Origin of
Polysaccharide Depolymerase Associated
with
Bacteriophage Infection
PASQUALE F. BARTELL AND THOMAS E. ORR
Department ofMicrobiology,NewJerseyCollege ofMedicine andDentistry, Jersey City, New Jersey07304
Received for publication9 December 1968
Analyses, by construction of phage growth curves, indicated that the
poly-saccharide depolymerasewassynthesizedby Pseudomonasaeruginosastrains Band
BI after infection with phage 2.The kinetics of biosynthesis of the depolymerase
were foundtoparallel closelytherateofformation ofphage-directed virions, and
alterations intheexperimentalconditions of infectionwerereflected byalterations
in theproductionofenzyme.Infection with other Pseudomonasphages,84and1197,
didnotresult in thesynthesis ofdepolymerase. The enzymewasnot detectable in uninfected cultures, and no evidencewas obtained for the existence ofinhibitors
or activators ofenzyme activity in extracts of uninfected or infected cells. The results of experiments employing chloramphenicol or an auxotorphic mutant
(BI arg-) suggested that protein synthesisdenovowasessential for productionof
theenzyme. Variousmutants ofphage2
(pdp1,
pdp2), which alter the synthesisofthe polysaccharide depolymerase, have been isolated. These experimental results
stronglysupportthe role of thephagegenomein thesynthesis of thisenzyme.
Inaprevious paper,evidencewaspresented for
theexistence ofapolysaccharide depolymerasein
phage lysates of Pseudomonas aeruginosa (4).
Re-action of this enzyme with polysaccharide
ob-tained from the slimelayer of this organism
re-sulted inadecreasedviscosityandincreased levels
ofhexosamines, hexoses, andreducingsubstances.
Subsequently, the polysaccharide depolymerase
was purified 1,688-fold from crude lysates by
conventional methods of protein fractionation
(3). Maximal enzymatic activity of the purified
enzymewasfoundatpH 7.5,and,onthe basis of
gel filtration data, the molecular weight was
estimated to be 180,000. We have begun to
in-vestigate the problem of the origin of this hy-drolytic enzyme and its functional role in the virus life cycle. Evidence presented in this paper
strongly suggests that thepolysaccharide
depoly-merase, which isrecovered from crude lysates of
P.aeruginosaafter infection with phage 2, results
from de novo synthesis ofa phage-directed
pro-tein.
MATERIALS AND METHODS
Bacterial and phage strains. P. aeruginosa strainsB
andBI weredescribedpreviously(4). Isolation ofthe
auxotrophicmutantofstrain BI used in these
experi-mentsisdescribed below. Pseudomonas phages 2and
29were isolated fromlysogeniccultures of strainsB
andBI, respectively, and phages 84 and 1197 were isolated from other lysogenic cultures ofP.aeruginosa.
Media.Trypticase SoyBrothandAgar(BBL)were employedascompletemedia forbacterial cultivation and phage propagation. The minimal medium de-scribed by Freese (7) was also employed in certain experiments. It had the following composition: glucose, 5 g; disodium hydrogen phosphate, 7 g; potassium dihydrogen phosphate, 3 g; ammonium chloride, 1 g; FeCl3-6H2, 0.0003 g; CaCl122H20,
0.015 g;magnesiumsulfate,0.25g;distilledwater, 1 liter;agar (whenrequired),15 g.
Isolation ofauxotrophic mutant strain BI arg-. A modification of thedelayed-enrichment technique (9)
was employed for the isolation of a nutritionally deficient mutant of strain BI. An 18-hr culture of strain BI was centrifuged at 3,020 X g for 20 minandwaswashed andresuspendedin 0.1 M sodium phosphate-buffered saline, pH 7.5. Approximately 7 ml of thesuspension,containing 4.8X 107viable cells per ml, wasexposed to ultraviolet (UV) irradiation (Sylvania germicidallamp, G15T8) at anintensityof 120jgw/cmO. Irradiation was carried out with agita-tionof thesuspensionfor 15sec.Viablecountswere
made before and after irradiation. These conditions resulted inasurviving fraction of 0.003 of theoriginal suspension. The exposed bacterial suspension was incubatedinthe darkat roomtemperature for 2 hr, then at 37C with aeration for approximately 5 hr; 0.1-mlamounts of various dilutions were placedon sterilecellulose membranes whichwerepositionedon thesurface of minimal agarplates.After 48 hr of incu-bationat37C,thecellulose membranescarryingthe bacterial coloniesweretransferredtoplates containing complete medium, and the position of the colonies
wasmarked. Theplateswereagainincubatedat37C. 290
on November 11, 2019 by guest
http://jvi.asm.org/
POLYSACCHARIDE DEPOLYMERASE
Isolates which grew oncompletebut not onminimal medium within a 48-hr period werecharacterized as auxotrophs.Qualitative identificationofgrowthfactor requirements was made auxanographically as de-scribed by Lederberg (9).
Mutants ofphage 2. Suspensions of phage2 were treated with 1%S ethyl methanesulfonate (Eastman Organic Chemicals) in solution with 0.5 M sodium acetate and 0.001 M magnesiumsulfate (pH 7.2) as
describedbyFreese (8). At variousintervals oftime afterincubation at 37C,samples ofthismixturewere
diluted 10-fold in Trypticase Soy Broth containing 2% sodium thiosulfate and were plated onlawns of P. aeruginosa BI. Mutants were picked, plaque-puri-fied, andpropagated. Thesemutantsweredesignated pdptosignify alterationin,or absenceof, the phage polysaccharidedepolymerase,accordingtothescheme suggestedby Eisenstark(6).
Production of enzyme in phage-infected cultures. Theelaborationof phage andpolysaccharide depolym-erase by phage 2-infected bacteria was studied in phage growth experiments. Bacteriophage titrations wereperformed bysoftagar-layer, andothergeneral phage procedures were performed according to
Adams (1). Lysogenic cultures were induced by UV irradiation. Bacterialcellswerefirst grown in Trypti-caseSoy Broth to aconcentration of S X 108/ml,and then were centrifuged and resuspended in 0.1 M sodiumphosphate-buffered saline (pH7.5). Approxi-mately7mlof thebufferedsuspension, inanexposed petridish, wasirradiated withagitationat10juw/cm2
for 20sec. Afterirradiation, the bacterialsuspension was added to an equal volume of Trypticase Soy Broth (2X) and wasincubated at 37 C. In other
ex-periments, approximately 5 X 10° cells/ml, in the exponential phase of growth, were usually infected with phage at a multiplicity ofinfection (MOI) of
3 to 4. Adsorption, at 37 C for 5 min with gentle
shaking,wasfollowedbycentrifugationand resuspen-sion ofsedimented cells inanoriginalvolume of pre-warmed broth. Incubation was at 37 C in a shaker waterbath.Sampleswereobtainedatvariousintervals oftime, immediatelychilled, andcentrifugedat6,000 X gfor 10 min at4 C. Supernatantfluids were as-sayedfor free phage and enzyme. The percentageof uninfected cells was measured directly by plating samples of the culture 5 min after infection. The MOI wasdetermined bytheratio ofinput phageto
bacteria (assuming 90% adsorption) or by counting the number of surviving bacterial colony formers (assuming a Poisson distribution of input phage). Values obtained by these two methods agreed to within 15 to 20%o. The intracellular appearance of phageand enzymewas determined bythe disruption ofinfected cells. Atvarioustimes,30-mlsampleswere
withdrawn,chilled, andcentrifuged. The sedimentwas
frozen and during thawingwas ground in 0.5 ml of distilled water containing 0.5 g of alumina powder (Gamal, Fisher Scientific Co.). Extraction was
con-tinued inatotal of5ml of colddistilledwaterfor 15
min. The extracts were then centrifuged, and the supernatant fluids were assayed. Extracts of unin-fected cells were prepared in the same manner. In
certain experiments, the effect of chloramphenicol
(Calbiochem) on depolymerase synthesis was deter-mined by adding chloramphenicol (100 ,ug/ml) to phage-infected cultures at various times.
Estimation of enzyme activity. Assays were based on the release ofhexosamines from slime polysaccharide substrate aspreviously described (4). The amount of hexosamine was determined by the method of Elson andMorgan (11) as modified by Boas (5). Progress curves were constructed, and the initial velocity was determined. A unit of enzyme activity is defined as nanomoles of hexosamine released per minute per milligram of protein (or per milliliter of sample). Astandard ofD-glucosamine hydrochloride (Eastman OrganicChemicals) wasincludedin each hexosamine determination. A method of assay described by Adams and Park (2) was also useful in estimating enzymeactivity.Thedetails, asapplied tothisstudy, have been described (4).Briefly, various dilutions of enzyme were observed fortheir ability to produce a perceptibleclearingofthe bacteriallawn.
RESULTS
Infection by phage2as aspecificrequirement for the synthesis of polysaccharide depolymerase. Formation of the polysaccharide depolymerase
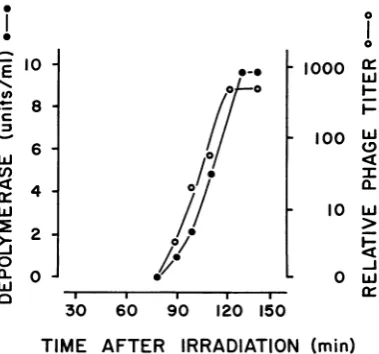
was routinely observed in lysates of P. aerugi-nosastrainB, which carriesphage 2 as prophage, after induction by UV irradiation (Fig. 1). Depolymerase activity was detected 90 min after irradiation and sharply increased to a maximal concentration during the following 30 to 40 min. During the same period oftime,there was almost a 3-log increase in the number of infective phage 2particles released by theinduced cells.
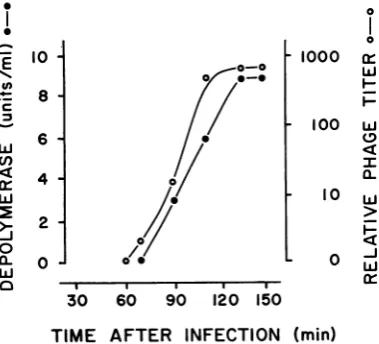
A similar response was observed when P. aeruginosa strain BI was infected with phage 2 (Fig. 2). Depolymerase activitywas not detected
-I0
£0
-E 10
-._
8-c
6-cn
4
-w
_
2-
0-a.0
-J
a
0
l
-000oo
w
-100o
I
a.
10
wOw
30 60 90 120 150
[image:2.487.250.440.437.614.2]TIME AFTER IRRADIATION (min)
FIG. 1.Appearanceof depolymeraseand phage 2 in
culturesofstrain Bafterinduction by UVirradiation
(10,lwpercm2, 20sec).
VOL. 3, 1969 291
on November 11, 2019 by guest
http://jvi.asm.org/
60 min after infection, but a signific
was present in the 90-min sample
increasing to high concentrations.
induced culture, theproduction ofde
was paralleled by a significant incr
number of infective phage, reachin
concentrations within 40 min after
appearance.
It was of interest to determine w]
phage and enzyme were first detec cellularly. The disruption of infecti various time intervals after infectic the presenceofdepolymerase and ms in the 50-min sample (Fig. 3). The
enzyme and maturephage in the
40-was further confirmed by concentr
samples 10-fold before assay. Supern
0
E
C
w
w
-0
a-w a
10
8
4-
2-0
0-0 0 -0
0
0
0
/
00
30 60 90 120 150
TIME AFTER INFECTION
FIG. 2. Appearanceof depolymerase a,
infectedculturesofstrain BI.
0
E
).
CK w
CLI c
w
0
10
-8
-
6-
4-
2-
0-30 50 70 90
ant amount and extracts prepared from uninfected cultures
thereafter of strain BI in the logarithmic phase ofgrowth
As in the and early in the stationary phase of growth
-polymerase were repeatedly examined and found to be
ease in the devoid of depolymerase activity. However,
ig maximal cultivation of strain B eventually resulted in
their initial detectable levelsofdepolymerase. This, of course,
could be related to the spontaneous induction
hen mature of strain B, resulting in the synthesis of both
table intra- phage and depolymerase. Experimentswere also
ed cells at conducted to determine whether phage 2
in-rn revealed fectionmight inducethe synthesis ofa substance
ature phage which, in turn, causes the activation ofa latent
absence of orrepressed enzyme. Whenextracts ofuninfected
min sample cells were mixed, in varying proportions, with
*ating these extracts ofphage 2-infected cells, no increase or
atant fluids decrease of depolymerase activity was observed.
Thus, no inhibitorof the enzyme was detectable
o in extracts of uninfected cells, and no evidence
was obtained to suggest a mechanism whereby
o0 the enzyme was released from a bound state
-1000 cr
u in extracts of uninfected cells upon incubation
- with anextract from infected cells.
F These results suggested that infection with
- 100 w phage 2 was a requirement for the synthesis of
< the polysaccharide depolymerase and that it
I
might
be aphage-directed synthesis
de novo.10 This
hypothesis
waspartially
corroboratedby
> thefollowingexperiments.
Infection of P. aeruginosa strains B and BI
L < by other Pseudomonas phages was studied to
0 W determine whether depolymerase synthesis was a
a specific responsetophage2infection, or whether
itmightbeamoregeneral response of these host (min) cells to phage infection. Therefore, strain BI
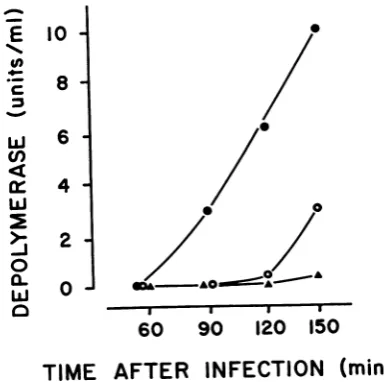
was infected with phage 84. Although high
con-id
phage 2 in centrations of phage 84 were produced, nodepolymerase activity was detectable (Fig. 4).
0
0
*1000 w E
C
*100
< w
I cn
a. <
10 W
S:--;X
-0lJ
>-lJ0c: w
0
10
-8
-6
-
4-2
60 90 120 150
0
-1000 cr
- 100 LLw
I
L:1
- 0 w
TIME AFTER INFECTION
(min)
FiG.3. Intracellularappearanceof depolymeraseand
phage2ininfectedculturesofstrainBI.
TIME AFTER INFECTION (min)
FIG. 4. Absence of depolymerase in phage
84-infectedBIcultures. ,h
O J
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.487.56.246.250.422.2] [image:3.487.58.445.458.635.2]POLYSACCHARIDE DEPOLYMERASE
In other experiments, a lysogenic
strain BI, which carries phage 29 as
wasexposed to UV irradiation. Altk
numbers of phage 29 were syntt
detectable quantities of polysacci
polymerase were present. Similarly, i
strain B with phage 1197 failed t
synthesis of the depolymerase, alti
concentrations of phage 1197 were
(Fig. 5).
The nature of the phage 2-ass4
polymeraseactivity was characterized
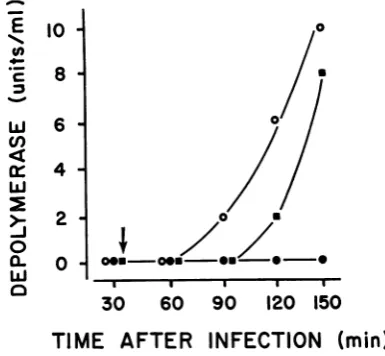
studying the effects of altering the N exposing phage 2 to UV irradiation MOI was drastically reduced, there longeddelay in the appearance ofde
(Fig. 6). At an MOI of3, approximal
T
E
._
w (n
c:
w cn
>111
0 a. w 0
10
-
8-
6-
4-
2-0
-0-0
0
30 60 90 120 150
TIME AFTER INFECTION
(min)
FIG. 5. Absence ofdepolymerase in phage
1197-infectedB cultures.
E 10
w
6-48
w
6
2
0
0-w
0
I
(min)
30 60 90 120 150
TIME AFTER
INFECTION
(min)
FIG. 6. Production of depolymerase in phage
2-infectedB! culturesatMO! of3.0 (0) and0.03 (0).
culture of the bacteria were infected and significant
con-s prophage, centrations of depolymerase were measured
iough great in the 90-min sample. Onthe other hand, when
iesized, no the MOI was reduced to 0.03, approximately 4
haride de- to 5%ofthe bacteria were infected and
depolym-infection of erase activity was not detected until 130 min
o result in after infection. Thus, the time ofappearance of
iough high depolymerase in infected cultures and the
quan-e produced tity of depolymerase produced appeared to be
influenced by the proportion of bacterial cells
ociated de- initiallyinfected by phage2.Suspensionsof phage
lfurther by 2wereirradiated for various periodsoftime,and
40I and of samples were used to infect cells at a ratio of
When the virus tocells of 3 to 4 (Fig. 7). With the lower
was a pro- doseofUV irradiation (5. 5% survival), the
ap--polymerase pearance of depolymerase activity was delayed
tely94% of until 120min afterinfection, and the
concentra-tion ofdepolymerase finally attained at 150min
0 was considerably lower than that of the
unir-0 radiated controls. Athigherdoses of UV
irradia-100 tion
(0.3%
phage survival),
an even morepro-w longed delay was observed before depolymerase
E activity could bemeasured. Based on a Poisson
distribution, approximately 5% of the bacterial
100 W0
cells
were initially infected with the phagesus-< pension receiving the higher UV dose, whereas
a.
approximately 15%
wereinitially
infected with10
w phage exposedtothelowerUVdose.In thecon-> trol cultures, that is, phage not previously
ex-! posedtoUV,approximately95 % ofthe bacterial
oJ cellswere
initially
infected.cr
Dependenceofdepolymeraseactivity
on proteinsynthesisde novo. Two typesofexperimentswere
carried out in order to establish that protein
E
0
4-
F-cn
Cl w
w -J
0
10
4-0 oo S
o A
0Ji
--I I .
60 90 120 150
TIME
AFTER INFECTION
(min)
FIG. 7. Production of depolymerase by strain B!
afterinfectionwithphage 2, UV-irradiatedto asurvival of5.5 X 102 (0) and 3.0 X 10-3 (A). Control cultures were infected with unirradiatedphage (0).
VOL. 3, 1969 293
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.487.44.238.244.634.2] [image:4.487.250.443.427.618.2]synthesis de novo was essential for the
appear-ance of depolymerase activity. In the first
ex-periment, chloramphenicol (100 jg/ml) was
added to cultures of strain BI at the time of
infection withphage2. Theresults (Fig. 8)
indi-cated that synthesis of the depolymerase and of
the phage was completely inhibited. The
ob-served effect could not be attributed to a defect
inthe adsorptiveprocess, since more than 99%
adsorption occurred in the presenceof
chloram-phenicol. When chloramphenicol was removed
from infected cultures after a 10-min exposure,
synthesis of depolymerase and phage was
re-sumed after a slight delay. This effect was also
demonstrable when chloramphenicol was
re-movedfrom infected cultures after a more
pro-longed exposure of 30 min. In other
experi-ments, the addition of chloramphenicol was
delayed until40, 65, and 90 minafter infection.
Depolymerase activity was not detectable in
infected cultures when an inhibitory
concentra-tion of chloramphenicol was added 40 minafter
infection (Fig. 9). However, when the addition
of chloramphenicol was further delayed to 65
min, a small quantity of depolymerase was
detectable, and with greaterdelay (90 min) even
higher concentrations of depolymerase were
detected in culture fluids.
In the second type of experiment, an
auxo-trophic mutant of strain BI was infected with
phage 2 in the absence of the necessary amino
E
4-cn
w
-J
cn
0.
(LL
w
a
10
-
8-4.
2
0
-30 60 90 120 150
TIME
AFTER
INFECTION
(min)
FIG. 8. Effect ofchioramphenicol onappearance of depolymerase in phage 2-infected BI cultures. Chlor-amphenicol (100 ,g/ml) addedat the time of
infec-tion(0); infectedcellswashedfree from
chlorampheni-colafter10 min (A);controlcultures infectedin the
absence ofchloramphenicol (0).
E
._-3
C]
w
w
i-J
0
a.
w
10-
8-6
0
-0
0 0
AO*--A
O::~-I I I T
30 60 90 120 150
TIME
AFTER INFECTION
(min)
FIG. 9. Appearance of depolymerase in phage
2-infected BI cultures treated with chloramphenicol at
various times after infection. Chloramphenicol (100
jug/mi) introduced at 40 mini (A), 65 min (0), and
90 min (0) after infection.
E 0 C
a..
w cn
CL
Ld
10
-
8-
4-0
a
0
0 a
I
'-o+/+/
0030 60 90 120 150
[image:5.487.255.448.56.258.2]TIME
AFTER INFECTION
(min)
FIG. 10. Depolymerase production by an arginine auxotroph (BI arg-) infected with phage 2 in the presence of arginine (2mg/ml; 0),in the absence ot arginine (0), andafter the addition of arginine (I) at 35 min after infection (U).
acid (Fig. 10). WhenstrainBIarg- was infected
in the absence of arginine, no depolymerase
activity was detectable throughout the 150-min
period of observation. However, when the
re-quired amino acid was added to the culture
35minafterinfection, depolymerase activitywas
detectable after a slight period of delay. The
addition of arginine to uninfected,
arginine-O
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.487.253.446.321.499.2] [image:5.487.53.240.390.579.2]POLYSACCHARIDE DEPOLYMERASE
starved bacteriahad no effect inbringing about
theappearanceofdepolymeraseactivity.
Mutants ofphage 2. Plaques of the wild-type
phage 2 are surroundedby large halos when this
phage is plated on lawns of the sensitive
indi-cator strainBI. The halo effect iscaused by the
action of the polysaccharide depolymerase,
which diffuses from the plaque area and
hy-drolyzes the slime polysaccharide of the bacteria
which surround the plaque (4). Additional
evi-dence supporting the role of the phage genome
in the synthesis of the polysaccharide
depolym-erase was obtained through the isolation of
mutantphage. Theexposureofphage2 to ethylm-ethanesulfonate resulted in the isolation of
threetypes of mutantswhichweredistinguishable
on lawns of strain BI (Fig. 11). The wild-type
phage 2 produced a turbid plaqueand was
sur-roundedbya largehalo. Aclear plaquemutant,
Cir, also produced depolymerase and was
sur-rounded by a large halo. However, the mutant
phagedesignatedaspdp1 producedaclearplaque
butwashalo-less. The absence ofapolysaccharide
depolymeraseinlysates producedbythismutant
was confirmedby enzyme tests with slime
poly-saccharide as substrate. Another mutant, phage
pdp2, also produced a clear plaque which
ap-peared to be equivalent in sizeto that ofphage
[image:6.487.43.239.369.590.2]pdp1,but the size of the surrounding halo was
FIG. 11. Plaques produced bymutantsofphage2on
alawnofBI. Wildtype, wild, turbidplaquewithhalo;
mutantclr,clearplaquewithhalo;mutantpdp,, clear
plaque, no halo; mutant pdp2, clear plaque, reduced halo.
smallerthan theclrorwild-type halos, suggesting
an alteration in the quantity or structure of the
synthesizedenzyme.
DISCUSSION
The experimental results which have been pre-sented strongly support the hypothesis that the
polysaccharide depolymerase was synthesized
de novo as a phage-directed enzyme protein after infection.
Analyses by construction of phage growth
curves indicated that the depolymerase was
synthesized by bacterial cultures infected from
without or after induction by UV irradiation.
In both cases, the appearance of enzyme was
preceded by a period of latency during which
no enzyme was detectable. Subsequent to its
intracellular appearance at approximately 50
min, therelease ofenzymeby infectedcells was
observed to occur by 90 min after infection.
In these studies, the kinetics of biosynthesis of
the depolymerase closely paralleled the rate of
formation of phage-directed virions. Although
determinations of phage deoxyribonucleic acid
synthesis have not been made, it is likely that
the polysaccharide depolymerase is a "late"
enzyme.
The depolymerase was not detected in
unin-fected cultures. However, theenzyme and phage
were recovered after cultivation of strain B at
37 C, but thisis attributabletothe spontaneous
induction of strain B which carries phage 2
as prophage. A substrate-controlled induction
of the depolymerase was not observed, nor was
there any experimental evidence for the
exist-enceof inhibitorsoractivators ofenzymeactivity
in extracts ofuninfected or infected cells.
Infec-tion of strains B and BI by other Pseudomonas
phages, 1197 and 84, respectively, did notresult
in the synthesis of a detectable depolymerase.
The induction of strain BI, which carries phage
29 inthe prophage state, byUV irradiation also
failedtoresult in the synthesis ofdepolymerase.
Thus, theproduction ofenzyme appears to bea
highly specific response following infection of
the hostcellbyphage2andcannotbe considered
ageneral responseof thecelltophageinfection.
When experimental conditions were altered to
reduce the number of bacterial cells initially
infectedwithphage, aproportionatedelayinthe
appearance of enzyme and a decrease in the
quantity of enzyme produced, within the time
period ofthe experiments, was observed. Thus,
alterations in experimental conditions of
infec-tion werereflectedby alterations inthesynthesis
ofenzyme. The results ofexperimentsemploying
chloramphenicol or an auxotrophic mutant of
VOL.3, 1969 295
on November 11, 2019 by guest
http://jvi.asm.org/
strain BI indicated that protein synthesis de novo was essential for synthesis of the depoly-merase.
However, the present study does not exclude the possibility that somehost cell protein might be incorporated into the final active form of the enzyme. In thepresent case, in which activity is not detectable in uninfected cells, a similar mechanism could account for the appearance of enzymatic activity afterinfection. Such a mecha-nism would postulate the existence, in uninfected cells, ofa preformed specific inactive precursor. Mathews, Brown, and Cohen (10) addressed themselves to this problem inthecase of deoxy-cytidylatehydroxymethylase. Cells prelabeled by growth in the presence of methionine-methyl-'4C were infected in nonradioactive medium. The hydroxymethylase purified from these cells was virtually nonradioactive and provided direct evidence that the enzyme was synthesized de novo after infection, and that the host cell did
notcontribute any preformed enzyme precursor. However, such findings do not exclude the pos-sibility that the host cell possesses the genetic information for enzyme synthesis and that the
infection itself activates this process through
some process of derepression. In view of our experimental findings, this possibility must be considered remote in the Pseudomonas phage 2 system. Furthermore, theexposure ofphage2 to
ethyl methanesulfonate resulted in the isolation
of variousmutantswhich altered thesynthesis of
polysaccharide depolymerase and thereby
pro-vided strong support for the role of the phage genome inthe synthesis ofthis enzyme.
ACKNOWLEDG MENTS
Thisinvestigationwassupported bygrantGB7891 fromthe NationalScience FoundationandbyPublic Health ServiceGrant AI-08504 from theNational Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages.Interscience Publishers, Inc., NewYork.
2. Adams, M. H., and B. H.Park. 1956. An enzyme produced by a phage host-cell system. II. The properties of the polysaccharide depolymerase. Virology 2:719-736. 3. Bartell, P.F.,G.K. H. Lam, and T. E. Orr.1968. Purification
andpropertiesofpolysaccharide depolymerase associated with phage-infected Pseudomonas aeruginosa. J. Biol. Chem.243:2077-2080.
4. Bartell, P.F.,T.E. Orr, and G.K. H. Lam. 1966. Polysac-charide depolymerase associated withbacteriophage infec-tion.J. Bacteriol. 92:56-62.
5. Boas, N. F. 1953. Methodforthedetermination of hexosa-minesintissues. J. Biol. Chem. 204:553-563.
6. Eisenstark, A. 1967. Bacteriophage techniques, 449-524.In K. Maramorosch and H. Koprowski (ed.), Methodsin virology, vol. 1. Academic Press, NewYork.
7. Freese, E. 1959.Thespecificmutagenic effectof baseanalogues onphage T4.J. Mol.Biol. 1:87-105.
8. Freese, E.1963. Induced and spontaneousmutations in bac-teriophage, p. 3-18.In W. J.Burdette (ed.), Methodology inbasic genetics. Holden-Day, Inc.,Publisher, San Fran-cisco.
9. Lederberg, J. 1950. Isolation and characterization of bio-chemicalmutantsofbacteria. MethodsMed.Res.3:5-22. 10. Mathews, C. K., F.Brown, and S. S. Cohen. 1964. Virus inducedacquisition ofmetabolic function. VII.
Biosynthe-sisde )?ovOofdeoxycytidylate hydroxymethylase. J. Biol. Chem.239:2957-2963.
on November 11, 2019 by guest
http://jvi.asm.org/