JOURNAL OF VIROLOGY, June 1978, 615-629 Vol. 26, No. 3 0022-538X/78/0026-0615$02.00/0
Copyright © 1978 AmericanSociety forMicrobiology Printed in U.S.A.
Genome Organization of RNA
Tumor Viruses
I. In Vitro Synthesis of Full-Genome-Length Single-Stranded and
Double-Stranded Viral DNA Transcripts
INDER M. VERMA
Tumor Virology Laboratory, The Salk Institute, San Diego,California92112
Received for publication 29 December 1977
Genome-length complementaryDNA (cDNA) transcripts were synthesized in vitro by using purified virions of avian myeloblastosis virus, Moloney murine
leukemia virus, and clone 124 mouse sarcoma virus. The size of the
genome-length cDNA transcripts was measured on either alkaline sucrose gradients or
alkaline agarosegels. The longest cDNA transcripts synthesized by using avian
myeloblastosis virus, Moloney murine leukemia virus, and clone 124 mouse sarcoma virus were 7, 9, and6 kilobases (kb), respectively. The in vitro system
used was capable of synthesizing double-stranded DNA, but the plus strands
(same polarity as the viral RNA) were only0.5 to 1.2 kb long. Lone Moloney
murine leukemia viruscDNA transcripts were used astemplates to synthesize
thesecondplus strand. Essentially two strategies wereemployed as follows. (i)
The 3' ends of the cDNA transcripts were extended by addition of 50 to 100
dAMPresiduesbyterminaldeoxynucleotidyl transferase. The
(dA)n-tailed
cDNAtranscriptswere usedas templates alongwith anoligomerof dT as primer and
Escherichia coli DNA
polymerase
to synthesize the plus strands. (ii) DNase-digested calf thymus DNA wasused to prime the synthesis of plus strands onlong cDNA withE. coliDNA polymerase I. In both cases, the synthesis of the
plus strands was monitored by increased resistance of the cDNA templates to
single-strand-specific
S,
nuclease. The double-strandedDNA wasfractionatedonneutralsucrosegradients. Analysisofthe double-stranded DNAsynthesized by
using oligo(dT) primer showed the plus strands to be about 5 to 6 kb long, whereastheplusstrandssynthesized by using DNase-digested calfthymus DNA
primerswereonly0.3 to 0.5kblong. Double-strandedDNAsynthesizedby either
method has an average size of 6 x 106 daltons. Double-stranded DNA was also
synthesized by usingcDNAtranscriptsastemplates withouttheadditionof any
primers. In this case, the plus strands were covalently linked to the template strand andwere notrepresentative of the wholeparent strand.
Infection byRNA tumorvirusesrequires the positive strand
(same polarity
asthe viralRNA)
conversion of theirgenetic
material into DNA consists ofsmallDNAtranscripts (13,
38, 47, 48,(2, 43-45, 50, 57), whichcanbe foundinthecell
59).
The conversion of the viralgenetic
material after infection(13,
48,59).
Theviral RNAisfirst into cDNA is carried outby
reversetranscrip-transcribed into
complementary
DNA(cDNA), tase, which is encodedby
the viral RNA(50,57).
which in turn acts as atemplate
to synthesize Viralmutantsdefectiveinthis functionareun-the secondstrand
(17;
H.E.Varmus,
S.Heasley,
abletoestablish infection(24,
46, 49, 53, 56,
57).
H.-J. Kung, H.Opperman,
V. C.Smith,
J. M. Whenpurified
virions arelysed
with nonionicBishop, andP. R.
Shank,
J. Mol.Biol.,
inpress).detergent
and incubated withsubstrates,
DNA In cells infected with either avian or murine issynthesized
that has the nucleic acid sequence retroviruses, thesupercoiled, circular,andlinearcomplexity
of the entire viral genome(8, 11).
forms ofviral DNA have been identified(13, 15,
However,
the average size of the cDNA tran-16, 30). The linear form of the viralDNA,how-scripts
on alkaline sucrosegradients
is about4 ever, represents a majority of the viral DNA to 5S(ca.
500nucleotides)
(11, 44).
Several re-identified in the infectedcell (13, 15,38, 48,59). search groups haverecently
been able to syn-Thenegative strand (DNA strandcomplemen-
thesize small amounts ofnear-genome-length
tarytothe viral RNA) of thelinearviral DNA cDNA
transcripts.
Collett and Faras(6)
ob-appearstobe offull genomelength,whereas the served thatataconcentration of200,um of each615
on November 10, 2019 by guest
http://jvi.asm.org/
616 VERMA J. VIROL.
oftheprecursors, cDNA transcripts larger than single-strand-specific nuclease
SI
(40, 58). The 16S (5 kilobases [kb]) can be synthesized by reaction is stopped when the template is overusing purified virions of Rous sarcoma virus. 70%
SI-resistant,
and double-stranded DNA is Junghans et al. (21), however, showed that the isolated from the unreacted DNA bysedimen-synthesis of long DNA transcripts complemen- tation onneutralsucrosegradients.(ii) The sec-tary to Rous sarcoma virus can be synthesized ond approach is essentially that described by
if optimal detergent concentrations are used. Taylor et al. (41, 42) to utilize DNase-digested Rothenberg and Baltimore (32, 33) have dem- calf thymusDNA asprimers.The minus strand onstrated that at highdeoxynucleosidetriphos- isincubatedwith calfthymusDNAprimers,and
phate precursor concentrationsnearfull-length E. coliDNApolymerase I and the reactionare cDNA transcripts to murine leukemia virus monitored as described above. The
double-(MLV) RNA can be synthesized. Meyers et al. stranded DNA is then isolated on neutral su (29) have reported the synthesis of large cDNA crosegradients. The advantage of the first pro-transcripts byusing the reconstructed system of cedure is that theplusstrand isnearlythe
length
avianmyeloblastosisvirus(AMV) RNA as tem- ofthetemplate strand, whereas the plus strands
plate, oligo(dT) as primer, and purified reverse synthesized by using calfthymusDNAprimers
transcriptase. Recently, Rothenberg et al. (34) areonly 0.5 to 1.0 kblong. However, the second have reported that the in vitro-synthesized approach is more efficient and involves less ma-cDNA using MLV isinfectious. nipulations. The results obtainedby these two
Over the lastyears, wehave been interested approaches and the usefulness of the double-in studydouble-ing theorganization of the genomes of stranded DNA in constructing physical mapsby
avian and murine RNA tumorviruses. We have bacterial restriction endonucleases (31) are
dis-taken the following two approaches: (i) to syn- cussed.
thesize in vitro viral DNA and employ the use
ofrestriction endonucleasestoconstruct a phys- MATERIALS AND METHODS
icalmapof the genome, and (ii) to form heter- Viruses. AMV was obtained from Life Sciences
oduplexes
between thefull-length
cDNA tran- Inc. through the courtesy of the Office of Programscripts ofa given virusandRNA from another Resources and Logistics of the National Cancer Insti-virus and analyze them under anelectron micro- tute.Cloned isolates of M-MLV clone 1 obtained from scope. Both of these approaches require the HungFan and stocks of clone 124 MSV obtained from
availability ofrelatively large amounts of full- Karen Beemon were grown as described previously
genome-length cDNAtranscripts. In this man- (12). The medium was harvested between 24 and 72 h
uscript, wedescribethe conditions of synthesis and, after a low-speed spin (2,000 rpminICE), was oflargecDNAtranscripts fromAMV,acloned stored at -70°C. Virions were
purified
as described isolateofMoloney MLV(M-MLV) andMoloney before (51). The viral protein concentration was de-murine sarcoma virus(M-MSV), clone 124. The terminedas described before (51), and reversetran-invitrosyste
descibedereicapascriptase
activitywasassayedbyemployingexogenousin vitro system described here is capable Of template as described
(51). Purified virions
from clonesynthesizing both strandsofDNA; however, the 1 M-MLV often showed two bands in discontinuous
second strand (plus strand) is synthesized in sucrose
gradients.
Both bands had reverse transcrip-smallfragments.Furthermore, longcDNA tran- tase activity. In general, viruses that showed lessthanscriptsappear tobe entirelysingle stranded. Our 1,000 units ofpolymerase activity (1 unit defined in efforts to cleavefull-genome-length cDNA tran- reference52) permgofviralproteinwerenotusedto
scripts with restriction endonucleases like Hae makecDNA.
III orHpa II, known to cleave single-stranded Unlabeleddeoxyribonucleosidetriphosphates were DNA (3, 18), were not reproducible. To over- obtained from P. L. Biochemicals; 3H-labeled dTTP comethisproblem,wedecidedtosynthesizethe was obtained from Schwarz/Mann; and
P-labeled
comethis
problem,
we decided tosynthesizethedeoxyribonucleoside
triphosphates were obtained second strand ofDNA, using the cDNA strand from ICN, Irvine, Calif. Oligo(dT)-cellulose (type 3) as template. We have used two approaches to wasobtained from CollaborativeResearch, Inc. Ter-synthesize double-stranded viral DNA as fol- minaltransferasewas a gift of John Abelson and for lows. (i) The 3' end of the minus strand is later experiments was obtained from Boehringer extended by 50 to 100 dAMP residues by em- MannheimCorp. Chymotrypsin-treatedE. coli DNAploying the use of terminal deoxynucleotidyl polymerase I was also obtained from Boehringer transferase (terminal transferase [14, 20,
25,
Mannheim. Calf thymus DNA was obtained from35]).
The dA-tailed cDNAtranscripts
are se- Miles Laboratories, Inc.lected by binding to oligo(dT)-cellulose column Synthesis of cDNA. A typical reaction mixture andsedsteplat, wtha oliomerofd as contained50mMTris-hydrochloride (pH 8.3), 10mM
and used as template, with anoligomer of dT as
dithiothreitol,
6 mMmagnesium
acetate,
60mM
NaCl,
primer, andEscherichia coli DNA polymerase 2 mM each of dATP, dCTP, dGTP, dTTP, 0.02 to 1(23).Thereaction ismonitored bydetermining 0.2% Nonidet P-40 for avian viruses and 0.01% for theincreased resistance of the cDNA strand to murine viruses, and 2 to 3 mg of viral protein per 1.0
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 617
ml of the reaction mixture. Reactions werecarried out stranded). The reactionwasterminated eitherby ad-in glass tubes and were flushed with N2 to prevent dition of4
pl
of 0.5 M EDTA or0.25ml ofbinding oxidation of the reducing agent. At the end of the buffer(see below).reaction,portions were withdrawn and the amount of Oligo(dT)-cellulose chromatography. This was acid-precipitable material was determined as de- performed as follows. About 100 mg of oligo(dT)12.18-scribed.Ifthereactionvolumewasless than 2.0 ml, it cellulose (type 3) was equilibrated in binding buffer was chromatographed as described. If the reaction containing 0.01 M Tris-hydrochloride (pH 7.4), 0.4 M volume was less than 2.0ml, it was chromatographed NaCl (chelated), 0.001 M EDTA, and 0.5% sodium on a G-75 Sephadex column to separate unincorpo- dodecyl sulfate. About 50,ugof yRNA was adsorbed ratedradioactivity and substrates. The column buffer tothe column to prevent any nonspecific binding. The contained0.2 MLiCl,0.01MTris-hydrochloride (pH column was washed with 5 volumes of binding buffer, 7.5), and 0.005 M EDTA. Portions from the effluent andthe sample was loaded and allowed to adsorb for werewithdrawn, andpeaksamples with radioactivity 30min. The column was washed with 5 column vol-werecombined andadjustedto afinal concentration umes of binding buffer and eluted withelution buffer of0.4MNaCl, 1% sodiumdodecyl sulfate, and0.01M containing 0.01 M Tris-hydrochloride (pH 7.4) and EDTA.Equal volumes ofphenol saturated with Tris- 0.5%sodium dodecyl sulfate buffer. The material elut-hydrochloride buffer, pH 8.0, were added to it and ingwith elution buffer was brought to a salt concen-shakenvigorously. One volume ofchloroform-isoamyl- tration of 0.2 M LiCl, carrier yRNA was added to a alcoholwasadded, and the mixturewasshaken and finalconcentration of 40
tig/ml,
and nucleicacids were centrifuged in a bench top clinical centrifuge. The precipitated with ethanol.aqueouslayer was reextracted withphenol and chlo- E.coli DNA polymerase I reaction. The reaction roform-isoamylalcohol, followed by extraction twice mixturein0.1 ml contained 20 mM Tris-hydrochloride with chloroform-isoamylalcohol. The aqueous phase (pH 7.5), 4 mM dithiothreitol, 10 mM magnesium was withdrawn, adjusted to 40,ug of yeast RNA as acetate, 60 mM NaCl, 1 mM each of the four deoxy-carrier perml, and nucleic acidswereprecipitated with ribonucleoside triphosphates, and desired amounts of 2.5 volumes of chilled ethanol. Nucleic acids were labeled precursor and template. About 200 pmol of recovered bycentrifugation in 15-ml Corex tubes at (dT),oor(dT)1218 was added as primer when up to 400 15,000xgfor20min.Thepelletswereresuspended in pmol of
(dA).-tailed
cDNA transcripts was used as standard buffer (102 M Tris and 10'- MEDTA, pH template. About 200jg
of DNase-digested calfthymus 7.5). If the reaction volumewas larger than 2.0 ml, DNAprimerwasaddedtoareactioncontaining up to then the nucleic acids were extracted with phenol- 400pmol of cDNA transcripts. The DNase-digested chloroform asdescribed above andprecipitated with calfthymusDNAprimersweremadeasdescribed(41, ethanol. The precipitate was dissolved in small 42). The reaction was carried out at 22 to23°C, and amountsof standardbuffer andchromatographedon incorporation wasmeasured by determining trichlo-G-75 Sephadex column. Restriction endonuclease, roacetic acid-precipitable radioactivity. The double-Bam I-treated '4C-labeled polyoma DNA, and Hae strandedness of the material was measured by SI III-digestedsingle-stranded phage4X174DNA frag- nucleaseasdescribed in thehybridization section. The mentsdid notundergo anychangeinsizeduringthe reaction was stopped by addition of4 1l of 0.5 M incubationperiod for cDNAsynthesis,thussuggesting EDTA, and the unincorporatedradioactivitywas re-that the in vitro system is free of nucleases. movedby chromatographyonG-75Sephadexcolumns Addition of(dA)nresidues. The reaction mixture as described above. Ifno radioactive precursorwas in 0.05ml contained0.2Mpotassium cacodylatebuffer added,the reaction was directly layered on a gradient. (pH 7.4), 1.6 mM ,B-mercaptoethanol, 2 mM cobalt Alkaline sucrose gradients. The DNA sample chloride,0.08mM3H-or32P-labeleddATP,and cDNA wasadjusted to 0.3 MKOH and centrifuged in5to transcript. The mixturewasincubated at37°Cfor5 20%alkalinesucrosegradients pieparedinbuffer con-min, and 1,ulofsamplewaswithdrawntodetermine taining0.7MLiCl,0.3MKOH,and0.005MEDTA, zerotimeincorporation. About8 to 10unitsof enzyme pH 12.4.Aftercentrifugation forappropriatelengths wereused if less than0.2,ig of cDNAtranscriptswas oftime, the gradients were collected bypuncturing addedin thereactionmixture.(The operationaldefi- the bottom of the tube.Polyallomertubes treated with nitionofaunit of terminal transferase obtained from asolution containing 3 ,igofbovine serum albumin the lab of John Abelsonwasthe amountof enzyme perml,10mMTris-hydrochloridebuffer(pH7.4), and requiredto add50dA ordT residues per ,ug of Eco 1mM EDTAtoreduce nonspecific stickingof single-RI-digestedlambda DNAfragment [about0.4pmolof stranded cDNA transcriptswereused for centrifuga-ends] in5 to 15minat 37°C.)The reaction mixture tion. If the cDNA synthesized waslabeled with 32P, wasincubatedat37°Cforperiods of time,depending the amountofradioactivity in each samplewas deter-onthe amountof cDNAtranscriptadded tothe re- mined by counting Cerenkov radiations. For 3H-la-action. The number of nucleotides addedwasdeter- beled DNA samples, portions were withdrawn and minedbythefollowingformulaprovided byJ.Beck- countedinscintillationfluidasdescribed(54). Labeled man of the University ofCalifornia at San Diego: form IIofpolyomaDNA (a gift from M. Vogt) wasnumberofnucleotides=[c/(numberof endsxd)]x usedroutinelyasthestandard forestimatingthe size [b/(a x 106 g)] where a = micrograms of DNA of the cDNAtranscripts.Theopen circles ofpolyoma sample, b = molecular weight of thefragment, c = DNAsedimentedat16S, correspondingto anaverage countsperminuteincorporation,d=specific activity size of 5kb,asdeterminedbytheequationofStudier in counts per minute per picomole, and number of (39).The closed circles sedimentedat18S, represent-ends = 1 (as our long cDNA transcripts are single- ing the sizeofapproximately6.5 kb. All conversions of
on November 10, 2019 by guest
http://jvi.asm.org/
VERMA J. VIROL.
S values tomolecular weightswere doneby using the DNA digested with Hind III and adeno-2 DNA di-equation of Studier (39). Standards were either in- gested with Sma Iwereusedasstandards for deter-cluded in the gradient containing the sampleor runin mining molecular weights. The gelwas dried under parallel gradients. The samplesofinterest were neu- vacuum andsubjectedtoautoradiography. The stan-tralized with acetic acid and precipitated with ethanol. dards werevisualized bystaining the gel with ethidium BothLiCl andpotassiumacetate arehighly soluble in bromide. Details of thegel electrophoresisprocedure
ethanol. will bepublished in the accompanying paper (52).
Neutral sucrose gradients. Sucrose gradients (5 Hybridization.Hybridizationswerecarriedoutin to 20%) wereprepared in buffers containing 50mM buffercontaining0.5MNaCl,0.01M TESbuffer,pH Tris-hydrochloride (pH 7.4), 1 M LiCl, and 5 mM 7.4, 0.001 MEDTA, and 0.1% sodiumdodecyl sulfate. EDTA. Thesampleswerelayered inavolume of 200 All solutions were chelated by chromatography ,lI, andcentrifugationwascarriedoutinpolyallomer throughacolumn containingChelex resin. Incubations tubesintheSW50.1 rotorfor the desired amount of werecarriedoutin 5- to20-/U1 samples at 68°C. The time. Forms I and IIpolyomaDNA(gift of M. Vogt) samples werelayeredwithparaffin oil to prevent loss sedimenting at20and 16S(45)andfull-genome-length ofmaterial. After incubations for appropriate lengths cDNAtranscriptsedimentingat38Swereincluded as oftime, thesampleswerewithdrawnand addedto a standards inparallel gradients. The gradientwasfrac- 10-volumeexcess ofbuffercontaining0.25 M potas-tionated bypuncturing the bottom of the tube, and sium acetate (pH 4.5), 0.01 M ZnSO4, 20
jig
of dena-the distributionofDNA wasdeterminedby measuring turedcalf thymus DNA per ml, and 5 ,ug of native calf radioactivity in various fractions. Fractions with peak thymusDNAperml. The samplesweredividedinto radioactivitysedimenting between18 to21S (or lower twoparts. To one half was added 2HtL of S, nuclease S values if smaller cDNA transcripts were used as (1unit), and it wasincubated for 90 min at 45°C. The templates) were combined and precipitated with other halfwasincubated at 45°C without enzyme.Atethanol. theend of the incubations, residualacid-precipitable
Agarosegel electrophoresis. (i) Alkaline gels. radioactivity wasdetermined. To determine the de-Thehorizontalgel system described by McDonnell et greeof hairpins formed, samples were incubated for1 al. (26)wasused with fewmodifications. Thegelwas to 2 min at 45°C and then immediately assayed for made in buffercontaining 30 mM NaCl and 2 mM sensitivitytoSInuclease.
EDTA, and thesamplewasloadedinthesamebuffer.
Therunning buffer contained 30 mM NaOH and 2 RESULTS
mMEDTA,pH 12.6.Theelectrophoresiswascarried
out in a cold room (4°C) at 2 V/cm for 19 h. The Sze of the cDNA
transcrilpts.Filgure
sA
buffer in the two chamberswas circulatedduring the showsthesedimentation profilesinalkaline su-run tokeep constant pH.At the end ofelectrophoresis, crose gradients of the DNAsynthesized at op-the marker wasvisualized withstainingwithethidium timum conditions by usingpurified
virions of bromide(100,g/ml),and then thegelwasdried under AMV, clone 1 MLV, and clone 124 MSV. The vacuum and autoradiographed. Bam HI-treated pol- majority of cDNA transcripts sediment atabout yomaDNA and HindIII-digestedlambda DNAwere 8 to lOS. Thelargest fraction (fraction I) ofthe boiled in 0.3 N NaOHfor 10minand used as standards. total cDNA transcripts synthesized by using (ii) Neutral gels. Samplesweresubjectedto elec- AMV or clone 1 MLV (shown in Fig. la) were trophoresis in 1.2% agarosegelsmade in Tris-borate resedimentedonalkalinesucrosegradients,
and buffer, pH 8.0, as described (36). A horizontal gel proflesaeon
in Fig.gradion
and apparatus was used, and the gelwas43 cmlong and 4 theirprofiles are shown in Fig.lB.
Fraction Iof mM thick. Samples were loaded in 25t1
of buffer AMV cDNA sediments slightly ahead of iS containingbromophenolblueasdye marker. Lambda marker,representingasize ofabout 7kb,whichFIG. 1. Sizeanalysis of cDNA transcripts onalkalinesucrosegradients and alkaline agarose gels. (A) Analysisof total cDNA. The reaction conditions described in thetextwereemployedtosynthesizecDNA to AMV, clone 1 M-MLV, and clone 124MSV. Either
I"H]dTTP
or[fP]dTTPwas includedas the labeled precursor(specific activity200to2,000cpm/pmol),and thereactionwasincubated for 8 to 16 h at37'C.The cDNAwasprocessedasdescribedin thetextandanalyzedonalkalinesucrosegradients. Small portions from each gradient fraction werecounted to determineradioactivity. Labeled polyoma DNA form II was includedas asize marker.(B)Resedimentation of long cDNA transcripts. The regions of the gradient markedasfractionIin(A)werepooled,neutralized, and precipitated with ethanol. In the panel showing MSV cDNA, regionIIofthegradientwaspooled. The samples were resuspended in alkaline buffer and centrifuged on 5
to20%1valkaline sucrosegradientsin SW41 rotorfor15h at 32,000 rpm at22'C.Thegradients were collected and assayedasdescribed before (50). Labeledpolyoma DNA form II was included as a size marker. (C) Electrophoresisof fractionIof clone1M-MLV cDNA onalkaline agarose gels. 32P-labeledfraction I (about
800 to1,000cpm) obtainedfromalkaline sucrosegradients was neutralized, and the sample was taken in 20 ,ulofbuffer containing30mMNaCl and 2 mM EDTA. Panels (A) and (B) represent fraction I from two different experiments.The molecular weights are determined by using phage A DNA digested with restriction endonuclease Hind III andpolyomaDNAdigested with Bam I. A molecular weight of 3 x 10i shown in the figureisdenatured Hind IIIBfragment ofphage DNA, and 1.5 x ldi molecular weight isdenatured linear formofpolyomaDNA.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26,1978 IN VITRO SYNTHESIS OF VIRAL DNA 619
AMV M-MuLV MSVclone 124
K# fi I
--O 4 _ .jP1 _ _
IEBS 16S 95S 'AS 95 Z.
OWX 106A
B)
IBC lBS --5
4
x~
2 II
0 20 40 0 20 40 0 20 40
FRACTION NUMBER
C
on November 10, 2019 by guest
http://jvi.asm.org/
620 VERMA VIROL.
isvery close to the 7.5-kb subunit sizeofAMV Table 1also showsthe amounts ofvarious frac-RNA (9). On the otherhand, fractionI ofclone tions of cDNAtranscriptssynthesized byAMV 1 MLV cDNA sedimentsatabout 19.5S,repre- and clone 1 M-MLV. Similar datawereobtained senting a size of about 9 kb (19), which is in from cDNA transcripts synthesized using
puri-agreement withthe molecularweightofclone 1 fied virions of MSV (data not
shown).
The MLV RNAof about3 x10'.
The averagesizeof amount offull-genome-length
cDNA transcriptthegenomic RNA of clone 124 MSV is6kb (7, varies from experimentto
experiment,
butit is 19). Resedimentation offraction II of cDNA to alwaysgreater than 5% of the total cDNA syn-clone 124 MSV (Fig. 1B) appears to have an thesized.average size greater than 16S (>5kb).Theviri- Ithas been
reported
thatlarge
amountsofin onsofclone 124 MSV have been shown to con- vivo-synthesizedproviral
DNA are linear (59).tain 1 to7% of the genome ofhelperMLV (7). It Thenegative strand of thelinear
proviral
DNA canbe observed from theprofileof total cDNA appears to be ofgenomiclength,
whereas the synthesized from clone 124 MSV that there is positive strands (having thepolarity
of the ge-somematerialsedimentingahead of18S marker nome) have anaverage size of0.8 to 1.2 kb (59;(6.5kb). Varmus etal., J. Mol.
Biol.,
inpress). We wereFigure IC shows the size of fraction I ofclone interested in
determining
whether the second 1 MLV cDNA made from two different prepa- strandonDNAcanbesynthesized
invitro. The rations (Aand B) onalkalineagarosegels.Frac- long cDNA transcripts appear to be entirelytion I appears to have two distinct bands of single stranded; however, the smaller cDNA averagemolecularweightsof 2.8 x
10"
to 2.9 x transcriptsappeartobequiteSI resistant(Table 106and 3.0 x106
to 3.2 x10".
The ratiosof the 1). If the entire second strand of DNA was twobands differ in differentexperiments.Roth- synthesizedbuthadanaveragelength
of0.5 toenberg et al. (34) have also recently reported 1.2 kb, then on alkaline sucrosegradients most that on alkaline agarose gelsfull-length cDNA of the second strand will sediment in fractions
transcriptsof M-MLV have twosizeclasses,one IV to VI (Fig. 1A). To testthis possibility, we of which is about 600 nucleotides shorter than decided to anneal AMV cDNAfraction V with the other. Ourdata appear to be in agreement fractions I and II. If fractions I and II were with those ofRothenberg etal. (34). transcribedtomakesecond strands of DNA and Nature of the cDNA
transcripts.
Various thetranscriptsweresmall insize,thenannealingfractions of cDNA transcripts obtained from fractions I and IIwith fraction V should render alkaline sucrose gradientswere reannealedand them
SI
resistant. Table 2 shows theresults ofassayed for resistance to
single-strand-specific
suchannealingexperiments. It can be seen that nucleaseSi.
Table 1 registersthedegreeof sin- bothfractions I and II have now becomecom-gle-strandedness and the amounts of various pletely
SI
resistant, suggesting that they were cDNA transcripts synthesized by using AMV transcribed to make the second strand of DNA, andclone1M-MLV. Itcanbe seen thatmaterial but the second strand was small in size. cDNA [image:6.501.68.462.493.588.2]sedimenting largerthan 16S onalkalinesucrose transcripts made in the presence ofactinomycin gradients islargely
SI
sensitive. However, frac- D(100/ig/ml)
areessentially all single stranded tions III to VIbecomeincreasinglySi resistant. (data notshown).TABLE 1. Natureand amountsofvariousfractionsof AMV and clone1M-ML V DNA transcripts"
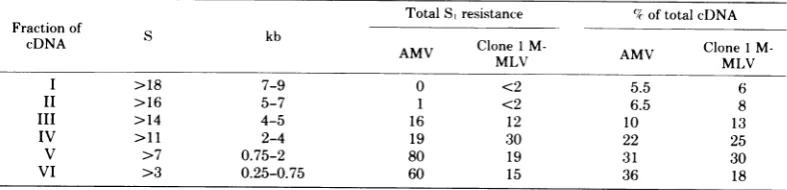
TotalS,resistance %7oftotalcDNA
Fraction of
cDNA S kb AMV CloneIM- AMV Clone 1
M-AV MLV AV MLV
I >18 7-9 0 <2 5.5 6
II >16 5-7 1 <2 6.5 8
III >14 4-5 16 12 10 13
IV >11 2-4 19 30 22 25
V >7 0.75-2 80 19 31 30
VI >3 0.25-0.75 60 15 36 18
"Approximately2to3 ng (400-500 cpm) ofvarious fractions indicated in Fig. IA were pooled, neutralized with acetic acid, andannealed to aCot of 2.4 x 10-2mol s/liter under the conditions described in the text. Sampleswereassayed forS, resistanceasdescribed.The degree of hairpins formed was scored and was found
tobe less than 2 to5%.The values shown in the table are given after subtracting the values of the hairpins formed. The percentage of each fraction present in the total cDNA was determined by dividing the amount of DNAin various fractions marked inFig.IAwith total cDNApresent in the gradient. The values are an average ofatleast 10different experiments.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 621
Addition of
(dA).
residuesatthe3'endof TABLE 3. Selection of(dA),-tailed
cDNAtranscripts
long cDNA transcripts by terminal trans- onoligo(dT)-cellulosecolumn"ferase. Figure 2 shows the kinetics of incorpo- % Bound to oligo(dT)-cel-ration of [32P]dAMP molecules at the 3' end of
Sample
lulose column3H-labeledfull-genome-length cDNA transcripts
amH
esynthesized byusing purified virions of clone 1 'H___ _ _
M-MLV.It can beseen that the incorporation is ['H]cDNA transcript 0
linearwithtime and after incubation for 15 min (>7.5 kb)
anaverage of 50 to 60 nucleotides are incorpo- [ P](dA)r,
['H]cDNA
>90 >90 rated. Unincorporated substrate was removedtranscripts
bychromatographyon a G-75 Sephadex column, "3H-labeled cDNA transcripts were bound to
and the materialelutinginthe void volume was oligo(dT)-cellulose (type 3) column before and after
precipitated with ethanol. The [32P]dA-tailed addition of(dA)n residuesdescribed in the text. The
[3H]cDNA transcripts were selected from the bound
material
was eluted with low-salt buffer asunreacted molecules or
those
[3H]cDNA tran- described.Theamountof thematerial bound to thescripts having
less than 20to 30(dA)n
residues column remained constant even after two cycles of,. ,n ~~bindingandelutionfromthe column.
by chromatography on an
oligo(dT)-cellulose
column. Table3shows that over 90% of the [image:7.501.249.446.91.171.2]'H-and P-labeled material was bound to the col- scripts
contained
at least 30 or more dAMP nsuggesting that a majority of cDNA tran-residues.
The parent template does not bindto umn,suggestingthat a majorityofcDNA tran-oligo(dT)-cellulose
column,
suggesting
that itTABLE 2. Reannealing
of:"p-labeled
AMVDNA in hasnostretches of dA residuesgreater
than 30.fraction Vto 'H-labeledAMV DNA in fractions I
Figures
3a and b show the alkaline sucrose andIi" gradient profiles ofthe [3H]cDNA transcripts before and after addition of 32P-labeled(dA).
Fraction no.
labeled
cDNA residues by terminal transferase. Because the3H-labeled
parentstrand beforeand afteraddi-I 1 tion of 32P-labeled
(dA)n
residues cosediments,I+V 100 we conclude that the [32P]dAMP residues are
II 2
covalently linked to the
[3H]cDNA
transcript.
II +V 98 Furthermore,thedata indicatethattheintegritya3H-labeled fractions I and II (0.6 ng, 1,200cpm) of the template molecules remains unaffected
were annealed with 15 ng of fraction V (low specific during the
reaction.
activity, '2P-labeled, about 200 cpm) in the buffer
described in the text at 68°C to a Cot of 7.2 o10-2
Synthesis
of second strand of DNA by mol s/liter. The percent annealing was assessed by using(dA)n-tailed
cDNAtranscripts
as tem-analyzingwith S, nuclease. Fractions I and II, b plates. The second strand of DNA issynthe-self-annealing,showed only 1 to 2%Ft resistance. sized by using3H-labeled cDNA transcripts
con-taininganaverage of 50 to 60 (dA)nresidues at
o their 3' end as templates,
oligo(dT),21,4
asprimer,W 200 deoxyribonucleoside triphosphates including a
labeledprecursor,and E. coliDNApolymerase
I.
Figure
4shows the rateofincorporation
andz / thepercent
SI
resistance oftemplate at variousw / time intervals during the reaction. It appears
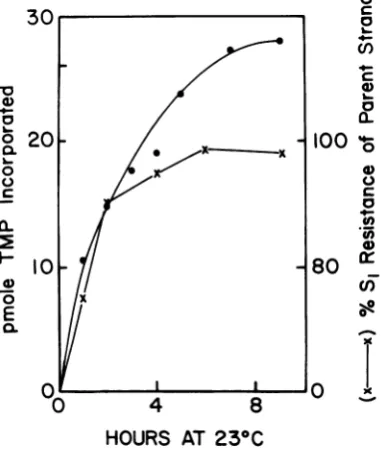
ina
loo / that maximum levels ofincorporation
arereached after 12 h of incubation at room
tem-perature
(230C).
At thispoint,
thetemplate
isox 50-1
over 70%
Si
resistant. The32P-labeled
pluso r < strand ofDNA is over 95%
SI
resistant,suggest-zo
ing that the parent and thedaughter
strands0 5 10 15 20 25 30 remain hydrogen bonded. Both the amount of
MINUTES AT 370C incorporation and the
SI
resistance of the tem-FIG. 2. Kinetics of incorporation of[a-_'2P]dATP plate do not change significantly for the next 10 by terminal deoxynucleotidyl transferase. The reac- to 11 h of incubation. Addition ofenzyme or tion mixturedescribedinthe text was used. About 0.2 substrates after 23 h of incubation stimulatespgofthegenome-lengthcloneIM-MLV cDNAtran-
substre
aftera23oh
Atcomltion
of te scripts obtainedfrom alkaline sucrose gradients (Fig. some more incorporation. At completionof the IB) was used as substrate, and 5-gilportions were reaction, the template is over 80%S,
resistant,withdrawn at various time intervals to determine and the plus strand is about 85% Sl resistant. trichloroacetic acid-insolubleradioactivity. Uponloneperiodsofincubation, theplusstrand
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.45.236.284.374.2] [image:7.501.46.238.452.582.2]622 VERMA J. VIROL.
00 ) 18S 16S
HU5
A0~~~~~~~~~~~~~
51000 _ 1000_ by using (dA)wcDNA and oligo(dT) as
template-U~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~U
500 If0
I ~~IEL FIG4.blKainecticsifty, ndthesi ofplusswothradsofDNA
. t6~~~~~~~~primer.aTheractiondcodtorlnscesdesrbdinsthnetex
K~~~~~~~~~~~~~~~~~~~~~~
.~~~~~~~~~~wremploed.theparentwnstrandh ecn tasdoDNA.labeed
{~~~~~~ i- ando inHicabele dTTPe wasdincuenterationalEoiDAplm
various tm inevl a o i o
E *
5w10 20 30 a
FIG. -
4t
FIG.prnetistran
yndthesis
second strandofDNA.
Tarro
usingfdiAte
o(dTiias
spolyte-where andE.ig
D [image:8.501.276.467.352.579.2]fracion.f3.HSizelanablysi o cDNA transcripts bee _ / _
fracionf3Hlabeed MMLV DNA ransript be- Hlble TPwsinlddi terato
fore and afterthe addition of 32P-labeled (dA) resi- v o m i
dueswas analyzed by centrifugation on alkaline su- 0r ap
crose gradients. 3H-labeled form II ofpolyoma DNA D
the
wasusedasmarkermindparallelgradients. 20 xe 100 o
appears to fall off theparent strand. This is not °prn 8
due to the 5'-3' exonuclease activity associated E
30~~~~~~~
with E. coli DNA polymNerase, as protease-b
treated E. coli DNA pol I (4, 22, 23) gives E
Rfl0)
essentially similar data (data not shown). Thesl80l
net amount of plus strand ofDNA synthesized
be-underthese conditions is about 80% of the input o x amount of the cDNA template. onal s
Synthesis of the second strand ofDNA
by utilising DNase-digested calf thymus o T
DNA primers. Figure 5 showsthe kinetics of o
incorporation and S1 resistance of the template
0)
0by using calf thymus DNA primers and cDNAs
transcript as template. The incorporation is con- HOURS AT
23°C
siderably faster than that observed in the pre- N
viouscase and the reaction essentially reaches by ui Dne-igested cftyus
DNtrim
ers'. .,. ..by..
utilzig
DNase-digested
calfthymus DApiessNAturaion
in.4itu5e.
Trinitor
he0P-labeled
cDNA transcripts were used as template byincreasedSresistance
I ofthetemplateandis in thereaction mixture described in the text, alongterminated when the template is over 95D SA with fHdTTPasthelabeledprecursorandDNase-resistant. If the reaction is not terminated, in- digested calf thymus DNA primers. Portions were
corporation continues,andthetemplateappears withdrawn at various timeintervals and assayed for tobecome less
Si
resistant. incorporationand resistance toSInuclease.on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 623
Isolationand characterization ofdouble- primer and Fig. 6b displays the profile of double-strandedDNA. It has been reported that linear stranded DNA synthesized by calf thymusDNA
double-stranded DNAsynthesized in vivo sedi- primers. It can be seen that a majority of double-ments between 18 and 21S on neutral sucrose stranded DNA synthesized by calf thymus gradients, whereas the parent strand sediments primers sediments as a sharppeak, whereasthe at 36 to38S (13, 59). The double-stranded DNA oligo(dT)-primed double-stranded DNA has a
synthesizedbyeither of the two above methods broader peak. Thetemplate DNA is includedin was fractionated on neutral sucrose gradients, the gradient shown in Fig. 6a and sedimentsat and the material sedimenting between 18 and predictedvaluesof38S (13).
21S was isolated. Figure 6a shows the profile of Analysis of the material sedimenting faster double-stranded DNA synthesized by oligo(dT) than the 18 to21Sregion of thegradient inFig. 6a shows that the parentstrand is full genomic la
length
butthe secondstrand is smaller in size.a) a The
larger
the S value of thefraction,
the 38S 21S 18S smaller the secondstrand (datanotshown)._)
Figure 7 shows the analysis of the size oftheS -_ double-stranded DNA
synthesized by
eitherog method on agarose gels. Figure 7a and b show
N400
thesize of thefull-genome-lengthparent strandO
4
before and after the synthesis of the secondx Effi strand
by
usingcalfthymus
DNAprimers.
The2.5 - r0 double-stranded DNA wascompletely resistant
Z
200T
toS1
nuclease. Ithas been shown before(Var-p,i$l
l mus et al., J. Mol. Biol., in press) thatsingle-taf
l; stranded DNA migrates faster on neutralaga-<{ . \ t l rose
gels
than double-stranded DNA.Double--0*'~~
0 ~ 1 stranded DNA synthesized by utilizing eitherO
20
40 calf thymus DNA primers (Fig. 7b) or byz b) 2 S 18S oligo(dT) priming (data not shown) has a
molec-Z ular weightof 5.9x
106
to 6.1x106,
which is thew
cc s , s
,4observed
sizeof the in vivo-isolated linearDNA.C.) Figure 7c andd, show the electrophoretic
pro-10- files offractional-lengthcDNAtranscripts
(mo-lecularweight about3x
106
to3.6 x 106) before andafter thesynthesisofthe second strand. The distribution of DNA inFig.
7d isquite
broadbecause a broad size class of the parent DNA
5 - wasusedastemplate (Fig.7c). Thus, itappears
that
regardless
of the size oftheparent
cDNA transcripts, both methodsarecapable
of synthe-sizingdouble-stranded viralDNA.The peak fractions ofdouble-stranded DNA
isolated from neutral sucrose
gradients
appear00 tobeover95%S1 resistant.
Although
thedouble-020 40 strandedDNAappears tosedimentat
predicted
FRACTION NUMBER values and has an average molecular weight of
FIG. 6. Fractionationofdouble-stranded DNA on 6 X
106,
the dataobtained byusingeitherneutral neutral sucrosegradients. Uponcompletion ofE.coli sucrose gradients or agarose gels do not show DNApolymeraseIreaction, the sampleswerefrac- that bothstrandsareoffullgenomic length. To tionated on 5 to 20% neutral sucrosegradients as answerthisquestion,wehaveanalyzed the dou-described in thetext. Inparallelgradients,unreacted ble-strandedDNA onalkalinesucrosegradients. parentcDNAtranscriptsandformsIandIIofpoly- Figures 8a and b show the profiles of DNA omaDNAsedimentingat38S(13),20, and 16S(45), synthesized either by oligo(dT) primer or by respectively, were usedassize markers. Thevalues DNase-digested calf thymus DNA primers. It of 21 and18Sshown in thefigurewereobtainedby can be seen that the template DNA(Fig.
8a) using 20 and 16Spolyoma DNA markers asstan-dards. (a)Double-stranded DNAsynthesized byus- sedimentsat about
b9S,
whereas theplusstrand ing oligo(dT) asprimer. (b) Double-stranded DNA of DNA synthesized by oligo(dT) primer sedi-synthesized by using DNase-digested calf thymus mentsheterogeneouslywithanaveragesizeof5DNAprimers. to6 kb. Some fraction ofthe
plus
strandsalsoon November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.44.230.202.539.2]624 VERMA J. VIROL.
reverse
transcriptase
from AMV and E. coliMW X 0o6 DNA polymerase I can make second strandsof
double- - DNA, using the 3' end of the hairpin asprimer
stranded (28, 55). We were interested in determining
DNA whether thelong cDNA
transcripts
of MLV can6.0 - also be
self-primed
tomakeplus
strands of DNA.O
Figure
9a shows the kinetics ofincorporation
3 6 A 1 and SI resistance of|1
large
single-stranded
cDNA5
6
transcripts
used astemplate along
with E. coli* DNA polymerase I. Maximum levels of incor-W
poration
areachieved after 8to 10 h ofincuba-tion, and little
incorporation
is observed if the incubations are carried out for another 10 h. After 8 h ofincubation,
over50% of thetemplatebecomes Si resistant. Further incubation with
0.3
a)
18S 16Sx
0.2 2
10 o jX~
~~\
~0
x xx
x x
[image:10.501.69.258.73.355.2]cn x c
FIG. 7. Sizeanalysis ofDNAbyelectrophoresison = 0.1 |k°
agarosegels. Theparent strand obtainedfrom al- 0 c 1 o
kalinesucrosegradientswasmade double-stranded a 0 0 20 30 40 0
by using DNase-digested calf thymusDNAprimers. XL b) 16S 5S 0
It wasfractionatedonneutralsucrosegradients (see ° X - 6
Fig. 6) and analyzedon1.2%agarosegels. Electro- E x
phoresiswascarriedoutat2.5V/cmfor16h. Panels a. 2.0 I E
a and b show full-genome-length clone I M-MLV X
cDNA transcriptsbefore (a)andafter (b) beingcon- 4 '
vertedtodouble-strandedform. Panelscand d rep- I I
resent 3x Id;to 3.5x 10Plong cDNA transcript of
...l.
clone1M-MLVbefore (c)andafter (d)thesynthesis '
ofthesecond strand. Thesingle-strandedcDNAtran- 1.0 X
scripts migratedaheadofdouble-stranded DNA on r / 2 x
X
these gels. Autoradiographs ofthe driedgels are
shown here. The two molecular weight markers x-x
shown arefragment B of phage lambda DNA di- l /
gestedwithHindIIIandHindlll-digestedfragment I J
AofM-MLV DNA. 0 0 2 0 4
FRACTION NUMBER
sediments along with the template strand. In FIG. 8. Analysis ofdouble-stranded DNA on al-Fig. 8b, the template cDNAtranscript used was kaline sucrose gradients. Fractions of double-about3.5 x
106
daltons. Itseems to maintain its stranded DNA obtainedfrom neutralsucrose gra-integrity during the reaction. However, the plus dients wereanalyzedonalkalinesucrosegradients. strandsstradssynthesizedyntesizd
b cafbycalfthymusthmusDNAprimerDNAprimer(a)lntDouble-strandedd)-ald3PlbldcNDNA made byrncitusing genomen appear tobe no larger than 5S (0.3 to 0.4 kb). length-)n-tailed
'P-labeled
cDNAtranscriptsandSynthesis of double-stranded DNA with-
oligo(dT)12-1m
as primer. (b) Double-stranded DNASynthesis
Ofdouble-stranded DNA with- synthesized byusing4- to5-kb-long32P-labeled
cDNAout primer. It has been shown that various transcripts as template andDNase-digested calf thy-DNAscan form ahairpin and their 3' ends can mus DNA as primers. Labeled form II ofpolyoma then serve as aprimer to initiate DNAsynthesis DNA sedimenting at 18 and 16S was centrifuged in (10, 23). In the case ofglobin cDNA, both the parallelgradients.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.278.462.239.546.2]VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 625
more enzyme or substrate did not enhance S5
300 - ,> resistance. The plus strand remainscompletely
o a) la S resistant during the reaction. If, however,
!i
0O100
0 0 l cDNA transcripts of an average length of 5 kbo _ . zF <,, are used as template, over 70 to 80% of the
0
_ 200 <'' resulting hybrid appears to be double stranded
0
,
(dataQ Q r ^
notshown).
Figure
9b shows the neutraljz
z1-ssucrose
wgradient profiles
of double-stranded2 100
-co5
- 1 DNA synthesized by utilizingapproximately7-kb-long cDNA transcripts as template primer
o / and E. coli DNA
polymerase
I. Atcompletion
E , . _2 of the reaction, the parent strand was over50%
0z
> 10 20 °0SI
resistant.Compared
withtwoprevious
meth-HOURS AT
230C
} ods ofsynthesizing
the second strand ofDNA,
600
E.
the double-stranded DNA sediments with ab)35S 21S 1S' more
heterogeneous profile.
Over20 to 30% ofDNAsediments ahead of 21S,andsomematerial
400F
sediments
at a density ofparentsingle-stranded1 DNA transcript. A large proportion of the
ma-< t t - terial sedimentedbetween14and
21S.
Figure
9cshows the alkaline sucrose gradient profiles of
200 materials
sedimenting slightly
behind(fraction
z / t
28)
andslightly
ahead of 18S(fraction 25)
Dw marker. The data indicate that in both fractions
Nc
. someof theDNAsedimentsataposition
wherec 25 28 the
parent
cDNAtranscript
sediments(ca.
19S).
c) 22
2X
22SES16S However, about 30 to 40%of the materialsedi-200 l mented ahead of
19S,
with aleading edge
atabout 22S.
Thus,
itappearsthat theplus
strandE of DNA is
covalently
linkedtotheparent
strand_ and, thus, sediments faster than parent cDNA
cN4 a on density gradients. Furthermore,this
double-Ls
Cm100- stranded DNAreanneals withzeroorderkinet-o } t
ics,
suggesting
that the two strands formhair-pins. We have, however, so farneverbeen able
tU
to make double-stranded DNA of an averageO . , _ _ size of 18 kb by this procedure. The largest
jo- 0 10 20 30 40 double-stranded DNA sediments on alkaline
su-FRACTION
NUMBER
crose gradients at about 22S(11
to 12 kb). Upon FIG. 9. Kinetics of synthesis and analysis of dou- analysis on agarose gels, this double-stranded ble-stranded DNA synthesized withoutexogenously DNA migrates as a broadband with an average addedprimers. (a) Rate of reaction. The reaction molecular weight of 4.5 x106
to 5 x106
(data conditionsdescribed in thetextwere used exceptthat notshown). Ifthisdouble-strandedDNAis used noprimer was added. The cDNA transcripts usedwere 32P-labeled, and
PH]dTTP
was included as as substrate forrestriction
endonucleases, the labeledprecursor to monitor the reaction. Portions patternof fragments generatedisquite
different
were withdrawn at various time intervals, and a from that obtained by using double-stranded portion wasassayed for trichloroacetic acid-precipi- DNA made by the other two methods. Some tableradioactivity to determine the amount of incor- fragments, however, do appear tohave thesame poration. The other portions of the samples were molecular weights as those obtained from dou-assayed for resistance to
Si
nuclease as describedin ble-stranded DNA made by other methods (un-thetext andthelegend to Fig. 4. (b) Fractionation of published data).the double-stranded DNA on neutral sucrose
gra-dients.Asdescribedinthetextand the legend toFig. DISCUSSION 6.LabeledformsI andII ofpolyoma DNA and
8-kb-long cDNA transcripts were used as standard Genome-length cDNA transcripts were syn-markers for size analysis. (c) Analysis of double- thesized in vitrobyusingpurifiedvirionofAMV,
stranded DNAonalkalinesucrosegradients.
Frac-tions 25 and 28 from neutral sucrose gradients (b) M-MLVand clone124 MSV.The longM-MLV wereanalyzed on alkaline sucrose gradients.Labeled cDNAtranscrpts were then used as templates polyoma DNAfornIIwasincluded as sizemarker tosynthesizethe secondplus strand of the viral in aparallel gradient. DNA. Thispaper describes the characterization
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.501.44.233.74.466.2]626 VERMA J. VIROL.
and conditions for the synthesis of double- unpublished data). Varmus et al. (J. Mol.Biol.,
stranded viral DNA transcripts. in press) have shown that in quail tumor cells
Synthesis ofgenomic-length cDNAtran- acutely infected with avian sarcoma
virus,
plus scripts.Itappears thatasmall butreproducible strands were detected 45minafterinfection. At amount ofgenome-length cDNA transcripts can this stage, the minus strand is only 1 to 2 kb be synthesized in vitro from various avian and long.Thus, contraryto the modelpresentedby murine RNA tumor viruses. The most crucial Haseltine and Baltimore(17),it is not necessary parameters required for the synthesis of long to complete the synthesis of the minus strand cDNA transcripts appear to be very high con- before initiating the synthesis of plus strands. centrations of precursors and an optimal concen- Furthermore, thelargestplus strands observedtration of divalent ion. In addition, it isimpor- by Varmus et al. (J. Mol. Biol., in press) were tant todeterminetheoptimal detergentconcen- about 0.5 to 1.3 kblong, similar to those synthe-trationfor different viruses.Junghans et al. (21) sized in vitro.Thus, the in vitro data ofsynthesis
have shown that full-length cDNA transcripts of double-stranded DNA are very similar to canbesynthesizedfromRous sarcoma virus at those reported for viral DNA isolated in vivo a low substrate concentration if low levels of from either avian sarcoma virus-infected or
detergentareused tolysevirions. We are unable MLV-infected cells.
tosynthesize full-length cDNAtranscripts at a Synthesisofdouble-stranded viralDNA.
concentrationlower than 1 mM each ofthe four Essentially two approaches were employed as precursors. The amount of full-length cDNA described below.
transcriptssynthesizedin a reactionreflects the (i) Extending the 3' end of cDNA tran-amount of intact full-length viral RNA mole- scriptwithdAMP. ThecDNAtranscript was culespresent in thevirion. The data shown here extended by addition of an average of 40 to 60 are obtained from viruses in which the media dAMP residues at the 3'-OH end with terminal were storedat-70°C beforepurifying the viri- transferase (14). The reaction is quite efficient;
ons. Once the virions were purified, however, over 90% of the substrate gets adenylated with
theywere not frozen. In a parallel experiment, at least 30 dAMP residues as judged by its virion 70S RNA was isolated and heatdenatured binding to oligo(dT)-cellulose columns (Table
to generate 35Ssubunits. Less than 10% of the 3). In someexperiments, we have also extended
viral RNA sedimented at regions greater than the 3' end by addition of dTMP residues. Al-35S.Thus, itappears that theefficiencyof tran- though cobalt isused as the divalent ion, it can
scription of genome-length cDNA transcripts be substituted by
Mg2".
The reactionwithter-dependsonthestate ofgenomic RNA. minal transferase appears to be much less effi-Theinvitrosystemiscapableofsynthesizing cient if large amounts of carrier are present.
both strands of the viral DNA. However, the Commercially available terminal transferase is plus strand appears to be synthesized in small generally very dilute, requires long periods of
fragments.Theseresults can beexplained ifone incubation, and, thus, enhancesthe chances of assumesthatasthe complementarycDNAmi- nicking of parent cDNA molecules.
nusstrandelongates, the viral RNAisdigested The
(dA)n-tailed
cDNAtranscript can betran-byRNase H associated with reverse transcrip- scribed intocomplementary strandby using an tase.This RNase Hdigestionleaves3'-OHends oligomer of dT as primer and E. coli DNA onthetemplate,which can now serve asprimer polymerase I. The primer can be as small as to initiate DNA synthesis. The viral reverse (dT)4, and the largest primer we have used is
transcriptase-associated
RNase H is an exonu-(dT)12-18.
In some experiments we have usedcleaseandgeneratesproductswhichhave3'-OH purified AMV reverse transcriptase to replace and5'-phosphate ends (50). Thiskind of mech- E. coli DNA polymerase I. However, our prep-anism will alsosuggest that thesynthesis of the aration ofpurifiedAMV DNApolymerase
intro-plus strand of the DNA will be observed only duced at least one or two nicks per 10,000 nu-after thesynthesis of the minus strand of DNA cleotides. Furthermore, for optimal DNA syn-has been initiated. Furthermore, the synthesis thesis, using purified AMV reversetranscriptase,
ofplusstrands will beinitiatedbefore the syn- high substrateconcentrations are required. One thesis of theminus strand iscompleted. We have of the purposes of making a second strandwas isolated DNA atvariousperiods after incubation tomake it of high specific activity so that we can and determined its degree ofdouble stranded- use it for constructing physical maps of the nessby annealing and assaying for
SI
resistance. genome. The requirement of highconcentrationsOur data suggest that the second strand of DNA of precursors made the use of AMV reverse does not initiate until theminus strand of cDNA transcriptase less attractive. However, the reac-is atleast 1 to 1.5 kb inlength (Fung andVerma, tion with AMV reverse transcriptase can be
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 627
carried out at 37°C and is essentially completed thermore, this is a very useful methodfor us, as
in4 to 5 h. The AMV reverse transcriptase, in we wouldlike to usethis double-stranded DNA addition, is devoid of 3'-5'or 5'-3' exonucleolytic togenerate deletion mutants by employing the activities and, thus, will not digest full-length methodology described for papova viruses (45). double-stranded DNA (50). In the experiments Another advantage ofthis methodturned out to describedhere, the concentration of precursors be theease with which the 5'-terminal restriction
used forE. coli DNA polymerase was quite high fragment of in vitro-synthesized viral DNA
(1 mM each of four dNTP's). However, this is could be located. If
32P-labeled
cDNA transcriptnot necessary, and the reaction can be carried wasextended with[3H]dATP,the fragment
con-out at aconcentration of 10 to 20
AM
of each of taining both 3H and 32P radioactivities mustthe four precursors. We have recently synthe- come from the 5' end of the viralgenome.
sized double-stranded AKR viral DNA, where Theadvantage ofDNase-digestedcalf thymus
the plus strand was labeled with high-specific- DNA as primers is the efficiency of thereaction.
activity 32P-labeled dTTP using 15 ,uM of the The single-stranded cDNA transcript can be
labeled substrate. The reaction with E. coli directly converted to double-stranded DNA
DNApolymerase I is carried out at room tem- without going through the manipulation of ad-perature (22°C) to avoid formation of triplex ditionof
(dA).
residues.The recoveries aregen-structures. Because E. coli DNA polymerase erallyhigher, and the reaction reaches
comple-does containverypotent 5'-3' exonuclease activ- tionmuch faster. However, this methodsuffers ity(23),which utilizesdouble-stranded DNA as from the fact that the secondstrand consists of
substrate,weused chymotrypsin-treatedE.coli small DNAfragments. It is perhaps possible to
DNApolymerase (4, 22, 23), which is devoid of makelongsecond strand by using DNase ligase 5'-3' exonuclease activity. Nosignificant differ- (5, 37) tojointhe fragments.
ence in the data was found, except that with Self-priming of cDNA transcripts. All protease-digestedE. coli DNApolymeraseI the DNA polymerases require primers to initiate rateof reactionwasslower (data not shown). DNA synthesis (1, 23, 44). It was first shown
(ii) Use of DNase-digested calf thymus that T4 DNA canform hairpins andserve as a
DNA primer. This method has
successfully
primertosynthesize thecomplementarystrandbeen used to synthesize representative cDNA (10).Subsequently, it has been shown thatmany
probes from avian and murine RNA tumor vi- in vitro-synthesized cDNA's can self-prime to ruses (41). TheDNase-digestedautoclaved calf make double-stranded DNA (27,28,55).
Appar-thymus DNA is about 4 to 8 nucleotides long ently the3'end of theDNAfoldsback onitself
(41). Themostlikely mode of synthesisof second to reanneal to some complementary part and
strand of DNA is by displacement of newly provides a free 3'-OH end to forn a phospho-synthesized strands.Ifthe reaction isallowedto diester bond with theincoming deoxynucleoside proceedpastwhere the single-strandedtemplate
triphosphate.
Thehairpin loop remains,
how-strand has become SI resistant, the amount of ever,single-stranded.
Itappearsthat the size of incorporation continues and a yield of second thehairpin loop
in case offull-length
cDNA strand greater than that ofinput DNA is ob-transcript
astemplate
is ratherlarge.
Thismaytained. Althoughwehavenottested it directly,
explain why
the maximumSI
resistance seenthe displaced strandscanalsoact astemplates. with over 7-kb cDNA
transcripts
is not moreRecently Varmusetal. (J. Mol.Biol., in press) than 50%.
Furthermore,
theanalysis
of thisco-have also successfully used this approach to
valently
attached double-stranded DNA shows synthesize double-stranded DNA by utilizingge- that thelargest
cDNAtranscript
isonly
1.5 nome-length minusstrandisolatedin vivo, after times the size of the parent molecule. If this infecting quail tumorcells with avian sarcoma double-stranded DNA is treated with Si, the virusesastemplate. parent strand sediments atapproximately
half(iii)
Relative advantages of these two its size(unpublished data). Finally,
if theself-methods. The major advantage of the (dA)n- primed double-stranded DNA is used as sub-tailing method is that verylong transcripts of strate for
digestion
with restrictionendonucle-plus strand of DNA can besynthesized. This is ase, Bam
HI,
a discrete band ofapproximatesofartheonlymethod to obtain biochemically molecular
weight
of 1.25x10'
is observed (datausablequantitiesof lineardouble-stranded viral not
shown).
Thisfragment corresponds
in sizeto DNA where both strands are of near genome theBamICfragment (52)
whichrepresents the length.The other available method is toconvert 3' end of the M-MLVgenomic RNA. Thus, it in vivo-isolated circular viral DNA into linear will appear thatonce thesynthesis
of theplus
form by restriction endonucleases, which have strand
initiates,
itwill
continuetoelongate
untilonecleavagesite in theproviralDNA(60). Fur-
completion.
When cDNA transcripts ofanon November 10, 2019 by guest
http://jvi.asm.org/
628 VERMA J. VIROL.
erage length of4 to 5 kbareusedastemplates, cation of virus-specificmessengerRNA. J. Mol. Biol. over 75% of theresulting hybrid is Si resistant 83:93-117.
and
upo,.alyisoflklinscre
12. Fan, H., andM. Paskind. 1974. Measurements of theand upon analysis ofalkaline sucrose gradient sequence complexity ofclonedMoloneymurine
leuke-has an average size of4 to 8 kb. We are now mia virus60 to 70S RNA:evidence for ahaploid
ge-examining the size of the loop by electron mi- nome.J. Virol. 14:421-429.
croscopy and by restriction endonucleases. 13. Gianni, A. M., D. Smotkin, and R. A. Weinberg.1975.
Thesesd,
wMurine
leukemiavirus:detectionofunintegrateddou-These studies, however, will not distinguish ble-stranded DNA forms ofthe provirus. Proc. Natl. whether the in vivo mode ofsynthesisof second Acad. Sci.U.S.A.72:447-451.
strand of proviralDNA isbyself-primingofthe 14. Goulian, M. 1971. Biosynthesis of DNA. Annu. Rev. minus strand or by use of template RNA as Biochem. 40:855-892.
15. Guntaka,R.V.,B.W. J.Mahy, J. M. Bishop, and H.
primer. E.Varmus. 1975.Ethidium bromide inhibits
appear-ACKNOWLEDGMENTS ance of closed circular viral DNA and integration of
virus-specificDNA induckcells infected by avian sar-I thank LeslieJerominskiand Lisa Dee for thepreparations coma virus. Nature(London) 253:507-511.
of theviruses, and Marianne McKennettandChristine Rob- 16. Guntaka, R. V.,0.C.Richards,P.Shank, H.-J.Kung, ertsforhelpduring this work.IthankM.Vogtforagenerous N.Davidson, E. Fritsch, J. M. Bishop, and H. E.
supplyoflabeledpolyomaDNA markers. IthankJ.Beckmann Varmus. 1976. Covalently closed circular DNA of avian
forvaluabletipsregarding theuseofterminal transferase. I sarcoma virus: purification from nuclei of infected quail
also thank Harold Varmus for communicating his results tumor cells andmeasurement by electron microscopy beforepublication.I thank the members of the Tumor Virol- and gelelectrophoresis. J. Mol. Biol. 106:337-357. ogy Laboratory for constant help and encouragement and 17. Haseltine, W. A., and D. Baltimore. 1976. In vitro
Carolyn Gollerfortypingthemanuscript. replication of RNA tumor viruses. p. 175-213. In D. Thiswork wassupportedbyPublic HealthServiceresearch Baltimore, A. S. Huang and C. Fred Fox, (ed.), ICN-grant CA 16561 and CA 21408 from the National Cancer UCLASymposium on Molecular and Cellular Biology,
Institute andCoreGrantCA 14195 from theNational Cancer Animal Virology, vol. 4. Academic Press Inc., New York. Institute. 18. Horiuchi, K., and N. D. Zinder. 1975. Site specific
LITERATURE CITED cleavageofsingle-strandedDNAbyaHemophilus
re-striction endonuclease. Proc. Natl. Acad. Sci. U.S.A.
1. Baltimore, D., andD.F.Smoler.1971.Primerrequire- 72:2555-2558.
mentsandtemplate specificityof theRNAtumorvirus 19. Hu, S.,N.Davidson,and I.M.Verma.1977.A
hetero-DNA polymerase of RNA tumor viruses. Proc. Natl. duplex studyof the sequencerelationship between the Acad. Sci. U.S.A. 68:1507-1511. Moloneymurine sarcoma virus RNA andMoloney mu-2. Bishop, J. M., and H. E. Varmus. 1975. Molecular rine leukemia virus complementary DNA. Cell
biologyof RNA tumorviruses,p. 3-48.In F. F.Becker 10:469-477.
(ed.), Cancer, vol. 2, Plenum Publishing Corp., New 20. Jackson, D. A., R. H. Symons, and P. Berg. 1972. York. Biochemical method for inserting new genetic infor-3. Blacksley, R. W., and R. D. Wells. 1975. "Single- mationinto DNA simian virus 40:circularSV40 DNA stranded" DNA from OX 174 and M 13 iscleavedby molecules containinglambdaphagegenesand the ga-certain restriction endonucleases. Nature (London) lactoseoperon ofEscherichiacoli. Proc. Natl. Acad. 57:421-423. Sci. U.S.A.69:2904-2909.
4. Brutlag, D., M.R.Atkinson,P.Setlow,and A.Korn- 21. Junghans,R.P.,P.H.Duesberg,andC. A. Knight. berg. 1969. Anactive fragment ofDNA polymerase 1975.Invitrosynthesisoffulllength DNAtranscripts
producedby proteolytic cleavage. Biochem. Biophys. ofRoussarcomavirus RNAby viralDNApolymerase. Res.Commun. 37:982-989. Proc.Natl. Acad.Sci. U.S.A.72:4772-4776.
5. Carbon,J.,T.Shenk,and P.Berg.1975.Construction 22. Klenow, H.,andI.Henningsen. 1970.Selective elimi-invitro ofmutantsof simian virus 40: insertion of a nationofthe exonuclease activityofthe deoxyribonu-poly(dA dT)segmentattheHemophilus parainfluenza cleicacidpolymerasefromEscherichiacoliB by lim-IIrestriction endonuclease cleavage site.J. Mol. Biol. ited proteolysis. Proc. Natl. Acad. Sci. U.S.A. 98:7-15. 65:168-175.
6. Collett,M.S.,and A. J.Faras.1975.In vitro transcrip- 23. Kornberg, A. 1974. DNA Synthesis. W. H. Freeman & tion of DNA from the70SRNA of Rous sarcoma virus: Co., San Francisco.
identification and characterization of various size 24. Linial, M., and W. S. Mason. 1973.Characterization of
classesof DNAtranscripts.J. Virol.16:1220-1228. twoconditional early mutants of Rous sarcoma virus. 7. Dina,D.,K.Beemon,and P.H.Duesberg.1976.The Virology 53:258-273.
30S Moloney sarcoma virus RNA contains leukemia 25. Lobban, P. E., and A. D. Kaiser.Enzymatic end-to-end virusnucleotide sequences. Cell 9:229-309. joiningofDNA molecules. J. Mol. Biol.78:453-471. 8. Duesberg, P.,and E.Canaani.1970.Complementarity 26. McDonnell, M. W., M. N. Simon, and F. W. Studier.
between Rous sarcoma virus (RSV) RNA and the in 1977. Analysis of restriction fragments of T7 DNA and
vitro-synthesized DNA of the virus-associated DNA determinationofmolecular weights byelectrophoresis polymerase. Virology42:783-788. in neutral alkaline gels. J. Mol. Biol.110:119-146. 9.Duesberg, P.,and P. K.Vogt.1973.Gelelectrophoresis 27. McReynolds, L. A., J. J. Monahan, D. W. Bendure, S.
of avianleukosis and sarcoma viral RNA informamide: L. C. Woo, G. V.Paddock,W.Salser, R. E.Moses,
comparison with other viral and cellular RNA species. and B. W.O'Malley. 1977. The ovalbumin gene: inser-J.Virol.12:594-599. tion ofovalbumin gene in chimeric bacterial plasma. J. 10. Englund,P. T. 1971.Theinitial step of in vitro synthesis Biol. Chem.252:1840-1843.
ofdeoxyribonucleic acid by the T4deoxyribonucleic 28. Maniatas, T., S. G. Kee, A. Efstratiadis, and F. C. acidpolymerase.J. Biol.Chem. 246:5684-5687. Kafatos. 1976. Amplification and characterization of a 11.Fan, H.,and D.Baltimore. 1973. RNA metabolism of B-globingenesynthesized in vitro. Cell8:163-182.
murineleukemia virus: detection of virus-specific RNA 29. Meyers, J. C., S.Spiegelman, and D. L. Kacian. 1977. sequences in infected anduninfected cell and identifi- Synthesis of full-length DNA copies of avian
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 IN VITRO SYNTHESIS OF VIRAL DNA 629
blastosisvirus RNA inhigh yields. Proc. Natl. Acad. A.Aaronson.1975. Thermolabile reversetranscriptase Sci.U.S.A. 74:2840-2843. of a mammalian leukemia virus mutant temperature
30. Ringold, G. M., K. R. Yamamoto, P. R. Shank,and H. sensitive in its replication and sarcoma virus helper E.Varmus. 1977. Mouse mammary tumorvirus DNA function.J.Virol.16:1476-1482.
in infected rat cells: characterization ofunintegrated 47. Varmus, H.E., and P. R. Shank. 1976.Unintegrated
forms. Cell 10:19-26. viral DNA is synthesized in the cytoplasm of avian 31. Roberts,R. J. 1976. Restrictionendonucleases.CRC Crit. sarcoma virus-transformed duck cells by viral DNA
Rev. Biochem. 4:123-164. polymerase. J.Virol.18:567-573.
32. Rothenberg,E.,andD.Baltimore.1976.Synthesisof 48. Varmus, H. E., R. V.Guntaka,C.T.Deng,andJ. M. long,representative DNAcopies of themurineRNA Bishop. 1975. Synthesis, structure, and function of tumorvirus genome.J. Virol.17:168-174. avian sarcoma virus-specific DNA in permissive and 33. Rothenberg, E., and D. Baltimore. 1977. Increased non-permissivecells. Cold Spring Harbor Symp.Quant.
length of DNAmade by virions of murine leukemia Biol.39:987-996.
virus at limitingmagnesiumionconcentration.J.Virol. 49. Verma,I. M.1975.Studies on the reversetranscriptase
21:168-178. of RNA tumor viruses. I.Localization ofthermolabile
34. Rothenberg, E.,D.Smotkin, D. Baltimore, and R. A. DNA polymerase and ribo-RNase H activities onone
Weinberg. 1977.In vitrosynthesisof infectious DNA polypeptide.J.Virol. 15:121-126.
of murine leukemia virus. Nature (London) 50. Verma, I. M. 1977. The reverse transcriptase. Biochim.
269:122-126. Biophys.ActaRev. Cancer473:1-38.
35. Roy-choudhary, R.,E.Joy, andR. Wu. 1976.Terminal 51. Verma,I. M., and D.Baltimore. 1973. Purification of
labeling and additionofhomopolymertracts toduplex the RNA-directed DNA polymerasefrom avian mye-DNAfragments byterminal deoxynucleotidyl transfer- loblastosis virusand itsassay with polynucleotide
tem-ase.NucleicAcid Res.3:101-116. plates.MethodsEnzymol.29:125-131.
36. Sharp,P., S.Sugden, and J. Sambrook.1973.Detec- 52. Verma,I. M., and M. A. McKennett. 1978. Genome
tion of two restrictionendonucleaseactivities inHae- organizationof RNAtumorviruses. II.Physical maps mophilus parainfluenzae using analytical agarose- of invitro-synthesized Moloneymurineleukemiavirus
ethidium bromide electrophoresis. Biochemistry 12: double-stranded DNA by restrictionendonucleases.J.
3055-3063. Virol.26:630-645.
37. Shenk, T.E.,C.Rhodes,P. W. J.Rigby, andP.Berg. 53. Verma, I. M., W. S. Mason, S. D. Drost, and D. 1975.Mappingofmutational alterationsinDNAwith Baltimore. 1974. DNApolymeraseactivity from two
S, nuclease: the location ofdeletions, insertionsand temperature-sensitivemutantsof Roussarcomavirus is
temperature-sensitivemutations in SV40. ColdSpring thermolabile.Nature(London) 251:27-31.
HarborSymp.Quant.Biol. 39:61-67. 54. Verma,I. M., G. F.Temple, H. Fan,and D.Baltimore.
38. Smotkin, D., F.K.Yoshimura,and R. E.Weinberg. 1972. In vitro synthesis of DNA complementary to 1976. Infectious,linear,unintegratedDNA ofMoloney rabbit reticulocyte 1OS RNA. Nature (London) New
murine leukemiavirus.J.Virol. 20:621-626. Biol.235:163-167.
39. Studier, F. W. 1965. Sedimentation studiesof the size 55. Verma, I.M., G.F.Temple,H.Fan,and D.Baltimore. andshapeofDNA.J.Mol.Biol. 11:373-393. 1973.Synthesisby reverse transcriptase of DNA com-40. Sutton,W. A. 1971. Acrude nucleasepreparation suitable plementary toglobinmessengerRNA,p. 355-372. In B. for use in DNA reassociation experiments. Biochim. B. Biswas, R. K. Mandal, A. Stevens, and W. Cohen
Biophys.Acta240:522-531. (ed.),SymposiaonControlofTranscription. Calcutta, 41. Taylor, J.M., R. Illmensee,and J. Summers. 1976. India. PlenumPress,New York.
Efficient transcription of RNA into DNA by avian 56. Verma, I. M., H. E. Varmus, and E. Hunter. 1976. sarcoma virus polymerase. Biochim. Biophys. Acta Characterization of "early"temperature-sensitive mu-442:324-330. tants ofavian sarcoma viruses: biological properties,
42. Taylor, J. M., R. Illmensee, L. R. Trusal, and J. thermolabilityof reversetranscriptase in vitro and syn-Summers. 1976.Transcriptionofavian sarcoma virus thesis of viral DNA in infected cells. Virology 74:16-29.
RNA,p. 161-174. In D.Baltimore,A.S.Huang,and C. 57. Vogt,P. K. 1977. Thegeneticsof RNA tumorviruses. In
FredFox(ed.), ICN-UCLA Symposium onMolecular H.Frankel-Conratand R. R.Wagner(ed.),
Comprehen-andCellularBiology,AnimalVirology, vol.4,Academic sivevirology.PlenumPublishingCorp.,NewYork. PressInc.,NewYork. 58. Vogt,V. 1973.Purification andfurther propertiesofsingle
43. Temin, H.M. 1974. On theoriginof RNAtumorviruses. strand-specificnuclease fromAspergillus oryzae. Eur. Annu. Rev.Genet.8:155-177. J.Biochem. 33:192-200.
44. Temin, H.M., andD.Baltimore. 1972.RNA-directed 59. Weinberg, R.W. 1977. Structure of the intermediates
DNAsynthesisof RNA tumor viruses. Adv. Virus Res. leadingtothe integrated provirus. Biochim. Biophys.
17:129-186. ActaRev. Cancer 473:39-55.
45. Tooze, J. (ed). 1973. In Molecular biology of tumor 60. Yoshimura,F.K.,and R. A.Weinberg. 1978.
Restric-viruses, chap. 10-11. ColdSpring Harbor Laboratory, tionendonuclease cleavageof linear andclosed circular ColdSpring Harbor,N. Y. murineleukemia viral DNAs: discovery of a deleted
46. Tronick,S.R.,J.S.Stephenson,I. M.Verma, andS. circular form.Cell,inpress.
![FIG. 2.pgofscriptswithdrawntionbytrichloroaceticIB) terminal Kinetics of incorporation of [a-_'2P]dATP deoxynucleotidyl transferase](https://thumb-us.123doks.com/thumbv2/123dok_us/1528956.105485/7.501.46.238.452.582/fig-pgofscriptswithdrawntionbytrichloroaceticib-terminal-kinetics-incorporation-datp-deoxynucleotidyl-transferase.webp)