JOURNALOFVIROLOGY,Jan. 1994. p. 187-194 Vol.68,No. 1 0022-538X/94/$04.00()+
Copyright
© 1994,AmericanSociety for MicrobiologySelective Sites of Adenovirus
(Foreign)
DNA
Integration
into
the
Hamster
Genome: Changes
in
Integration
Patterns
GERTRAUD OREND, ANDREAS LINKWITZ,t ANDWALTER DOERFLER*
Institiute
for Genetics, University ofColognze,
D-50931Colognze,
Germany Received28 June 1993/Accepted 29September 1993Weinvestigatedwhether, upon the integration of multiplecopies of adenovirus type 12 (Adl2)DNA into an established mammalian (hamster) genome, the pattern of foreign DNA insertion would remain stable or change withconsecutive passages of cells in culture. By the injection of purified Adl2 into newborn hamsters, tumors were induced, cells from these tumors were cultivated, and five independent cell lines, HT5, H201/2, H201/3, H271, and H281, wereestablished. These cell lines carried different copy numbers ofAdl2 DNA per cell in an integrated form anddiffered inmorphology. Cell line HT5 had been passed twice through hamsters astumor cells and wassubsequently passaged in culture. Patterns ofAdl2 DNAintegration were determined by restrictioncleavage of the nuclear DNA withBamHI, EcoRI,HindIII,MspI,orPstI followed by Southern blot hybridization using
32P-labeled
Adl2DNA or its cloned terminal DNA fragments as hybridization probes. In this way, theoff-sizefragments, which represented the sites of linkage betweenAdl2 andcellular DNAs, were determined. At early passage levels in culture, the integration sites of Adl2 DNA in the hamster genome, as characterized by the positions of off-size fragments in agarose or polyacrylamide gel electrophoresis, were different in the fivedifferenttumor cell lines. Upon repeated passage, however, theoff-sizefragment patterns generated by the five restriction endonucleases became very similar in the five tumor cell lines. This surprising result indicates that under cell culture conditions,Adl2-transformed tumor cell lines that carry the foreign(Adl2)genome in selective, probably very similar sites of the cellular genome evolve.
Studies onthe integration offoreign (viral) DNA intothe preexisting genomes ofmammalian cells can serve an
exem-plary
function in the analysis of the mechanisms and the outcome ofinsertional recombination between foreign DNA and an established genome. These interactionsare of interest not only in tumor virology and gene therapy but also formodels about the evolutionary role of foreign DNA integra-tion. A largenumber ofadenovirus-transformed cell lines or adenovirus type 12 (Adl2)-induced tumor cells or cell lines
havebeeninvestigatedforthesites ofviral DNAinsertion (for
a review, see reference 1). For a number of junction sites
between adenovirus and cellular DNAs, the nucleotide se-quences have been determined (2,6, 12). Inthese cell lines,a
specific cellular nucleotide sequence at the insertion sites for
adenovirusDNAhas not beenfound. However,allintegration sites of adenovirus DNA investigated in that respect have
proved tobe transcriptionally active both in the transformed cells and in the original cells prior to adenovirus infection, integration, and transformation (3, 18). Short patch homolo-gies betweenviral and cellularjunction sequenceshaveoften,
but not always, been found.
Recently,we have initiated work oncell-free systems, using
nuclear extracts from hamster or insect cells to elucidate details of the mechanism of insertional recombination (5, 17, 24, 25). The recombinants generated in the cell-free in vitro system arealsocharacterized bypatchhomologiesbetween the
recombinationpartnersatthesites of
junction (24,
25). Several times,the hamster cell sequence CCTCTCCG has been found tobe therecombination target, or to belocated closetoit,
with adenovirus DNA in several independent cell-freerecombina-tionexperimentsin which extracts
purified
todifferentextentsCorresponding author. Mailing address: Institute for Genetics, UniversityofCologne,Weyertal 121,D-50)931K6ln, Germany.Phone: 49-221-470-2386. Fax:49-221-470-5163.
t Present address: BayerAG, Leverkusen, Germany.
have been used (24, 25). The same hamster cell nucleotide
sequence has been identified at the site of linkage to Adl2
DNA in the Adl2-induced hamstertumor CLACI (21).
In a series of
Ad12-induced
hamster tumor cell lines (11,14), we have now analyzed the dynamicsof the sites of viral DNA insertion.
MATERIALS ANDMETHODS
Most of the techniques used in this study have been de-scribed previously (14).
Adl2-induced hamster tumor cell lines. Hamster tumors were induced by the injection of Adl2, and the cell lines H201/2, H201/3, H271, and H281 were established from tu-mors T201/2, T201/3 (two different tumors in the same ani-mal), T271, and T281 and passaged in Dulbecco medium enriched with 10% fetal calfserum(14). Conditioned medium
was obtained by growing BHK-21 hamster cells in regular
Dulbecco medium with 10% fetal calfserum for24h, sterile filtering this medium, and then using itin a 1:1 mixture with fresh Dulbecco medium
supplemented
with 5% fetal calfserum for the cultivation of Adl2-induced tumor cells. The hamster tumorT313 was induced, and the cell line H313was established morethan 10 years ago
(11).
The H313 cellswere passagedtwicethrough hamsters thatdevelopedtumorsuponinjection of H313 cells. From the second tumor passage,cell
line HT5 was derived. The tumors and cell lines T-H201/2, T-H201/3a, T-H271, T-H281, H313, and HT5 contained
ap-proximately 10, 3, 2 to 3, 5, 30, and 10 Adl2 genome
equivalents percell,
respectively
(14).
The cell lines in culture werepassagedatdifferentsplit
ratiosranging
from 1:2to 1:10. Analyses by Southern blot (19) and DNA-DNA hybridiza-tion. In differentexperiments,
DNAwas cut with one of five different restrictionendonucleases,
BamHI,
EcoRI,
HindlII,
MspI,
and PstI. Thegenerated
fragments
were transferred inalkaline medium(0.25M
NaOH,
1.5 MNaCl,
or0.4 MNaOH)
187
on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 1. Characteristics ofAdl2-induced tumorsandof hamster tumorcell linesestablished from them
Tumororcell line No.of
Adl2
genome SimilarAdl2
Denovomethylationof Cellmorphologyequivalents/cell' integration
patternsb
integratedAdl2DNAC
Tumor
T313c 30 + +
T5d 10 +
T191 3-5 + +
T211 3 + +
T201/2 10 - +
T201/3 3 - +
T271 2-3 - +
T281 5 - +
Celllinee
H313 30 + + Trapezoid
HT5 10 + - Trapezoid
H191 3-5 + Fibroblastic
H211 3 + Fibroblastic
H201/2 10 + + Trapezoid
H201/3
a 3 + + Trapezoid
b NDf ND ND Fibroblastictotrapezoid&
H271
Grown in normal medium 2-3 + + Fibroblastictotrapezoid
Grownin conditioned ND ND ND Fibroblastictotrapezoid
medium
H281 5 + + Fibroblastic totrapezoid
Data takenfrom reference 14.
bFor tumors,symbols represent similarities among the tumors and corresponding celllineslisted. For cell lines, symbols represent similarities observed in HT5 at p6,H201/2 atplO, H201/3aatp12,andapproachinglate stagesof cell lines H271 atp13and H281 at pll.
cDatataken in part from reference 10.
dAgift of Ingrid Kuhlmann, Universityof Konstanz. The integrated adenoviral DNA is already heavily de novo methylated in tumor T5. The methylation state does notfurther increaseduringcultivation of cell line HT5.
e In cell lines H313, HT5,H191, H211, and H201/2, the described parameters applied to several passages. Cell lines H201/3a, H271, and H281 were analyzedin p3, p5,and pl,respectively.
fND,CompleteAdl2genomes are not detectable. Themajority ofpreviously integrated Adl2 sequences is deleted. gUponcultivation,cellmorphologychangedfrom fibroblastic totrapezoid.
toHybond N+ nylon membranes. As a32P-labeled hybridiza-tion probe, Adl2 DNA or one of the terminal fragments of Adl2 DNA, the EcoRI C (left end) or the BamHI E (right
end)
fragment, was used. Hybridization and wash conditionswere described previously (7, 14). These probes could be repeatedly removed by boilingin 2 x SSC(1x SSCconsists of 0.15 M NaCl and 0.015 M sodium citrate) and 0.1% sodium dodecyl sulfate.
In some
experiments,
DNA fragments were separated by electrophoresison5% denaturing polyacrylamide gels contain-ing 7 M urea, electrotransferred to HybondN+ nylonmem-branes, and then hybridizedto
32P-labeled
probesasdescribed previously (14).Restriction maps ofAdl2 DNA. The complete nucleotide
sequenceof Adl2DNA wasdetermined inourlaboratory
(20).
The restriction maps presented in Fig. 2fwere derived from thissequence.
RESULTS AND DISCUSSION
Alterations in cell morphology and integration patterns of
Adl2 DNA in Adl2-induced hamster tumor cell lines. The
Adl2-inducedhamster tumor cell lines H201/2, H201/3, H271,
H281, and HT5 were described previously (14), and their
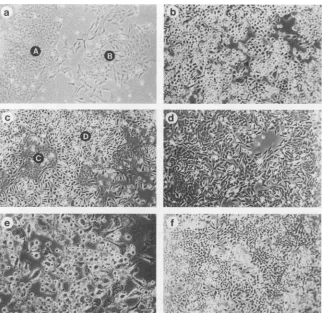
characteristics are summarized inTable 1. Both the morphol-ogy ofthe tumorcells in culture (Fig. 1) and the patterns of
viralDNAintegration underwent distinct changesas cells were
explanted
from the tumor-bearing animals and repeatedlypassaged in culture. Changes in integration patterns were
manifestedby alterations in the positions ofthe off-sizeAdl2
DNAfragments upon agarose orpolyacrylamide gel
electro-phoresis. These fragments constituted the junctions between
the termini of Adl2 DNA and the abutting cellular DNA sequences.
Patterns ofAdl2DNAintegration inAdl2-inducedhamster tumor cells. The DNA extracted from the original tumors, T201/2, T201/3, T271, and T281, was analyzed by using one of
the five restriction endonucleases BamHI, EcoRI, HindlIl,
PstI, and MspI and by Southern blotting. The integration
patterns of Adl2 DNA in tumors T201/2 and T271 are
reproduced inFig. 4a and 3, respectively (seebelow). Some of
the integration patterns ofAdl2 DNA in these tumors were
described in detail previously(14). As a probe for
hybridiza-tion, the
32P-labeled
Adl2 DNA orthe terminal Adl2DNA fragment EcoRI C (left) or BamHI E (right) was used insuccessive hybridization experiments. The autoradiograms in Fig. 2a through d present the cleavage patterns of the
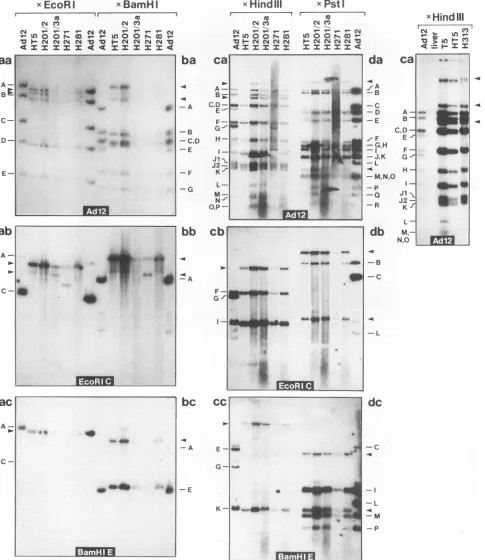
inte-grated Adl2genomes generated byEcoRI (Fig. 2a),BamHI (Fig. 2b), HindIII (Fig. 2c), PstI (Fig. 2d), andMspI (Fig. 2e)
with the DNA of cell lines HT5, H201/2, H201/3a, H271, and H281. The restriction maps for AdI2DNA foreach of these
restriction endonucleases are reproduced in Fig. 2f. These maps were drawn according to the complete nucleotide se-quence of Adl2 DNA that was recently determined in this
laboratory (4a, 20). As expected, the internal viral DNA
fragments comigratedwith the Adl2DNAmarker fragments and were identical in the DNAs from the different tumors,
indicating that the Adl2 genomes were integrated in an orientation colinear with that inAdl2virion DNA. Theoff-size
fragmentswith sequence homologies to the left- or the
on November 9, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS DNA INTEGRATION AT SELECTIVE SITES 189
FIG. 1. Changes in cell morphology upon continued passage in culture. (a) Cell line H271 at an early (A) or intermediate (B)stage; (b) cell lineH271 at alate stage.(c)cell line H281 atintermediate stages, early (C) and late (D) cell morphology; (d) cell line HT5; (e) cell line H201/3b (inthis cell line, completeAdl2genomes are no longerdetectable[Table 1]); (f) cell line H201/3a. Photographs were taken under a Leitz inverted microscope at x100magnification.
terminal Adl2DNAfragment or toboth terminiaremarked by arrowheads inthe autoradiograms.
It was apparentfrom the datapresented in Fig. 2ato ethat the off-sizepatternsdifferedamongtheDNAsamples from the different tumorsand from those of cell lines H313 and HT5 (Table 1). The cleavage patternsof theDNAsfrom the latter
two lines are presented for comparison. However, their pat-terns wereidentical tothose approached by cell linesH201/2, H201/3a, H271, and H281 aftercontinuous cell culture
(Fig.
2ca). After cleavage withoneof the five restriction
endonucle-ases EcoRI, BamHI,
HindIll,
PstI, and MspI(Fig. 2),
the integration patterns of Adl2 DNA in the five different cell linesinvestigatedwereverysimilar; in particular, the distribu-tions of the off-size bands(arrowheads
inFig. 2)were practi-cally identical. Thisfinding
suggested that the sites ofviral DNA integration were highly selective at this stage. Asex-plained above, cell line H313 was generated about adecade ago
(11),
and cell line HT5 was derived from it after two hamster tumor cell passages. Cleavageandoff-sizepatternsof these celllineswereidentical for theDNAsfromH313andT5.However, thecopynumber of30Adl2genome equivalents in cellline H313 wasdecreased to 10incell line HT5(Fig. 2ca).
For each of the panels in Fig. 2, the hybridization probe is indicated foreasyappraisal ofthe results.
Intermediate stages ofintegrationpatterns in the transition from tumor cells to tumor cell lines.Upon continuouspassage
of theAdl2-induced hamster tumor cellsin
culture,
the size distribution of the off-sizefragments of theintegrated
Adl2genomes began to change. These alterations first became
apparentin passage 1(pl), p3, p5, andplforcell lines H201/2, H201/3a, H271, and H281, respectively. We have not
illus-trated these
changes
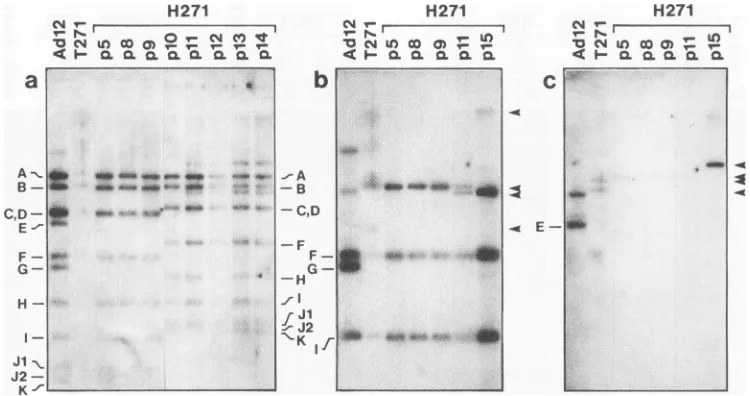
for all four cell lines but ratherpresenta typical example. Theautoradiograms in Fig. 3 demonstrate the HindlIl cleavage patterns of integrated Adl2 DNA in theDNAfrom thetumorT271and from celllineH271in different
passages. It was apparent that starting with p1 1, the distribu-tion of the off-sizefragments
(designated
by arrows)began
tochange and, after passage
p15,
finally
attainedapatternsimilarto the patterns in the other Adl2-induced tumor cell lines H201/2, H201/3a, and HT5. Similar resultswereobtained for theDNAfrom cell line H281 after
cleavage
of theDNAwith HindIll. Some of the Adl2 off-size bands in the HindlIl cleavage patterns ofDNAfrom cell lines H271 andH281 inhigher passage numberswere identical in their size distribu-tions. Similar
findings
were made with the restrictionendo-nucleasesBamHI,EcoRI, and MspI
(data
notshown).
Apparentsimilarities in the patterns of off-sizefragmentsof Adl2 DNA in five different Adl2-induced hamster cell linesas documented by cleavage with five different restriction endo-nucleases. As demonstrated
by
the data inFig.
2 and3,
thecleavage
patternsofDNAfrom the fiveindependently
estab-lished cell lines
generated
with fivedifferentrestrictionendo-nucleases showedverysimilar distributionsofmanyof theleft
and
right
terminal off-size bands. It wasparticularly striking
that these off-size bands had thesamesizes in cell linesH201/2,
H201/3a, H271, and H281 in
comparison
with the tumor cell VOL.68, l1994on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.146.469.73.386.2]190 OREND ET AL.
x
BamH
I
N
cm N v-o O- T-0_
1.-N N N
t Z,x
0
41
S
_*.Y' 4t
AD-ba
ca
A--A
C,D-F
--B
G--C,D
H--E
I_
J1% J2
--F
K- M-N
-O,P
-: 7 bb
..4
..4
4_40
*b
-ExHindIII x
Pst
I
rU
NC) NC)
l-N N NN-FN NNN n
4Zz--- -S-S-4
...
x
HindIII
'a > LO
F-I=
F4h
)da
,rA -B
-c
ca
-D
A--E
B-C,D
-rF E
--:G,H
=I
F--J,K G-'
-L
.4 H--M,N,O
_ p I
-a J1
-R
J2--R
K1s
db
M,-N,O
-.4
-B -C
__
low _ ___
_0m
-
-_ mli I
_
,:_
_.
_I.4
-L
dc
-c
.4
-I
-L
.4
-M
-P
FIG. 2. Patternsofintegrationof Adl2 DNA inthegenomesfrom five different Ad12-inducedhamstertumorcelllines. The DNAs fromcell lines HT5 atp6,H201/2 atplO,H201/3aatp12, H271 atpl1 (aand b) and p13 (c and d), and H281 atpIl (aandb) and p14 (c andd)were
extracted and cleaved withrestriction endonucleasesasdescribed in the text.Thefragmentswereseparatedby electrophoresison0.8% agarose
gels (ato d)andhybridizedto32P-labeled Adl2 DNA(panelsegmentsa),to the left-terminal EcoRI Cfragment (panel segmentsb),ortothe right-terminal BamHI-E fragment of Adl2 DNA (panel segments c). Amounts of authentic, virion-extracted Adl2 DNA cleaved with the corresponding restriction endonuclease andrepresentingapproximately 10(leftmarkerlane)or20(rightmarkerlane)genomeequivalentsper
cellwerecoelectrophoresed asfragmentsize references. Genomeequivalentswerecalculatedonthe basis oftheamountof cellular DNA used
perslot: aandb,10,ug;candd,40fig; ca, 10 jigforcell lines H313 andHT5;e,50 jLg.(a) CleavagewithEcoRI, usingAdl2 DNA(aa)orthe EcoRl C(ab)orBamHI E(ac) fragment of Adl2 DNAasthe hybridization probe. (b) CleavagewithBamHI. (c) Cleavage with HindIll.(d)
xEcoRI
N
cem CO__
N4 .r T- '-. Y-l
V1-N NN N
<=--z :r
aa
A B
C-ab
A-
C-ac
C-
w---RZZ-tillossm
J. VIROL.
"Mi
V,4
_*.tmmw:A <
7iw
!r
-i..
I.V.
.i.,
Ai.
:
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.73.558.77.637.2]ADENOVIRUS DNA INTEGRATION AT SELECTIVE SITES 191
H271
I I
a-
'-m.
a.CL
xN
4c
H201/2
I LO
HT5 a
Q C>J I0) I
NM x
N
2
-1
-2
-3
-4
H271 a
I I
S
WI. ax xQQ
H201/2
I
In
2 x
HT5 m
0)
ND
a.
-o
-1 -2
-3
-4
-5 -5
f
C
BJO
F
|Q|N
K A DE
G HPI
M
LC D B B F A
A G H D I F C B E
G I F B C D M |NL A H K |
a
x
N
cm
4r
-2
-3
-4
-5
Psti EcoRI BamHI Hindlll CleavagewithPstI. (e)Cleavage withHpaIIorMspI,using theEcoRI C(ea) orBamHI E(eb)fragment as the hybridization probe. Fragments 5 represent the very terminal 143-bpMspIfragments at both ends of theAdl2genome. Off-size bands are designated byarrowheads. In panelca, HindIll-cuthamster liverDNAwascoelectrophoresed as a negative control. In panels a and b, DNA from cell line H271 was examined before theintegrationpatternofAdl2DNAhad reached the final stage. In panel ca, equal amounts of DNA were used in all lanes. The autoradiograms for T5 and HT5wereexposedfour timeslongerthan that for H313. (f) Restriction maps of theAdl2genomefor the restriction endonucleases PstI, EcoRI,BamHI, andHindIll.
line HT5. Since thecell lines investigatedwere derived from tumorsinducedby Adl2inhamsters, itwas notsurprisingthat mostof the cell lines contained multiple copiesof
Adl2
DNA(Table 1).Inaddition,someofthecell linescarried additional, submolar fragments of Adl2 DNA which gave rise to weak bands inpositions not identical with anyof the Adl2 marker
DNAbands(Fig. 2). The nature of these weak bands was not
investigatedin detail.
Apartfromthe fact thatthe five cell lines harbored different copy numbers of Adl2 genomes,the increase in the de novo methylation ofthe integrated Adl2 genome in the different cell lines followed different kinetics in the transition from VOL.68, 1994
AMW
on November 9, 2019 by guest
http://jvi.asm.org/
H271
C'
r--<1_p% I Q
Q-
Q0
H271
I_ Q
.ea.%k Qj*:0
vr-,:f
0O C<a
c
F-A_G^v:+; _ 0 _,
[image:6.612.111.486.76.274.2]K tL
FIG. 3. Changes in the distribution of off-size bandsintheDNAfrom the Adl2-inducedhamster tumor T271 and the hamster tumor cell line H271. For details, seethe textand legendtoFig. 2. As the hybridization probe, Adl2 DNA (a)orthe left-terminal EcoRI Cfragment (b) or
right-terminal BamHI Efragment (c) of Adl2 DNAwasused.Inpanels b andc,lanesp15containedmorethan 10 p.gofDNA,anamountthat
wasloaded in all of the other gel slots. Thesignals in this lanewerethereforemoreintense. The data showninpanelawerederivedfrom two differentelectrophoresis experiments and aligned accordingtothepositions of markerDNAfragments.
tumor tocell line.Moreover,the patterns ofmethylationinthe integrated Adl2 genomes in later passages ofthe cell lines
were not identical (Fig. 4) (14, 14a) for the cell lines that exhibitedsimilarintegrationpatternsandsites,asdocumented
by the size distributions of the off-size fragments. An example ispresented inacomparison between theHpaII (H) and MspI
(M) cleavage patterns of DNA from cell lines HT5 and H201/2. The HpaII cleavage patterns of Adl2 DNA were
distinctly different in thetwocell lines(Fig. 4b and c). Similar results were obtained after HhaI cleavage (data not shown). Thus, thepatternsofmethylation in the integrated Adl2 DNA
weredifferent, attesting tothe independent origins of thetwo cell lines.
Intwoother cell lines also derived from the sameseriesof
Adl2-induced hamster tumors, H191 and H211, the integra-tion patterns of Adl2 DNA clearly differed from those just described,evenafter extensive subcultivationof these celllines (datanotshown). We also analyzed thestateofmethylationin several subclones oftheAdl2hamster tumorcell lines H191 andH281, which differedintheir Adl2integrationpatterns.By usingsubfragments of Adl2DNAashybridization probes, cell
line-specific methylation patterns of integrated Ad12 DNA
were found (datanot shown).
It is concluded thatpossiblyas aconsequenceof thetransfer from cells from afreshly induced hamster tumor to thevery
different conditions of cellculture, patternsofforeign (Adl2) DNAintegration can change, leading either to striking simi-laritiesof Adl2DNAintegrationpatternsortoapartial loss of Adl2genomes(Table 1). The datasuggestthat viral DNAcan
be rearranged to sites of linkage to cellular DNA that are
similar or identical after fiveindependent integration events. Apparently, under certain, so far unidentified conditions,
rearrangementscanbehighly selective, perhapsevendirected
tothesamecellular DNAsequences.The latterpossibilitywill have to be further investigated by cloning some of these secondary junction sites and bycomparing the cellular nucle-otide sequences. The data argue for the chromosomal
insta-bilityof the Adl2genomes,which have becomeintegrated into
the cellulargenomeduringtumorinduction,upontransfer and propagation oftumorcells under cell culture conditions.
Evaluation of results. There is much evidence thatarguesin favorofnonspecific insertion of adenoviralDNAorretroviral
genomesintocellulargenomes. It islikely thatmanydifferent cellular DNAsequences can serve asintegrationtargets,with
apossible preference for transcriptionally activesequences or
possibly for origins of DNA replication. On the other hand, it has been observed that Adl2 genomes frequently become
associated with human chromosome 1 (13, 15) or that the genome of adeno-associated virus can become linked to
hu-man chromosome 19 (8, 9, 16). In the present report, the evidence suggests that Adl2 DNA can become integrated at
verysimilar sites ofhamster cell DNA intumorcells that have been explanted from the tumor and have subsequently been carried in culture for longer periods of time. Apparently, initially the Adl2 integration siteswere different in the
differ-ent tumors analyzed. However, upon cultivation of cells, integrated viral DNAsequencesmusthave beenrearrangedor
transferred to cellular sites such that the junction fragments appearedverysimilarwhenanalyzed with five different restric-tion endonucleases. Thereare numerous previouslydescribed examplesinwhich thisremarkablereorganization didnotseem
to occur (1, 22, 23), perhaps because the cell culture and
passage conditions used in earlier workas well as the devel-opmentalstagesof the animalsmayhave been different.
Thepossibility existed that inthecourse ofourstudies,we
would have selected for a subpopulation of Adl2-induced
tumor cells from each of the tumors in which Adl2 had originally been insertedataspecific site which then would have
been thesame orverysimilarin alltumors investigated here. The following observations argue against this possibility and demonstratetheindividuality of the cell lines. (i) The cell lines analyzed exhibit different morphologies (Fig. 1). (ii) They contain different amounts ofintegrated Adl2 DNA per cell (Table 1). (iii) The five different cell lineswere derived from
fourtumorinductionexperiments, anddifferent passage
pro-tocols have been followed. (iv) One of the cell lines was
H 271
N
_,-w° cs,. I na m- cI XtI
la1
coi
Qfir
fia
Qr
ffi ffia
on November 9, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS DNA INTEGRATION AT SELECTIVE SITES 193
N4 H201/2 C4 x
o Q ) 01-t
-A
-B
-C,D -E
-F
-G
-H -1
_rJl -J2 - K
[image:7.612.82.278.68.736.2]-L
FIG. 4. (a) HindIII restriction patterns of integrated Adl2 ge-nomes in the Adl2-induced hamster tumor T201/2 and the corre-sponding cell line H201/2. As the hybridization probe, 32P-labeled Adl2 DNA was used.The Adl2integration patterns differed between tumor andcelllines,asindicatedby arrowheads and the rectangle. In lane pl, less than 10 ,.g of DNA was loaded on the gel. Lane SS contained 10 p.g of sonified salmon sperm DNA. (b) Methylation patterns of integrated Adl2 genomes in the Adl2-induced hamster tumor lines HT5 and H201/2 were different. The high degree of 5'-CCGG-3' methylation in the integrated genomes of T5 remained stable, whereas upon cultivation of cell lineH201/2,anincrease in the extent of 5'-CCGG-3', approaching a comparably high degree of methylation in Adl2 DNAof the tumor T5, was observed. The DNA from the two celllines or from different passages of cell line H201/2 or from Adl2 were cleaved with MspI (M) orHpaII (H), as indicated. Experimental procedures for the generation of Southern blots and theseautoradiograms aredescribedin the text andlegendtoFig.2. As the hybridization probe,32P-labeledAdl2 DNA was used. The HpaII fragments ofauthenticAdl2marker DNA are designated A to X. (c) Tabulationof the increase in 5'-CCGG-3' methylation of Adl2 DNA in tumorT201/2and in cell line H201/2 in comparison with the high degree of Adl2 DNAmethylationin tumor T5.Values indicate no (0), partial (1), or complete (2) methylation of 5'-CCGG-3' sequences.
A-
B-C,D
- E,F-G
- Hj-J
- K,L- MN-N,O
-P
-Q H201/2
X o
'_
oX N - _)
,-< 2 x2 2x
-b.z.nE~~~~~~1~T~
AOA.v
:i i
70
..4
Adl2 T201/2 H201/1 T5
x x x x
HpaII HpaII HpaII HpaII
p1 p5 plO
A 0 0 1 1 1
B 0 0 1 2 1
C/D 0 1 2 2 2
E/F 0 2 2 2 2
G 0 1 1
established completely independentlymorethan 10years ago and has been subjected to two successive hamster tumor passages. (v) The rates of de novomethylation as well as the final methylation patterns of theintegrated Adl2 genomes are similar butnotidentical
(Fig.
4b).(vi)
The transition patterns of Adl2DNAintegration for the cell lines have been different, and in different celllines,
some of the off-size bands are in nonidentical positions.The experimental conditions that can lead to changes in the Adl2 integration patterns, as evidenced by the approach to
similar off-size band patterns
generated
with five different restriction endonucleases and thenot infrequent loss of Adl2 genomesfrom Adl2-transformed cells orfrom Adl2-inducedtumor cells
passaged
in culture(4, 10),
are not understood. The presence ofapparently
similar patterns of Adl2 DNAintegration in five independently isolated hamster tumorcell linesmight reflect
genomic instability
of theforeign DNA in these cell lines. We have initiated work to clone the sites of junction between Adl2 DNA and theadjacent
hamster cellDNA from different Adl2-induced tumor cell lines. In this way, wehope to assess thepossibility of selective sites ofAdl2
DNA
integration
and document itat thenucleotide level. ACKNOWLEDGMENTSWe thank Ingrid Kuhlmann, University of Konstanz, Konstanz, Germany,formakingcell line HT5 available. WeareindebtedtoPetra Bohmfor excellent editorial work.
This research was supported by the Deutsche Forschungsgemein-schaft through SFB274-TP1 and bythe Bundesministerium fur For-schungundTechnologie through GenzentrumKoln,TP2.03.
REFERENCES
1. Doerfler, W. 1982.Uptake,fixation,andexpressionofforeignDNA inmammalian cells: theorganizationofintegratedadenovirus DNA sequences.Curr.Top. Microbiol. Immunol. 101:127-194. 2. Doerfler, W., R.Gahlmann, S. Stabel, R. Deuring, U. Lichtenberg,
M.Schulz, D.Eick, and R.Leisten. 1983. On the mechanism of recombination between adenoviral and cellular DNAs: the struc-ture ofjunction sites. Curr. Top. Microbiol. Immunol. 109:193-228.
3. Gahlmann,R., M.Schulz,and W.Doerfler. 1984. Low molecular weight RNAswithhomologiestocellular DNAatsites of
adeno- A- B-C,D
co as x
T5 4
b
C VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
virus DNAinsertion in hamsteror mousecells.EMBOJ. 3:3263-3269.
4. Groneberg, J., D. Sutter, H. Soboll, and W. Doerfler. 1978. Morphologicalrevertantsof adenovirustype12-transformed ham-stercells.J. Gen.Virol. 40:639-645.
4a.Heuss-Neitzel, D., B. Schmitz, J. Sprengel, C. Zock, and W. Doerfler.Unpublished data.
5. Jessberger, R., D.Heuss,and W.Doerfler.1989.Recombination in hamster cell nuclear extracts between adenovirus type 12 DNA and twohamsterpreinsertion sequences.EMBO J. 8:869-878. 6. Jessberger, R., B. Weisshaar, S. Stabel, and W. Doerfler. 1989.
Arrangement and expression of integrated adenovirus type 12 DNA inthetransformed hamster cellline HA12/7: amplification of viral DNA and evidence for hybrid viral-cellular transcripts. Virus Res. 13:113-128.
7. Koetsier,P., J.Schorr, and W. Doerfler. 1993.Arapid optimized protocol for downward alkaline Southernblotting of DNA. Bio-Techniques 15:260-262.
8. Kotin,R. M., J. C.Menninger, D. C. Ward, and K.I.Berns. 1991. Mapping and direct visualization ofa region-specific viral DNA integration site on chromosome 19ql3-qter. Genomics 10:831-834.
9. Kotin,R.M., R.M.Linden, and K.I. Berns.1992. Characteriza-tion ofapreferred siteonhuman chromosome 19q for integration of adeno-associated virusDNA by non-homologous recombina-tion. EMBO J. 11:5071-5078.
10. Kuhlmann, I., S. Achten, R. Rudolph, and W. Doerfier. 1982. Tumorinduction by human adenovirustype12inhamsters: loss of the viralgenomefrom adenovirustype 12-induced tumorcells is compatible withtumorformation. EMBOJ. 1:79-86.
11. Kuhlmann, I., and W. Doerfler. 1982. Shifts in the extent and patternsofDNA methylation upon explantation and subcultiva-tionof adenovirustype12-induced hamstertumorcells.Virology 118:169-180.
12. Lichtenberg, U., C. Zock, and W. Doerfler. 1987. Insertion of adenovirus type 12 DNA in the vicinity of an intracisternal A particle genomein Syrianhamstertumorcells. J.Virol. 61:2719-2726.
13. McDougall, J. K.,A. R. Dunn, and K. W. Jones. 1972. In situ hybridization of adenovirus RNA and DNA. Nature (London) 236:346-348.
14. Orend, G., I. Kuhlmann, and W. Doerfler. 1991. Spreading of DNAmethylation acrossintegrated foreign(adenovirus type 12)
genomesin mammalian cells. J. Virol. 65:4301-4308. 14a.Orend,G., A.Linkwitz,and W. Doerfler.Unpublished data. 15. Rosahl, T., and W. Doerfler. 1988. Predominant association of
adenovirus type 12 DNA with human chromosome 1 early in productive infection. Virology 162:494-497.
16. Samulski, R. J., X. Zhu, X. Xiao, J. D. Brook, D. E. Housman, N. Epstein, andL.A. Hunter. 1991.Targeted integration of adeno-associated virus (AAV) into human chromosome 19. EMBO J. 10:3941-3950.
17. Schorr, J.,andW.Doerfler. 1993.Non-homologousrecombination between adenovirus and AcNPV DNA fragments in cell-free extracts from insect Spodoptera frugiperda nuclei. Virus Res. 28:153-170.
18. Schulz, M., U.Freisem-Rabien, R. Jessberger, and W. Doerfler. 1987.Transcriptional activities of mammaliangenomes atsites of recombination withforeign DNA. J.Virol.61:344-353.
19. Southern, E. M. 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
20. Sprengel, J., B. Schmitz, D. Heuss-Neitzel, C. Zock, and W. Doerfler.1994.Nucleotidesequenceofhuman adenovirustype12 DNA:comparative functional analysis.J. Virol. 68:379-389. 21. Stabel, S.,and W.Doerfler.1982. Nucleotidesequence atthesite
of junction between adenovirus type 12 DNA and repetitive hamster cellDNAintransformed cell line CLACI. Nucleic Acids Res. 10:8007-8023.
22. Stabel, S., W.Doerfler, and R. R. Friis. 1980.Integration sites of adenovirustype 12 DNAin transformed hamster cells and ham-stertumorcells. J. Virol. 36:22-40.
23. Sutter, D., M. Westphal, and W. Doerfler. 1978. Patterns of integration of viralDNA sequencesin thegenomesof adenovirus type 12-transformed hamster cells. Cell 14:569-585.
24. Tatzelt, J., K. Fechteler, P. Langenbach, and W. Doerfler. 1993. Fractionated nuclearextractsfrom hamster cells catalyze cell-free recombination at selective sequences between adenovirus DNA and a hamster preinsertion site. Proc. Natl. Acad. Sci. USA 90:7356-7360.
25. Tatzelt,J., B.Scholz,K. Fechteler, R.Jessberger, and W. Doerfler. 1992. Recombination between adenovirus type 12 DNA and a hamsterpreinsertionsequenceinacell-freesystem.Patch homol-ogiesandfractionation of nuclearextracts.J.Mol. Biol.
226:117-126.