahn.1999. Origin of HIV-1 in the chimpanzeePan troglodytes troglodytes. Nature397:436–441.
20.Gao, F., L. Yue, A. T. White, P. G. Pappas, J. Barchue, A. P. Hanson, B. M. Greene, P. M. Sharp, G. M. Shaw, and B. H. Hahn.1992. Human infection by genetically diverse SIVsm-related HIV-2 in West Africa. Nature358:495– 499.
21.Gao, F., L. Yue, D. L. Robertson, S. C. Hill, H. Hui, R. J. Biggar, A. E. Neequaye, T. M. Whelan, D. D. Ho, G. M. Shaw, P. M. Sharp, and B. H. Hahn.1994. Genetic diversity of human immunodeficiency virus type 2: evidence for distinct sequence subtypes with differences in virus biology. J. Virol.68:7433–7447.
22.Georges-Courbot, M. C., C. Y. Lu, M. Makuwa, P. Telfer, R. Onanga, G. Dubreuil, Z. Chen, S. M. Smith, A. Georges, F. Gao, B. H. Hahn, and P. A. Marx. 1998. Natural infection of a household pet red-capped mangabey (Cercocebus torquatus torquatus) with a new simian immunodeficiency virus. J. Virol.72:600–608.
23.Grimm, T. A., B. E. Beer, V. M. Hirsch, and K. A. Clouse.2003. Simian immunodeficiency viruses from multiple lineages infect human macro-phages: Implications for cross-species transmission. J. Acquir. Immune Defic. Syndr.32:362–369.
24.Hasegawa, M., H. Kishino, and T. Yano.1985. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol.22:160– 174.
25.Hirsch, V. M., B. J. Campbell, E. Bailes, R. Goeken, C. Brown, W. R. Elkins, M. Axthelm, M. Murphey-Corb, and P. M. Sharp.1999. Characterization of a novel simian immunodeficiency virus (SIV) from L’Hoest monkeys ( Cer-copithecus l’hoesti): implications for the origin of SIVmnd and other primate lentiviruses. J. Virol.73:1036–1045.
26.Hu, J., W. M. Switzer, B. T. Foley, D. L. Robertson, R. M. Goeken, B. T. Korber, V. M. Hirsch, and B. E. Beer.2003. Characterization and compar-ison of recombinant simian immunodeficiency virus from drill (Mandrillus leucophaeus) and mandrill (Mandrillus sphinx) isolates. J. Virol.77:4867– 4880.
27.Jin, M. J., H. Hui, D. L. Robertson, M. C. Muller, F. Barre-Sinoussi, V. M. Hirsch, J. S. Allan, G. M. Shaw, P. M. Sharp, and B. H. Hahn.1994. Mosaic genome structure of simian immunodeficiency virus from West African green monkeys. EMBO J.13:2935–2947.
28.Jolly, C. J., J. E. Phillips-Conroy, T. R. Turner, S. Broussard, and J. S. Allan. 1996. SIVagm incidence over two decades in a natural population of Ethi-opian grivet monkeys (Cercopithecus aethiops aethiops). J. Med. Primatol. 25:78–83.
29.Lemey, P., O. G. Pybus, B. Wang, N. K. Saksena, M. Salemi, and A.-M.
on November 8, 2019 by guest
http://jvi.asm.org/
Vandamme.2003. Tracing the origin and history of the HIV-2 pandemic. Proc. Natl. Acad. Sci. USA100:6588–6592.
30.Peeters, M., V. Courgnaud, B. Abela, P. Auzel, X. Pourrut, F. Bibollet-Ruche, S. Loul, F. Liegeois, C. Butel, D. Koulagna, E. Mpoudi-Ngole, G. M. Shaw, B. H. Hahn, and E. Delaporte.2002. Risk to human health from a plethora of simian immunodeficiency viruses in primate bushmeat. Emerg. Infect. Dis. 8:451–457.
31.Peeters, M., W. Janssens, K. Fransen, J. Brandful, L. Heyndrickx, K. Koffi, E. Delaporte, P. Piot, G. M. Gershy-Damet, and G. van der Groen.1994. Isolation of simian immunodeficiency viruses from two sooty mangabeys in Cote d’Ivoire: virological and genetic characterization and relationship to other HIV type 2 and SIVsm/mac strains. AIDS Res. Hum. Retrovir.10: 1289–1294.
32.Posada, D., and K. A. Crandall.2001. Selecting the best-fit model of nucle-otide substitution. Syst. Biol.50:580–601.
33.Saitou, N., and M. Nei.1985. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol.4:406–425. 34.Simon, F., P. Mauclere, P. Roques, I. Loussert-Ajaka, M. C. Muller-Trutwin,
S. Saragosti, M. C. Georges-Courbot, F. Barre´-Sinoussi, and F. Brun-Ve
´zi-net.1998. Identification of a new human immunodeficiency virus type 1 distinct from group M and group O. Nat. Med.4:1032–1037.
35.Souquie`re, S., F. Bibollet-Ruche, D. L. Robertson, M. Makuwa, C. Apetrei, R. Onanga, C. Kornfeld, J. C. Plantier, F. Gao, K. Abernethy, L. J. White, W. Karesh, P. Telfer, E. J. Wickings, P. Maucle`re, P. A. Marx, F. Barre ´-Sinoussi, B. H. Hahn, M. C. Mu¨ller-Trutwin, and F. Simon.2001. Wild
Mandrillus sphinxare carriers of two types of lentivirus. J. Virol.75:7086– 7096.
36.Swofford, D.1999. PAUP*: phylogenetic analysis using parsimony (*and other methods), version 4. Sinauer Associates, Sunderland, Mass. 37.Thompson, J. D., D. G. Higgins, and T. J. Gibson.1994. CLUSTAL W:
improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res.22:4673–4680.
38.Wolfheim, J. H.1983. Primates of the world. University of Washington, Seattle.
39.Yamaguki, J. Y., S. G. Devare, and C. A. Brennan.2000. Identification of a new HIV-2 subtype based on phylogenetic analysis of full-length genomic sequence. AIDS Res. Hum. Retrovir.16:925–930.