JOURNAL OF VIROLOGY, Apr. 1990, p. Vol. 4 0022-538X/90/041764-07$02.00/0
Copyright© 1990,AmericanSocietyforMicrobiology
Mutation of
a
Consensus
Purine
Nucleotide
Binding Site in the
Adeno-Associated Virus
rep
Gene
Generates
a
Dominant Negative
Phenotype for DNA
Replication
NOR CHEJANOVSKYAND BARRIEJ. CARTER*
Laboratory of Molecularand CellularBiology, National InstituteofDiabetes andDigestive andKidneyDiseases,
Building8, Room304, Bethesda, Maryland 20892
Received6September1989/Accepted 21 December 1989
Adeno-associated virus(AAV) containsamultifunctionalnonstructural gene, rep, which isrequired forAAV DNA replication and has pleiotropiceffects on positiveandnegative regulation of geneexpression. All of the parvovirus nonstructural genes contain a region of highly conserved amino acid homology. Within this conservedregionis the consensussequence forapurine nucleotidebinding site.We constructed a mutant AAV having a mutation in this site by converting lysine 340 to histidine. The resulting mutant AAV genome, pNTC23, overproduced themutantRep proteins, indicating that theseproteins areautoregulated. Further-more, the mutant gene was unable to replicate butwas able toinhibitin transwild-typeAAV DNAreplication. Thus, pNTC23 representsa dominant negative mutant of AAV. Theseresults suggest thatrephas separate functional domains important forDNAreplication.
Site-specific mutagenesis canbe used readily togenerate mutations in specific genes. Generally,this results in inacti-vation of the gene product. However, some mutations may resultin overexpression of a mutantpolypeptidewhich can inhibit the activity of the wild-type gene product. Recently,
it was pointed out that such dominant negative mutations may be especially useful for the analysis of regulatory
proteinsor enzymeswhich have severalfunctionaldomains oractivities (22). We have constructedadominantnegative
mutationinthe adeno-associated virus (AAV) rep gene. AAV, like all other parvoviruses, contains a
multifunc-tionalgene, rep,which is required for AAV DNA replication
(7, 8, 21, 31, 33, 35) and has pleiotropic effects on positive
andnegative regulation of gene expression (25, 26, 36, 38). The AAV rep gene uses two transcription promoters to encodefouroverlapping Repproteins:Rep78 and Rep68 are
encodedby transcripts fromtheP5promoter,and Rep52 and
Rep4Oareencoded by transcripts from the P19 promoter (29,
38). Rep78 and Rep52 are encoded by unspliced mRNAs, whereas Rep68 and Rep4O have altered carboxyl termini,
reflecting splicing ofthetranscripts (29, 38). Rep78 or Rep68 isrequired for AAV replicating-form (RF) DNA replication
apparently by bindingtothereplication origin (1, 24).Rep52
andRep4Oarenotrequired for RF DNA replication but may
be required for the accumulation of single-stranded DNA
(ssDNA) and infectiousparticles (8).
Allofthe parvovirus nonstructural genes contain a region ofhighlyconservedamino acid homology (2, 9, 32). Notable within this region is the consensus sequence G(X)4GKT/
S(X)5_6I/L/V
for a purine nucleotide binding site present in a variety of other eucaryotic and procaryotic proteins,includ-ingthelargeTantigens of simian virus 40 and polyomavirus (2,
5,
9, 13, 16, 19). In simian virus 40 and polyomavirus an ATPase activity that is associated with this domain is required for viral DNA replication (10-12).In AAVtheputative purine nucleotide binding site (2, 33) is common to all four Rep proteins. In this work, we constructed a mutant AAV having a mutation in this site.
* Correspondingauthor.
Thecodonforlysine340in theconsensusnucleotidebinding
sequence was changed to histidine. The resulting mutant AAVgenome (pNTC23)overproducedthemutantRep
pro-teinsbutwasunabletoreplicate.Themutantgenewasable to inhibit in trans wild-type AAV DNA replication. Thus,
pNTC23 representsa dominantnegative mutantof AAV.
MATERIALS ANDMETHODS
Cells andviruses. An established humanembryonic kidney
cellline transformedbyadenovirus 293-31(293 cells; 18)was grown at 37°C in 5% CO2 in monolayer cultures in Eagle
minimalessentialmediumsupplementedwith antibiotics and 5% fetal calfserum. Human adenovirus type 5 and AAV type 2 (AAV-2) were grown and assayed as described previously (6).
Plasmid construction and mutagenesis. Plasmid construc-tion andmutagenesis wereperformed asdescribed in detail elsewhere (7, 8). Plasmid pNTC244 contains the
wild-type
AAV-2genome inserted viaBglII linkers(27)atthe BamHI
sitein thepolylinkersequenceof thephagemid pTZ19U
(8).
pNTC3 is an amber mutant of pNTC244 having a point
mutation in the rep geneatmapposition22(7).pNTC23was
generated from pNTC244 by oligonucleotide site-directed mutagenesis of thewild-typeAAV-2 genomewiththe
oligo-nucleotide 5'-ACTACCGGGCACACACCAACATC-3' by
use of the Muta-gene system (BioRad Laboratories, Rich-mond, Calif.) as previously described (7, 8). pNTC41 was
produced by deleting the SstI fragment between the SstI sites in the polylinker of pNTC244 and at nucleotide 814 (mapposition 17.4)of the AAV-2 sequence(8).Thisdeletion
removes theP5 transcription unit but leaves the P19
tran-scriptionunit intact. pNTC41awasgenerated frompNTC41 by introducing the same amber mutation as in pNTC3 (8). pNTC8 was constructed from pNTC23 by deleting the p5 transcription unit by use of the SstI sites as described for pNTC41. pTS18 isaRep-p40-catconstruct(36,37) grown in Escherichia coli HB101 andpurifiedby standardprocedures
(28, 36). Stocks of pTZ19U and the pNTC plasmids were grownin E. coli MV1190 and purifiedby alkaline lysis and
1764
on November 10, 2019 by guest
http://jvi.asm.org/
Gly GGG
1338 1340
1 1
Pro Ala Thr Thr Gly Lys Thr Asn lle
CCT GCA ACT ACC GGG MG ACC AAC ATC
., ,~~~~~
I I
CAC
-- His -'
5 19 40 "
2 10
120
301,4148
50Rp I 1 I . 1,
Rop78 -11*
60 70 80 90 , 100
I I I
I
96'9 4.2kb
Rep68 :|
Rep52
Rep4O
I ;'993 ; 2183
* II
aI II
993 ' 1 2252
1
.1907
I 3.9kb
*, 3.6kb
" 3.3kb
pNTC244
M*.
pNTC23
E.-pNTC3
pNTC41
pNTC41a
pNTC8
a
a
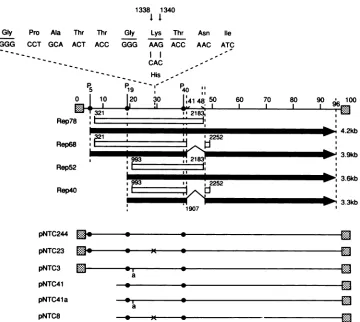
FIG. 1. AAV rep gene structure and consensuspurine nucleotide binding site sequence. The AAV-2 genome (33) is shown on a scaleof 100mapunits(1unit is approximately 47nucleotides).Control regions are shown asstippledboxes (DNAreplication origins;20) andsolid circles(p5,Plg,andP40transcription promoters). Heavy arrows show p5 orp19mRNA,with theintron (mapunits41to48) shownbythe carets and thepolydenylation siteatmapunit 96 shownby the arrowheads.The
p4o
transcriptswhich codeforcapsid proteins andthe minor RNA species derived by alternate splicingareomittedfor clarity. The coding regions forthefourproteins, Rep78, Rep68, Rep52,andRep40 (29, 38),areshownbyopenrectangularboxes. Theputative purine nucleotide binding regionwith consensusamino acids underlined(2)atmap position28.6is indicatedatthetop, as arenucleotides 1338 and 1340, which were altered to convert the consensuslysine340codonAAG to ahistidinecodon,CAC. Inthelowerportionareshown theregions ofAAV-2 DNAcontainedin severalplasmids. pNTC244, pNTC23, andpNTC3 containtheentireAAV-2genome. InpNTC41, pNTC41a,andpNTC8,nucleotides 1to814 of AAV-2aredeleted. pNTC23and pNTC8 contain the mutation in the purine nucleotide binding site indicated by x. pNTC3 and pNTC41a contain a nonsense mutation (indicatedby "a") atnucleotide1033generated by changing a serine codon, UCG,to an ambercodon, UAG(7). Thenucleotidenumbering scheme is thatofSrivastavaetal. (33). kb, Kilobases.pZ523columns(5Prime-3Prime, Inc., West Chester, Pa.) in accordance with manufacturerprotocols (7, 8, 28).
DNA transfectionandviralDNAreplication. 293cells(5 x
105/35-mm
dish)were transfected by thecalciumphosphateprecipitation method (35). For AAV replication, the cells were infected with adenovirus type 5 (5 PFU per cell) 1 h prior to transfection as described before (27, 35). For indi-vidual transfection experiments, the total amount ofDNAin each transfection mixture was adjusted to the same level with control plasmid pTZ19U.
IntracellularAAV DNAreplication forms wereextracted
40 h aftertransfection bythe modified Hirt sodiumdodecyl sulfate-high-salt lysis procedure (27, 35). The DNA forms were analyzed by agarose gel electrophoresis followed by
blotting to nitrocellulose filters and hybridization with
[32P]AAV
probes radiolabeled by the random priming pro-cedure (17). The amount of radioactivity hybridized wasquantitated byscintillationcountingof individual bands from theblots.
Proteinanalysis. Cell lysates foranalysis ofRepproteins
wereprepared 48 h aftertransfection ofadenovirus-infected
293 cells and electrophoresed in 12% polyacrylamide gels. AAV Rep proteinswere detected with anti-Repantiseraby
the immunoblotting procedure (29, 38) exactlyasdescribed
previously (8).
CAT assays. 293 cells
(106)
were transfected with 1jig
ofpTS18,aRep- AAVp40-catvector(36, 37),and upto10 ,ug ofacomplementing
plasmid.
Cell extracts wereprepared
at 48 h after transfection and assayed forchloramphenicol
acetyltransferase (CAT)
activity
as describedpreviously
(36).
RESULTS
Mutagenesisof theputativenucleotidebindingsite in therep reading frame. The structure of the AAV genome and the
organization ofthe rep gene are shown in
Fig.
1. The rep gene is transcribed fromtwodifferent promoters,designated
atp5andP19(at map
positions
5 and 19,respectively).
Four Repproteins were identified withspecific
anti-Rep
antibod-ies (29, 38). The predominant
proteins
wereRep78
andNo
1 fth4 1 1 1 . --l-
p-"i-)
1
-
u--_
>s 2252
--9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.133.493.66.388.2]c\j
Rep78_
Rep68-
Rep52-
Rep4O-1
2 3 4
5
6
.0
CO '- :
c'ij
-to~
<...
MP
.1
,A [image:3.612.366.514.72.289.2]."p
FIG. 2. Production of Rep proteins by mutant AAVgenomes.
Adenovirus-infected 293 cells (5 x 105 cellsper35-mmdish) were
transfected with the AAV plasmidsdesignated by the numbersatthe top(thepNTC designation is omitted). Ad indicates control adeno-virus-infected cells. Proteinextractswereanalyzedby immunoblot-ting withananti-Rep antibody.
Rep52, whereas Rep68 and Rep4O were produced at much lower levels (29, 38; Fig. 2).
The consensus purine nucleotide binding site in the rep
reading frame (2) was at map position 28.6 and was thus
common to all four Rep proteins (Fig. 1). To produce a
mutation in the putative nucleotide binding site, we
intro-duced atwo-base change in the lysine 340 codon (Fig. 1).
Thus, the codon AAG was convertedto CAC (nucleotides 1338 and 1340), coding for histidine, to generate mutant pNTC23.
SeveralothermutantAAVplasmidsare alsodiagrammed
in Fig. 1. pNTC3 contains an amber mutation at map
position 22. pNTC41, pNTC41a,andpNTC8 areanalogous
topNTC244, pNTC3, and pNTC23, respectively, but retain onlytheP19transcription unit.
Repprotein production by AAVmutants. The expression ofthe Rep proteins from mutant and wild-type AAV ge-nomes wasdetermined by immunoblot analysis ofextracts prepared fromadenovirus-infected 293 cellstransfected with the appropriate plasmids. pNTC23 produced all four Rep proteins at a higher level than did the wild-type AAV plasmid pNTC244, suggesting that the production of Rep proteins is autoregulated (Fig. 2). It is possible that the overproductionby pNTC23representsanincreasedstability
of Rep proteins. However, we have previously shown by
pulse-chaseanalysisthatRep proteinsarehighlystable(30).
Figure 2 also shows, as wereportedpreviously for pNTC41
andanalogous plasmids (8, 29, 38), that mutationordeletion
of the p5promoterortheamino terminus ofRep78orRep68
resulted in the overproduction of Rep52 and, to a lesser
extent,ofRep4O.Incontrast,pNTC8, which is analogousto pNTC41 but which has thesame mutation intheconsensus
nucleotide binding site as pNTC23, did not overproduce Rep52. This resultsuggeststhat the expression ofRep52 (or Rep4O) may be independently autoregulated in addition to
being regulated by Rep78 or Rep68. Elsewhere we showed
thatpNTC3and pNTC41a,whichhaveanamber mutationat
mapposition22, produced no Repproteins (7, 8).
Blockage of AAV DNA replication by a mutation in the
consensus nucleotide binding site. The Rep proteins are
essentialfor AAVDNAreplication(7,8, 21,24, 31, 35). The effectof themutation intheputative nucleotide binding site of rep was analyzed by isolating intracellular replicating
AAV DNA from adenovirus-infected 293 cells transfected with pNTC23 or pNTC244. pNTC23 was defective for
[image:3.612.104.256.73.200.2]23
244
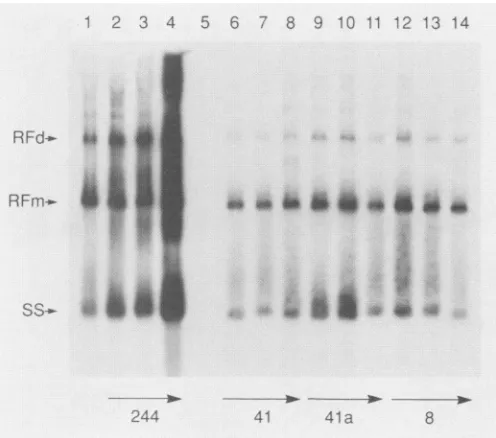
FIG. 3. Replication of AAVgenomes. Adenovirus-infected 293 cellsweretransfected withpNTC244orpNTC23 DNAasfollows. Intracellular AAV DNA was extracted, electrophoresed in an agarose gel, and detected by Southern blotting with an AAV [32P]DNA probeasdescribedin Materials and Methods. Lanes: 1, 6 jigofpTZ19U;2,0.1 ,ug of pNTC23;3, 1 jigofpNTC23; 4, 6 jig of pNTC23; 5, 0.1 ,ug of pNTC244; 6, 1 ,ug of pNTC244. All transfec-tion mixtureswereadjustedto6 ,ug with control plasmidpTZ19U. RFd, RFm, and SS indicate replicating dimer DNA, replicating
monomerDNA, andprogenyssDNAs,respectively.
replication,and the normal AAV DNA intracellularspecies (4, 34), includingmonomeranddimer RFs, (RFm andRFd,
respectively) andprogenyssDNA, were notobserved(Fig.
3). We reported elsewhere that pNTC3, pNTC41, and pNTC41a were all unable to replicate AAV DNA (7, 8). Similarly, pNTC8, which is both an ori mutant and a rep
mutant, like pNTC41 and pNTC41a, didnotreplicateAAV DNA(datanot shown).
Inhibition of AAV DNA replication bymutantRepproteins. Detection of AAV DNA replication in adenovirus-infected 293 cells is a sensitive assay, and RFs resulting from transfectionof less than 10ngofanAAV-containing plasmid canbe observed(unpublished results).We used thisassayto determine if the mutant Rep proteins from pNTC23 could inhibit wild-type DNA replication. AAV DNA replication
was easily detected when 50 ng of wild-type AAV, pNTC244,wastransfectedintoadenovirus-infected293cells (Fig. 4, lane 1). Increasing amounts of wild-type AAV plasmidincreased theamountof intracellular DNAsynthesis (Fig. 4,lanes 2to4). However,when50ngofpNTC244was
cotransfected with increasing amounts ofpNTC23, AAV DNA replication was severely inhibited (Fig. 4, lanes 8 to 10). Indeed, significantinhibitionby pNTC23 wasseenwith
only a 1:1 ration of pNTC23 to pNTC244 (Fig. 4, lane 8). When the amber mutant pNTC3 was cotransfected with pNTC244 (Fig. 4, lanes 5 to 7), RF DNA levels were
decreasedbyno morethan2-fold,evenata20-foldexcessof themutant.Sincethe amberplasmid pNTC3 suppliesnoRep
protein, the drastic inhibition by pNTC23 must have been
duetothemutantRep proteinsandnot totheeffect ofadding
additional AAVtemplates containing replication origins. Additionalexperiments(Fig. 5, lanes 7to12) showed that inhibitionby pNTC23wasnotobserved when theamountof
RFd
RFm
Ss
VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
12 3 4 5 6 7 8 9 10 11 1 2 3 4 5 6 7 8 9 10 11 12
RFd
RFm4
Ss
-.4~~~~~~~~~~~~~~~~~~~~~~~~
3 23
FIG. 4. Inhibition of wild-type AAV DNA replication by a trans-dominant mutant. Adenovirus-infected 293 cells were trans-fected with0.05 jigof pNTC244(lane 1) and cotransfected with an additional0.05, 0.1, or 1jigof pNTC244(lanes 2 to 4), pNTC3 (lanes 5 to 7), or pNTC23 (lanes 8 to 10), respectively. Lane 11 shows adenovirus-infectedcellstransfected withthe controlplasmid alone. Alltransfection mixtureswereadjustedto 1 ,ugwithcontrol plasmid pTZ19U. AAV DNA was extracted and analyzed as for Fig. 3. RFd, RFm, and ssDNA species are indicated. The additional species in lanes 7 and 10 areinputpNTC3 andpNTC23DNAs,respectively. mutantplasmid was decreased to about one-tenth that of the wild-type plasmid. Similarly, inhibition could be overcome when the amount of wild-type plasmid was increased 10-fold (Fig. 5, lanes 2 to 5). Also, pNTC23 could be complemented forreplicationwhen wild-typerep was suppliedby
cotrans-fectinga10-to20-foldexcessofanOri- AAV plasmid (data not shown).
Themutation in pNTC23 affected both the Rep78 or Rep68 and Rep52 or Rep4O reading frames. Thus, it was conceiv-able that the mutant Rep52 or Rep40 protein alone might inhibitwild-typeAAV DNAreplication.This possibility was tested by cotransfecting the wild-type AAV plasmid with
pNTC8, aswellaswith pNTC41 and pNTC41a(Fig. 6). No
significantinhibition of AAV DNAreplicationwasobserved with any of these plasmids, even at a 20-fold excess over the wild-type plasmid, pNTC244 (Fig. 6).
The datafrom
experiments
similartothose inFig.4and 6 werequantitated by cutting the blots afterhybridizationand determining theamountof[32P]DNA
probe annealedto the RFmspecies. A 1:1ratio ofpNTC23topNTC244 produced20-fold inhibition, whereas even a 20:1 excess of plasmids
pNTC3, pNTC41, pNTC41a, andpNTC8producednot more than2- to2.5-fold inhibition(Fig. 7). This lower inhibitionby
the latter plasmids may reflect simply a dilution effect of
providing a large number of substrates (i.e., replication origins) thatpotentially bind Rep proteins. This appears to be thecase evenforpNTC3,whichcanbecomplemented by
RFd-
RFm-em.
w
a
.6
pNTC244
pNTC23FIG. 5. Effect of relative concentration ofwild-type and mutant plasmidsonAAV DNAreplication. Adenovirus-infected 293 cells weretransfected with DNA as follows: lane 1, 0.1 jigofpNCT23; lanes 2to5,0.1 jig of pNTC23 plus0.1jLg(lane2), 0.5 ,ug(lane 3), 1jig(lane4), or 2 ,ug(lane5)ofpNTC244;lane 6,pTZ19U control DNA; lane 7, 0.05 jig of pNTC244; lanes 8 to 12, 0.05 jig of pNTC244 plus0.1 jig(lane 8), 0.05 ,ug(lane9), 0.01 jig (lane 10), 0.005jig(lane11),or0.001 ,ug(lane 12) ofpNTC23.Alltransfection mixtureswereadjustedto 2jigof total DNA with pTZ19U DNA. AAV DNAreplicationwasanalyzedasfor Fig.3.
the wild-type plasmid to generate mutant genomes that competefor, but donotsupply, Rep proteins. Data similarto those inFig. 7werealso obtained when the radioactivityin RFd or ssDNA was quantitated, except that ssDNA was moresensitivetoinhibition thanwasRFd DNA(e.g.,Fig.4, lanes 5to 7). Thisprobablyreflectsacascade effect in that decreased ssDNA accumulation may result both directly
fromdecreased RFd DNA synthesis andperhaps indirectly
from decreased wild-type protein synthesis. Both capsid proteins and the Rep52 or Rep4O protein are apparently required for efficientssDNA accumulation(8, 21, 35).
trans-regulation by pNTC23-encoded Rep proteins. Previ-ous studies showed that in uninfected 293 cells the Rep proteinsexert pleiotropic effects on the level ofexpression fromchimericp40-catconstructs(36, 37).Thus,Repproteins increasedbytwo- tothreefold the accumulation ofchimeric
P40 RNA
transcripts
but inhibited theexpression
of CATactivity (37). The ability of the mutant Rep proteins from
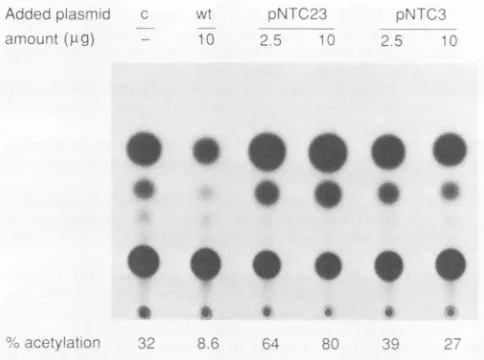
pNTC23 to trans-regulatep40-cat expression was analyzed
with thechimeric Rep-p40-catconstructpTS18(Fig. 8) (36, 37). Cotransfection ofwild-type pNTC244 with the reporter
plasmidresulted inafourfoldinhibitionof CATactivity (Fig.
8). In contrast, with pNTC23 there was about a twofold
stimulation, whereas the rep amber mutant
pNTC3
had a null effect (Fig. 8). This result suggests that the mutantpNTC23may haveretainedthetrans-activation property but
apparentlyhas lostthe negative regulatoryeffect of rep.
DISCUSSION
Alterationof the
putative purine
nucleotidebinding
site in the AAV rep gene resulted in a mutant,pNTC23,
whichoverproduced all four Rep
proteins
but was defective for AAV DNA replication. Furthermore, the mutantproteins
SS-l
I
a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.78.290.73.349.2] [image:4.612.316.561.74.277.2]41..8_._ . ...
[image:5.612.55.303.64.283.2]41 41ta 8
FIG. 6. Replication of wild-type AAV DNA in thepresenceof
mutant AAV genomes producing only p19-encoded Rep proteins.
Adenovirus-infected 293 cells were transfected with 0.05 ,ug of
pNTC244 (lanes 1 to 4 and 6 to 14) and cotransfected with an
additional 0.05, 0.1,or1,ug, respectively, of pNTC244 (lanes 2to4), pNTC41(lanes 6to8),pNTC41a (lanes 9to11),orpNTC8 (lanes 12
to 14)or1 p.gof controlplasmid pTZ19U (lane 5). All transfection
mixtureswereadjustedto1 pLgwith the controlplasmid. AAVDNA replicationwasanalyzed asfor Fig. 3.
wereinhibitory for wild-typeAAVDNAreplicationintrans.
However, this inhibitionwas overcomebya 10-fold excess
ofwild-type AAVgenomes.Presumably, thisresult reflects
theneedfor increased wild-type Rep proteininthepresence
ofoverproduced mutantprotein. pNTC23fulfills the criteria foradominant negativephenotype (22), suggestingthat the putative purine nucleotide binding site may be important
both for AAV DNA replication and for some of the
pleio-tropic properties ofrep.
TheRep78orRep68 proteinisnecessary(7, 21, 31,35, 36) andsufficient (8) forinvivo amplificationof AAVduplexRF DNA. Consistent with this, Rep78 or Rep68 binds
specifi-cally to the AAV palindromic termini which constitute the replication origins (1, 24). Furthermore, recent evidence from in vitro assays indicates that the Rep78 or Rep68
proteinappears tocontain an ATP-dependent, site-specific,
strand-specific nuclease activity involved in the resolution of theAAVDNA termini(32a). In in vitroassays,resolution of
thetermini butnotbinding of Rep78orRep68tothe origins requires ATP (24; Snyderet al., in press). Thus, the DNA-negative phenotype ofpNTC23 provides in vivo evidence consistentwiththepossibility thatafunctional ATP binding
site inRep78 orRep68is requiredforRepfunction inAAV
DNA replication. That the DNA-negative phenotype of pNTC23 isdominant intrans suggeststhatotherfunctional domainsarestill presentin the mutantproteins.
The most obvious explanation ofthe dominant negative phenotype of pNTC23for AAV DNA replication is that the mutantRep78 and Rep68 proteins have retainedtheir origin-binding ability but havelost their enzymatic activity requir-ingATP. Thus,themutantproteinscouldcompete withthe wild-type protein. However, inhibition could also be medi-ated by protein-protein interactions between mutant and wild-type Rep proteinsorbyinteraction withcellularfactors
5
1
la
pNTC23
100 1000
AddedDNA(ng)
FIG. 7. Effect of cotransfectedmutant AAVgenomesonAAV DNAreplication. The data werederived from experiments
analo-goustothose inFig. 4 and 6. Theamountofradioactivityhybridized
tothe RFmspecies afterblottingwasdeterminedby cutting the filter and liquid scintillation counting. The effect of cotransfecting
ge-nomesis shownby plotting theamountsofradioactivitypresentin the RFm band as a percentageof the control amount present in adenovirus-infected293 cells transfected with 0.05,ugofpNTC244.
important forAAV replication. It is also possible that the dominantphenotype of pNTC23 could be mediated by the mutantRep52orRep4O proteinthatwasalsooverproduced. This possibility is less likely, however, because a 20-fold
excessofpNTC8,whichproducedmutantRep52andRep4O proteins but not Rep78 and Rep68 proteins, showed no
specificinhibitoryeffect.However, pNTC8didnot
overpro-duce the mutantproteins, so evenatahigh excess ofinput
genomesit maynotgenerate sufficient mutantprotein. The overproduction of the Rep proteins by pNTC23 providesthe first directevidence that theexpressionofthese proteins is autoregulated. This regulation is complex. We previously reported that Rep78 orRep68 apparently
down-regulates the expression ofRep52 or Rep 40 (29, 38). The
present report suggests that Rep78 and Rep68 may also down-regulate their own expression. Preliminary evidence (B. Antoni and B. Carter, unpublished data) suggests that this negative regulation is mediated at least in part at the level of RNA accumulation. Recent evidence also suggests thatRep proteinsdownregulatetheexpressionof CATfrom thep5 or P19 promoter and the expression ofRep mRNA
from a simian virus 40early promoter (3). In contrast, in adenovirus-infected KB cells, Rep proteins apparently
in-1 2 3 4 5 6 7 8 9 10 11 12 13 14
5001
S-0-24 2
0
0
0
10
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.320.554.68.400.2]Aa *cilc]p,uo
1p3s
:; ; LITERATURE CITEDI2~t %'i ~ C w
s1.
tot !Ashktorab, H.,C and A.Srivastava.1989. Identification of nuclear proteins that specifically interact with adeno-associated virus type 2 inverted terminal repeat hairpin DNA. J. Virol. 63: 3034-3039.2. Astell, C. R., C. D. Mol, and W. F. Anderson. 1987. Structure andfunctional homology of parvovirus and papopavirus poly-peptides.J. Gen. Virol.68:885-893.
3. Beaton, A.,P.Palumbo,and K.I. Berns. 1989.Expressionfrom * * *^ * the adeno-associated virus p5 and p19 promoters is negatively W W regulatedin transbythe repprotein.J. Virol.63:4450-4454.
4. Berns, K. I., and W. W. Hauswirth. 1984. Adeno-associated virus DNA structure and replication, p. 1-31. In K.Berns(ed.),
5 5 ^ 5^ ^ Theparvoviruses. Plenum Publishing Corp.,New York.
@
___w_ 5. Bradley, M. K., T. F. Smith, R. H. Lathrop, D. M. Livingston, and T. A. Webster. 1987. Consensus topography in the ATP * * * * * ,,, binding site ofthesimian virus40andpolyomaviruslarge tumor
antigens. Proc. Natl. Acad.Sci. USA84:4026-4030. .4Q.
lis 6. Carter,B.J., C. A.Laughlin,L. M.de laMaza,and M.Myers.
1979. Adeno-associated virus auto-interference. Virology 92: FIG. 8. trans-Regulation by rep of expression from a p40-cat 449462.
plasmid (pTS18) in uninfected293 cells. Cells were transfected with 7. Chejanovsky, N., and B. J. Carter. 1989. Replication of a human pTS18(1 ig)and10,ugofwild-type (wt) DNA (pNTC244) or 2.5 or parvovirus nonsense mutant in mammalian cells containing an 10 ,ugofmutant DNA (pNTC23 and pNTC3) or 10 ,ug of control inducible amber suppressor. Virology 171:239-247.
plasmid pBR322 (c). CAT activitywas measured 48 h after trans- 8. Chejanovsky, N., and B. J. Carter. 1989. Mutagenesis of an fection. The percent acetylation in each reaction is shown at the AUG codon in the adeno-associated virusrepgene: effects on
bottom. viral DNA replication. Virology 173:120-128.
9. Chen, K. C., B. C. Shull, E. A. Moses, M. Lederman, E. R. Stout,and R.C. Bates.1986.Complete nucleotidesequenceand genome organization ofbovine parvovirus. J. Virol. 60:1085-creased the transcription of mRNA from all three AAV 1097.
promoters(26). Furthermore, we previously showed that for 10. Clark, R., D. P. Lane, and R. Tjian. 1981. Use of monoclonal
acatgeneexpressedfromanAAVP40promoterin 293 cells, antibodies as probes of simian virus 40 T antigen ATPase
Rep79 orRep68 simultaneously mediateda modest activa- activity. J. Biol. Chem. 256:11854-11858.
tion of two- tothreefold inp40-cat mRNA expression but a 11. Clark, R., K. Peden, J. M. Pipas, D. Nathans, and R. Tjian.
fivefolddecrease in netCAT activity, apparently because of 1983. Biochemical activities of T-antigen proteins encoded by
a moredrastic10-to20-foldposttranscriptional inhibition of simian virus 40 A gene deletion mutants. Mol. Cell. Biol.
CAT
more y h eprmn n i.8sgessta e 3:220-228.CAT activity. The experiment in Fig. 8 suggests that Rep 12. Clertant, P., and F. Cuzin. 1982. Covalent affinity labeling of
proteinsfrompNTC23 may have lost the negative regulation periodate-oxidized[a-32P]ATP of the large-T proteins of poly-but have retained the positive activation of p40-cat expres- oma and
SV40
viruses. J. Biol. Chem. 257:6300-6305.sion. 13. Clertant, P., andI. Seif.1984. A common function for polyoma
In summary,ourresults, together withthoseofothers (24, viruslargeT andpapillomavirusEl proteins.Nature(London)
32a), suggest that the Rep proteins may have separate 311:276-279.
functional domains, including an ori binding domain and 14. Cole, C. N., J. Tornow, R. Clark, and R.Tjian. 1986. Properties
probablyanATPbindingdomain. Apreliminary analysis(J. of the simian virus 40(SV40)large T antigens encoded bySV40
Trempe,R. Owens,N. Chejanovsky,andB. Carter, unpub- mutants with deletions in gene A. J. Virol.
57:539-546.
lisheddata)suggeststhatthemutantRep78protein produced 15. Denhez, R., B. Heiman, L. d'Anriol, T. Graf, M. Coquilland,J.
bypNTC23isafunctionalAAV oribindingprotein. Defin- Coll, F. Galibert, K. Moelling, D. Stehelin, and J. Ghysdase.
by p
oTC23
iS atunctilonal
AAVourifinding
protein.Defin-
1988. Replacement ofIys622in the ATP binding domain ofpl00
itiveproofofthis will require purification and characteriza- gag-mil abolishes the in vitro autophosphorylationof the protein tion of mutant Rep proteins. Nevertheless, these observa- and biological properties of thev-miloncogene of MH2 virus. tions indicate that rep may beanalogous to other viral genes EMBOJ. 7:541-546.
such as the simian virus 40 T-antigen gene, which has 16. Doolittle,R. F. 1986. Proteinsequencedata banks: the continu-separatedomainsmediatingoribinding andATPbindingor ing search for released structures. Protein Eng. 1:15-27. ATPase (10-14, 19). Finally, we note that in a variety of 17. Feinberg, A. P., and B. Vogelstein. 1982. A technique for
protein kinases,notdirectlyrelatedtoRep butcontainingan radiolabeling DNA restriction fragments to high specific activ-ATPbinding site, mutation of the conservedlysine specifi- ity. Anal. Biochem.
132:6-13.
caTPy
b.'. . c eo. 18. Graham, F. L., J. Smiley,W. C.Russell, andB. Naiva. 1977. cally alters biological activity. For instance, conversion of Characteristics of a human cell line transformed by DNAlysine721 ofthe humanepidermal growthfactor receptorto adenovirus type 5. J. Gen. Virol. 36:59-72.
alanine abolished tyrosine kinase
activity,
altered receptor 19. Griffin, B. E. 1981. Structure and genomic organization ofSV40recycling,and abolishedsignal transduction(23). Similarly, and polyoma virus, p. 61-124. In J. Tooze (ed.), Molecular the conversion of lysine 623 of the retroviral mH2 v-mil
biology
ofDNAtumorviruses. ColdSpringHarborLaboratory, oncogene protein,plOOga-"',
to methionine abolished Cold Spring Harbor, N.Y.serine-threonine in vitro autophosphorylationandbiological 20. Hauswirth, W. W., and K. I. Berns. 1977. Origin and
termina-properties of the protein (15). tion of adeno-associated virus DNA replication. Virology 79: 488-499.
21. Hermonat, P., M. A. Labow, R. Wright, K. I. Berns, and N. ACKNOWLEDGMENTS Muzyczka. 1984. Genetics of adeno-associated virus: isolation andpreliminarycharacterization of mutants of adeno-associated Wethank N. Muzyczkaand K. Bernsforuseful discussion and virus type 2. J. Virol. 51:329-339.
communicationofdatapriortopublication. 22. Herskowitz,I. 1987. Functional inactivation ofgenesby
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.57.299.71.251.2]CHEJANOVSKY AND CARTER
nantnegative mutations. Nature(London)329:219-222.
23. Honegger, A. M., T. J. Dull, S. Felder, E. Van Obberghen, F.
Bellot,D. Szapary, A.Schmidt, A. Ullich, and J. Schlessinger. 1987. Point mutation atthe ATPbinding site of EGF receptor
abolishes protein-tyrosine kinase activity and alters cellular routing. Cell51:199-209.
24. Im, D.-S., and N. Muzyczka. 1989. Factorsthatbindto adeno-associated virus terminalrepeats. J.Virol. 63:3095-3104. 25. Labow,M.A.,L. H. Graf,Jr., andK. I.Berns. 1987.
Adeno-associated virusgeneexpression inhibits cellular transformation
byheterologousgenes. Mol.Cell. Biol. 7:1320-1325.
26. Labow, M. A., P.L. Hermonat,andK. I. Berns. 1986.Positive andnegative autoregulation of the adeno-associated virustype2
genome. J.Virol.60:251-258.
27. Laughlin, C. A., J.-D. Tratschin, H. C. Coon, and B. J. Carter.
1983. Cloning of infectiousadeno-associatedvirus genomes in
bacterialplasmids. Gene 23:65-73.
28. Maniatis, T., E. F. Fritsch,and J.Sambrook. 1982. Molecular cloning:alaboratorymanual. Cold Spring Harbor Laboratory,
ColdSpring Harbor, N.Y.
29. Mendelson, E., J. P. Trempe,andB.J. Carter. 1986. Identifi-cation of thetrans-acting Repproteins of adeno-associatedvirus by antibodiestoasynthetic oligopeptide. J. Virol. 60:823-832.
30. Redemann, B. E., E. Mendelson, and B. J. Carter. 1989. Adeno-associated virus Repproteinsynthesis during productive infection. J. Virol.63:873-882.
31. Samulski, R. J., L. S. Chang, and T. Shenk. 1987. A
recombi-nant plasmid from which aninfectious adeno-associated virus
genome can be excised in vitro and its use to study viral replication. J. Virol. 61:3096-3101.
32. Shade, R.O., M. C. Blundell, S. F. Cotmore, P. Tattersall, and C. R. Astell. 1986. Nucleotide sequence and genome organiza-tionof human parvovirus B19 isolated from the serum of a child during aplastic crisis.J.Virol.58:921-936.
32a.Snyder, R. O., R. J. Samulski, and N. Muzyczka. 1990. In vitro resolution ofcovalently joined AAV chromosome ends. Cell 60:105-113.
33. Srivastava, A., E. W. Lusby, and K.I.Berns. 1983. Nucleotide sequenceandorganization of adeno-associated virus2genome. J.Virol. 45:555-564.
34. Straus, S. E., E. D. Sebring, and J. A. Rose. 1976. Concatemers of alternating plus and minus strands are intermediates in adenovirus-associated virusDNAsynthesis. Proc. Natl.Acad. Sci. USA 73:742-746.
35. Tratschin,J.-D., I. L. Miller, and B. J. Carter. 1984. Genetic analysis of adeno-associated virus: properties of deletion mu-tantsconstructedin vitroand evidence foranadeno-associated virusreplication function. J. Virol. 51:611-619.
36. Tratschin, J. D., J. Tal, and B. J. Carter. 1986. Negative and positive regulation in trans of gene expression from adeno-associatedvirus vectorsinmammaliancellsbyaviralrep gene product. Mol. Cell. Biol.6:2884-2894.
37. Trempe, J. P., and B. J. Carter. 1988. Regulation of adeno-associated virusgeneexpression in293cells:control of mRNA abundance and translation.J. Virol. 62:68-74.
38. Trempe, J. P., E.Mendelson, and B. J. Carter. 1987. Charac-terization ofadeno-associated virusrepproteinsin humancells byantibodies raised against repexpressed inEscherichia coli. Virology 161:18-28.