JOURNALOFVIROLOGY, Jan. 1994,P. 34-47 0022-538X/94/$04.00+0
Copyright © 1994, American Society for Microbiology
Requirement for
Double-Strand
Breaks but Not for
Specific
DNA Sequences in Herpes Simplex Virus Type
1
Genome
Isomerization
Events
ROBERT T. SARISKY12AND PETER C. WEBER2*
Departmentof Microbiology and Immunology2 and GraduateProgram in Genetics,1 The Milton S. Hershey
Medical Center, ThePennsylvania State University College of Medicine, Hershey,Pennsylvania 17033 Received 16 June 1993/Accepted 22September1993
Herpes simplex virustype 1(HSV-1) genomeisomerizationoccursas aresultofDNAreplication-mediated
homologous recombination between several sets of inverted repeat sequences present in the viral DNA.The frequency with which this recombinationoccurshasbeendemonstratedtobedependentuponDNAhomology length rather than specificsequences.However, the smallestof the viral invertedrepeats,thea sequence,has been showntofunctionas arecombinational hotspot,leadingtospeculationthatthissequencemayrepresent
a specific element through whichgenomeisomerization ismediated. To investigate this apparent paradox,a
quantitative transient recombination assay system was developed and used to examine therecombinogenic properties ofapanel ofasequencemutants.Thisanalysis revealed that thepresenceof both thepaclandpac2 elementswas bothnecessaryand sufficient for the induction ofhigh-frequency recombinationeventsby thea sequence.However,itwasthedouble-strand break promoted by pacl and pac2 during cleavage and packaging
atthea sequence, andnot the DNAsequences of the elements themselves, whichappearedtobecritical for recombination. Thiswas illustrated (i) by the inability of the same pacl and pac2 sequences to mediate inversioneventsincellsinfected withanHSV-1mutantwhichwascompetentforDNAreplication-dependent recombination but defective for the cleavage and packaging process and (ii) by the ability of double-strand
breaksgeneratedinnon-HSV-1 DNAbyaninvivo-expressed restriction endonucleasetosignificantly stimulate theinitiation ofrecombinationeventsinvirus-infected cells. Thus, theasequence appearstoactasahotspot
forhomologous recombination simply because it happenstocoincide with the site of the double-strandbreak which isgenerated during the cleavage and packagingprocess,notbecauseit contains discretesequenceswhich are required for this activity. However, itwasfound that this enhancedrecombinogenicity disappeared when
the element was flanked by regions of extensive sequence homology, particularly that of the large inverted
repeatswhich flanktheasequenceatits natural site in theHSV-1genome.Thesefindingsareconsistent with amodel for HSV-1 genome isomerization inwhich recombination is initiatedprimarily by multiple random double-strand breaks which ariseduring DNAreplicationacross theinverted repeats of the genome, rather thanbya single specific break whichoccursatthea sequenceduring the cleavage and packagingprocess.
The herpes simplex virustype 1(HSV-1)genomeisa152-kb
linear double-stranded DNA molecule divided into two seg-ments, a 108-kb long (L) component and a 13-kb short (S)
component.TheL and Ssegmentsareflanked by theinverted
repeat sequences b (9 kb) and c (6.5 kb), respectively. The genomeitself isbracketedateither endbyoneormorecopies
ofadirectrepeat,theasequence(250to500 bp), and contains
onetoseveralinvertedcopies of theasequenceseparating the L and S components. The HSV-1 genome can therefore be
represented by the following arrangement (9, 26, 38): ab---L---b'a'c'---S---ca
Four equimolar isomers of viral DNA exist during produc-tive HSV-1 infection, since the L and S components invert relative to each other at a high frequency (7, 9). These
inversions were originally postulated to be the result of a
site-specific recombination eventoccurringat thea sequence, on the basis of two lines of evidence. First, deletion of the entire internal L-S junction, which includes the a sequence,
*Correspondingauthor. Mailingaddress:Parke-Davis
Pharmaceu-ticalResearch, Infectious Diseases Section,2800 Plymouth Road, Ann
Arbor,MI48105. Phone: (313) 998-3295. Fax: (313) 998-3318. Elec-tronic mail address: [email protected].
from the viral genome resulted in the appearance of
nonin-verting or"frozen" genomeswhich represented only asingle
isomeric form(10, 11, 21). Second, theduplication of genomic segments containing the a sequence, but not other unique HSV-1 sequences within the viral DNA, resulted in novel inversion events in the genome (4, 14, 15, 17). Since these
initial studiessuggested that only thea sequence wascapable
ofmediating inversion events in the HSV-1 genome, it was
concluded that the a sequence contained a signal for
site-specific recombination which initiated thisisomerization pro-cess. However, later evidence has indicated that HSV-1 ge-nomeisomerizationcanresult fromgeneralized recombination
between any of the inverted repeat sequences flanking theL andScomponents.Forexample,amutantvirus which had lost two-thirds of thebsequenceand theentirea andcsequences
of theL-Sjunctionwas still capable of undergoing L compo-nent inversion during productive infection (12). It was also
found that otherHSV-1sequenceswhich lackhomologytothe
a sequence promoted similar recombination events when duplicated in the viral genome (10, 11, 20-22, 37). Finally,
when sequences of theprokaryotic transposable element Tn5
were incorporated into the viral genome, theywere foundto undergo inversionina mannerwhichwasentirely analogousto
Land Scomponent inversion (39). This novel rearrangement event, which is typically not observed in the transposon's 34
Vol. 68, No. 1
on November 9, 2019 by guest
http://jvi.asm.org/
MECHANISM OF HSV-1 GENOME ISOMERIZATION 35
HSV-I (F) a sequence:
Ap
Ss Sm Sm AP
TV VV DRI paCI \
Ss Ss Ss TV V
DR2 DR4
Ap
Ss Sm TV
EJ=
pac2
DR1
440 440A1
440A2
440A8
440A10 a
440A11 { 440A12 440A13 440A14 474orAFsp
Wt
I
NON-HSV-1 SEQUENCE INSERT(GALA ORISS0) NOaSEQUENCE INSERT
ED EJ]
EJi D3 D3
pLR
recombination index 1.0 2.0 3.0
1
pLAR
recombination index 1.0 2.0 3.0 4.0
U I I
I I I
U
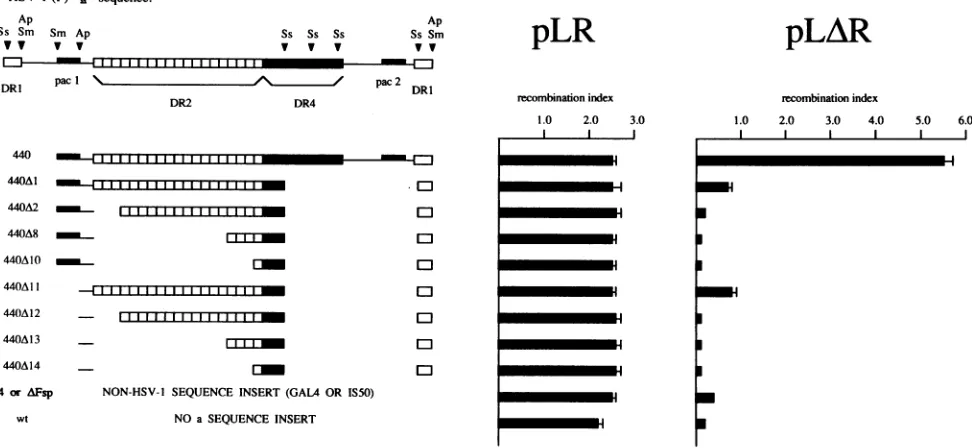
FIG. 1. Construction of HSV-1asequencederivatives containingdeletions in the DR2repeatarrayand quantitation of their recombinogenic
potential. At leftaresubfragments of the HSV-1asequencecontaining anisomorphicandnonanisomorphic DR2repeatarrays.440representsa
wild-typeasequencederivativewhichpossessesboth pacl and pac2 elementsaswellas acomplete DR2array.The eight deletion derivatives of
440 shownhave lostone orbothpacelements from variouswild-typeormutantversions of the DR2repeatarray.474 and AFsparetwonon-HSV-1
DNA insertswhicharecomposed of 474 bp of theyeastgeneGAL4 and600 bp ofTn5sequences,respectively.wt,wild-type plasmid withoutan asequence insert.Restriction sitesareshownfor Apal (Ap), SmaI (Sm), and SstII (Ss). At right the relative contributions of theseasequence
subfragmentstoTn5inversioninaplasmid transfection-superinfectionassay aresummarized. The recombination index (with associated standard
deviation) correspondstothe ratio ofinversion frequency in theasequence-containingTnSplasmid pLRorpLARtothe deletion frequencyin
the internal standardplasmid pGAL-STD. Each plasmid transfectionwasperformed in triplicate.
natural host, Escherichia coli, was shown to be caused by
high-frequencyhomologous recombination between the dupli-cated inverted IS50 elements of TnS. Furthermore, theseven
enzymeswhich constitute the HSV-1 DNAreplication
machin-ery werefound todirect these recombination events indepen-dent ofspecific DNAsequence requirements (39).
These results indicate that any inverted repeat homology should be sufficient for recombination to occurinthe HSV-1 genome. However, it remains unclear how the a sequence, which is apparently sufficient but not required for genome
isomerization, canactas arecombinationalhotspotby direct-ing inversion events over great distances within the viral genome(4, 27,37).Theasequencesof different HSV-1strains, although highly variable, contain a number of common fea-tures (5, 13, 37). These include a complex of short tandem
guanosine-cytosine-rich reiterations (the DR2 repeat array), two terminal reiterations (the DR1 repeats), and two sets of unique sequences (Ub and
Uj)
possessing highly conserved signals (pacl and pac2, respectively) that direct both the cleavageof concatameric viral DNA into unit-lengthgenomes and its packaging into capsids (6, 37) (Fig. 1). Additional strain-specificreiterations of theasequencewhichmaysome-times be found between the DR2 repeat array and the
Uc
sequences include the DR4 repeats of HSV-1 strain F (13) (Fig. 1) and the DR3.5 repeats ofHSV-1 strainJustin(16).
Two of these features of thea sequence, the DR2 repeat
array and the pac elements, may enhance the frequency of homologous recombination events in the viral genome by servingassites for theinitiationofrecombination. First,when
multiple copies of tandem DR2 repeats are present in a
negatively supercoiled plasmid, theymayadoptanovel
second-arystructuretermedanisomorphic DNA (42). This conforma-tion probably contains single-stranded regions as a conse-quenceofsupercoil-induced relaxation, since thesesequences
have been showntobe highly sensitive to cleavage by single-stranded nucleases.Moreover,aconformation-specific butnot sequence-specific nuclease present in both mock- and HSV-infected cells has beenshowntopreferentially cleave the DR2 repeatarrayin vitro(41). Although the role that this nuclease
plays in recombination in HSV-1 hasnotyetbeendetermined, theseproperties of the DR2 arrayindicate that itmay poten-tiallyact as a site for the initiation of recombination events. Alternatively, double-strand breaks which are created during
cleavage and packaging(cleavage/packaging)eventspromoted by the pacl and pac2 elements (6, 19, 37) may serve to stimulate thefrequencyofhomologousrecombination events. Smiley et al. (27) have provided compelling evidence in supportof this ideaby demonstrating adramatic decrease in the frequency ofa sequence-mediated recombination events uponthe deletion of eitherpacelement.
Thus, nicks withinan anisomorphic secondary structureor
double-strand breaks resulting from the genome cleavage eventmayindividuallyortogethercontribute tothe observed enhanced recombination whichoccursat thea sequence. The potential role that these featuresplayinstimulating recombi-nationevents attheasequence wasexamined in this work. The previously describedabilityof transfected plasmid-borne TnS
sequences to undergo inversion in HSV-1-infected cells (39)
was developed into a convenient system for quantitatively
analyzing the relative contributions of specific a sequence
elements to enhanced recombinogenicity. This approach in-volved inserting wild-type and mutant derivatives of a se-5.0 6.0
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.67.553.82.306.2]36 SARISKY AND WEBER
tandem HSV-1 (KOS) a sequences:
Ss Sm DrSm Mnl Ss Ss Sm DrSm Mnl Ss Ss Sm
VT TV 1IIIIIIIIIIT TV TV llIllIllIlY VT
DRI pacI pac2 pac l. pac2DRI
DR2 DR4 t DR2 DR4
cleavage-packaging site
4
193 _ : { 0
193A1 *
193A2 ---ra -i
193A3 193A4 *
474orAFsp NON-HSV-IINSERT(GALAOR IS50)
wt NOaSEQUENCE INSERT
pLR
recombination index 1.0 2.0 3.0
pLAR
recombination index 1.0 2.0 3.0 4.0
U
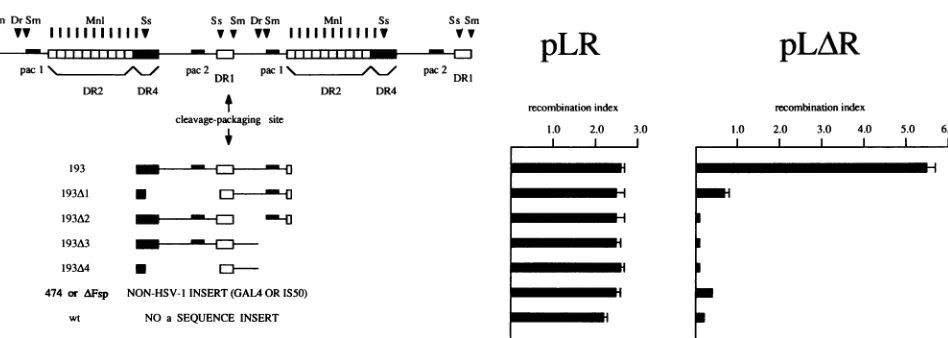
FIG. 2. Construction of HSV-1asequencederivatives whicharedefectiveforcleavageandpackagingandquantitationof theirrecombinogenic potential. At leftarecleavage/packaging-proficientandcleavage/packaging-deficient subfragmentsofatandem HSV-1 asequencejunction. 193 possessesboth the pacl and pac2 elements andrepresentsthe minimalsignal sufficientformediating cleavageandpackagingofviral DNA.The four deletion derivatives of 193 shown lackone orboth of thepaclorpac2elements. 474 and zFsparetwonon-HSV-1 DNA inserts whichare
composed of474bp of theyeastgeneGAL4 and 600bpof theprokaryotictransposonTn5sequences,respectively.wt,wild-type plasmidwithout anasequenceinsert.Restrictionsitesareshown for Dral(Dr),MnlI(Mnl), SmaI (Sm),and SstII(Ss).Atrightthe relative contributionsofthese
a sequence subfragments to Tn5 inversion in a plasmid transfection-superinfection assay are summarized. The recombination index (with
associated standarddeviation) corresponds tothe ratio of inversionfrequencyin theasequence-containingTn5plasmid pLRorpLARtothe
deletion frequency in the internal standard plasmid pGAL-STD. Each plasmidtransfectionwasperformedintriplicate.
quence subfragments into both IS50 repeatsofTnS andthen determining the relative increases in TnSinversion frequency
that theyconferred. The presence of the cleavage/packaging-proficient pac2-DR1-pacl arrangement wasfound tobe both
necessary and sufficient for the enhanced recombinogenic
properties of thea sequence,while the abilitytoform
aniso-morphicDNA intheDR2repeatarrayhad little influenceon
recombination frequency. The requirement for a
double-strand break createdduringthecleavage/packagingprocessin
inducingHSV-1 recombinationwasdramaticallyconfirmedby
the demonstration that a restriction endonuclease-generated
break in anonviral DNA sequence hadthesamestimulatory
effect.However, theability ofanisolatedasequenceto actas a recombinational hot spotwas completely abrogated in the presenceof extensive flankingsequencehomology, particularly
that ofthe b andc sequenceswhich normally surround thea sequence at the L-S junction. These observations serve to
resolve thecontroversyovertherole that theasequenceplays
in HSV-1 genome isomerization by demonstrating that the high frequency of recombination events which arise during
DNAreplication through the b andcrepeatsacts todiluteout theenhancedrecombinogenicity exhibited by this element.
MATERIALS AND METHODS
Cells andviruses.HSV-1 (strain KOS) used in
superinfec-tionswaspropagated in Verocells. Stocks of the UL28gene
deletionvirus gCBwereprepared bypassageina
complement-ingcell line,Cl,whichexpressesthe UL28geneproduct(32).
Both cell lineswere grown in Dulbecco's minimum essential
medium supplemented with 5% fetal calfserum.
Plasmid constructions. (i) 440 derivatives. 440(Fig. 1)was
constructedby attachingEcoRI linkersontothe 440-bpSmaI
fragmentof theHSV-1 (strain F)a sequenceofpRB380(14)
and cloning it into the EcoRI site of pUC19. 440A1 was
generated by performing a 160-bp SstII collapse in 440; this
resultedin thedeletion of thepac2and DR4 elements of 440
but spared the complete DR2 array. 440A2 through 440A10
contain spontaneous deletions of integral numbers of DR2 repeats from 440A1 and were isolated by passaging this
plasmid in the recombination-proficient E. coli strain C600. 440A11, 440A12, 440A13, and440/14 represent 440 mutants that possess part or all of the DR2 array but lack both pac elements.Theywereconstructedby transferringtheApaI-SstII fragments of 440A1, 440A2, 440A8, and440A10, respectively,
into the ApaI-SstII sites of the 193A4 construct described below.
(ii) 193 derivatives. A HaeII fragment containing the two tandema sequences ofplasmidpCW103 (40) wasisolatedby gel purification, digestedwithMnlI, and shotgun cloned into
the SmaI site of pSG424 (24). One of the clones whichwas
isolated, 193A1,containedasmallsegmentof the bsequence,
aDR1 repeat, andacomplete Ub withpacl motif. An80-bp
SstIIfragmentfrom 440 which contained thepac2elementwas
then inserted into the SstII site of 193A1togenerate 193(Fig. 2);this derivative therefore containsanintactpac2-DR1-pacl
arrangement, whichrepresentsthe minimum signal necessary forcleavageand packaginginamplicon assays(19). Finally,a
series of SmaI- and Dral-generated deletions was used to createtheremainingmutantderivatives of193;these lackpacl (193A3),lack bothpacelements(193A4),orlack thesequences
adjacenttopaclwhicharenotconservedamongherpesviruses yet are essential for the cleavage/packaging process (193A2). All 440 and 193 derivatives were sequenced by the dideoxy
method toverifytheirauthenticity.
(iii) pL, pLR, and pLAR. Each of the 440 and 193 deriva-tives was inserted into the IS50-containing plasmid pL to facilitate their incorporation into the Tn5 shuttle plasmids pLR andpLAR. pLwasgenerated byfirstcloning the 2.4-kb
SmaIfragment containingthe left half of the Tn5sequencesin
pMC110::Tn5 (39) into the PvuII sites ofpUC19. An EcoRI linker was then inserted into the PvuII site of the IS50L
5.0 6.0
J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.77.551.83.252.2]MECHANISM OF HSV-1 GENOME ISOMERIZATION 37
element of this plasmid. The various a sequence derivatives
were subcloned via their flanking EcoRI sites into pL and screened by restrictionsite mappingto determine their orien-tation.
To create pLR-a, pMC11O::TnS was first cleaved at its
Sall
sitestogenerate twofragmentswhichrepresented the left and right halves of the Tn5 transposon. The 6.0-kb
Sall
fragment containing ISSORwasself-ligated tocreate pR*, and the 2.7-kb Sall fragment containing ISSOLwascloned intotheuniqueSall
site of pMC110 (3) to generate pL*. Next, the 1.3-kb
HincIl-BglII fragment from pL, which contains the entire IS50
ele-mentwith its440 or 193 derivative insert,was cloned into the HpaI-BglII sites of both pL* and pR*. These transposon halves, whichnowcontainedidentical a sequenceinserts, were then reassembledtocreate the intact TnS molecule in plasmid pLR-a. This involved ligating the ISSOL-containing
BglII-SstI
fragment of pL* into theBamHI-SstI sites of pR*. pLR and pLR-474wereidentical topLR-a, exceptthatthe former lacks
a sequence inserts in its IS50 elements, while the latter contains a 474-bp insert derived from the Saccharomyces
cerevisiae GAL4 gene as a non-HSV-1 DNA control. This insertwasobtained byfirst cloning the 0.5-kb GAL4-contain-ing
HindIII-BamHI
fragment ofpSG424 into pGEM7 (Pro-mega) and then transferring itinto pL as anEcoRI fragment for incorporation into pLR.pLAR-a
represents a derivative of pLR-a in which all but236 bp of the ISSOR element has been deleted; it was con-structed intwosteps. First, a 0.2-kbHindIIl-BsaHI fragment of the IS50 element ofpLwhich contains the a sequence insert
was transferred into the HindIII-AccI sites of pUC19. The
2.8-kb IS5OL-containing BglII-SstI fragment of pLR-a was then transferred into theBamHI-SstI sites of this plasmid, so that thefinalconstructcontained identicalasequencederivatives in inverted orientations. pLARisidentical topLAR-a but lacks a
sequenceinsertsin its IS50 elements. pLRAFsp, which
repre-sents a pLAR derivative with a non-HSV-1 DNA (i.e., Tn5-derived) insert,wasconstructed byreplacingthe 1.3-kb HpaI-BglII fragment of the IS5OR element of pLR with the 0.6-kb
FspI-BglII
fragment ofISSOR.(iv) pGAL4-STD. The plasmid used as an internal standard in recombination assays,pGAL4-STD,wasgenerated inthree
steps. First,
pSG424-ori,
was constructed by inserting the 0.2-kboris-containing
SmaI fragment of pMC110 into the SmaI sites of pSG424. Next, pMC-0.5HK was constructed by inserting the 0.5-kbHindIII-KpnI
fragmentofpSG424 into theHindIII-KpnI
sites of pMC110. Finally, pGAL4-STD wasassembled by inserting the 0.7-kb
BglII-SstI
fragment ofpSG424-oris
into the BamHI-SstI sites ofpMC-0.5HK;
this resulted in a construct which contains two tandem repeatcopies
of the yeast gene GAL4 linked to anHSV-1 origin ofreplication.
(v) p3.4Hin and p3.4Hin-E. To create p3.4Hin-E, a TnS molecule which contained
EcoRI
sites within both IS50 ele-ments first had tobe constructed. This was accomplished byfirst cleaving the transposon in pTn5A1 (39) into two halves,
each of which contained an IS50 element. The 3.0-kb
ISSOR-containing SalI-SstI
fragment ofpTn5Al
wasinsertedinto theSalI-SstI sites of pMC110 to generate pAIR. The 5.8-kb
IS5OL-containing
BamHI
fragmentofpTn5A1wasreligatedtoitselftogenerate
pAlL.
The 1.0-kbNotI-BglIIIS50-containingfragments
of bothpAlR
and pAlL were thenreplacedbythe 1.0-kbEcoRI linker-containing NotI-BglII fragment of pL toyield
pAIR-E
andpAlL-E,
respectively. Finally, the 5.8-kbSalI-SstI fragment of pAlL-Eand the 3.4-kbSalI-SstI fragment
of
pAlR-E
were ligated to createpTn5A1-E. p3.4Hin-E andp3.4Hin
(see Fig. 9) were then constructedby inserting the3.4-kb
HindIII
fragments ofpTn5A1-E
andpTn5A1,
respec-tively, into theHindlIl
site of a pMC110 derivative whose uniqueEcoRI
site had been destroyed. All but a 335-bp segment, which contained theEcoRI
linkers in p3.4Hin-E, wasdeleted from the
ISSOL
andIS5OR
elements during this final step. The construction of theEcoRI
gene-expressing plasmid pMENs has been described previously (18).(vi) pBaC andpBAaC. The first step in generating pBaC was to subclone the 2.7-kb
EcoRI
fragment ofpCW101
(40) into theEcoRI
site of pSG424, yielding pSG424-BaC. The con-struction of pBAaC involved first removing the a sequence from pCW101 by digesting this plasmid withBstEII-BspMI,
blunting the resulting ends with
Klenow
fragment, and religat-ing; the 2.2-kbEcoRI
fragment of this plasmid,pCW11Ala,
was then subcloned into the
EcoRI
site of pSG424 to generate pSG424-BAaC. TheHindIII-XbaI
fragments ofpSG424-BaCand pSG424-BAaC, which contain 474 bp of GAL4 DNA
sequences in addition to the L-S junction sequences, were then inserted into the
HindIII-XbaI
sites of pMC110 to create pMC110-BaC and pMC110-BAaC, respectively. The 2.7- and 2.2-kbEcoRI
fragments ofpCW101
andpCW1l1Aa,
respec-tively, were also subcloned into theEcoRI
linker of pL* to generate pL*-BaC and pL*-BAaC, respectively; theBglII-partial
SstI
fragments of these two plasmids were then ligated into theBamHI-SstI
sites of pR* to generate pLR-BaC andpLR-BAaC.
Finally, the 5.4- and 4.9-kbBglII-KpnI
fragments of pLR-BaC and pLR-BAaC, respectively, were ligated withthe 6.0- and 5.5-kb
BamHI-KpnI
fragments ofpMC110-BaC
and pMC110-BAaC, respectively, to generate pBaC and
pBAaC, respectively. In both plasmids, the only inverted repeat homology present was derived from the L-S junction of the HSV-1 genome.
Analysis of
TnS
inversion events in a transfection-superin-fection assay. Vero cells were transfected in triplicate with 6,ug
of each plasmid by calcium phosphate precipitation and sub-jected to a 15% glycerol shock after 4 h. The medium was changed after 18 h, and the cells were superinfected with HSV-1 (KOS) or gCB at a multiplicity of infection of 10 to 20. After 24 h, the infected-cell DNA was isolated and treated with RNase, proteinase K, phenol, and chloroform according to the procedure of Weber et al. (39) before being split equally into six Eppendorf tubes for precipitation overnight at -70°C.
The DNA from a single tube was then resuspended in 190RI
of water-21[lI
of10x restriction buffer, digested overnight with 30 Units each of the appropriate restriction endonucleases, reprecipitated, electrophoresed on a 0.8% agarose gel, trans-ferred to GeneScreen Plus (NEN DuPont), and hybridized toTn5-
or GAL4-specific probes which had been radiolabeled with [32P]dCTP with a random-primed labeling kit (Boehr-inger Mannheim). Southern blots were exposed to Kodak XAR5 film for various lengths of time without an intensifier screen, and the resulting autoradiograms were quantitated with a Hoefer GS 300 densitometer.Si nuclease sensitivity assay. Cleavage of a sequence sub-fragments cloned into pL by Si nuclease was performed by using a modification of the assay described by Wohlrab et al. (42). Briefly, 1.5
,ug
of each plasmid was treated with 3.5 U of Si nuclease (Bethesda Research Laboratories) for 30min at37°C in 50
[lI
of reaction buffer (40 mM sodium acetate [pH 4.6], 50 mM NaCl, and 1 mM ZnSO4); the reactions were stopped by the addition of EDTA to a final concentration of 30 mM. The samples were then subjected to phenol and chloro-form extraction, ethanol precipitation, digestion withPstI,and electrophoresis on a 0.8% agarose gel.VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
38 SARISKY AND WEBER
B
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
D. L
-PS50L
neo Pst Pst
* I <
I~-M ~Pstlfragment A(1.1 kb)
SI nuclease Pst
* 1
Pstl-S1IfragmentAlI( 0. 7kb) [i h
Pstl-SIfragment A2(-0.4kb) sI L3. |
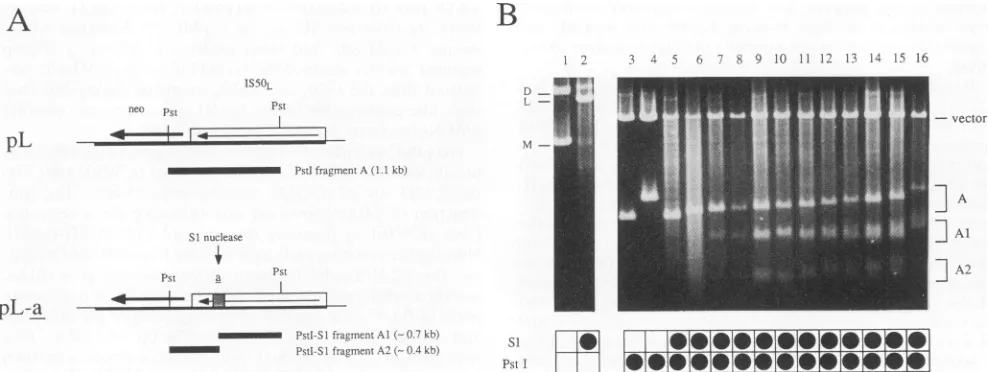
FIG. 3. Ability ofasequencesubfragmentstoform anisomorphic DNA. (A) 440 derivatives containing variable numbers ofDR2 repeatswere
inserted into the IS50 element of plasmid pLto generateapanel of pL-a derivatives. Recognition of anisomorphicDNA conformations inpL-a plasmids by Si nuclease will be evident by cleavage of theasequence-containing PstI fragmentAtoyieldtwosmallerfragments,Al and A2.(B) Agarose gel electrophoresis of SI and/or PstI digests of pL and pL-a plasmid DNAs. D, L, and M refertodimer, linear, andmonomerspecies, respectively,presentin thefirsttwolanes. In the remaining lanes, A, Al, andA2refertothefragments identifiedinpanel A, while thevectorband
referstothe 3.6-kb vector-containingPstIfragmentcommontoallpL-aderivatives. Plasmidsweretreated withPstIalone,with Si nuclease alone,
orwith Si followed by PstI,as indicated in the gridatbottom. Lanes: 1, 2, 4, and 15,pL-440A1; 3and 5, pL; 6, pL-44Oz10; 7, pL-440A9; 8, pL-440A8; 9, pL-440A7; 10, pL-440z6; 11, pL-440A5; 12, pL-440A4; 13, pL-440A3; 14, pL-440A2; 16, pL-440.
RESULTS
Construction and characterization ofa sequence mutants. The enhanced recombinogenic properties of the a sequence are thoughtto be dueto anovel anisomorphic DNA
confor-mation occurring at the DR2 repeat elements or to
double-strand breaks resulting from cleavage and packaging events
mediatedby the pacl andpac2motifs.Inordertoexamine the possible contribution of these featurestorecombinogenicity, a
series ofsubfragments of theasequencewhich contained both wild-typeandmutated componentswasconstructed, such that
all possible combinations of anisomorphic, nonanisomorphic, cleavage/packaging-proficient, and
cleavage/packaging-defi-cient elements could be prepared foranalysis in genetic and
biochemical assays.
Thefirst section of thea sequence which wasexamined is
found in 440, a SmaI fragment of the wild-type a sequence
which contains the full DR2arrayand bothpac sequences(Fig.
1).Apanel of deletionmutantswasconstructedwhich
repre-sented both anisomorphic and nonanisomorphic versions of 440 inwhichone orbothpacelements weredeleted (Fig. 1).
To furtherexploretheroleof thepacelements andtoestablish
apossiblelinkbetweencleavageandpackagingwith enhanced
recombinogenicity, asecondsubfragment which includedonly
thetwopac elementswas examined: 193 is anMnlI fragment
which contains the pac2-DR1-pacl arrangement from two
tandemly repeateda sequencesandcompletely lacks the DR2
array (Fig. 2). An analogous fragment from a closely related
HSV-1 strain has previously been shown to function as a
minimalcleavageandpackaging signal inampliconassays(19).
Mutations in 193 in which either orboth pac elementswere
deletedwere then created(Fig. 2).
Prior to examination of the recombinogenic activities of thesea sequencederivativesin aquantitative sequence
inver-sionassay,thephysical and biological properties of the 440 and
193derivativeswerecharacterized. First, the ability of variable
numbers of DR2 repeats toform anisomorphic DNA
confor-mationswasexaminedbyanSi nucleasesensitivityassay(42).
440 and a series of deletion mutants containing variable
numbers of DR2 repeats, which potentially represent both
anisomorphic and nonanisomorphic conformations (Fig. 1),
weresubcloned into the IS50 element ofplasmid pL (Fig. 3A)
and treated with SI nuclease. The 440 mutants analyzed included440LA1, 440A2, 440A8, and 440A10,which contain 19, 16, 4, and 1 DR2 repeats, respectively (Fig. 1), and 440A3, 440M4, 440A5, 440A6,440LA7,and440A9,which contain15, 13, 10, 6, 5, and 3DR2 repeats,respectively (not illustrated).
An Si-sensitive site was observed in plasmids bearing the
440 insert and its deletion derivatives and could bemappedto the DR2repeatarrayby digestionwith PstI(Fig. 3B)andother
restriction endonucleases(25). Cleavage by S1 wasevident in
theconstructspresentinlanes 7to16 ofFig. 3B,since theira
sequence-containingPstI fragments (fragment A in Fig. 3A) hadbeen cleaved by S1 toyieldtwosmallerproducts (Al and
A2). The breadth of the Al and A2 bands in those pL-a
derivatives which containedhigher-copy-number inserts (Fig. 3B,lanes 10to 16)indicated that the Si cleavage eventcould
occur atvariable sites within the DR2repeatarray.However,
plasmids with fewer DR2 repeats possessed sharper bands
(Fig. 3B, lanes 7 to 9) and thus contained fewer potential
nuclease-sensitive sites. Incontrast,constructswhich lackedan
insertion (pL; Fig. 3B, lane 5) or which contained either a
singleDR2 repeat(pL-440A10;lane6)or anon-HSV-1 DNA insert (25) were resistant to S1 cleavage. Thus, any 440 derivative which contained at least three DR2 elements was
found to facilitate the formation of an unwound nuclease-sensitive DNAstructure. Moreover, recognition of this struc-ture by S1 nuclease required negative supercoiling of the
plasmid template, since cleavage of the DR2 repeat array
could be prevented by linearization of the DNA prior to nucleasetreatment(25).Allof these observationswerefound to be in complete agreement with the earlier results of Wohlrab et al. (42). Thisobserved Si sensitivitysuggeststhat
A
pL
pL-a
-vector
] A ] Al ] A2
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.554.71.257.2]MECHANISM OF HSV-1 GENOME ISOMERIZATION 39
Ss
l
l1
i
l
ml
GAL4 GAL4 ori
(NruU/SstI = 3.9 kb)
recombination
(deletion) Ss
(NrulISstI = 3.4 kb)
a Nr a Ss
pLR-a
--.
m0
9
IS50 R IS50L ors
(Nrul/Sstl = 5.0, 2.1 kb)
recombination
(inversion) a Nr
I1
I41am-
Ss1(NruI/SstI = 4.6,2.5 kb)
a Nr a Ss
IS50L oris (Nrul/SstI = 3.6,2.1 kb)
recombination
(inversion) Nr
n
.a Ss
1
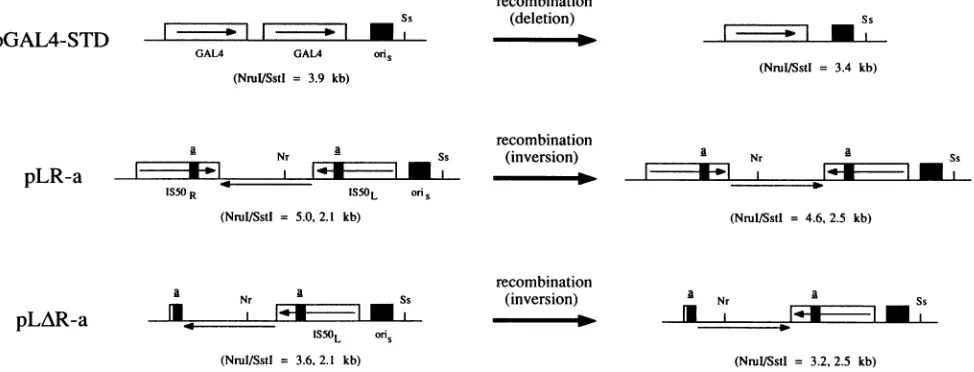
-FIG. 4. Plasmids used in the quantitative Tn5 inversionassay.pGAL4-STD undergoes deletion ofoneof itstwocopies of theyeastgeneGAL4
ataconstantfrequency when replicated in HSV-1-infected cells, making itauseful internalstandard for TnS inversionassays.pLRand pLARare
twoTnS-containing plasmidswhichdifferdramatically in theirabilitiestoundergo sequenceinversioneventsin HSV-1-infected cells, since the
latter haslostalmostthe entireIS5OR element through which recombinationmusttake place. Theasequencederivatives illustrated in Fig. 1 and
2wereinsertedininverted orientation into the IS50 elements of both pLR and pLARasshown,togenerateapanelofpLR-aand pLAR-aplasmids
whichwereanalyzed inTnSinversion assays.
the DR2sequencesof these 440 derivatives could besimilarly
cleavedby nucleases presentin HSV-1-infected cells andmay
therefore represent sites at which recombination events are
initiated.
Inaddition, theabilityofa sequencesubfragments
contain-ingmutations in thepacl andpac2motifstomediatecleavage/ packaging events was examinedby using an amplicon
propa-gation assay. Defective HSV-1 plasmid vectors or amplicons
minimallycontain both aviral DNAreplication origin and an a sequence and arereplicated as large concatamers, cleaved,
and packaged into virions in the presence of a helpervirus
(29).A number of 193 and 440 derivatives (Fig.1 and2)were
therefore inserted into an HSV-1 origin-containing plasmid
and tested for their abilitiestobepropagatedasampliconsby
cotransfection into Vero cells with infectious HSV-1 helper virusDNA. DNase-resistant virion DNAwas prepared from
the resulting viral stocks after multiple low-dilution serial passagesand examined for thepresenceofplasmid amplicons bySouthern blotanalysis. Onlythoseasequencesubfragments which contained both thepaclandpac2 signals,440 and 193,
were observed to promote replication and packaging into virions with high efficiencyand without rearrangementevents
(25).Incontrast, mutantderivatives of440 and 193 from which
one orbothpacelements had been deleted either failedtobe
propagatedasampliconsor werepropagated onlyafter illegit-imate recombinationeventshad restoredacompletesetofpac
elementstothe defectiveasequence. Thus,the double-strand break generated within the wild-type 440 and 193
subfrag-ments during cleavage and packaging enabled theiramplicon
vectors tobe passaged through multiple rounds ofinfection;
thesefindingsareincompleteagreementwithpreviousstudies ofasequencemutantsinamplicon propagationassays(6, 37).
The possibilitythat this double-strand break mayincrease the
frequency of recombination events at the a sequence by generating free DNA ends was explored in the
transfection-superinfection assaydescribed below.
Utilization ofaquantitativeTnS inversionassaytoanalyze
the recombinogenic properties ofa sequence mutants. The
relative contributions of DNA breaks ina sequence subfrag-mentsresulting either from anisomorphic structural
conforma-tionsorfromcleavageandpackagingeventstoHSV-1 recom-bination were ascertained by using a Tn5 shuttle plasmid
system.Thissystemreliesontheabilityof the TnStransposon linked in cis to an HSV-1 origin of replication to undergo
sequence inversion in a transient transfection-superinfection assay(39).Such inversioneventsaremediatedby generalized
recombination between the invertedrepeat IS50elements of TnS. To accurately quantitate increases in recombination
frequencywhichwould result from the insertion of variousa sequencesubfragmentsinto theIS50 elements of the
transpo-son, acotransfected standardplasmid (pGAL4-STD)
possess-ingaconstantrecombinationfrequencywasusedas aninternal
reference. This plasmid contained two tandemly arranged copies of the yeast gene GAL4 and an HSV-1 origin of
replication (Fig. 4). In the absence of HSV-1 infection, pGAL4-STD yielded alinearized fragmentthatwas3.9 kb in size.However,in thepresenceof HSV-1 infection,oneof the
copiesof GAL4wasdeletedby replication-mediated
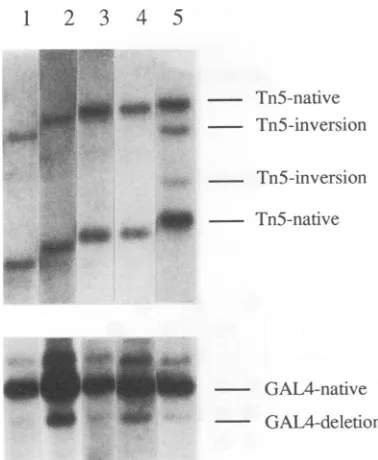
recombi-nation. This resulted in theappearanceofalinearized recom-bination productwhichwas 3.4 kb in size (Fig.4 and5). The internal standardwastested in cotransfectionexperimentswith the wild-type Tn5-containing plasmid pLR (Fig. 4), which
yieldstwonativefragmentsafter NruI-SstI digestionaswellas two novel fragments resulting from replication-mediated
se-quenceinversion. Bandscorrespondingtothe nativefragments
and recombination products of both pLR and pGAL4-STD
could be readily detected in transfected-cell DNA (Fig. 5); DpnI digestion was utilized to verify that only replicated specieswereanalyzed.
The differences in the levels of inversion and deletionbands
versus native bands between pLR and pGAL4-STD,
respec-tively, remained relatively constant over repeated transfec-tions. InversionproductsinpLRweredetectedatafrequency
ofapproximately25% inrepeatedtransfections.This indicated that the assay was only halfway to reaching recombinational equilibrium, such that systemsaturation would be markedby
pGAL4-STD
pLAR-a
(Nrul/SstI = 3.2, 2.5 kb) VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.60.547.79.265.2]40 SARISKY AND WEBER
pGAL-STD pGAL-STD
alone +pLR
1 2 3 4
-
TnS-native
- TnS-inversionGALA-native
GALA-deletion
-t
1-, - Tn-inversion
_t <i _ ~~TnS-native
HSV-1(KOS)superinfection
Digestion with: SstI DpnI
[image:7.612.319.554.70.252.2]NruI FIG. 5. Demonstration
I
of the quantitative Tn5 inversion assay. DNA was isolated fromVero cells which hadbeen first transfected
withpGAL4-STD aloneorwith bothpGAL4-STD andpLRand then
mockor HSV-1 infected.This DNAwasdigested with the indicated
enzymes and then analyzed by Southern blot, using radiolabeled
pUC19::Tn5DNAas aprobe todetect both the native and
recombi-nation-derived bands of bothplasmids.
the appearance ofinversion products at afrequency of 50%. This inversion assay therefore representsa convenient model
system for the quantitative analysis of HSV-1 replication-dependent recombination events and would be sensitive enough to detect any increase in Tn5 inversion upon the
insertion ofarecombinational hotspot into the IS50 elements ofpLR. In other words, ifanyof theasequencesubfragments described above (Fig. 1 and 2) can function as hot spots for recombination, they should be able to promote a discernible
increase in Tn5 inversion frequency in pLR relative to the GAL4 deletionfrequency inpGAL-STD. Thus, by quantitating differences between the levels of pLR inversion products and pGAL-STD deletion products within a given experiment, an accurate assessment of the recombinogenic potential of any
inserted a sequence subfragment can be made. Since the
addition of identical a sequence derivatives into both IS50 elementsresulted inasmall increase in thesequencehomology between inverted repeats which could theoretically increase the frequency ofgeneralized recombination events occurring inTn5, thepLRderivative pLR-474, which containeda474-bp
insert of non-HSV-1 DNA (i.e., the yeast GAL4 gene), was
used as acontrol.
The recombinogenic properties of the a sequence mutants by themselves (i.e., in the absence of flanking IS50 homology)
were also examined by using the plasmid pLAR, in which
almost all of the IS50R element has been removed (Fig. 4). This plasmid was designed to directly identify isolated hot spots of recombination, since it lacks the IS50 homology through which normalsequenceinversioneventsaremediated,
and apreviousanalysis demonstrated that Tn5 inversion does not occur at detectable levels in constructs which contain regions of invertedrepeathomology thatare assmall(i.e.,600
bporless)astheinserted 440or 193subfragments (39). Thus,
the extent to which an inserteda sequence mutant increases Tn5 inversion above its normal level in pLR or allows the
detection ofTn5inversion in pLAR willbeadirectindication
of the relative ability of its anisomorphic DNA conformation
2 3 4 5 6 7 8 9 10
- Tn5-native
- Tn5-inversion
- Tn5-inversion
- Tn5-native
- GAL4-na6ve - GAL4-deletion
:WX
FIG. 6. Analysis of the recombinogenic propertiesof 440 and its
derivatives in the pL-AR plasmid. Each pLAR-440 derivative was
cotransfected into Vero cells with pGAL4-STD, superinfected with HSV-1,reisolated, digestedwithNruI-SstI-DpsnI, electrophoresedona 0.8% agarose gel, and blotted onto a GeneScreen Plus nylon
mem-brane. Hybridizations were carried outwith two differentprobes to
circumvent the problem of quantitating overlapping pLAR- and pGAL4-STD-derived bands. The blotwasfirstprobedwiththe radio-labeled 3.4-kbHindIIIfragment ofTn5tovisualize thepLAR-specific bands (upperpanel). Thisprobewas then stripped,and the blotwas
reprobed with the radiolabeled 0.5-kb GAL4 gene-containing
HioldIlI-EcoRI fragment of pSG424 to visualize the pGAL4-STD-specific bands(lowerpanel).Thenative bands and therecombinationproducts for both cotransfected plasmids are indicated on the right of the autoradiogram.Lanes: 1,pLAR;2,pLAR-440A10;3,pLAR-440A14;4, pLAR-440A8;5,
pLAR-440A13;
6,pLAR-440A2; 7,pLAR-440A12;8, pLAR-440A1; 9,pLAR-440A11;
10,pLAR-44t).
or cleavage/packaging signals to enhance the frequency of
existing homologous recombination events in HSV-1. Analysis of therecombinogenicpropertiesof the a sequence mutants in
pLAR.
Duplicate copies of the 440 and 193 a sequencesubfragmentsaswellastheirmutantderivatives(Fig.1 and 2) were inserted in inverted orientation intopLAR to assay their recombinogenicity in the
transfection-superinfec-tion system.
Representative
Southern blots of transfectedpLAR-440
or pLAR-193 derivatives digested withNruI-Sstl-DpnI
and hybridized with Tn5-and GAL4-specific probesare shown inFig. 6and7, respectively. The intensities of bands in the autoradiogramswere quantitated by densitometryto nor-malizethe TnS inversion levels to the levels ofdeletion in theinternal standardpGAL4-STD; these values areexpressedas a recombination index in Fig. 1 and 2.
The wild-type 440 insert was found to possess significant recombinogenic potential in that it conferred a 50-fold in-crease in inversion frequency over a pLAR plasmid which
lacked any inserts (Fig. 1). However, all of the deletion
derivatives of 440 were defective for this enhancement in
recombination activity in pLAR, although a very low level of
inversion could be detected in
pLAR-440A1, pLAR-440A2,
pLAR-440A11,
and pLRAFsp, whichrepresents apLARplas-mid that contains a600-bpinsertofnon-HSV-1 DNA(Fig.
1).
Similarly, pLAR-193 also showed a50-fold increase in
inver-sionfrequencyoverpLAR whichlacked aninsert,butdeletion of eitherorboth pac sequences from the 193 insertresulted in the loss ofmost ifnot all of thisactivity(Fig. 2and7).These results demonstrated that both the wild-type 440 and 193
J. VlII(OL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.66.300.71.269.2]MECHANISM OF HSV-1 GENOME ISOMERIZATION 41
1
2
3
4
5
- Tn5-native
-
TnS-inversion
-.
Tn5-inversion
-
TnS-native
GAL4-native
[image:8.612.78.267.74.304.2]GAL4-deletion
FIG. 7. Analysis of the recombinogenicproperties of 193 and its derivatives in the pLAR plasmid. Each pLAR-193 derivative was cotransfected into Vero cells with pGAL4-STD, superinfected with HSV-1,reisolated, digested withNruI-SstI-DpnI, electrophoresed on a 0.8% agarose gel, and blotted onto a GeneScreen Plus nylon mem-brane. Hybridizations were carried out with two different probes to circumvent the problem of quantitating overlapping pLAR- and pGAL4-STD-derived bands. The blot was first probed with the radio-labeled3.4-kbHindlIl fragment of Tn5 to visualize the pLAR-specific bands (upperpanel). This probewasthenstripped, and the blot was reprobed with the radiolabeled 0.5-kb GAL4gene-containing HindlIl-EcoRI fragment of pSG424 to visualize the pGAL4-STD-specific bands(lowerpanel). The native bands and the recombination products for both cotransfected plasmids are indicated on the right of the autoradiograms. Lanes: 1, 193A4; 2, 193A1; 3, pLAR-193A3;4,pLAR-193A2; 5,pLAR-193.
A
pLAR-1 93
pLAR
a Nr a
T I [. - _
.4 IS50 L oris
Nr
IS50L oris
inserts could indeed function as hot spots of recombination.
Moreover, the mutationalanalyses of the 440 and 193 inserts
indicated that the presence of two functional pac sequences was critical for enhancing a sequence-driven recombination; however, the DR2 array, with its associated anisomorphic
DNAconformation, wasdispensable for this activity.
Role ofdouble-strand breaks generated during thecleavage/
packaging process in promoting recombination at the a se-quence. The finding that two functional pac sequences were both necessary andsufficient for mediating enhanced recom-bination at the a sequence suggested that the double-strand break generated at this site by the viral cleavage/packaging machinerywasinvolved in this process. Toexplore this
possi-bility further, recombination between a sequenceswas
exam-ined by using the quantitative Tn5 inversion assay utilized previously,except that thetransfected cells weresuperinfected with an HSV-1 cleavage/packagingmutant, gCB, rather than
the wild-type KOS strain. gCB lacks almost the entire UL28
openreadingframe,whichrenderstheviruscompletely
defec-tive for the cleavage andencapsidationofreplicatedviral DNA
(32). Since the UL28 gene product is essential for virus
propagation, stocks ofthe gCB mutant had tobe grown in a
UL28-expressingcelllinepriortotheir use inexperimentswith
noncomplementing Vero cells. Three constructs which had
been employed in the experiments described above, pLR, pLl\R, and
pLAR-193
(Fig. 8A), were transfected into Verocells, superinfected with gCB, and digested with
Nrul-Sstl-DpnI
priorto Southern blot analysis; arepresentative blot ofthetransfected-cell DNAis shown in Fig. 8B.
a sequence-independent recombination was unaffected by theconditions employedin thisexperiment, since pLR
exhib-ited the samehigh levels of inversion after gCBsuperinfection (Fig. 8B) thatwere observed after wild-type virus superinfec-tion (Fig. 5).The finding thatreplication-dependent recombi-nationwasuninhibited forpLRin gCB-infected cellswas not
unexpected, sincethe majorreplication defect inthis superin-fecting virus is in the process of cleavage and packaging and notin DNAsynthesis. However, noinversion eventscould be
detected in pLAR-193 superinfected withgCB (Fig. 8B); this
B
1 2 3-Tn5-native
Tn5-inversion
Tn5-inversion Tn5-native
*L.
II
I
- -Gal4nativeCn~~~~-nIArHoIptinnIS50R _ S5GL
Vw-FIG. 8. Analysisofrecombination in cells infected withanHSV-1 mutantdefective for thecleavage/packagingprocess. (A)Structuresofthe transfectedplasmids pLAR-193, pLAR,andpLR. Nr, NruI.(B)TheplasmidsinpanelAwere individually cotransfected withpGAL4-STDinto Verocells,superinfectedwith theHSV-1 mutantgCB, reisolated, digestedwithNruI-SstI-DpnI,electrophoresedon a0.8%agarosegel,blotted onto GeneScreen Plusmembrane, and hybridized with the radiolabeled 3.4-kb HindlIl fragment of Tn5 (upper panel). This probewas then
stripped,and the blotwasreprobedwith theradiolabeled 0.5-kbGAL4gene-containingfragment ofpSG424 (lower panel).Thenativebandsand therecombination productsfor bothcotransfected plasmidsareindicatedontheright ofthe autoradiograms.Lanes: 1,pLAR-193; 2, pLAR;3,
pLR.
PLR
V()L.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.130.480.494.654.2]42 SARISKY AND WEBER
A
B
San
i Km p3.4Hin
p3.4Hin-E
Sall
W
orisI- *
EcoRI EcoRl
Sall Sall
RKmr
|s1
|Sall: 4.3Kb, 1.9Kb Native Sall 4.9Kb. 1.3 Kb Inversion
PlasmiS'a!scl3 p3 4Hin
p34Hin-F pMENs
HSV1(KOS) Superintection.
D(gostionwith Sall
Dpni
1 2 3 4 5 6
-Tn5-inversion -Tn5-native
-Tn5-native
_Tn5-inversion
*I-le-*1 1-1
1- 1
-.-. .
0O
90
0
[image:9.612.67.556.72.298.2]"'555
FIG. 9. Ability of a double-strand break generated by an in vivo-expressed restriction endonuclease to promote recombination in aTn5 inversion assay. (A) Structures and predicted restriction fragments of the transfected plasmids p3.4Hin andp3.4Hin-E.(B) The plasmids in panel A wereindividually transfected with or without theEcoRI-expressingplasmid pMENs into Vero cells, mock or HSV-1infected,reisolated,digested withthe enzymes indicated in the grid, electrophoresed on a 0.8% agarose gel, blotted onto a GeneScreen Plus membrane, andhybridizedwith theradiolabeled 1.3-kb HpaI-BglII fragment ofpUC19::Tn5 DNA.This probe does not hybridize with the pMENs plasmid, therebyenabling visualization of only the test plasmids. The native bands and the recombination products are indicated on theright of theautoradiograms.
wasinsharpcontrast to the high levels ofinversion observed
for thissame plasmid in the presence ofsuperinfecting wild-typevirus (Fig. 2 and7).
Thus, enhanced recombinogenicity inanisolateda sequence
requires not only the presence of both the pacl and pac2
elements(Fig. 1 and2) but also the complete complement of viral proteins required for cleavage and packaging (Fig. 8). These results stronglysuggestthat it is the double-strand break generated in theasequenceduring the cleavage and packaging
process,andnotthesequencesofthe pac elementsthemselves, which isrequired forpromotinghigh-frequency recombination
events at the a sequence. One important prediction of this hypothesis isthat adouble-strand breakin any DNA sequence should be ableto exert asimilarstimulatory influence on the
initiation of HSV-1 recombination. To test this prediction directly, double-strand breaks which were generated in
non-HSV-1DNAby theexpression ofarestriction endonuclease in virus-infected cellswereexamined for theirabilitytopromote
frequencies ofrecombination eventswhichwerecomparableto
those observed for thea sequence.
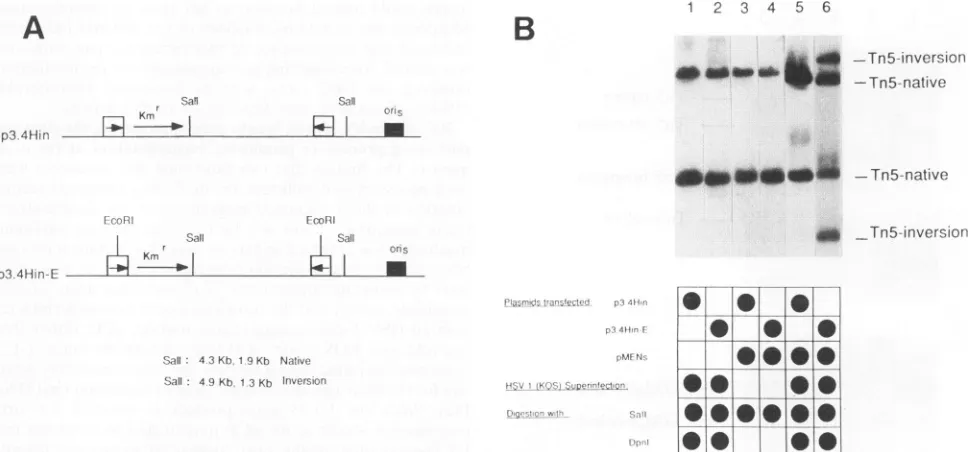
The plasmids used in these experimentswere two deletion
derivatives ofpLR, p3.4Hin and p3.4Hin-E (Fig. 9A), which differonlyin the presence of anEcoRI site centered within the invertedrepeatsof thelatterconstruct.Since itwaspreviously
established that a minimum of 600 bp of IS50 homology is essential todetectaninversion eventin theTn5-based
recom-bination assay (39) and since both p3.4Hin and p3.4Hin-E contain only 340 bp of IS50 homology, it was not surprising
that no sequence inversion events were detected in these
plasmids by Southern blot analysis (Fig. 9B, lanes 1 and 2).
However, pLAR-193 contains approximately thesame amount
ofsequencehomologyasthep3.4Hinplasmids but undergoes
significant
levelsof sequence inversion by virtueof itsrecom-binogenic
double-strand break created during thecleavage/
packaging process (Fig. 2 and 7). To generate a comparable
double-strand break in p3.4Hin-E as a means of potentially stimulatingrecombination,use wasmade ofpMENs,aplasmid which cantransientlyexpress theEcoRI restriction endonucle-ase gene in mammalian cells (18, 25). Cotransfection of
pMENs with p3.4Hin-E was found to have no effect on
recombination frequency in mock-infected cells (Fig. 9B,lane
4).However,thepresenceof both cotransfected pMENsDNA
and HSV-1 superinfection resulted in a dramatic increase in the p3.4Hin-E inversion frequency (Fig. 9B, lane 6). This
stimulatory effectwas notobserved with p3.4Hin,whose IS50
elementslack therecognitionsite for theEcoRI endonuclease, under identicalconditions (Fig. 9B, lane 5).
Thus, three components were essential for the appearance
ofhigh-frequency recombinationeventsin theseexperiments:
the pMENs plasmid, which expressed the EcoRI restriction
endonuclease that created a double-strand break within the
region of IS50homology; anEcoRIrestriction site, whichwas
cleavedby the endonuclease and thereforerepresents the site
of the double-strandbreak; and HSV-1 superinfection,which
provides in trans the viral factors necessary for
replication-mediated recombination.Moreover,these results demonstrate
thatabacterial DNA sequencewhichwasotherwise
recombi-nationally inertin HSV-1-infected cells could acquire
recom-binogenic properties which rivaled those of the a sequence
simply throughthe acquisition ofadouble-strand break. This
finding dramatically illustrates that, whilenospecificsequences were required for high-frequency recombination to occur in
HSV-1-infected cells, the creation of a double-strand break appearedtobecritical. Theimplication of this result is thatthe a sequence is able to act as a hot spot for homologous recombination simply because it happenstocoincide with the site of the double-strand breakwhich isgeneratedduringthe
cleavage/packaging process, not because it contains discrete J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
MECHANISM OF HSV-I GENOME ISOMERIZATION 43
1 2 3 4 5 6 7 8 9 10 11
Tn5-native Tn5-nversioi
GAL4-native
GALA-deletion Tn5-inversion
Tn-native
FIG. 10. Analysisof the recombinogenic propertiesof 440 and its derivatives inthepLR plasmid. EachpLR-440derivativewas cotrans-fectedinto Vero cells with pGAL4-STD, superinfected with HSV-1, reisolated, digestedwith NruI-SstI-DpnIl, electrophoresed on a 0.8% agarose gel, blotted onto a GeneScreen Plus nylon membrane, and
hybridizedwith radiolabeledpUCI9::Tn5DNA.The native bandsand
the recombinationproducts for bothcotransfected plasmidsare indi-cated on the right of the autoradiogram. Lanes: 1, pLR; 2, 440A10; 3, 440A14; 4, 440A8; 5, 440A13; 6, pLR-440A2; 7, pLR-440A12; 8, pLR-440A1; 9, pLR-440A1 1; 10, pLR-440; 11, pLR-474.
sequences which are required for high-frequency inversion events.
Analysisof therecombinogenic propertiesofthea sequence
mutants in pLR. Duplicate copies of the 440 and 193 a sequencesubfragmentsaswellastheirmutantderivatives(Fig.
1 and 2) were inserted in inverted orientation into pLR to assaytheirrecombinogenicitiesin the transfection-superinfec-tion system. Representative Southern blots of transfected pLR-440 and pLR-193 derivatives digested with NruI-SstI-DpnI andprobedwith pUC19::Tn5 (39) sequencesareshown
in Fig. 10 and 11, respectively. The intensities of the bands in
the autoradiogramswere quantitated by densitometryto nor-malize the Tn5 inversion levels to the levels ofdeletion in the internal standardpGAL4-STD;these valuesareexpressedas a
recombination index in Fig. 1 and 2.
Surprisingly,the introductionofthe 440 and 193 inserts and
theirdeletion derivatives into the intact IS50elementsofpLR resulted in only a minimal increase (13% or less) in Tn5 inversion frequency relative to the GAL4 deletion frequency (Fig. 1 and2).Thisslight stimulatoryeffect didnotindicate the
presence ofahot spotforrecombination but ratherappeared
1 2 3 4 5
TnS-native - Tn5-inversion
-. GAIA-native
- GAL4-deletion TnS-inversion
TnS-native
FIG. 11. Analysisof the recombinogenic propertiesof193 and its
derivatives in thepLR plasmid.EachpLR-193 derivativewas cotrans-fected into Vero cells with pGAL4-STD, superinfectedwith HSV-1, reisolated, digestedwith NruI-SstI-DpnI, electrophoresed on a 0.8%
agarose gel, blotted onto a GeneScreen Plus nylon membrane, and
hybridizedwith radiolabeledpUC19::TnSDNA. The nativebands and the recombination productsfor both cotransfectedplasmidsare indi-cated on the right of the autoradiogram. Lanes: 1, pLR-193A4; 2, pLR-193A1; 3, pLR-193A3; 4, pLR-193A2; 5, pLR-193.
to be the result of increasing the length of IS50 homology through which recombination events could occur, since the
plasmid containing the non-HSV-1 DNA insert (pLR-474) behaved similarly. Thus, the insertion of any a sequence
derivative, eventhe wild-type 440 and 193 inserts, which were shown to be highly recombinogenic in pLAR, had little
stimu-latory effect onthe level of homologous recombination which was already occurring between the IS50 elements of Tn5 in pLR.
Analysis of the contribution of the a sequence to HSV-1 genome isomerization. The simplest interpretation of the results obtained with the pLR-a constructs is that inserts such aswild-type 440 and 193could indeed function as hot spots of
recombination when examined in an isolated state (i.e., in
pLAR)but thattheylost this property whenflankedby regions of extensive homology, such as the 1.3 kb of IS50 sequences present in pLR. Since the a sequence is normallyflanked by much larger (approximately 16-kb)tractsof b and c sequence
homology at the L-S junction of the HSV-1 genome, this
elementmaysimilarly fail toact as arecombinationalhot spot in itsnaturalcontextintheviralDNA.Unfortunately, it is not
possibleto testthishypothesis directly by generatingavirusin
which all of the a sequences have been deleted, since such a mutant genome would be incapable of normal propagation
because of the loss ofthepac signals.Moreover, any attempts to characterize a genome in which the a sequence has been removed fromjustthe L-Sjunctionwould be hindered by the
efficient repair ofthe deleted elementbygene conversion-like
processes (23, 36).
To circumvent these difficulties in working with the viral genome, use was made of a modified Tn5 inversion assay in which the two IS50 elements of the transposon were replaced byeitheroftwoinvertedsegmentsoftheL-Sjunction.One L-S
junction fragment contained 698 bp of the b sequence, the
entire572-bpasequence,and 1,437bpof the c sequence; two
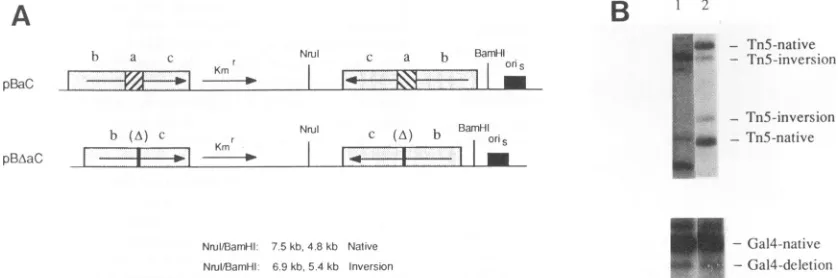
copies of thissegment wereused to constructpBaC(Fig. 12A). The other L-Sjunction fragmentwasidenticaltothat inpBaC
except that the a sequence had been deleted from both b-a-c
repeats; twocopiesofthissegment wereused to createpBAaC (Fig. 12A). These two plasmidswere cotransfected into Vero cells with the internal-standard plasmid pGAL4-STD,
super-infected with HSV-1, reisolated, cleaved with NrlI-BanHI-DpnI, and analyzed by Southern blotting (Fig. 12B). Quanti-tation of Tn5 inversion events afternormalizing for
pGAL4-STD deletion events revealed that the recombination indices for pBaC and pBAaC were nearly identical (3.68 and 3.62, respectively). Thus, the loss of the a sequence from the L-S junctionrepeats in pBAaC had noeffect onthe ability ofthis
plasmid to undergo recombination, and conversely, as in the
experiments utilizing
pLR-a,the presence of theasequence inpBaC failed to increase the frequency of existing sequence
inversionevents.Thesimplestinterpretationof theseresultsis that
high-frequency
recombination which arisesduring
DNAreplication throughthe b and c sequencesofthe L-Sjunction
is fully capable of dilutingout any
recombinogenic
contribu-tions of the a sequence.Thus,
the presence ofthis element most probably adds verylittleto theoverall poolof recombi-nation events which are responsible for HSV-1 genome isomerization.DISCUSSION
The phenomenon of genome isomerization is one of the most unique and yet enigmatic aspects of the life
cycle
of HSV-1. The L and S components ofthe HSV-1 genome are able to invert relative to each other at ahigh frequency,
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:10.612.59.298.75.185.2] [image:10.612.84.266.527.635.2]44 SARISKY AND WEBER
B
b a c K
I
I
Kmg
b (A) C r
I(--c
KmpBAaC
-|___
--_I_______-__
NruI c a b BanI-I
onBs
I
'I
G
M
-=NruI C (A) b or
1
-1 2
- Tn5-native
- Tn5-inversion
- Tn5-inversion
- Tn5-native
-Gal4-native
-Gal4-deletion
FIG. 12. Analysisofthe contribution of theasequencetorecombination betweenduplicatedL-Sjunctionsof the viralgenome.(A)Structures ofthe transfected plasmids pBaC and pB/aC.ThepredictedrestrictionfragmentsforpBaCarelisted; each of thecorresponding fragmentsin
pBAaCwill be 0.5 kb smaller because of the deletion ofthe a sequencesfrom this plasmid. (B) The plasmids inpanel Awere individually
cotransfected with pGAL4-STD intoVero cells, superinfectedwith HSV-1, reisolated, digested withNruI-BamHI-DpnI, electrophoresed on a
0.5%agarosegel, blotted ontoaGeneScreen Plus membrane,andhybridizedwith the radiolabeled 2.8-kbBglII fragmentofTnS(upper panel).
Asecondaliquotof thesameinfected-cell DNAwasdigestedwithSstI-DpnI, electrophoresedona0.8%agarosegel,blottedontoaGeneScreen
Plus membrane,and hybridized with the radiolabeled 0.5-kbGAL4gene-containing fragmentofpSG424todetermine the deletionfrequencyof
pGAL4-STD(lower panel). Therecombination index values for thetwopairs of plasmidswerethendeterminedasdescribedinthelegendtoFig.
1;thesearelistedin thetext.The native bandsandthe recombinationproducts for bothcotransfected plasmidsareindicatedontherightof the
autoradiogram. Lanes: 1, pBAaC; 2,pBaC.
resulting in the appearanceof four equimolar isomeric
popu-lations of DNA(7, 9). Althoughagreatdeal has been learned inrecentyears about HSV-1 genomepropagation, the
mech-anism by which the process ofisomerization occurs has been
somewhat controversial. The earliest studies suggested that these inversions were due to homologous recombination be-tween the entire set of inverted repeats flanking the two components (26), but later investigators implicated the a sequence as acis- acting element through which site-specific
recombination eventsoccurred (4,14, 15, 17). Themost recent studies, however, have indicated that thea sequence may be
sufficientbut is not necessary forgenome isomerization. The
overwhelming evidence insupportof this conclusion includesa
description ofa sequence-independent genome isomerization
(12), the characterization of genome recombination events thatare mediated by HSV-1 sequenceswhich lack any
homol-ogytotheasequence (10, 11,20-22, 37), the inability tomap
discrete elements within thea sequencewhich are absolutely
required for the alleged site-specific recombination event (4, 27), the inability ofanisolatedasequenceto promotethesame
highfrequency of inversioneventsasthe intact L-S junction (4,
27), the lack of enhancedintermolecular recombinationevents between L-Sjunction sequences compared with that between
thenonreiteratedsequencesof the viral DNA(35), and finally,
thedemonstrationthat inversion eventsin the HSV-lgenome canbedirectly mediated by the viral DNA replication machin-ery and not asite-specific recombinase and can occur in the
complete absence ofsequence specificity (39).
If HSV- I genome isomerization is indeed the result of
generalized recombination between the invertedrepeatswhich flank the L and S components, the idea that thea sequence acts as a site-specific recombination signal should be
aban-doned. However, ifgenomeisomerization is mediated strictly
by generalized recombination events which occur between
extensive diploid sequences, it is unclear how an element as
small as thea sequencecan function asan apparent hot spot
for recombination by directing homologous recombination eventsoversuchgreatdistances inthe viralgenome(4, 27, 37).
Toresolve thisapparent paradox, anovel quantitative
recom-bination assaywhich utilizes the inversion of bacterial trans-poson sequences by viral DNAreplication-mediated homolo-gousrecombinationwasemployedtoanalyze the role of thea sequence in HSV-1 genome isomerization.
Since thisapproach utilizedashuttleplasmidandnotactual viral DNAas arecombinationtemplate,caremustbe exercised inextrapolating the behavior of theasequencein this situation tothat in the viralgenome.However, shuttle plasmids and viral
DNA are indistinguishable in that they both contain viral
origins of replication, both are replicated to form a linear
concatamericstate,and bothareacteduponbythesamesetof proteins which mediate HSV-1 DNAreplication and recombi-nation. Moreover, the use of transient transfection assays
employing shuttle plasmids minimizes the extent to which a sequence mutantsmightrecombine with and berepairedbya
wild-type a sequence present at the L-S junction, which has beenfrequently observed when theyare incorporated into the viral genome (6, 27). Genomica sequence derivatives which
haveundergone repair will become amplifiedintheDNApool during the extended replication time requiredtogeneratesuch recombinant viruses; this should result in inaccurate assess-mentsof recombinationfrequency, since the mutantcharacter of these inserted sequenceswouldonly be temporary. This is best illustratedby the observation thatpacelement mutations in an a sequence inserted into the viralgenome resulted in a
reduction in inversion frequency which wasonly one-third of thewild-type levels because of recombination withawild-type a sequence (27). However, the set ofpac mutantswhich was
employedin this studyhadareduction in inversionfrequency
ofup to 1/50 of thewild-type levels (Fig. 1 and 2), since they
maintained the integrity of their deletions in a short-term assay.
The HSV-1 a sequence contains two features which were
postulatedtoenhance thefrequency of homologous recombi-nation events in the viral genome: the anisomorphic
confor-mationsof the DR2arrayand thepacl and pac2 elements. An extensive array representing all possible combinations of
ani-somorphic, nonanisomorphic, cleavage/packaging-proficient, andcleavage/packaging-deficient derivatives of theasequence
A
pBaC
NnuI/BanmHI: 7.5kb,4.8kb Native NnrUIBamHt: 6.9kb, 5.4 kb Inversion
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:11.612.98.517.74.213.2]MECHANISM OF HSV-1 GENOME ISOMERIZATION 45
(Fig. 1 and 2) was characterized in this work. Analysis of these subfragments in the pLAR construct, which lacked flanking IS50 sequence homology, established that the enhanced re-combinational activity of the a sequence was not dependent upon the length of thesubfragment tested or its ability to form anisomorphic DNA but only on the presence of both cleavage andpackaging signals. Thisconclusion is especially clear from the results obtained from thepLAR-193 mutants, which dem-onstrate that the absolute minimal sequences which are capa-ble of mediating a cleavage and packaging event (19) are indistinguishable from those required for detection of high-frequency recombination events (Fig. 2). These findings are supported by the earlier results of Smiley et al. (27), who showed that recombinant viral genomes containing insertions of mutant a sequence derivatives underwent a significant decrease in DNA rearrangements when one or the other pac element was deleted from the inserted a sequence. Thus, those mutations which reduced the ability of an a sequence to mediate cleavage and packaging events caused a concomitant decrease in its ability to promote recombination events (27).
Together, these results suggested that the double-strand breakwhich iscreated within thepac2-DR1-pacl arrangement during cleavage and packaging was critical for stimulating the initiation of recombination events. This interpretation was supported by thefindings that the enhanced recombinogenic properties of the a sequence, but not normal replication-mediated recombination, disappeared in cells which were infected by an HSV-1cleavage/packaging mutant (Fig. 8) and that a non-HSV-1 DNAsequence, whose only distinct feature was that it wascleaved in vivo by a restriction endonuclease, possessed the same high recombinogenic activity which was previously thought to be associated only with the a sequence (Fig. 9). The latter observation was especially remarkable in that it demonstrated that the viral cis- and trans-acting com-ponents which were responsible for the special properties of the a sequence could be functionally replaced by a DNA sequence and endonuclease which were both bacterial in origin. These results strongly suggest that a double-strand break, and not specific viral sequences (such as the pac elements of the a sequence), is all that is required for convertingasegment of the HSV-1 genome into a hot spot for homologous recombination.
Thus, recombination between a sequences occurs via the
DNA
replication-mediated
homology-dependent mechanism postulated previously (39) but can be stimulated by the free ends generated during the cleavage and packaging process. Thisconclusion is strengthened by the recent demonstrations that the actual site of crossover between two a sequences undergoing recombination maps in a region of homology which is distinct from the cleavage site (27) and that the ability of thecleavage andpackaging process to enhance recombina-tion is not manifest in the absence of homologous DNA sequences(28). It is therefore unnecessary to evoke additional models suchasasite-specific recombination system to explain the unique recombinogenic properties of the a sequence. Moreover, these results raise the interesting possibility that HSV-1 genome isomerization may be yet another biological system which utilizes double-strand break repair (31) as ameansofrecombination; in this model, free ends generated by
adouble-strand break are able to recognize an intact homol-ogoussequence and utilizeit as a template for repair synthesis. Such a mechanism could also explain the high frequency of gene conversion-like repair which is commonly observed in HSV-1-infected cells (23, 36). It is interesting that under certain conditions, the cleavage and packaging site of the bacteriophage lambda genome can similarly behave as a hot
spot for homologous recombination; this is also thought to involve a double-strand break repair-like mechanism (30, 33, 34).
The analysis of the 440 subfragment and its mutant deriva-tives in the pLAR construct clearly indicated that the ability of the a sequence to enhance the frequency of recombination events in the HSV-1 genome is independent of its ability to form anisomorphic DNA. This is perhaps best illustrated in a comparison of the recombinogenic properties of
pLAR-440
and pLAR-440A1. The latter plasmid exhibits only a residual level of inversion compared with the former, yet they both retain an identical DR2 array (Fig. 1) and therefore an identicalpropensity to form anisomorphic DNA in supercoiled plasmids (Fig. 3). Furthermore, the presence of anisomorphic DNAeven failed to enhance the stimulatory effects of the pac elements on recombination, since the recombination indices for pLAR-440, which contains the two pac elements and an anisomorphic DR2 array, and for pLAR-193, which contains thesame pacelements but lacks the DR2 repeats, were nearly identical (Fig. 1 and 2). Although there has been much speculation as to the role that this DNA conformation plays in recombination and although a host cell nuclease has been identified which specifically cleaves this DNA structure (41), the results in this work suggest that the anisomorphic DNA present in the a sequence, as well as any nucleases which recognize it, plays little if any role in HSV-1 genome isomer-ization. This isperhaps not surprising, since the a sequence is observed to form anisomorphic DNA only in a supercoiled circular DNA template, which represents a situation that is clearly distinct from the normal linear state of the HSV-1 genome. These results do not preclude the possibility that this unusual DNAconformation plays a role in recombination in other biological systems; for example, the DR2 repeat array appears to promote elevated levels of intramolecular recom-bination in E. coli plasmids (25) and may account for the elevated levels of intramolecular recombination observed in supercoiled circular simian virus 40minichromosomes contain-ing the L-Sjunction of HSV-1 (40).