0022-538X/94/$04.00+0
Copyright©D 1994, American Society for
Microbiology
Mutational
Analysis
of
Delta
Antigen:
Effect
on
Assembly
and
Replication
of
Hepatitis
Delta Virus
MING-FU
CHANG,'*
CHUN-JUNGCHEN,'
ANDSHIN C.CHANG2
Institutesof
Biochemistry'
andMicrobiology,2 College of Medicine, National Taiwan University, Taipei, Taiwan, Republic of ChinaReceived 27 August 1993/Accepted 2 November1993
Hepatitis delta virus requires a helperfunction fromhepatitisBvirus forpackaging, release, andinfection ofhepatocytes. The assemblyof large deltaantigen(HDAg) ismediated bycopackagingwith the small surface antigenofhepatitisBvirus (HBsAg),andtheassemblyof smallHDAg requiresinteractions withlarge HDAg. Toexamine the molecular mechanisms by which smallHBsAg, largeHDAg,and smallHDAg interact,wehave established a virionassemblysystem inCOS7cellsbycotransfecting plasmids encodingthe smallHBsAg,the smallHDAg, and largeHDAgmutants.Resultsindicate that sequences within theC-terminal 19-amino-acid domainflankingtheCxxxisoprenylation motif areimportantfor theassemblyoflarge HDAg.Inaddition, a large HDAg mutant bearing extra sequences separating the C-terminal 19-amino-acid domain from the commonregionsof the small andlargeHDAgsiscapable, likethewild-type largeHDAg,of
copackaging
with small HBsAg. The ability ofassembly is also demonstrated for a large HDAg mutant from which nuclear localization signals have beenremoved. Furthermore, acryptic signal within the N-terminal50 amino acid residues other than the putative N-terminal coiled-coil structure and a subdomain betweenamino acid residues 50 and 65 of the largeHDAg are important for the assembly of small HDAgas well as the trans-dominant negative regulation of largeHDAg inhepatitisdelta virusreplication.Hepatitis delta virus(HDV) isasatellite virus ofhepatitisB virus (HBV). HDV particles contain a negative single-stranded, covalently closed circular RNA genome and virus-specific delta antigens (HDAgs)(1, 2,6, 18, 24, 27, 34,35). The RNA genome possessesextensive intramolecular complemen-tary sequencesthat formanunbranched rod-likestructure(18) and isaribozyme similartoplant viroidRNA(3, 20, 26, 31, 36, 37). The envelope ofHDVis composed of three forms ofHBV surface antigen (HBsAg), L, M, and S, thatare presentinthe ratio of 1:5:95 (2). The S protein (small HBsAg) contains information sufficient for producingemptyparticles ofHDVin thepresenceof HDAgs(28, 32). TheLprotein is essential for infectivity (30), whereas the roles of M protein in HDV multiplication are unknown. In addition, encapsidation of HDVgenomic RNAis mediatedby interactionsbetween the RNA and HDAgs and requires protein-protein interactions betweenHDAgsand small HBsAg (9a).
HDAgsarenuclearphosphoproteins(6, 17) that exist intwo forms. The onlysequence difference between thetwoforms is thepresenceinthelarge HDAg ofanadditional 19-amino-acid domain at the C terminus resulting from a specific point mutation (uridineto cytidine) atthe stop codon of the small HDAg (4, 39, 42). However, they have very distinctive func-tions. ThesmallHDAg iscapable ofsupporting replication of HDV (19), while the large HDAg inhibits the replication of HDVand is essential for virion assembly (5, 9, 28). Recent studiesonthe functional motifs ofHDAgsrevealed that there exist multiple functions within the N-terminal regions of HDAgs. Twoindependent nuclear localization signals (NLS1 andNLS2) importantfor nuclear transport of the HDAgs are located between amino acid residues 35 and 88 (7). The N-terminal 65-amino-acid domain is important for the
inter-*Corresponding author. Mailing address: Institute of Biochemistry,
National Taiwan University College of Medicine, No. 1, Jen-Ai Rd., 1stSection,Taipei, Taiwan, Republic of China. Phone: (02) 397-0800,
ext. 8217. Fax:(02)341-1695.
actions betweenHDAgsboth in vivo and in vitro(7, 8, 33, 40). Furthermore, aputative coiled-coil structure oran unidenti-fiedsignal within the N-terminal 50 amino acid residuesof the large HDAg has been proven to be involved in
negative
regulation of viral replication and incopackaging
with the small HDAg(8,
10, 21,40).
Inaddition,
the Leu-115(8)
and twoarginine-rich
motifs in the middle domain(22)
of the small HDAg arecrucial forRNAbinding andHDVreplication. The isoprenylation motif at the C terminus of large HDAg is involved inHDVassembly(14).
Eventhough themanystudies examining the functional motifs of HDAgs havegreatly
con-tributed to ourunderstanding of the replication and assembly of HDV, the HDV multiplication process has not beenfully
elucidated.Previous studies have demonstrated that the large HDAg could be copackaged with small HBsAg to form virus-like particles in Huh-7 cells. The virus-likeparticles secreted into culture media could be isolated through sucrose
gradient
centrifugation
and observedunderanelectronmicroscope
(5,
28, 32,34).Inthecurrentstudy,wehavefurther established a packagingsystemforHDV inCOS7 cells. Detailed mutational analyses have revealed thatflanking
sequences of the Cxxx isoprenylation motif within the C-terminal 19-amino-acid do-main ofthe large HDAg are important for the assembly of HDV.Moreover,alarge HDAg lacking amino acid residues 50 to 75 failsto inhibit viralreplication
andtocopackage
small HDAg.MATERIALSAND METHODS
Cell lines and DNA transfection. Huh-7 cells
(a
human hepatoma cellline)
and COS7 cells(a
monkeykidney cellline)
were cultured at 37°C in Dulbecco's modified Eagle medium supplemented with 10% fetal calf serum
plus
100 U of penicillin and 100pug
ofstreptomycin perml. DNA transfec-tion was performed with cationic liposomes as previously described(7).
646
on November 9, 2019 by guest
http://jvi.asm.org/
Plasmid construction. (i) Plasmids pECE-d-BE, pECE-d-SM, and pSVD2. Plasmid pECE-d-BE encodes the wild-type large HDAg consisting of 214 amino acid residues, and plas-mid pECE-d-SMencodesthe wild-type small HDAg consisting of 195 amino acid residues. Both plasmids have beendescribed previously (6, 41). For construction of plasmid pSVD2, the 3,397-bpSacl-HindIll DNA fragment of plasmid pD2 (8)was
subcloned into modified pECE vector (13) from which the polylinkersequencesfrom theSacl sitetotheHindlIl site had been deleted. The resultant plasmid, pSVD2, contains a
dimeric HDV cDNA under the control of the simian virus 40 early promoter.
(ii) Plasmid pECE-C-ES. For construction of plasmid pECE-C-ES, plasmid pECE-C (a gift from James Ou, Univer-sity ofSouthern California), which contains monomeric HBV
cDNA,wasdigested with HindlIl and EcoRI and treated with the Klenow fragment of DNA polymerase I before self-ligation. The resultant plasmid, pECE-C-ES,encodesthe small form of HBsAg (Fig. IA).
(iii) PlasmidspECEL-d35/75andpECEL-d89/163. For
con-struction of plasmids pECEL-d35/75 and pECEL-d89/163, plasmid pECE-d-BE containing the BamHI-EcoRI fragment of HDVcDNA was digested withMroI and StuIl plus SmaI, respectively, before self-ligation. The resultant plasmids, pECEL-d35/75 andpECEL-d89/163, encode largeHDAg
mu-tantswith deletions from amino acid residues 35 to 75 and 89
to 163, respectively. Specific mutations in these plasmids and
the following constructs used in this studywere confirmed by
DNAsequencing, using the dideoxychain termination method (29). Positions ofmutations for each correspondingHDAgare
summarized in Fig. 2 and 5A.
(iv)Plasmids pECEL-d35/88and pECEL-CRLDK. For
con-struction of plasmids pECEL-d35/88 and pECEL-CRLDK, plasmid pECE-d-BEwasdigested withMroIplus StuI andSall
plus XbaI,respectively, and treated with the Klenow fragment of DNApolymerase I before self-ligation. The resultant plas-mid, pECEL-d35/88, encodes a large HDAg mutant missing the domain from amino acid residues 35 to 88, and plasmid pECEL-CRLDKencodes alarge HDAgwiththe Cxxx
isopre-nylation motif (14, 25) (211-Cys-Arg-Pro-Gln-214; numbers indicate thepositions of amino acid residues) attheextreme C
terminuschangedto211-Cys-Arg-Leu-Asp-Lys-215 (CRLDK;
Cxxxx).
(v) Plasmid pECEL-aCAT. Plasmid pT7L-aCAT was
cre-ated and used forconstructionof pECEL-aCAT.For
construc-tion ofpT7L-aCAT, plasmidpSV2CAT (15)wasdigested with
StuI plus RsaI. The StuI-RsaI fragment which contains the chloramphenicol acetyltransferase gene was then subcloned
into a modified pT7-d-BP (6) from which the 143-bp
SaclI-Sacl DNA fragment had been deleted and the resultant ends blunted with T4 DNA polymerase. For construction of pECEL-aCAT,plasmid pT7L-aCATwas furtherdigested with
HindIII for subsequent cloning of the HDAg-containing HindIII fragment into the unique HindlIl site of the pECE
vector (13). This resultant plasmid, pECEL-aCAT, encodes a
fusionprotein with the N-terminal 31 amino acid residues of chloramphenicol acetyltransferase linked to amino acid resi-dues 10to 214of the large HDAg.
(vi) Plasmid pECEL-dl96/210.Forconstruction of pECEL-d196/210, plasmid pT7-d-BP (6)wastreated withNcoIand the
Klenowfragmentof DNA polymerase I before SacII digestion. The 559-bp SacII-NcoI DNA fragment was then subcloned
into modified pECE-d-BE from which the 608-bp SacII-SalI DNA fragment had been deleted and the remaining DNA blunted at the Sall site with the Klenow fragment of DNA polymerase I. The resultant plasmid, pECEL-dI96/210,
en-codes a large HDAg mutant in which amino acid residues 196 to 210 have been deleted but the Cxxx isoprenylation motif (211-CRPQ-214) is unchanged.
(vii) Plasmid pECEL-dlO/55. A 495-bp DNA fragment that represents the HDV sequence from nucleotides 720 to 1214 was synthesized by PCR as previously described (8). After further digestion of the PCR productwithSall, a341-bp DNA fragment was generated and subcloned into modified pECE-d-BE from which the 608-bp
SacII-SalI
fragment had been deleted and the remaining DNA blunted at theSacII
site with T4 DNA polymerase. The resultant plasmid, designated pECEL-dlO/55, encodes the large HDAg with an internal deletion from amino acid residues 10 to 55.(viii) PlasmidpECEL-d164/195. For construction of plasmid pECEL-dl64/195, the 365-bp NcoI-HindIII DNA fragment of plasmid pT7-d-BP (6) was blunted at theNcoI site and inserted into a modified pBluescript KS(+) vector (Stratagene) that had been treated withEcoRI plus
HindIll
and blunted at the EcoRI site with the Klenow fragment of DNA polymerase I. The recombinant plasmid, designated pKSL(NH), was further treated withPstI to generate a 374-bp DNA fragment. Follow-ing a blunt-end reaction with T4 DNA polymerase, the 374-bp PstI fragment was further subcloned into modified pECE-d-BE from which the 519-bp SmaI-EcoRI fragment had been de-leted and the remaining DNA blunted at theEcoRI site with the Klenow fragment of DNA polymerase I. The resultant plasmid, pECEL-d164/195, encodes the large HDAg with an internal deletion from amino acid residues 164 to 195.(ix) Plasmids pECEL-d5O/75 and pECEL-d65/75. Recombi-nant plasmids pKSN50Z(SB) and pKSN65Z(SB) were first obtained for further construction of pECEL-d50/75 and pECEL-d65/75, respectively. To construct pKSN50Z(SB), a 125-bpSaclI-BamHIfragment generated from plasmid pN50Z (7) was subcloned into a modified pBluescript KS(+) vector that had been treated with SacII plus BamHI. A similar approach was taken to construct plasmid pKSN65Z(SB) except that the starting plasmid was pN65Z (7) instead of
pN5OZ,
and a 170-bp SacII-BamHI DNA fragment was generated from plasmid pN65Z. BothpKSN5OZ(SB)
and pKSN65Z(SB) were digested with SacII plus SmaI. The SacII-SmaI DNA frag-ments were then individually subcloned into modified pECE-d-BE from which the 317-bp SacII-MroI fragment had been deleted and the remaining DNA blunted at the MroI site with the Klenow fragment of DNA polymerase I to generate plasmids pECEL-d50/75 and pECEL-d65/75, respectively. Plasmid pECEL-d5O/75 encodes the large HDAg with an internal deletion from amino acid residues 50 to 75 whereas plasmid pECEL-d65/75 encodes the large HDAg deleted from amino acid residues 65 to 75. Both of the mutant proteins possess the extra amino acids GDPPR (Gly-Asp-Pro-Pro-Arg) located at the site of deletion, resulting from the processes of plasmid construction.(x) Plasmid pECES-aSR. For construction of plasmid pECES-aSR, plasmid pECE-d-BE was digested with
SalIl,
deleted with exonuclease III
(Exolll)
and mung bean exonu-clease (16), and further digested with XbaI and blunt ended with the Klenow fragment of DNA polymerase I before self-ligation. The resultant plasmid, pECES-aSR, encodes a small HDAg with two extra amino acid residues, Ser-Arg, at the Cterminus.(xi) PlasmidspECEL8-al95/196andpECEL7-al95/198.For construction of plasmids pECEL8-al95/196 and pECEL7-a195/198, intermediate recombinant plasmids were first gener-ated asfollows.Plasmid pT7-d-BP (6) was digested withNcoI and treated with
Exolll
and mung bean exonuclease (16) and then withHindIll. DNA fragments with sizes around 300 bpon November 9, 2019 by guest
http://jvi.asm.org/
wereinserted into modified
pBluescript KS(+)
fromwhich thepolylinker
sequences from the Hindlll site totheEcoRI site had been deleted and blunted at the EcoRI site with the Klenowfragment
of DNApolymerase
I. Recombinant plas-midspKSL8
andpKSL7
containing
the HDV sequencesfrom nucleotides 654 to 1012 and 654 to 1003(24),
respectively,
were
generated.
PlasmidspKSL8
andpKSL7
were further treated withBamHI andthe Klenowfragment
ofDNApoly-merase I before HindlIl
digestion.
The resultantHDV-con-taining
BamHI-HindIIIDNAfragments
werethenindividually
subcloned intomodified
pT7-d-BP
(6)
from which the365-bp
HindIII-NcoI
fragment
had been deleted and theremaining
DNA blunted at the NcoI site with the Klenow
fragment
of DNA
polymerase
I to generate recombinantplasmids
pT7L8-a195/196
andpT7L7-a195/198, respectively.
PlasmidspECEL8-al95/196
andpECEL7-al95/198
were then obtainedby replacing
the SacII-SalIHDV-containing
DNAfragment
in the
plasmid
pECE-d-SM
with their cognatefragments
frompT7L8-a195/196
andpT7L7-a195/198,
respectively.
Plas-midpECEL8-al95/196
encodes alarge HDAg
mutant withanextraamino acid
domain, Trp-Ile-Pro-Arg-Ala-Ala-Gly-Ile
(WIPRAAGI),
inserted between residues 195 and 196. Plas-midpECEL7-al95/198
encodesalarge HDAg
mutantlacking
amino acid residues 196 and 197 andcontaining
an extradomain, WIPRAAG,
inserted between amino acid residues 195 and 198.(xii)
PlasmidspECE-d-BE(pro)
andpECE-d-SM(pro).
PlasmidpECE-d-BE(pro)
has been describedpreviously (8).
It encodesalarge HDAg
withproline
substitutionsatLeu-37 and Ile-41. The strategy used forconstructing pECE-d-SM(pro)
wassimilartothat used forconstructing pECE-d-BE(pro) (8)
except that the mutated
234-bp
Sacll-Stul
fragment
wassubcloned into modified
pECE-d-SM
from which the234-bp
SaclI-Stul
fragment
had been deleted.Harvest of HBV- and HDV-like
particles.
Virus-like parti-cleswerecollectedasdescribedpreviously (38)
with modifica-tions. Inbrief,
culture mediawereharvested 3days
posttrans-fection and clarifiedby spinning
at9,000
rpm in an RA-4F rotor(Kubota)
for 10 min. Thesupernatantwaslayered
over a20%
sucrose cushion(20%
sucrose, 20 mM HEPES[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid] [pH 7.4],
0.1% bovine serum albumin
[BSA])
and thencentrifuged
at40,000
rpm in anSW41
rotor(Beckman)
for 5 h. Virus-likeparticles
secreted into the culture media were spun to the bottom. Thepellet
wasresuspended
inphosphate-buffered
saline
(PBS)
andanalyzed
for the presence ofHBsAg
andHDAgs.
Immunostaining
assay.Immunoblotanalysis
wasperformed
as
previously
described(8).
Inbrief,
whole-cell extracts andprotein lysates
from virus-likeparticles
wereprepared
3days
posttransfection, separated by
sodiumdodecyl
sulfate-polyac-rylamide gel electrophoresis
and electrotransferred onto Im-mobilon-P membrane(Millipore).
Themembranewas immu-nostained withantibodiesspecific
toHDAg
(23)
ortoHBsAg
(Dako).
Indirect immunofluorescence
staining.
Transfected COS7 cells were fixed oncoverslips by
treatment withprecooled
acetone-methanol(4:1)
at -20°C
for2h.Thefixed cellswereincubatedwith 1% BSAat
37°C
for15 minandthen ina37°C moistchamber for30 minwithanantibody specific
toHDAg(23)
which had beenpurified
on immobilizedprotein
Ggel
(Pierce).
Afterincubation,
the cells were washed with PBS. Fluoresceinisothiocyanate-conjugated
goat anti-rabbitimmu-noglobulin
G(Jackson
ImmunoResearchLaboratories, Inc.)
wasadded at a1:40
dilution,
and incubationwascontinuedat37°C
for another 30 min. After asubsequent
treatmentwithEvansblue counterstain solution (Sigma) for 1 min, the cells were washed with PBS, covered with mounting fluid (no. 9; Organon TeknikaCorp.), and photographedunder a fluores-cencemicroscope.
Northern (RNA) blot analysis. Isolation of total cellular RNAand Northern blot analysiswereperformedaspreviously described (8, 11). The probe usedwasthe antigenomicstrand of HDV RNA that was transcribed in vitro from the Sall dimeric HDV cDNA plasmid, pD2 (8), with T7 RNA poly-merase (Promega) in the presence of [cx-32P]UTP. The 28S rRNA detected by hybridization with a 32P-labeled cDNA fragment of 28SrRNA wasusedas an internal control
(8).
RESULTS
Large HDAg could be detected in culture medium when coexpressed with small HBsAg in both nonhepatic and hepatic cells. To elucidate the mechanisms by which HDAgs are involved in the assembly of HDV, we first established a cotransfectionsystemin COS7 cells withplasmid pECE-C-ES (Fig. 1A) that encodes the small HBsAg and plasmid pECE-d-BE that encodes the large HDAg. Two formsof the small HBsAg, p24 and gp27, were detected in culture medium by Western blot
(immunoblot)
analysis with anantibody specific to HBsAg when plasmid pECE-C-ES alone was transfected intoCOS7 cells(Fig. 1B, lane 2).Asimilarresultwasobserved for protein lysate isolated from the transfected COS7 cells (data not shown). Therefore, the small HBsAg could be expressed in COS7 cells and present in both cell lysates and culturemedia. Thisprovides amodel systemforstudyingthe assembly ofHDV.Cotransfection of plasmid pECE-C-ES with pECE-d-BE further demonstrated that the large HDAg ex-pressed in COS7 cells is capable of copackaging with the small HBsAg (Fig. 1C, lane 1). Nevertheless, small HDAg was detected in culture medium only in the presence of large HDAg (Fig. 1C, lanes 2 and 3). Similarpatterns were repro-duced in Huh-7 cells(Fig.
1C,lanes 7 to9).
Functional motifs of thelarge
HDAg essential for virion assemblywere further examined.The Cxxxisoprenylation motif is not the only prerequisite for virion assembly of the large HDAg. A series of mutant constructs for large HDAg were generated (summarized in Fig.
2)
and used in cotransfection experiments with small HBsAg to determine the functional motifs of large HDAg required for virion assembly. Neither the deletionnearthe N terminus encoded by pECEL-d35/88, the deletion nearthe C terminusencodedby pECEL-dl64/195,northe deletion in the middle domain encoded by pECEL-d89/163 affected the ability oflarge HDAgto form virus-likeparticleswith smallHBsAg (Fig. 3A, lanes3 to5), suggesting that the amino acid residues between35 and 195 aredispensable for the assembly of large HDAg. Todetermine whether the N-terminal 35 amino acid residues contribute to the assembly of HDV, we further generatedtwo mutantconstructs,dlO/55 and pECEL-aCAT, that encode large HDAgs with deletions from amino acid residues 10 to55 andof the first10amino acid residues, respectively. In plasmid pECEL-aCAT, the sequences for translation initiationwere derived from the chloramphenicol acetyltransferasegene.Results indicatethat theN-terminal 55 amino acid residues may only contribute to the efficiency of virion assembly of the large HDAg as both HDAg mutants were detected in theculture media (Fig. 3B, lanes 3 and 4). Takentogether, results from mutational analysis suggestthat the 19 amino acid residues at the C terminus of the large HDAgplay amajor role in theassembly ofHDV.Isoprenylation of large HDAgat the 211-CRPQ-214 motif
on November 9, 2019 by guest
http://jvi.asm.org/
A B assembly
(kD)
HBsAg
1 2
29-226 gp27,
p24,
14-Sac I
c
1 2 3 4 5 6 7 8 9
3 45
10 11 12
-L
FIG. 1. Westernblot analysis ofHDAgsfollowing cotransfection of plasmids encoding the small HBsAg and wild-type HDAgs. (A) Structure ofthe recombinant plasmid pECE-C-ES. Plasmid pECE-C-ES encodes the small HBsAg (p24) consistingof 226 amino acid residues. The p24 smallHBsAg canbeglycosylated to form the gp27 product.The solid circlerepresentsthe simianvirus 40 early promoter. The stippled domain represents the complete coding region of the small HBsAg, and the solid boxes represent sequences flankingthe coding region of the small HBsAg. (B) Western blot analysis of HBsAg.Plasmidsused in the transfection studieswere pECE vector (13) (lane1) and pECE-C-ES (lane 2), respectively.HBV-likeparticles were harvested from medium 3 days posttransfection, and protein lysateswereanalyzedwithanantibodyspecifictoHBsAg(Dako).Two formsof the smallHBsAg,p24andgp27,wereidentified.(C)Western blot analysisofHDAgs.Cotransfectionwasperformedwithplasmids pECE-C-ES andpECE-d-BE (lanes1, 4, 7, and10), plasmids pECE-C-ES andpECE-d-SM(lanes 2, 5, 8,and11),orplasmids pECE-C-ES, pECE-d-BE, andpECE-d-SM(lanes 3, 6, 9,and12). Plasmid pECE-d-BEencodes thelarge HDAgwhereasplasmid pECE-d-SMencodes the small HDAg. Protein lysates were prepared either from the virus-likeparticlessecretedinto culture media(lanes 1to3 and 7 to9)
orfrom transfected cells(lanes4to6 and 10to12)andimmunoblotted with an antibodyspecifictoHDAgs. Cell lines usedforthe cotrans-fection studieswereCOS7cells(lanes1to6)and Huh-7 cells(lanes7
to 12). Large (L)andsmall (S) HDAgsareindicated.
has been showntobe
important
fortheassembly ofHDV(14).
However, whether the Cxxx motif issufficient for thiseventhas notbeenstudied. Here,wefurther examined theparticipation
of thesurrounding
sequence in theassembly
ofHDV. Muta-tions that have either the Cxxx(21
1-CRPQ-214)
motif re-placed by Cxxxx (21 1-CRLDK-215, encoded bypECEL-CRLDK)
or a short extension of two unrelated amino acidresidues,
Ser-Arg, at the C terminus of the small HDAg(encoded
bypECES-aSR)
abolished theability
of HDAgto form virus-likeparticles
with small HBsAg as both of the HDAg mutants were notdetected in the culture media (Fig. 3A, lanes7and8).
Itisnoteworthy
that theCys-211
residue in the former mutant isnot located in the fourthposition
from the Cterminus,
whereas the latter mutant lacks both the 211-CRPQ-214isoprenylation
motif and theflanking
sequence1 214
1 195
35 88
pECEL-d35/88 _ ,__
89 163
164 195 10 55
10
210 CRLDK pECELd89/163
pECEL-d164/195 pECEL-dlO/55 pECEL-aCAT
=
pECEL-CRLDK pECELd196/210 pECES-&SR pECEL8-a195/196 pECE-d-BE pECE-d-SM
+
+
+
196 210 195 MSR
195/196
WIPRAAGI
+
195/198
pECEL7-a195/198 +
WIPRAAG
FIG. 2. Structure of large HDAg mutants and their characteristics in the assembly of virus-like particles. Plasmids pECE-d-BE and pECE-d-SM encode thewild-typelarge HDAg consistingof214amino acid residues and smallHDAgconsisting of195 amino acid residues, respectively. Structural representations of individual mutants are shown. Numbers denote thepositionsofaminoacid residuesflanking eachdeletionandinsertion.Specific amino acid residuesthatreplaced the cognateresiduesin thewild-typelargeHDAgor extraamino acid residues that were insertedintowild-typeHDAg are indicated. The abilityof each large HDAgmutant to form virus-like particleswith smallHBsAgis indicatedbyaplusor aminussign.
from amino acid residues 196 to 210. Interestingly, in the presence of small HBsAg, an HDAg mutant (encoded by
pECEL-d196/210)
that possesses the211-CRPQ-214 motif but lacks the 15 amino acid residues from 196 to 210 was not detected inthe culture medium either(Fig.
3A, lane6).
These results strongly suggest that sequences within the C-terminal 19amino acid residuesflanking
the Cxxxmotif oflarge
HDAg areimportant
forvirionassembly.
Toexamine whether conformation of theextreme C-termi-nal
domain
oflarge
HDAg would affect virionassembly,
we constructed two insertion mutants, pECEL8-al95/196 and pECEL7-al95/198. The insertions resulted inseparation
of the C-terminal 19-amino-acid domain of thelarge HDAg
from the common 195amino
acid residues present in both small and large HDAgs. It was found that insertion ofeight
or seven unrelated amino acids between amino acid residues 195 and 196orbetween aminoacid residues 195and198,
respectively,
did notaffectlarge
HDAg incopackaging
withsmallHBsAg
(Fig.
3B, lanes 5 and6).
Takentogether,
theseresults indicate that the Cxxxisoprenylation
motifcoupled
with theon November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.58.295.75.329.2] [image:4.612.315.544.77.411.2]1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
a
1 2 3 4 5 6 7 8 9 10 11 12
FIG. 3. Western blot analysis of large HDAg mutants following cotransfection ofplasmids encoding wild-type small HBsAg and large HDAgmutants.Cotransfection studieswerecarriedoutinCOS7 cells. HDAgs expressedwere thewild-type large HDAg (panel A, lanes1
and9; panel B, lanes 2 and 8), the wild-type small HDAg (panel A, lanes 2 and 10; panel B, lanes 1 and 7), and large HDAgmutants.
Plasmids encoding large HDAg mutants used in this study were
pECEL-d35/88 (panel A, lanes 3 and 11), pECEL-d89/163 (panel A, lanes 4 and12),d164/195 (panel A, lanes 5 and 13), pECEL-d196/210 (panel A, lanes 6 and14), pECEL-CRLDK (panel A, lanes 7 and 15), pECES-aSR (panel A, lanes 8 and 16), pECEL-aCAT (panel B, lanes 3 and 9), pECEL-d10/55 (panel B, lanes 4 and 10),
pECEL8-al95/196 (panel B, lanes 5 and 11), andpECEL7-al95/198
(panelB, lanes 6 and 12). Western blot analysiswasperformedwithan
antibody specifictoHDAgs asdescribed in Materials andMethods. Protein lysates were prepared either from the virus-like particles
secreted into culture media(panel A, lanes 1to8; panel B, lanes1 to
6)orfrom transfected COS7 cells (panel A, lanes9 to 16; panel B, lanes 7to12).Wild-type HDAgs and large HDAgmutantsdetected in the culture mediaareindicatedby asterisks.
ing sequence located within the C-terminal 19 amino acid residues isimportant for the assembly of largeHDAg.
More-over, the 19-amino-acid domain doesnot havetobe immedi-ately adjacent to the sequences common to both small and largeHDAgs for virion assembly.
Alarge HDAgmutantlacking nuclear localization signals is capableofcopackaging withsmallHBsAg. We have previously shown that the large HDAg isa nuclear phosphoprotein (6).
Two nuclearlocalizationsignalsarepresentwithinthe domain from amino acid residue 35 to 88 (7). However, HBsAg is localized in the cytoplasm (12). Therefore, where and how nuclear HDAg and cytoplasmic HBsAg interact have been puzzles. Our previous results showed thatan
HDAg-,B-galac-tosidase fusion protein, N163Z(MS), from which amino acid residues 35 to 88 ofthe HDAg had been deleted, localized exclusively in the cytoplasm of transfected cells (7). Current studiesdemonstrate thatdeletion ofthis domain(pECEL-d35/ 88) has little effect on the assembly oflarge HDAg to form virus-like particles (Fig. 3A, lane 3). Indirect
immunofluores-cence staining shows that the plasmid pECEL-d35/88 does
encode a cytoplasm-localized large HDAg (Fig. 4c). These
findings suggest that the nuclear localization signals of large HDAg are not prerequisites for assembly of virus-like parti-cles.
b
_
FIG. 4. Indirect immunofluorescence
staining
of thelarge
HDAg in transfected COS7 cells. COS7 cells were transfected withpECE
vector
(a), pECE-d-BE (b),
orpECEL-d35/88
(c).
At48h posttrans-fection, cells were fixed andprocessed
for detection ofHDAg
by
indirect immunofluorescence
staining
as described in Materials and Methods. Nuclearstaining
wasobserved for thewild-type large
HDAg encodedby
pECE-d-BE
whereas thepECEL-d35/88-transfected
COS7 cellswerestained in the
cytoplasm.
PlasmidpECE
wasusedas anegative
control.A
signal
within the N-terminal50 amino acid residues anda subdomain between amino acid residues 50 and 65 of the
large HDAg
areinvolved in theassembly
of smallHDAg.
Ithas been shown that thelarge HDAg
isessential for theassembly
of small
HDAg
(10, 28).
Asignal
within thefirst50amino acid residues of thelarge HDAg plays
animportant
role for the event(10, 21). Interestingly, proline
substitutions at Leu-37 and Ile-41 oflarge HDAg
affect the interactions betweenHDAgs
but retain theability
oflarge HDAg
to act as anegative regulator
in HDVreplication
(8).
Here,
we further demonstrate that thelarge HDAg
withproline
substitutions is stillcapable
ofcopackaging
thewild-type
smallHDAg
(Fig.
5C,
lane5).
Moreover,
copackaging
of smallHDAg
couldproceed
whether theproline
substitutions weregenerated
in thelarge HDAg
(Fig.
SC,
lane5),
inthe smallHDAg
(Fig.
SC,
lane
4),
orinboth(Fig. SC,
lane6),
eventhough
theefficiency
may differ between
wild-type
andmutantHDAgs.
Inaddition,
wefound thatthe N-terminal 50-amino-acid domainisnotthe
only
determinant for theassembly
ofsmallHDAg.
Plasmidson November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.65.304.72.278.2] [image:5.612.353.530.78.433.2]M
1
2
3
4
5
A
pECE-d-BE I
35 75
pECEL-d35/75 _
50 75
pECEL-dSO/75 _ _ 6575
pECEL-d65/75
1 2 3 4 5 6 7 8
c
1 2 3 4 5 6
[image:6.612.323.541.72.277.2]_L*
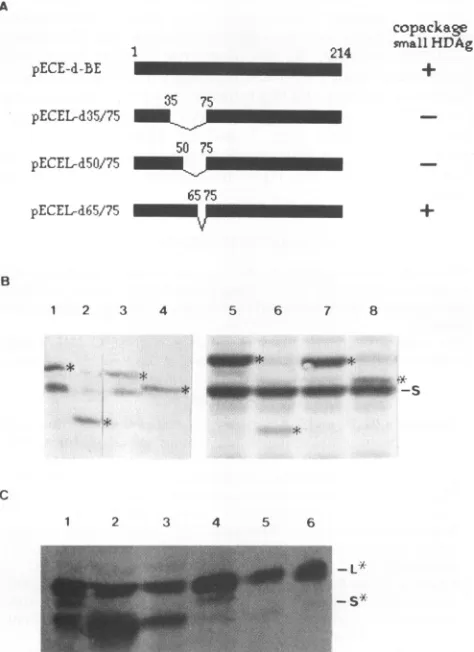
FIG. 5. WesternblotanalysisofHDAgs following cotransfection of plasmids encodingthe smallHBsAgandHDAgmutants.(A) Diagram showingthestructureofwild-typeandmutantlarge HDAgsand their characteristics in virionassembly.Numbers shownon individual
mu-tants denote the positions of amino acid residues flanking each deletion. Theabilityof each large HDAgmutant tocopackage small
HDAgisindicated byaplusorminussign. (B)Western blotanalysis
ofHDAgsfollowing cotransfection withplasmids C-ES, pECE-d-SM, and either the wild type (pECE-d-BE) (lanes 1 and 5) or a mutant construct ofthe large HDAg (lanes 2 to4 and 6 to8).The mutantconstructsoflarge HDAg usedwerepECEL-d35/75 (lanes2 and 6),pECEL-d65/75 (lanes 3 and 7),and pECEL-d50/75 (lanes4 and8).Western blotanalysiswasperformedwith theantibody specific toHDAgsasdescribed in Materials and Methods. Proteinlysateswere
prepared either from the virus-like particles secreted into culture media (lanes I to4)orfrom transfected COS7 cells (lanes5 to8).
Wild-type large HDAg and its mutated forms are indicated by
asterisks. S denotes thewild-type smallHDAg. Twolighter signalsin lane 2 representing wild-type small and large HDAgsresulted from
overflowingofasamplefrom thenextlane. (C)Western blotanalysis
ofHDAgswithprolinesubstitutionsatLeu-37 and Ile-41. Plasmidsof
HDAgsusedwerepECE-d-BEandpECE-d-SM(pro) (lanes1 and4),
pECE-d-BE(pro) and pECE-d-SM (lanes 2 and 5), and pECE-d-BE(pro) and pECE-d-SM(pro) (lanes 3 and 6). Plasmids pECE-d-BE(pro) and pECE-d-SM(pro) encode large and small HDAgs,
re-spectively, with proline substitutions at Leu-37 and Ile-41. Protein
lysateswerepreparedfrom either transfectedCOS7 cells(lanes1to3)
orvirus-likeparticles secreted into culture media (lanes4 to 6).L*
denotes thewild-type and proline-substituted large HDAgs. S* de-notesthewild-type andproline-substitutedsmallHDAgs.
FIG. 6. Northern blot analysis of HDV RNA following cotransfec-tion of Huh-7 cells with dimericHDV cDNAandaplasmid encoding
large HDAg.Plasmids usedinthe cotransfection studiesrepresentthe dimericHDVcDNAonly (lane 1) and thedimeric HDV cDNAplus
either thewild-type large HDAg (lane 2)orlarge HDAgmutantsthat
weredeleted between amino acid residues65 and 75 (lane3), 50 and
75(lane 4),or35and75 (lane 5).Theprobe usedwastheantigenomic
strand of HDV RNAandwaspreparedasdescribedinMaterialsand
Methods. Invitrotranscripts representing the 1.7-kb monomeric and
3.4-kb dimeric HDV genomic RNAs hybridized to the antigenomic RNAprobewereusedasmarkers (M). 28Srepresentsthe 28S rRNA
as an internal control.
pECEL-d35/75, pECEL-d5O/75, and pECEL-d65/75, which encode mutant large HDAgs with deletions from amino acid residues 35to75,50to75, and 65to75,respectively,wereused
incotransfectionexperiments. Data show that the small HDAg could be copackaged and secreted into culture media only whencotransfectionwascarriedoutwith the plasmid pECEL-d65/75 (Fig. 5B, lane 3), butnot theothertwo plasmids (Fig. SB, lanes2 and4). Takentogether, these results indicate that
asubdomain from amino acid residues 50 to 65 aswell as a
signal within the N-terminal 50 amino acid residues of large HDAg may be important for the interactions between small and large HDAgs and is involved in virion assembly of the smallHDAg.
Alarge HDAgmutantlackingaminoacid residues 50to75 fails torepress thereplication of HDV.To learn whether the subdomain from amino acid residues 50 to 65, which is important for the assembly of HDV, also contributes to the negative effect of large HDAg on HDV replication, we
co-transfected Huh-7 cellswith the plasmid pSVD2, which
con-tains dimericHDVcDNA, andaplasmid that encodeseither thewild-typeoramutatedlarge HDAg.Aswehavepreviously shown(8),thedimeric cDNA alone couldundergo replication (Fig. 6, lane 1). Interestingly, wefound thatlarge HDAgwith adeletion of amino acidresidues 50to75 failedtorepressthe replication of HDV (Fig. 6, lane 4) whereas a deletion from
amino acid residues65to75 retainedinhibitory activity (Fig. 6, lane 3). These findings indicate that the subdomain between
amino acid residues50 and 65 of the large HDAgisinvolved
in the negative regulation of HDV replication. copackage
214
small
HDAg
U
+3.4-
1.7-B
+
28S-0
#Olb* --m^Aw* qmlp*
4m ANIML4s
44%%bb.* -VOWOO#*
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.54.291.96.422.2]DISCUSSION
In this study, we have examined subdomains of the large HDAg
involved
in theassembly of
HDV-likeparticles.
De-tailed mutationalanalyses
demonstrate thatthe
C-terminal 19-amino-acid domaincontaining
theisoprenylation
motif isimportant
for theassembly
oflarge HDAg. Furthermore,
a subdomain from amino acid residues 50 to 65 and asignal
within the N-terminal 50 aminoacid residues
of thelarge
HDAgareinvolved in theassembly
of smallHDAgaswellas the trans-dominantnegative regulation
ofHDVreplication.
The process by which nuclear HDAg
(6)
andcytoplasmic
HBsAg(12)
meet toform HDVparticles
is unclear. An earlierstudy
correlated theassembly
ofHDVwithisoprenylation
of large HDAg at the 211-Cxxx-214 motif(14).
A serine substi-tutionatCys-211 abolished
theassembly
oflarge HDAg
(14).
Therefore,
itis possible
thatisoprenylation of large
HDAg results in membraneanchoring
which,
in turn,facilitates
the interactions betweenlarge HDAg
and smallHBsAg
on the membrane of theendoplasmic
reticulum.Interestingly,
we found that alarge HDAg
from which the nuclear localizationsignals
have beendeleted iscapable
ofcopackaging
with smallHBsAg
(Fig. 3A,
lane3).
HDAgsarenuclearphosphoproteins
(6,
17).
The role of nuclear localizationsignals
in virionassembly
is unclear.However, nuclear localization ofwild-type
large HDAg is likely to be involved in the assembly of both smallHDAg and HDVgenomic
RNAlocatedin the nucleus in order to form infectious HDVparticles.The C-terminal 15 amino acid residues
adjacent
to the 211-Cxxx-214isoprenylation
motifareinvolved in theassembly
oflarge HDAg.
Alarge HDAg
mutant, encodedby
pECEL-d196/210,
that retains the Cxxxisoprenylation
motif but lacks itsflanking
amino acid residues from 196 to 210 failed toundergo assembly (Fig.
3A, lane6).
Theeffect of the 15-amino-acid deletiononassembly may be dueto adisruption
of directinteractions
between thelarge HDAg and small HBsAgordue to aninfluenceonisoprenylation. Alternatively, the conforma-tion of theflanking
sequencestogether with theisoprenylation
motif may becritical forassembly.
However, the latter hypoth-esis isnotsupported by
ourresultsshowing
thatalarge HDAg
mutant in which the extreme C-terminal 19-amino-acid do-mainwas
separated
from thecommonN-terminal 195-amino-aciddomainby
an8-amino-acid
peptide
retained theability
to form virus-likeparticles
(Fig.
3B,
lane5).
Inaddition,
alarge
HDAg
mutant in which a seven-amino-acidpeptide
was in-sertedat thesameposition
and thewild-type
amino acids 196 and 197weredeleted could bedetected in the culture medium(Fig.
3B,
lane6). Furthermore,
alarge
HDAgmutantin which the C-terminalPro/Gly-rich region
betweenamino
acid resi-dues 164 and 195wasdeleted stillretained theability
toform virus-likeparticles
(Fig.
3A, lane5). Therefore,
we propose that theisoprenylation
motiftogether
with theflanking
amino acid residues from 198to210 isimportant
for theassembly
oflarge
HDAg and may be involved in the interactions betweenlarge
HDAg and small HBsAg. The question of whether cellularproteins
areinvolved in the interactions is unanswered.Protein-protein
interactionsbetween the HDAgs have beenhypothesized
toplayakey role in virion assembly as well as thenegative
regulation ofHDVreplication (8, 10, 21, 40). These studies concluded that a putative coiled-coil structure or an unidentifiedsignal
located betweenamino acid residues 13 and 48 isimportant
for both events. Our present study on theassembly
of smallHDAg further suggests that a cryptic signal within the N-terminal 50 amino acid residues other than the N-terminal coiled-coilstructureisresponsible for the interac-tions betweenHDAgs; small HDAgwascapable ofcopackag-ing with a large HDAg containing proline substitutions at Leu-37 andIle-41,eventhoughtheefficiency maybedifferent
(Fig.
SC).
Aspredicted by
computeranalysis (PepPlot
ofGenetics
Computer Group programs), proline
substitutions break the a.-helix and are likely to destroy the N-terminal coiled-coil structure. Results from the presentstudy
also suggest that a subdomain located between amino acid residues 50 and 65 ofthelarge
HDAg
is involved in the virionassembly
of small
HDAg
and thenegative
effect oflarge HDAg
onthereplication
of HDV(Fig.
5 and6). However,
the exact conformation of the N-terminal 65 amino acid residues of HDAgs involved in the HDV multiplication remains to be elucidated.ACKNOWLEDGMENTS
Wethank J.-C.Chengand C.-W. Tsai fortechnical assistance.We
aregratefultoJames Ouforproviding plasmid pECE-CandtoJeffrey Yenand SteveRoffierforhelpfulcommentsand criticalreadingofthe manuscript.
This workwassupported byaresearchgrant (NSC82-0419-B-002-438-MB) from the National Science Council of theRepublicof China.
REFERENCES
1. Bergmann, K.F.,andJ.L.Gerin. 1986.Antigensofhepatitisdelta virus in the liver andserumof humans andanimals.J.Infect. Dis. 154:702-705.
2. Bonino, F.,K. H.Heermann,M.Rizzetto,and W. H. Gerlich.1986. Hepatitisdelta virus:protein compositionof deltaantigenandits hepatitisBvirus-derivedenvelope.J.Virol.58:945-950. 3. Branch,A.D.,and H. D. Robertson.1991.Efficienttranscleavage
and a common structural motif for theribozymes of thehuman hepatitis 8agent. Proc.Natl. Acad. Sci. USA 88:10163-10167. 4. Casey, J. L.,K. F.Bergmann,T. L.Brown,andJ.L.Gerin.1992.
Structural requirements for RNA editing in hepatitis 8 virus: evidence forauridine-to-cytidineeditingmechanism.Proc. Natl. Acad. Sci.USA 89:7149-7153.
5. Chang, F.-L., P.-J. Chen,S.-J. Tu, C.-J. Wang,andD.-S. Chen. 1991. The large form of hepatitis delta antigen is crucial for assembly of hepatitis delta virus. Proc. Natl. Acad. Sci. USA 88:8490-8494.
6. Chang, M.-F.,S. C.Baker,L. H.Soe,T.Kamahora, J.G.Keck,S. Makino, S. Govindarajan, and M. M. C. Lai. 1988. Human hepatitis delta antigen is a nuclearphosphoprotein with RNA-binding activity.J.Virol.62:2403-2410.
7. Chang, M.-F.,S. C.Chang,C.-I.Chang,K.Wu,and H.-Y.Kang. 1992. Nuclearlocalizationsignals,butnotputativeleucinezipper motifs, are essential for nuclear transport of hepatitis delta antigen.J.Virol.66:6019-6027.
8. Chang, M.-F., C.-Y. Sun, C.-J. Chen, and S. C. Chang. 1993. Functional motifsof deltaantigen essentialfor RNAbindingand replicationofhepatitisdeltavirus.J.Virol. 67:2529-2536. 9. Chao, M.,S.-Y.Hsieh,andJ.Taylor. 1990. Role oftwoforms of
hepatitis delta virus antigen: evidence for a mechanism of self-limitinggenomereplication.J.Virol. 64:5066-5069.
9a.Chen, C.-H.,and M.-F.Chang.Unpublishedobservation. 10. Chen, P.-J.,F.-L.Chang, C.-J. Wang, C.-J. Lin,S.-Y.Sung,and
D.-S. Chen. 1992. Functional study ofhepatitisdeltaviruslarge antigeninpackagingandreplicationinhibition:role of the amino-terminal leucinezipper.J.Virol. 66:2853-2859.
11. Chomczynski, P.,and N.Sacchi. 1987.Single-step method ofRNA isolation by acid guanidium thiocyanate-phenol-chloroform
ex-traction. Anal.Biochem. 162:152-159.
12. Eble,B.E.,D. R.Macrae,V. R.Lingappa,and D.Ganem.1987. Multiple topogenicsequencesdetermine the transmembrane ori-entation ofhepatitisBsurfaceantigen. Mol. Cell.Biol.7:3591-3601. 13. Ellis, L., E. Clauser, D.0. Morgan, M. Edery, R. A. Roth, and W. J. Rutter. 1986. Replacement of insulin receptor tyrosine residues 1162 and 1163 compromises insulin-stimulated kinase activityanduptakeof2-deoxyglucose. Cell 45:721-732.
14. Glenn, J. S., J.A.Watson,C. M.Havel, andJ.M.White. 1992.
on November 9, 2019 by guest
http://jvi.asm.org/
Identification of a prenylation site in delta virus large antigen. Science 256:1331-1333.
15. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombi-nantgenomes which expresschloramphenicol acetyltransferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
16. Henikoff, S. 1984. Unidirectional digestion with exonuclease III createstargeted breakpoints for DNA sequencing. Gene 28:351-359.
17. Hwang, S., C. Z. Lee, and M. M. C. Lai. 1992. Hepatitis delta antigen expressed by recombinant baculoviruses: comparison of biochemical properties and post-translational modifications be-tweenthelarge and small forms. Virology 190:413-422. 18. Kos, A., R. Dijkema, A. C. Arnberg, P. H. van der Merde,and H.
Schelekens. 1986. The HDV possesses a circular RNA. Nature (London)323:558-560.
19. Kuo, M. Y.-P., M. Chao, and J. Taylor. 1989. Initiation of replication of the human hepatitis delta virus genome from cloned DNA:role of deltaantigen. J. Virol. 63:1945-1950.
20. Kuo, M. Y.-P., L. Sharmeen, G. Dinter-Gottlieb, and J. Taylor. 1988. Characterization of self-cleaving RNA sequences on the genome andantigenomeof humanhepatitisdelta virus. J.Virol. 62:4439-4444.
21. Lazinski, D. W., and J. M. Taylor. 1993. Relating structure to function in thehepatitisdeltavirusantigen.J.Virol. 67:2672-2680. 22. Lee, C.-Z., J.-H. Lin, M. Chao, K.McKnight, and M. M. C. Lai. 1993.RNA-binding activity of hepatitis deltaantigeninvolves two arginine-richmotifs and isrequired for hepatitisdeltavirusRNA replication.J.Virol. 67:2221-2227.
23. Lin, J.-H., M.-F. Chang, S. C. Baker, S. Govindarajan, and M. M. C. Lai. 1990. Characterization ofhepatitis delta antigen: specificbinding to hepatitis delta virus RNA. J.Virol. 64:4051-4058.
24. Makino, S., M.-F. Chang, C.-K. Shieh, T. Kamahora, D. M. Vannier, S. Govindarajan, and M. M. C. Lai. 1987. Molecular cloning and sequencing of human hepatitis delta virus RNA. Nature (London)329:343-346.
25. Maltese, W.A.1990.Posttranslational modificationofproteins by isoprenoids inmammalian cells. FASEB J. 4:3319-3328. 26. Perrotta, A. T., and M. D.Been.1991. Apseudoknot-likestructure
required for efficient self-cleavage ofhepatitis delta virus RNA. Nature (London)350:434-436.
27. Rizzetto, M.,B.Hoyer,M.G.Canese, J.W.-K.Shih,R. H.Purcell, and J.L.Gerin. 1980. Delta agent: association of delta antigen with hepatitis B surface antigen and RNA in serum of delta-infectedchimpanzees. Proc. Natl. Acad. Sci.USA77:6124-6128. 28. Ryu, W.-S., M. Bayer, andJ. Taylor. 1992.Assemblyofhepatitis
delta virusparticles.J.Virol.66:2310-2315.
29. Sanger, F., S.Nicklen, andA. R.Coulson.1977. DNAsequencing
with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
30. Sureau, C., B. Guerra, and R. E. Lanford. 1993. Role of the large hepatitis B virus envelope protein in infectivity of the hepatitis delta virion. J. Virol. 67:366-372.
31. Uhlenbeck, 0. C. 1987. A small catalytic oligoribonucleotide. Nature(London)328:596-600.
32. Wang, C.-J., P.-J. Chen, J.-C. Wu, D. Patel, and D.-S. Chen. 1991. Small-formhepatitisBsurface antigen issufficienttohelp in the assemblyof hepatitis delta virus-like particles. J. Virol. 65:6630-6636.
33. Wang, J.-G., and S. M. Lemon. 1993.Hepatitisdelta virusantigen forms dimers and multimeric complexes in vivo. J. Virol. 67:446-454.
34. Wang, K.-S., Q.-L. Choo, A.-J. Weiner, J.-H. Ou, R. C. Najarian, R.M. Thayer,G. T. Mullenbach, K. J. Denniston, J. L. Gerin, and M. Houghton. 1986. Structure, sequence and expression of the hepatitisdelta virus genome. Nature (London)323:508-513. 35. Weiner, A. J., Q.-L. Choo, K.-S. Wang, S. Govindarajan, A. G.
Redeker, J. L. Gerin, and M. Houghton. 1988. A single anti-genomicopenreading frame ofthehepatitis delta virusencodes the epitope(s) of both hepatitis delta antigen polypeptides p248 andp278.J.Virol. 62:594-599.
36. Wu, H.-N., and Z.-S. Huang. 1992. Mutagenesis analysis of the self-cleavage domain ofhepatitis deltavirus antigenomic RNA. NucleicAcids Res. 20:5937-5941.
37. Wu, H.-N., Y.-J. Lin, F.-P. Lin, S. Makino, M.-F. Chang, and M. M. C. Lai. 1989. Humanhepatitis delta virus RNA subfrag-mentscontain an autocleavageactivity.Proc.Natl. Acad.Sci.USA 86:1831-1835.
38. Wu, J.-C.,P.-J. Chen,M. Y.-P.Kuo, S.-D.Lee,D.-S. Chen,and L.-P. Ting. 1991.Productionofhepatitisdeltavirusand suppres-sion ofhelper B virus in a human hepatoma cell line. J. Virol. 65:1099-1104.
39. Xia, Y.-P., M.-F. Chang, D.Wei, S. Govindarajan,and M. M.C. Lai. 1990. Heterogeneity of hepatitis delta antigen. Virology 178:331-336.
40. Xia, Y.-P., andM. M.C. Lai. 1992. Oligomerizationofhepatitis delta antigen is required for both thetrans-activating and
trans-dominant inhibitory activities of the delta antigen. J. Virol. 66:6641-6648.
41. Xia, Y.-P., C.-T. Yeh, J.-H.Ou,and M.M. C.Lai. 1992. Charac-terization of nuclear targeting signal of hepatitis delta antigen: nuclear transportas aprotein complex.J.Virol. 66:914-921. 42. Zheng, H.,T.-B.Fu, D.Lazinski, and J. Taylor. 1992.Editingon
the genomic RNA of human hepatitis delta virus. J. Virol. 66:4693-4697.