Vol.45, No. 1
JOURNALOFVIROLOGY, Jan. 1983, p. 291-298 0022-538X/83/010291-08$02.00/0
CopyrightC1983,American SocietyforMicrobiology
Nucleotide Sequence of the 3' End of MCF 247
Murine
Leukemia Virus
MARGARET KELLY,1 CHRISTIE A.HOLLAND,1 MARIA LI LUNG,t SISIR K. CHATTOPADHYAY,2'3 DOUGLAS R. LOWY,3ANDNANCY H. HOPKINS'*
Centerfor Cancer Research and DepartmentofBiology, Massachusetts Instituteof Technology, Cambridge,
Massachusetts 02139,1 and PediatricOncologyBranch2 andDermatology Branch,3National Cancer
Institute, Bethesda, Maryland20205
Received1 June1982/Accepted27September1982
We isolated DNA clones of MCF 247, aleukemogenic, recombinant type C virus obtained from the thymus ofanAKRmouse.We determined the nucleotide sequence of the virallong terminal repeat(LTR) and the 3' end ofenv, andwe
compared thesequencestocorrespondingsequences of thegenomeof Akvvirus, the putative ecotropic parent of MCF 247. By analogy with Moloney leukemia virus, we identified the amino terminus of Prpl5E, the C-terminal proteolytic
cleavage product ofenv and precursor to mature virion pl5E. In MCF 247 the presumptive Prpl5E is encoded by a 603-nucleotide open reading frame. The
majority of thissequence is identical tothatof Akv. However, arecombination
event near the 3' end of the Prpl5E-coding region introduces nonecotropic sequencesinto MCF247, and these extendtothe3' endthrough the U3 portion of the LTR. TheU3 regions of Akv and MCF 247areabout 83%homologous. The R and U5 regions of the LTR of MCF 247 and Akvareidentical. Large RNase
T1-resistant oligonucleotides analyzed previously innumerous ecotropic and MCF
viral genomeswerelocated within the Akv and MCF 247 DNA sequences. The
resulting precise
T,
oligonucleotide maps of the 3' ends of.MCF
viral genomesreveal that the biologically defined, leukemogenic class I MCFs isolated from thymic neoplasms of inbred mice all share thesequencepatternseeninMCF247, arepresentative of thisgroup;theypossessrecombinantPrpl5Egenesand derive U3 from theirnonecotropic parents.
Mink cell
focus-forming
(MCF)typeC virusesthatdiffer in
oncogenicity
can bedistinguished
from one another and from their nononcogenic
ecotropic progenitors by
RNaseTl-resistant
oli-gonucleotides
located in U3 and the 3'portion
ofenv(4, 16, 23,24, 33). To determine the
precise
structural differences in this region of the
genomebetween
leukemogenic
MCF 247 and its non-leukemogenic ecotropic progenitor, Akvvi-rus, we determined the nucleotide sequence of
the3' 1.3 kilobases ofaDNAclone of MCF 247.
This region includes sequences encoding the
long terminal repeat (LTR),the primer binding
site forplus-strand DNA synthesis,Prp15E, the
C-terminal
proteolytic cleavage
product ofenvandprecursor to mature virion
pl5E
(9, 19, 27,38), and the C terminus of
gp7O.
The LTR ofAkv had been sequenced previously by Van
Beveren etal. (36), and Lenzet al. (21)and V.
Corbin and W. Herr
(unpublished results)
havesequenced the env gene of Akv. The DNA
sequences reveal the precise location of large
tPresent address:UniversityofHongKong,HongKong.
RNase T1-resistant oligonucleotides analyzed
previously
inviral RNAfingerprints
(23, 24, 31,32). Together, the T1
oligonucleotide
data andthe DNAsequencesreveal that the
amino-termi-nalhalf of
pl5E
and the Cterminus ofPrp15E,
as well as U3, distinguish biologically distinct
MCF and
ecotropic
viruses of inbred mice.Previous
biological
and biochemical studieshave emphasized differences in
gp7O
betweentheseviruses (7, 16).Together, the studies
indi-cate that particular sequences of
gp7O, pl5E,
and theLTR may allbe important in determining
the leukemogenic phenotype of MCF type C
virusesof inbred mice.
MATERIALSANDMETHODS
Molecular cloning of MCF 247 virus. To obtain circular DNA intermediates of MCF 247, Mus dunii cellswereinfected withMCF 247 virus obtained from
chronically infected cells (2). Eighteen hours after
infection, DNA wasextractedbythe Hirtprocedure (18)asdescribedpreviously (30). Toenrichfor geno-mic-lengthmolecules,DNAfrom the Hirtsupernatant
wascleaved withHindIlI andelectrophoresedon a1%
291
on November 10, 2019 by guest
http://jvi.asm.org/
agarose gel. DNA migrating between 6.7 and 9.5 kilobases was isolated (2). A 2- to 20-fold excess of enriched DNAwasligatedtoHindIII-cleaved pBR322
at a DNA concentration of 25 to 50 ,g/ml. After ligation, 100 to 500 ngof DNA was usedtotransform the RR-1 strain of Escherichia coli(5). Colonies that grew on agar plates containing ampicillin were screenedfor viral DNA sequences by filter hybridiza-tion (14) with 32P-labeled cDNA prepared from 70S MCF 247 RNAas aprobe (2). Singlecolonies which hybridized to cDNAwereamplified inliquidculture, andplasmid DNAwasisolated(3).Plasmid DNAwas
analyzed by restriction endonuclease cleavage with KpnI, SmaI, and EcoRI. Plasmid inserts which
con-tained restriction sites similartothoseof MCF 247 (1) werepurified frompBR322 afterHindlIl digestion of plasmid DNA. From 1 to 5 1Lg of each insert was
ligated and transfectedby the calciumphosphate pre-cipitation method (12) ontoNIH 3T3, MCT,orSC-1 cells (15). Cellswere passagedfor 2to6weeks,and then culture fluidswereassayed forreverse
transcrip-taseactivity.
Sequence determination. The pMCF247-lb DNA clone was cleaved with restriction endonucleases
(from New England Biolabs or Bethesda Research Laboratories), andfragmentswerelabeledattheir 5' ends with[-y-32P]ATP(fromTheRadiochemical Cen-tre, Amersham, England) and polynucleotide kinase
(from BoehringerMannheimCorp.)afterelution from agarose preparativegels. Fragmentswere recleaved,
and their sequencewasdeterminedby usingthe chem-ical modification procedures of Maxam and Gilbert (25). Reactions were run on 15,8, or 6%
polyacryl-amide-8 Mureagels, 0.4mmthick. Sequenceswere
compiledandanalyzedwith the computer programsof
Gingeras etal.(11).
RESULTS
Characterization of molecular clones of MCF
247 virus.
Molecular
clones of MCF 247 wereprepared
by
cleavage
ofcircular viral DNA atthe
unique
HindIlI
site and insertion intopBR322 or other
cloning
vectors as describedabove. At the time the DNA
sequencing
studieswere
initiated,
eight
molecular clones had beenobtainedin this way. Of
these,
six hadrestric-kb i
Rl
tion patterns similar to those determined by Chattopadhyay et al. (1, 2) for proviral DNA of
MCF247-infected cells,yetnone was infectious
when transfected into appropriate mouse cells.
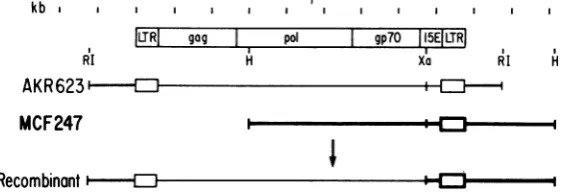
To determine whether the 3' end of one of
theseclones was biologically functional, we
gen-erateda recombinant in vitro between an MCF
clone, designated pMCF247-lb, and an
infec-tious DNAclone of integratedAkv murine
leu-kemia virus (MuLV),AKRMuLV 623 (22).The
MCF DNAwasdigested with XbaIandHindIII,
andthe fragment encodingthe 3' end of the viral
genome was isolated and ligated to an EcoRI-XbaI fragment containing the 5' remainder of the genome from AKR MuLV 623 (Fig. 1). When
transfected, this DNA was infectious, yielding
ecotropic, XC plaque-forming virus, whose
RNase T1 fingerprint was consistent with its
genomic structure and with previously
deter-mined
T,
oligonucleotidemaps(data not shown;32).Therefore,wewereconfidentof the
biologi-cal integrity of pMCF 247-lb intheregionfrom
XbaItothe3'endofthe MCF 247 genome, and we proceeded to sequence this DNA.
Recently, Holland (unpublished data)
ob-tained
infectious
DNAclones ofMCF 247, andthe sequenceof the region from XbaI to the 5'
endof thepl5E-codidg region,presented below,
was determinedagain with oneof these clones.
One nucleotide
difference
(outof173bases)wasfound between thetwoclones within thisregion.
DNA sequencing. All DNA sequences were
determined by the Maxam and Gilbert
proce-dure (25). The strategyfor generating
appropri-ate
fragments
for sequencing the 3' end of theMCF 247 DNA clone is shown in Fig. 2. The
nucleotidesequencesobtainedare shown in Fig.
3 and 4, where the Prpl5E- and LTR-coding
regions of MCF 247 are compared with the
corresponding
sequences of Akv obtained byVan Beverenetal. (36)(LTR), Lenz et al. (21)
(Prpl5E), and V. Corbin andW. Herr(Prpl5E;
unpublished results).
II
ILIRI
gag I pol gp7 |15EILTRI
H Xa RI H
AKR623'
C1 tCMCF247
Recombinant
iFIG. 1. Schematic representation of DNA clones of MCF 247 and Akv and arecombinant between them
constructed in vitro. AKR 623wascloned fromintegratedAkvproviruspresent in infected NIH/3T3 cells after
EcoRI cleavage of the cellular DNA. This viral clone therefore containsmouse cellular sequences(22). The molecular clone of MCF 247wasobtainedby HindIIIcleavageofacircular moleculeand ispermutedasshown. Restriction enzyme sites usedto constructtherecombinant betweenAkvand MCF247areindicatedatthetop of thefigure. Abbreviations: RI,EcoRI; H,Hindlll;Xa,XbaI.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.489.109.393.525.623.2]VOL.45,1983 NUCLEOTIDE SEQUENCE OF 3' END OF MCF 247 293
.4gp7O
I
Prp15E1+
I
U3I RI
U5 ~-1022 -420 -382 +142
GOx
Inverted TATAAAA Inverted8 (35)1 9 18 47 33 101 26Repet (14) 108 \106 CAATAAA Repet
ag
p
mm_
a
a
U*
i
T H X B 0 RHIRHSo PRI M SmnK
-1100 -984 -854 -797 -694 -528 -486 -446 -344 -304 -206 +31 +37
- --- - --- ---
4-T=
Taql
B:=BglII So=Sau3A M=MspIH= HinfI D:= Ddel P: Psi! Sm:Smul
[image:3.489.53.444.70.224.2]X= Xbul RII=EcoRII RI =EcoRI K=
Kpnl
FIG. 2. StrategyforsequencingDNAencodingthe 3' end of the MCF247 genome. Arrowsindicate regions
of the DNA thatweresequencedfrom the restriction sitecorrespondingtothetail of thearrow. Symbols: (4----)
regions sequencedfrom minus-strandDNA;(--*)regions sequencedfromplus-strand DNA; (+)originof
plus-strandDNAsynthesis; (U)
T,
oligonucleotides analyzedpreviouslyinfingerprintsof MCF247genomic RNA;(LI)positionsoftwo
T,
oligonucleotidesthatarepresent inAkv,butnotinMCF247.Theboundaries ofPrpl5Eand the U3, R, and U5 regions were identified as described in the text. The numbers above the
T,
oligonucleotidesarethosegiveninpublished fingerprints(23, 31,32).G1x,
T1oligonucleotide8 hasbeen showntobegeneticallylinkedtothe presenceof thegp7Oantigen
G1x.
The first base pair inR is consideredas 1.G:
rC~ ~ ~ ~ ~ I
C~ ~ ~ ~ ~ ~ A
jC CC,7 C CAT 'TCCGAA:AA,
A ~~~~~T.A: :-''L%-TL-`T GA" -- - TTGTA' A'
Thr As
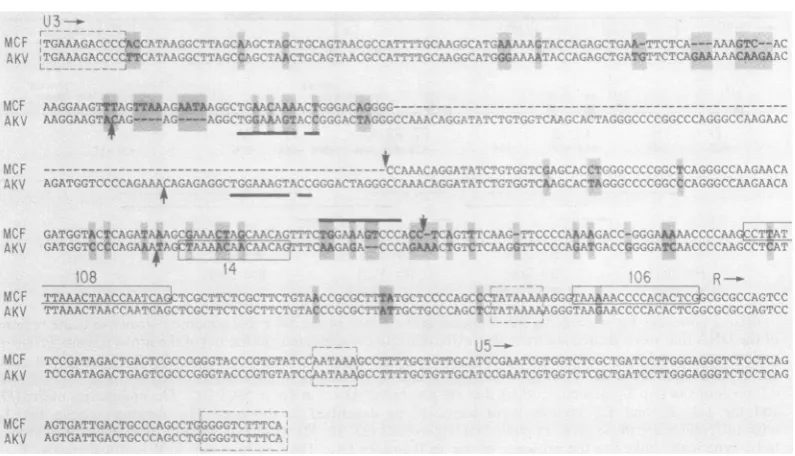
FIG. 3. Comparisonof nucleotideseqencesofDNAclonescorrespondingtothe 3' ends ofAkvand MCF247
viralgenomes. Nucleotide sequencesencoding
Prpl5E
andaregionbetweenPrpl5E
and the LTRbelieved tocontain theoriginofplus-strand DNAsynthesis. Theamino terminus ofPrp15Ewas identifiedby comparison
with dataforMoloneyleukemia virusasdescribedin thetext.Openboxes enclosesequencescorrespondingto
large RNase
T,-resistant
oligonucleotides analyzed previously in Akv and MCF 247 fingerprints (23, 32).Nucleotide differences betweenthevirusesareaccentuatedbygrayboxes andarrowheadsabovethe sequences.
Amino acids ofPrp15Ethatwould differ betweenthetwovirusesareindicatedwithin grayboxes below theAkv sequence. Dotted lines indicate thatthePrp15EofAkvis twoamino acidsshorter than that of MCF 247. The
largearrowheadabove the sequencesindicatestheputative cleavagesiteforgeneratingmature
piSE
(seetext).The last nucleotide inthe figureis thefirstprecedingthe LTR.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.489.53.450.320.582.2]P,PC.AAC'TTI,.-rrTAA,.-AATA` AlC. PAA:4
;AG-:.--M .CCPilC *A... A TA: .C
rT~'GGACTC.ACATAAAC.GAtiCT G;'.: TC.AG( C A: '.T - AA :A 'TV
MC;TTAAC,AAC,-CC'CAA.,-iAC:' AAC. A2%.* TA ..A '..'.G: G T7.C.t 7.. .2
[image:4.489.52.450.73.301.2]; I'TtrAAACTAAC
.--.AV
ATTA T <G:...ATr".T;AG.'
C»VqCAACTG( ATT C'P2C-C
FIG. 4. Nucleotidesequencesofthe LTRsofAkvversusMCF 247. Theboundaries ofU3, R,and U5were
determined as described in the text. Dotted line boxes enclose the inverted repeats of the LTR, a putative
transcriptional signal,and thepolyadenylicacidadditionsignal.Otherboxesare asinFig.3.The smallarrows
indicatethe boundaries ofadirectrepeatinAkv andasequencethat appearsas adirectrepeatinother DNA
clones of MCF247.Heavyblack lines withindirectrepeats underlineasequenceshared with simianvirus 40(see
addendum).
Large
T,
oligonucleotides
analyzed
previous-ly by
Rommelaereetal.(32)
andLung
etal.(23,
24)
were first located in the DNA sequences ofMCF247
by scanning
thesequencing
gels
for14-to
35-base-long
gapsin the G(or
C)
lane(s)
(Fig.
5).
Comparison
ofthe DNA sequences within these gaps with thepancreatic
RNasedigestion
products
oftheT,
oligonucleotides,
determinedpreviously
by
Rommelaere et al.(31,
32)
andLung
et al.(23),
orto thecomplete
nucleotidesequences of the
T,
oligonucleotides
ofAkv,
determined
by
Pederson and Haseltine(29),
allowed identification of the
oligonucleotides.
TheT,
oligonucleotides
studiedpreviously
areoutlined
by
boxes inFig.
3 and 4.In
Fig.
3 we have indicated thepredicted
amino acid sequences of thePrpl5Es
andpiSEs
of Akv andMCF 247.Identificationof theamino terminus ofPrpl5E
and theC terminus ofpiSE
issolely
by
comparison
with data obtained forMoloney
virus. Henderson and hiscolleagues
(17)
determined the amino andcarboxy
terminal residues ofpiSE
ofMoloney
virus, andSutcliffetal.
(35)
and Shinnicketal.(34)determined the nucleotide sequence ofaDNAcloneofMoloney
virus,
allowing
placement
of thePrpl5E-coding
portion
ofenv. Acomparison
ofour data with thatforMoloney
virusPrpl5E
is shown inFig.
6. InMoloney virus, Prpl5E
isproteolytically
cleaved toyield
mature virionpiSE (13, 34).
Thiscleavage
involves the removalof the 16C-terminal amino acids from
Moloney Prp15E.
Theanalogous
cleavage point
isindicated inFig.
3 and 6forthepresumptive
Prpl5Es
of Akv andMCF 247. In
Akv,
thiscleavage
would releasea19-amino-acid
peptide;
in MCF247,
it wouldrelease a 21-amino-acid
peptide.
It is apparentfromthe datain
Fig.
3 that sequencescoding
forthe C terminus of
Prpl5E
of MCF 247 werederived from its
nonecotropic
parent, whereasthe
majority
of thepl5E-coding
sequenceswereshared with Akv.
Itisapparentfrom
Fig.
4that the U3portion
of the LTR of MCF 247 was derived from its
nonecotropic
parent. The R andU5
portions
of the LTR are identical to those of Akv. The inverted repeat sequence,Hogness
box, andputative polyadenylic
acid additionsignal
occu-py similar
positions
in Akv and MCF 247(36).
DISCUSSION
We determined the nucleotide sequence of
approximately
1.3 kilobases of DNAcorre-sponding
tothe 3' terminus of the MCF 247 viral genome.By analogy
withMoloney
leukemia virus(13, 17,
34),
we identified sequencesen-coding
the aminoterminus ofPrp15E,
aprotein
which in MCF 247 is
specified
by
a 603-nucleo-tide openreading
frame. The termination codonat the end of this sequence is followed
by
34 basespresumably containing
theorigin
ofplus-strand
DNAsynthesis
(10, 26)and thenby
on November 10, 2019 by guest
http://jvi.asm.org/
NUCLEOTIDE SEQUENCE OF 3' END OF MCF 247 295
the LTR, defined in MCF 247 by 11-base-long inverted repeats at its termini. The particular clone of MCF 247 whose complete 3' end se-quence was determined possessed an LTR524
base pairs long. This LTR did not possess an
internal directrepeatsequence ashas beenseen
in LTRs of several other retroviruses (6, 20, 34, 35). However, the LTR of another MCF DNA clone, whosesequence waspartially determined
(data not shown), did contain two copies ofa
105-base-long direct repeat sequence. Several
aspectsof thesequences,of their comparisonto
corresponding sequences of Akv and Moloney viruses, and their use in identifying genomic
hallmarks of biologically distinct MCF viruses
are ofnote.
Prp15Esequences. Thepresumptive Prpl5Es
of MCF 247 and Akv are 201 and 199 amino acids long, respectively, and the nucleotide se-quences encoding them are 96% homologous.
Moloney Prpl5E is 1% amino acids long (34). Although the nucleotide sequence ofMoloney
Prpl5E is only about 77% homologoustothat of MCF247, the predicted amino acidsequencesof their respective proteins are 90% homologous.
Figure 6 shows a comparison of the predicted
amino acid sequences of the Prpl5Es of the three viruses. As noted by Lerner and his col-leagues (34), the C terminus of Prpl5E, aregion
that is proteolytically removed to yield mature
virionp1SE, is variable among different murine typeC viruses.
Lenzetal.(21) havepresentedamodel for the
placement of theenvprecursor, and itsPrpl5E portion in particular, relativeto the membrane. Inthese models amino acids 134through 164 of Prpl5E extend through the membrane, and the remaining C-terminal tail is cytoplasmic. The
rulesemployed todevelop these models would apply with similar resultstothepredicted MCF 247Prpl5E amino acidsequence.
It isapparentfromFig. 3 that themajority of thePrpl5E-coding region of MCF 247 is shared with Akv virus, although sequences encoding
theC terminus, commencingatnucleotide507,
mustbe derived fromthenonecotropicparent of
MCF247. Because thenonecotropic parents of
MCF viruses have not been isolated, wecould
not, from these data alone, have been certain
that the majority of the plSE of MCF 247was
derivedfrom Akv. However, Chattopadhyayet
al.(1, 2) found that the XbaI cleavage siteinthe Prpl5E gene(Fig. 3), asite sharedbyAkv and
MCF247, is found only in the Akv endogenous virus of AKR mice. Thus, this region of MCF 247 must be ecotropic in origin, and it seems
reasonable to suppose that the majority ofthe
Prpl5E gene of MCF 247 was inherited from
Akv. There arethreenucleotide differences
be-tweenAkv and MCF247 in theamino
[image:5.489.327.378.75.450.2]terminal-A4 TIC GO
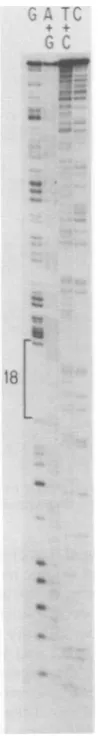
FIG. 5. Sequencing gel showing aregionof DNA
encodingPrp15E thatcontains the large
Tl-resistant
oligonucleotide18.coding
portion of thePrpl5E
gene. One oftheseleadstothedisplacement ofan Akv T1
oligonu-cleotide (35 inFig. 3) in fingerprintsof MCF 247
(32). These three base changes may simply be
point
mutations; alternatively, they could havebeenintroduced by recombination. Inthe latter
case, since sequences encoding the Cterminus
of
gp7O
of Akv and MCF 247 are identical(unpublishedresults) and since the
XbaI
site inMCF 247 is derived from Akv, two crossover
events would probably be needed to introduce the three nucleotide changes.
A major motivation for our studies was to
localize ecotropicandMCF-specific Tl
oligonu-cleotides whose presence or absence in MCF
genomic
T,
fingerprintshadbeen found by Lunget al. (23, 24)to becharacteristicof
biologically
distinctgroupsof MCFs(4, 33).Figures 3 and 4,
in conjunction with the previously determined
VOL.45, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
Phe. . . . - . . . . .. : .: ;< - . :
*i".:t. 'irai.~~~Vo A^rg er .
Va~~~~~~~I Sr
Aoi
Arg Me~st
ser Asp Pro GiuiVol GIU
*Tr
AS CY-*. Lysyr Gn Pro .
FIG. 6. Comparisonofthe predictedaminoacidsequencesof the
Prpl5Es
ofAkv, MCF247, andMoloneyleukemiaviruses. The arrowhead indicates thepositionofproteolytic cleavageto generatematurep15.
T1 fingerprints,reveal that classI,leukemogenic
MCFsisolatedfromthymic neoplasmscan
prob-ably deriveallormostof the amino-terminal half
ofp1SE fromtheirecotropic parentsince almost
all of the viruses possessthe threeecotropic
T,
oligonucleotides 35, 19, and 18 in Fig. 3. Abouthalf of the viruses then resemble MCF 247 in
continuing with ecotropic sequences almost up
totheC terminusofenv.Like MCF247, howev-er,allpossesstheMCF-specific
T,
oligonucleo-tide 101 attheveryend of thisregion.Theotherhalf of theleukemogenicclass I MCFs
apparent-Z
gP70 Prpl5E A U3SC/ossIMCf:
C/assIMCF
as
::~~~~~~~~~~~~~~~~~----::
---::::
[image:6.489.50.243.488.597.2]Derivedfromecotropicparent 1 Derived fromnon-ecotropic * Derived fromeitherparent prent
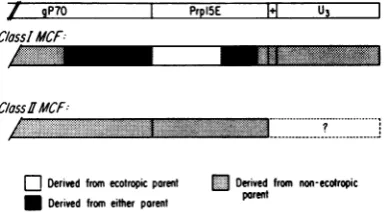
FIG. 7. Diagrammaticrepresentationofthe 3'ends
of class I andclass IIMCFviruses, indicatingwhich
regionsarederivedfrom theecotropicand nonecotro-picparents. The conclusion that the patterns shown
are representative of viruses belonging to the two classesof MCFs is baseduponthe assignmentofT, oligonucleotidestoparticular genetic elements, deter-mined fromDNAsequencing,andontheT1
oligonu-cleotidemapsobtainedby Lungetal. (23,24).
ly undergo recombination in the middle of the p15E gene and thus lose the two ecotropic T1
oligonucleotides 47 and 33 (Fig. 3) andacquire
oligonucleotide 101. This general pattern is sum-marized schematically in Fig. 7. Interestingly,
oligonucleotide analysis of a group of MCFs
isolated from thymus ofAKR mice by O'Don-nell etal. (28) suggests that if the recombination event in the middle of Prpl5E that generates someclass IMCFsoccurstoofarto the 5'
side,
sothatoligonucleotide18 islost, then the
result-ing MCF viruses are non-leukemogenicor only weakly so in the AKRaccelerationassay (M. L. Lung, P. O'Donnell, and N. Hopkins, unpub-lished results). In other words, ecotropic
T,
oligonucleotide 18 in Prpl5E-coding sequences appears to be genetically linked to the highly
leukemogenic phenotypeofclass I MCFs.
Class II MCFsisolated from nonthymic neo-plasms lack all of theecotropic
T,
oligonucleo-tidespresent inPrpl5Eandthusmayderive this
entire portion of env from their nonecotropic
parents (4, 23). These viruses all possess the
MCF-specific
T,
oligonucleotide101 encodingaC-terminalregion ofPrp15E.
The termination codonatthe end of Prp15E is followedby 34 nucleotides that are highly con-served between MCF 247, Akv, and Moloney
viruses and which may encode the origin of
plus-strandDNA synthesis(10, 26, 34).
LTR. As noted above, the MCFDNA clone we sequenced possessed only one copy of a
potentiallydirectrepeatsequence.Identification
on November 10, 2019 by guest
http://jvi.asm.org/
NUCLEOTIDE SEQUENCE OF 3' END OF MCF 247 297
ofthis sequencewasmade
by partially
determin-ing thenucleotide sequenceof another MCF 247
clone, clone p247-13,possessingadirect repeat.
Khan et al. (20) also sequenced an MCF LTR
containing adirect repeat. It is apparentthat the
direct repeats of Akv and MCF are staggered
relative to one another (seearrowsin
Fig.
4).All class I, thymic MCFs whose
genomic T1
fingerprints havebeenexamined lack the
ecotro-pic T1 oligonucleotide 14 in U3 (23, 24). Like
MCF 247, all of the highly oncogenic isolates
alsoacquire
oligonucleotides
106and 108 inU3,
whereas one non-leukemogenic isolate did not.
However, isolates which possess the pattern
oligonucleotide 14 absent, 106 and 108 present
are not necessarilyleukemogenic, sinceasnoted
above, loss ofecotropic
oligonucleotide
18in thepl5E gene is associated with low oncogenicity.
Class II MCFs, all of which are
non-leukemo-genic upon injection (4, 33), share
oligonucleo-tide 14 in U3 with their
ecotropic
parents, andnone
possesses
oligonucleotide
108(23,
24).These observationsare
probably insufficient
todetermine theparental
origin
ofthe U3s of classII MCFs, although theyare consistent with the
possibility that they are inherited from the
eco-tropic parents.
Conclusion.These
studies,
inconjunction withT1
fingerprinting, have suggested that particularnucleotide and amino acid sequences of
Prpl5E
and U3 contribute to the oncogenicity of MCF
type C viruses ofinbredmice. The role ofgp7O,
pl5E, and the LTR indetermining this complex
viral phenotype remain unclear.
ACKNOWLEDGMENTS
WethankTom Gingeras for his generous help in setting up computer programs toanalyze the sequence data. We thank Elaine Rands for expertassistance in themolecular cloning of MCF 247 and Joe Sambrook for many helpfuldiscussions. We thankWinship Herr and Victoria Corbin for providing us with sequences of thePrpl5E-codingregion of Akv virus during the course of this work and Jack Lenz and Bill Haseltine for a preprint of theirsequence of the Akv env gene.
This work was supported by Public Health Service grant CA19308 and Program Projectgrant CA26717 to N.H.H. from the National Institutes of Health and partially by Public Health Service core grantCA14051 to S. E. Luria. C.A.H. acknowledges the support of the Charles A. King Trust, Boston, Mass.,and the Muscular DystrophyAssociation, and M.L.L. acknowledges Public Health Service postdoctoral fellowship CA06511 from the National Institutes ofHealth.
ADDENDUM
We noted above that when the LTR sequences of
Akv and MCF 247arealignedasshownin Fig. 4, their
direct repeat sequences within U3 are staggered
rela-tive to one another. Recently, Laimons et al. (Proc.
Natl. Acad. Sci. U.S.A., inpress)obtained evidence
thatasequence criticalforactivator(enhancer)
func-tion in simian virus 40 is TGG' G, which occurs
withinthesequenceTGGAAAGTCCCinsimianvirus
40 and is also found in other viruses and suspected
enhancer regions. An identical 11-base sequence is
found in thedirectrepeatoftheMCF 247LTR,and 10
bases of the 11-basesequenceare presentin the Akv
directrepeat. However, in Akv the sequence appears 10 nucleotides from the 5' end of the direct repeat, whereas in MCF 247 it appears four basesfrom the 3' end of the direct repeat. Thus it is apparent (Fig. 4)
that the staggered positions of the direct repeats of Akv and MCF 247 ensure that in both viruses the directrepeatincludesthepotentially critical sequence
notedbyLaimonsetal.
LITERATURE CITED
1. Chattopadhyay, S. K., M. W.Cloyd, D. L. Linemeyer, M.R. Lander, E. Rands, and D. R. Lowy. 1982.Cellular
origin and role of mink cell focus-forming viruses in
murine thymiclymphomas. Nature (London) 295:25-31. 2. Chattopadhyay,S.K., M. R. Lander, S. Gupta, E. Rands,
and D.R. Lowy. 1981. Origin of mink cytopathic focus-forming (MCF) viruses: comparisonwith ecotropic and xenotropic murine leukemia virus genomes. Virology
113:465-483.
3. CleweUl, D. B. 1972. Nature of ColElplasmidreplication
inEscherichia coli in the presence of chloramphenicol. J. Bacteriol.110:667-676.
4. Cloyd, M. W., J. W. Hartley, and W. P. Rowe. 1980.
Lymphomagenicity of recombinant mink cell
focus-induc-ingmurine leukemia viruses.J. Exp. Med.151:542-552.
5. Cohen, S. N., A. C. Y. Chang, and L. Hsu. 1972. Nonchro-mosomal antibiotic resistance in bacteria; genetic trans-formation of Escherichia coli by R-factor DNA. Proc. Natl. Acad. Sci. U.S.A. 69:2110-2114.
6. Dhar, R., W. L. McClements, L. W. Enqulst,and G. F. VandeWoude. 1980. Nucleotide sequences of integrated Moloneysarcoma provirus long terminal repeats andtheir
host and viraljunctions. Proc. Natl. Acad. Sci. U.S.A. 77:3937-3941.
7. Elder, J., F. Jensen, R.Lerner,J. W.Hartley,and W.P.
Rowe. 1977. Biochemical evidence that MCF murine leukemia viruses are envelope (env) gene recombinants. Proc.Natl. Acad. Sci. U.S.A. 74:4676-4680.
8. Faller, D. V., and N. Hopkins. 1977. RNaseT1-resistant
oligonucleotides of B-tropic murineleukemia virusfrom BALB/c and five of its NB-tropic derivatives. J. Virol. 23:188-195.
9. Famulari, N. G., D. L. Buchagen, H. D. Kienk, and E. Fleissner. 1977. Presence of murine leukemia virus enve-lopeproteinsgp7O andpl5(E)ina commonpolyproteinof
infected cells. J. Virol. 20:501-508.
10. Gilboa, E., S. W. Mitra, S. P. Goff, and D. Baltimore. 1979. Adetailed model of reverse transcription and tests ofcrucial aspects. Cell18:93-100.
11. Gingeras, T. R., J. P. Milazzo, D. Scaky, and R. J. Roberts. 1979. Computer programs for the assembly of DNA sequences. Nucleic Acids Res. 7:529-545. 12. Graham, F. L., and A. J. van der Eb. 1973. A new
technique for the assay ofinfectivity of human adenovirus S DNA.Virology 52:456-467.
13. Green, N., T. M.Shinnick, 0.Witte, A. Ponticelli, J. G.
Sutcliffe,and R. A. Lerner. 1981. Sequence-specific anti-bodies show that maturation ofMoloney leukemia virus envelopepolyprotein involves removal of a COOH-termi-nalpeptide. Proc. Natl. Acad. Sci. U.S.A.78:6023-6027.
14. Grunstein, M., and D. S. Hogness. 1975. Coloney hybrid-jzation: a method for the isolation of cloned DNAs that contain a specific gene. Proc. Natl. Acad. Sci. U.S.A.
72:3961-3965.
15. Hartley, J. W., and W. P. Rowe. 1975. Clonal cell lines from a feral mouse embryo which lack host-range restric-tions formurine leukemia viruses. Virology 65:128-134. 16. Hartley,J. W.,N. K.Wolford,L. J.Old, and W. P. Rowe.
1977. A new class of murine leukemia virus associated with development of spontaneous lymphomas. Proc.
Natl. Acad. Sci. U.S.A. 74:789-792.
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
17. Henderson, L. E., T. D. Copeland, G. W. Smythers, H. Marquardt, and S.Oroszlan. 1978.Amino-terminal amino acid sequence andcarboxyl-terminal analysis of Rauscher murine leukemia virusglycoproteins. Virology 85:319-322.
18. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369. 19. Karshin, W. L., L. J.Arcement, R. C. Naso, and R. B.
Arlinghaus.1977.Common precursorfor Rauscher leuke-miavirus gp69/71, p15(E), and p12(E). J. Virol. 23:787-798.
20. Khan, A. S., R.Repaske, C. F. Garon, H. W. Chan, W. P. Rowe, and M. A.Martin. 1982. Characterization of pro-viruses cloned frommink cell focus-forming virus-infect-edcellular DNA. J.Virol. 41:435-448.
21. Lenz, J., R. Crowther, A. Straceski, and W. Haseltine. 1982.Nucleotide sequenceof the Akv env gene. J. Virol. 42:519-529.
22. Lowy, D. R., E. Rands, S. K. Chattopadhyay, C. F. Garon, and G. L. Hager. 1980. Molecular cloning of infectious integrated murine leukemia virus DNA from
infected mouse cells. Proc. Natl. Acad. Sci. U.S.A. 77:614-618.
23. Lung, M. L., J. W. Hartley, W. P. Rowe, and N. H. Hopkins. 1982. Large RNaseT1-resistantoligonucleotides
encoding plSE and the U3 region of the long terminal repeat distinguish two biological classes of mink cell
focus-forming type C viruses of inbred mice. J. Virol. 45:275-290.
24. Lung, M. L., C.Hering, J. W. Hartley, W. P. Rowe, and N.Hopkins. 1979. Analysis ofthegenomes of mink cell focus-inducing murine type C viruses: a progress report.
ColdSpringHarborSymp. Quant.Biol. 44:1269-1274. 25. Maxam, A. M., and W. Gilbert. 1977. A newmethod for
sequencingDNA. Proc.Natl. Acad. Sci. U.S.A. 74:560-564.
26. Mitra,S. W., S. Goff, E. Gilboa, and D. Baltimore. 1979.
Synthesis of a 600nucleotide-long plus-strandDNA by
virions of Moloney murine leukemia virus. Proc. Natl. Acad.Sci. U.S.A. 76:4355-4359.
27. Naso, R. C., L. J. Arcement, W. L. Karshin, G.A. Janjoom,and R.B.Arlinghaus. 1976. Afucose-deficient glycoprotein precursor to Rauscher leukemia virus
gp69/1.Proc.Natl. Acad. Sci. U.S.A. 73:2326-2330. 28. O'Donnell,P.V., E.Stockert, Y. Obata,A. B.DeLeo,and
L. J. Old. 1979. Murine-leukemia-virus-related cell-sur-faceantigens as serological markersof AKR ecotropic, xenotropic, and dual-tropic viruses. Cold Spring Harbor Symp.Quant. Biol. 44:1255-1264.
29. Pedersen, F. S., and W. A. Haseltine. 1980.Analysisof the genome of an endogenous ecotropic retrovirus of the AKRstrain of mice: micromethod for detailed character-ization of high-molecular-weight RNA. J. Virol. 33:349-365.
30. Rands,E., D. R. Lowy, M. R. Lander, and S. K. Chatto-padhyay.1981. Restriction endonucleasemapping of
eco-tropic murineleukemia viral DNAs: size and heterogene-ity of the long terminal repeat. Virology 108:445-452. 31. Rommelaere, J., D. V. Failer, and N. Hopkins. 1977.
RNaseTI-resistantoligonucleotides of Akv-1 and Akv-2 typeCviruses of AKR mice. J. Virol. 24:690-694. 32.Rommelaere, J., D. V. Faller, and N. Hopkins. 1978.
Characterization andmapping of RNase Ti-resistant oli-gonucleotides derived from the genomes of Akv and MCF murineleukemia viruses. Proc. Natl. Acad. Sci. U.S.A. 75:495-499.
33. Rowe, W. P., M. W. Cloyd, and J. W. Hartley. 1979. Status of theassociationof mink cellfocus-forming virus-es with leukemogenesis. Cold Spring Harbor Symp. Quant. Biol. 44:1265-1268.
34. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981. Nucleotide sequence ofMoloney murine leukemia virus. Nature(London) 293:543-548.
35. Sutcliffe, J. G., T. M. Shinnick, I. Verma, and R.A.
Lerner. 1980.Nucleotide sequence ofMoloney leukemia virus 3' end reveals details of replication, analogy to bacterial transposons, and an unexpected gene. Proc. Natl.Acad. Sci.U.S.A. 77:3302-3306.
36. VanBeveren, C., E. Rands, S. K. Chattopadhyay, D. R. Lowy, and I. M. Verma. 1981. Long terminal repeat of murine retroviral DNAs: sequenceanalysis, host-proviral junctions, and preintegration site. J. Virol. 41:542-556. 37. van deVen, W. J. M., D. van Zaane, C. Onnekink, and
H. P. J. Bloemers. 1978.Impaired processing of precursor
polypeptides of temperature-sensitive mutants of Rauscher murine leukemia virus. J. Virol. 25:553-561. 38. Witte, 0. N., A. Tsukamoto-Adey, and I. L. Weissman.
1977.Cellular maturation ofoncornavirusglycoproteins: topological arrangement of precursor and product forms in cellular membranes. Virology 76:539-553.