Copyright© 1976 American Society for Microbiology Printed in U.S.A.
Electron
Microscope
Studies of
Temperature-Sensitive
Mutants of Herpes Simplex Virus Type 2
GUY A. CABRAL* AND PRISCILLA A. SCHAFFERDepartmentofVirology andEpidemiology,BaylorCollege of Medicine, Houston, Texas 77030
Receivedfor publication 15 December 1975
Ninetemperature-sensitivemutants of herpes simplex virus type 2 represent-ing eightcomplementationgroupswere assigned to two classes as aconsequence of the virion forms and virus-specific cellular alterations observed in thin sections ofmutant-infected humanembryonic lung cells grown at the nonper-missive temperature. Mutants in class A, one DNA- and one DNA+, failed to
synthesizedetectablevirus particles. Mutants in classB,4DNA-and 3 DNA', produced moderate to large numbers of empty nucleocapsids. Dense-cored
nu-cleocapsids were not observed in thin sections of cells infected with any of the nine mutants at thistemperature. Virus-specific cellular alterations consisted
primarily ofmargination ofchromatin and nuclear membrane thickening and
duplication.
Theapplication oftemperature-sensitive (ts) mutants with defects in various stages of repli-cation as tools for the elucidation of the se-quence of events in viral assembly has been well established. Previous reports from this
laboratory have described the isolation of ts mutants of herpes simplex virus type 1 (HSV-1) and the classification of 22 of these mutants into 15complementationgroups (12). An
ultra-structural description of defectsof virion mor-phology, assembly, and maturation exhibited
bymutants representing each of these comple-mentation groups in thin sections at the non-permissive temperature has been published (13). Thisstudydemonstrated thatthemutants
varied in their ability to synthesize viral
nu-cleocapsids, in the kinds of nucleocapsids
syn-thesized, andintheirabilitytoproducemature
enveloped virions.
Thus,
thesemutants areap-parently blocked in various stagesofvirus
as-sembly and maturation at the nonpermissive
temperature.
Further investigations have dealt with the isolation, complementation, and phenotypic characterization of ts mutants ofherpes
sim-plex virus type 2 (HSV-2), in which nine ts mutants wereclassified intoeight
complemen-tation groups (1, 4).
Inthepresent report we describe the results of thin-section analysis of HSV-2 ts
mutant-infected cells maintained at thenonpermissive temperature. This study demonstrates that mutants representing eight complementation
groups differ markedly from each other, from the wild-type virus, and from ts mutants of HSV-1 with regardto the products of morpho-genesisatthistemperature.
MATERIALS AND METHODS
Cell cultures and media. Humanembryonic lung
fibroblasts,strain670,inpassage 9,weregrownin
8-ounce (ca. 0.24-liter) prescription bottlesor60-mm
petridishesusing Eaglemediumsupplemented with
10%fetal bovineserumand0.075 or 0.225%NaHCO3
for stoppered vessels or plates, respectively. They
were maintained in the same mediumcontaining5%
fetal bovineserumand0.150 or 0.225%NaHCO3for
bottlesorplates.
Virus and virus assays. HSV-2, strain 186 (11),
and nine HSV-2 ts mutants derived from strain186
were usedin this investigation. The mutants,
as-signedto eight complementation groups, were
iso-lated and characterized as describedpreviously at
permissive(pT°)andnonpermissive(npT°)
tempera-tures of 34 and 38C,respectively (1, 4). Preparation
of virus stocks(2, 13)andinfectivity assays (3)were conducted in accordance withprevious reports.
Di-lution of virus suspensionsandwashing of
monolay-ers wereperformed using Trisphosphate buffer at
pH7.4containing 1%fetal bovine serum. Cultures
inbottles were incubatedinwaterbaths (Blue M.
Electric Co., Blue Island, Ill.) having temperature
variations of ±0.1 C, whereas petri disheswere
in-cubated inhumidified CO2 (5%) incubators (Wedco
Inc., Silver Spring, Md.) with temperature
varia-tionsof ±0.2 C.
Infection of monolayers. Subconfluent
monolay-ers in 8-ounce prescription bottles containing
ap-proximately3 x106 cells were inoculated with1mlof
cell-freewild-type (WT)or tsmutant-virus
contain-ing 5 PFU/cell. Replicate cultures were incubated
for1hat37Ctoallowfor virusadsorption.Cultures
were then washedtwiceand incubatedwith 10 ml of
freshmaintenancemediumat 34and38C inwater
baths. At the indicated times, cells were scraped
intothe medium. Forinfectivity assaya1.5-ml
sam-ple of infected cell suspension wasfrozen, thawed,
andthensonicated for1 minat10kcycles/switha
727
on November 10, 2019 by guest
http://jvi.asm.org/
728 CABRAL AND SCHAFFER
Raytheon sonic oscillator. After clarification of the
suspension by low-speed centrifugation at4C, the
supernatant fluidwasassayedat34 Ctodetermine
the total yield of infectious virus and at 38C to
determine theyield ofts+ revertantvirusas
previ-ouslydescribed (13). The remaining cell suspension
wasprocessed for electron microscopy as described
below.
Electron microscopy. Cellsuspensionswere
cen-trifuged at 1,500 rpm at4 C, resuspended in cold
(4 C) Tris,andcentrifuged again. The resultant
pel-letswerefixedincold2.5%glutaraldehyde buffered
with Sorensen solution, pH 7.2. Pellets were
washed, postfixed, dehydrated, and embedded as
previously described (5). Thin sections were
exam-inedinaHitachi HU-11B electron microscope
oper-atingatanaccelerating voltage of75kV.
RESULTS
Growth of WT virus at pT' and npT'. The replicative cycle of the WT virus in human
embryonic lung cellsgrown atthepT' andnpT'
wasexaminedbeforeultrastructural studies of
ts mutants were initiated. For this purpose, WT virus-infected and control, mock-infected
cultures wereharvested,processed, assayed for
infectious virus, and examinedby thin-section
analysis atintervals from0through50 h post-infection (p.i.). Conditions ofone-stepinfection
were achieved when cultures were inoculated
with 5 PFU/cell (Fig. 1). Progeny virus was
8 * 7 0 E L-6 5 4 3
10 20 30 40 50
HOURS POST-INFECTION
FIG. 1. Growthcurveof HSV-2 (strain 186) WT
virus in monolayer cultures of human embryonic
lungfibroblasts. Cells wereinfectedatamultiplicity
of5PFUlcell, and virus infectivity was assayedas
described in Materials and Methods. Symbols: 0,
infectivityat thepermissive temperature (34C); 0,
infectivity atthe nonpermissivetemperature (38C).
J. VIROL.
firstevidentatapproximately 6 p.i. and
contin-uedtobesynthesized through24 h at38 C and
through 50 h at 34C. Titers of virus at 24 h
were6 x 107PFU/mlat38 C and 8 x 107PFU/ mlat34C. Thegrowthcurvesof WT HSV-2at
thetwo temperatures differedin two respects.
(i) Virus was produced more rapidly at 38 C
between 7 and 13 h p.i. as compared to 34 C; and (ii) at 38C production of infectious virus decreased after 24 h p.i.-presumably due to
inactivation at thistemperature-whereas
vi-rus production at 34C continued to increase slightly.
Thin-section analysis of cells infected with
WTvirus and maintainedat34 C demonstrated thepresenceof nakedand enveloped viral
par-ticles from 7 through 30 h p.i. (Table 1). The
relative number of virus particlesper nucleus was maximum from 12 through 16 h p.i.,
al-though large numbers continuedtobeobserved after thatperiod. Sections of WT virus-infected cells incubatedat38 C,ontheother hand,
con-tained naked viral particles in 25% of nuclear profiles examined asearlyas 5 hp.i., whereas
envelopedparticleswerenotobserved until 3h
later. The maximum relative number of virus particles pernucleus at 38 C preceded thatat
34 Cby2 to 4h. Large numbers of intranuclear
particleswereproduced, nevertheless, through
20hp.i. Prominent clearing of nuclei and
occa-sional aberrantnucleocapsids wereobservedin
sections of cells cultured after 20 hp.i.;
conse-quently, accuratequantification of virus parti-cles was not possible at either temperature
after this time.
On the basis of these data, an incubation
period of 18hwasselected forexamination ofts
mutants, sinceapproximately equal infectivity
titerswereproducedatboth 34 and 38 Catthis
timeand because thin-section analysis demon-strated that all cells infected with the WT virus and incubated at 34 or 38 C contained both
naked and enveloped viral particles in large andnearly equal numbers (Table 1).
Furthermore, growthcurvesof theWT virus
andthe mutants at34 Chave beenshowntobe
similarin boththe timeof first appearance of
infectious virus andinthe timeatwhich maxi-mum yields were reached (unpublished data).
No adjustments in the time of harvest were, therefore, made foranyofthe mutants.
Yields ofinfectious WT andts mutantvirus after18 h atpT° andnpT°. Yields of infectious virusfrom human embryonic lung cell cultures infected withthe WT virus and with the ninets
mutants at the pT° and npT° after incubation
for 18hareshowninTable2. At34 C yieldsof
fourof fiveDNA-mutants andoneDNA+
mu-tant(tsF3) approximated those of the WTvirus,
whereas three of four DNA+mutantsproduced
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.63.253.371.600.2]TABLE 1. Quantification of virus particles at different time intervals in thin sections ofcells infected with the
WTvirus at 34 and 38 C
%Cells with virus % Cellswith naked %Cellswith enve- Relativeno. of virus Time p.i. particles viralparticles lopedviral particles particlespernucleus'
(h)
34C 38C 34C 38C 34C 38C 34C 38C
4 0 0 0 0 0 0 0 0
5 0 25 0 25 0 0 0 1+
6 0 50 0 50 0 0 0 1+
7 100 100 100 100 33 0 2+ 2+
8 100 100 100 100 50 40 3+ 4+
10 100 100 100 100 100 100 3+ 4+
12 100 100 100 100 100 100 4+ 3+
14 100 100 100 100 100 100 4+ 3+
16 100 100 100 100 100 100 4+ 3+
20 100 100 100 100 100 100 3+ 3+
24 100 100 100 100 100 100 3+ ND'
30 100 100 100 100 100 100 3+ ND
aPercentage of positive cells observedin 50profiles examined.
b Numbersof particlespernucleusweregradedasfollows: 4+, >50;
[image:3.503.51.242.310.443.2]" ND, Notdone.
TABLE 2. Yields of infectious virus from cultures
infected with the WT virus and ts mutants at 34 and
38C
Viral Infectious virus
Mutant Virus DNA (PFU/ml)
class
pheno-type"
(38C) 34C 38CA WT + 6 x 107 1x 108
tsC2 - 4 x 107 2 x 103
tsD6 + 2 x 107 2 x 103
B tsAl - 5 x 107 6 x 103
tsA8 - 5 x 107 6 x 103
tsB5 - 8 x 107 1 X 104
tsH9 - 2 x 107 3 x 103
tsE7 + 1x 107 7 x 103
tsF3 + 4 x 107 1 X 104
tsG4 + 1x 107 2 x 104'
aViralDNA phenotypes from Esparza et al. (4).
b Mutantswere grown at 34 and 38 C; assays were
performed at34C.
c 103PFUof ts+ revertant virus per ml was
pres-entintheyield of tsG4at 38 C.
slightly less infectious virus. At the
higher
temperature, however, all mutants produced significantly less virus, with titers
approxi-mately 10-4 that of the WT virus. Leaky (ts) virus was produced by all mutants, and one
mutant (tsG4) yielded 103 PFU of revertant
(ts+) virus per ml.The absence ofts+revertant
virus in the progeny ofeight ofnine mutants
and the low revertant
yield
found with tsG4substantiate that ultrastructural observations
were indicative ofreplicative events ofts
mu-tantvirusandnot ofts+revertantvirus.
Electron microscope observations.
Quanti-fication of virusparticles and evaluation of vi-rus forms and specific cellular alterations in
thin sectionswere
performed
after examinationof75 to 100cell
profiles
of eachsample.
3+, 20-50; 2+, 10-20; 1+, 1-9.
TABLE 3. Quantification of virus particles in thin
sectionsof cells infected for 18 h with theWTvirus
andts mutants at 38C
Viral %Cells %Cells Relative DNA with na- withen- no.of
vi-Mutant D
-
velopedclass
Virus
pheno- kedvirus virus rus par-type parti- part ticles per (38C) dlesa parti- nucleusA WT + 100 100 4+
tsC2 - 0 0 0
tsD6 + 0 0 0
B tsAl - 80 0 2+
tsA8 - 75 2c 2+
tsB5 - 60 0 2+
tsH9 - 30 0 1+
tsE7 + 90 0 3+
tsF3 + 90 3c 3+
tsG4 + 80 0 2+
aPercentage of positive cells observedin75 to 100
profiles examined.
bNumbers of particles per nucleusweregradedas
follows: 4+, >100; 3+, 20-50; 2+, 10-20; 1+, 1-9.
c No enveloped particles were seen at the cell
surface of these cells. Enveloped forms were
ob-served inthe cytoplasm and at the nuclear
mem-brane.
Cellsinfectedfor18hwith the WT virusand incubated at 34 and 38 C and cellsinfected with
ts mutants at 34Cdemonstrated little
qualita-tive difference.
(i) WT virus. The relative number ofvirus
particles pernucleusinWTvirus-infectedcells
grown at 18 h at the npT° was significantly
higher than that oftsmutant-infected cells
(Ta-ble 3). Characteristic features of WT
virus-in-fected cellsinthin sections includedthickening
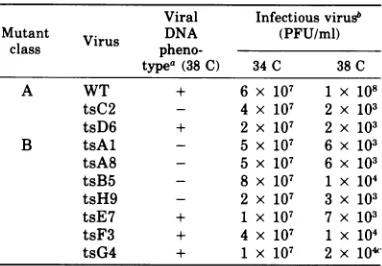
andduplicationof thenuclear membrane (Fig.
2Aand B) and condensation andmarginationof
chromatin (Fig. 2AandC). (Abetter represen-tationofthe pattern ofmarginated chromatin
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.254.448.311.450.2]730 CABRAL AND SCHAFFER
observed in WT virus-infected cells is demon-strated in
Fig.
4A,
which illustrates atsmu-tant-infected cell.) Intranuclear fibrillar bun-dles (Fig. 2D) werealsoobserved in WT virus-infectedcells (Table4)butnotinmock-infected cells. Fine-grained nuclear inclusionswerealso present, distributed either as
single
electron-.
~~~~~~~~~~~~~
dense roundmassesor as
irregular
anastomos-ing
arrays(Fig.
3A). All nuclearprofiles
con-tained corelessor
empty
nucleocapsids, capsids
surrounding
a 45- to50-nm,
ring-shaped
core(partial capsids),
andcapsids
containing
densecores
(i.e.,
presumably
containing
viral DNA)(Fig.
3B).
Occasionally,
particles
without densei%. 7
-.
I -n
WAL-6-I'I'
77
4
;R
n
.
44Al!
blat'
,f#si'
e
eFIG. 2. Humanembryoniclungcellsinfected with the WT virusafter18hat38C.(A)Nucleus(nu)and
cytoplasm (cy) of infected cell demonstrating margination ofchromatin (mc) and thickening ofnuclear
membrane (closed arrow). Note the presence of partial nucleocapsids adjacent to and buddingfrom the
nuclearmembrane(open arrow). An envelopednucleocapsidwithadensecore is seen inthecytoplasm.(B) Duplication of the nuclear membrane. (C) Nucleus (nu) and cytoplasm(cy) of infectedcellshowing
partially
enveloped dense-coredcapsidand margination of chromatin(mc).(D) Nucleus(nu) of infected cell
showing
fibrillarbundle(arrow). x59,800.
J. VIROL.
I.!r.
A.,,k
,., -, 19
01-.;%% I4,46.*
44-ICy
,A,
.,tI,.
.11
-SAIL .zI?
A .
I..
L'.. .; .4.
A&..PA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.95.425.155.586.2]TS MUTANTS OF HSV-2 731
TABLE 4. Virion forms and virus-specific alterations in cells infected with WT virus and ts mutants at 38Ca
CYTOPLASM AND/OR
NUCLEUS EXTRACELLULARSPACE
lIl
MUTANT VIRUS VIRAL FIBRILLAR RING-LIKE MEMBRANE MEMBRANE ''PER/CHROOATIN- NUCLEOCAPSIDS ENVELOPEDNUCLEOCAPSIDS CLASS DNA BUNDLES COMPONENTS THICKENING DUPLICATION LIKE''GRANULES
38- , EMPTYPARTIAL DENSE EMPTY/PARTIAL DENSE
WT + + + + + ! +
A tsC2 +
tsD6 + --*
B tsAl + + + +
tsAR - - + +
-tsBS - + + _
ts H9 - + +
Is E7 F _+_ _
tsF3 + - - + ++
Is G4 + - -+
a Evaluation based on the frequency of virion forms and the degree of alteration observed in virus-positive
cellsonly: -, absent; ±, presentinsmall amounts or numbers; +, present in moderate amounts or numbers;
++, present in large amounts or numbers.
cores were observed budding from the inner
nuclear membrane (Fig. 2A). Ring-like
intra-nuclear components 25 to 27 nm in
diameter,
similartothose describedpreviously forHSV-1
(13), were also observed. Cytoplasmic
altera-tions included vacuolization and formation of
concentric arrays of membranous cisternae.
Numerous enveloped dense-cored particles
were seen in most WT-infected cells (Fig. 3C).
Inaddition, moderate numbers ofvirions were
localizedatthe cellsurface andinthe extracel-lularspace (Fig. 3D).
(ii)Tsmutants.The ninemutantsexamined
were subdivided into two classes as a conse-quence of (i) the cellular alterations
they
in-duced and (ii) their ability to
synthesize
viral particles at38 C(Tables
3and4). ClassAmu-tants included one DNA- mutant, tsC2, and one DNA+ mutant, tsD6. These mutants were maximally defective, at leastat 18 hours p.i.,
since cells infected with them and maintained
at38C exhibited fewvirus-induced alterations
(Table4; Fig. 4A). Cell profiles of class A mu-tant-infected cultures contained no detectable viral particles. Although nuclear membrane thickeningwaspresentto amoderate
degree
in tsC2-infected cells (Fig. 4A) and, to a lesser extent, in tsD6-infected cells, nuclearmem-brane
duplication
and intranuclear fibrillar bundles were not observed. Margination of chromatin was observed in nuclei of cells in-fected with bothmutants.The remaining seven mutants, four
DNA-(tsAl, tsA8, tsB5, andtsH9) and three DNA+ (tsE7, tsF3 andtsG4),wereassignedtoclassB.
The incidence of class Bmutant-infected cells containing virus particles variedfrom 30% for
tsH9to 90%for tsE7and tsF3 (Table3).
Gener-ally speaking, the greaterthe number ofcells containing virusparticles, thegreaterthe
num-ber of particles observed per nucleus. Cells in
which90% of nuclear profiles contained naked viral particles (i.e., tsE7- and tsF3-infected cells) contained 20 to 50 particles per nucleus, and cells in which 60 and 75% of nuclear
pro-filescontained nakedviral particles (i.e., tsB5-andtsA8-infected cells,respectively)contained 10 to 20 particles per nucleus. In comparison, cells infected withtsH9contained naked parti-cles in only 30% of nuclei and each nucleus
contained fewer than 10particles (Table3).
Nuclear membrane modificationswere prom-inent features of cells infected with different
mutants of class B. Thickening of the nuclear membraneoccurred incells infected with three
DNA- mutants (i.e.,tsAl, tsA8,andtsB5)
(Ta-ble4) and with one DNA+ mutant, tsF3(Table
4). Whereas tsD6 (class A)-, tsAl-, tsA8-, and tsB5-infected cells exhibited only short seg-ments of membrane thickening, in cells
in-fected withtsF3thethickeningexceeded that of any of the other mutant-infected cells and
re-sembledchangesobservedin WTvirus-infected cells(Fig. 4B).Membraneduplication,a
promi-nent feature of cells infected with WT virus,
was limited to small segments of the nuclear membraneincellsinfected with tsAl and tsF3. Like mutantsinclass A, intranuclear fibrillar bundles were not observed in any of the cells infected with classBmutants.
on November 10, 2019 by guest
http://jvi.asm.org/
732 CABRAL AND SCHAFFER
.Y#.
7:
i4p
_L*
oHe0
_I1
I ,
D
4 $4
4
I'>''.t$4i''" X s ' A.~
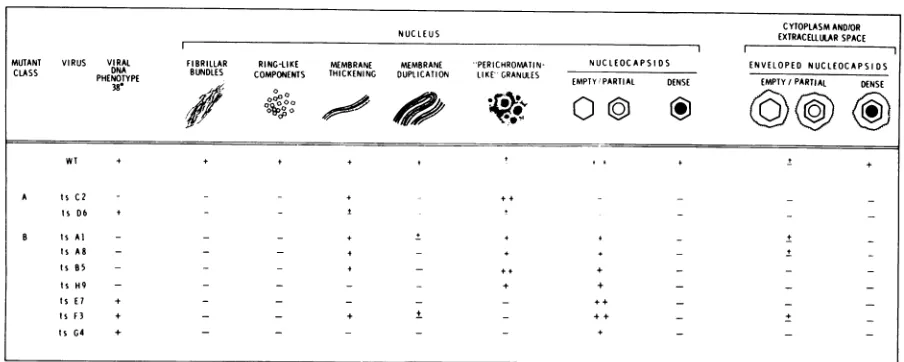
FIG. 3. Humanembryonic lung cells infected with WT virus after 18 h at 38 C. (A) Irregular anastomising
inclusion body (ar) found within the nucleus (nu). Note the presence of marginated chromatin (mc). (B)
Nucleocapsids with dense cores (long arrow), partial cores (short arrow), andempty capsids (open arrow) in
the nucleus. (C) Enveloped nucleocapsids with dense cores present in the cytoplasm. (D) Enveloped
nucleocap-sids withdense cores are seen at the cell surface and in the extracellular space.(A-C) x59,800; (D) x27,000.
Nuclei of cells infected withfive of the classB
mutants contained small to moderate numbers of empty and partial nucleocapsids (Table 4). The nuclei of tsF3 and tsE7 mutant-infected
cells, however, included large numbers of pre-dominantly partial nucleocapsids (Fig. 4C). In no case were nucleocapsids containing dense
coresobserved. In instances where capsids
con-training partial cores were adjacent to the
nu-clear membrane (e.g., tsA8) thickening ofthe
membrane and budding of particles were
occa-sionally observed (Fig. 5A). Cytoplasmic alter-ations, in comparision to WT-infected cells, were minor. In three cases (tsAl-, tsA8-, and tsF3-infected cultures) enveloped particles
con-taining empty nucleocapsids were located in
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.103.429.66.508.2]TS MUTANTS OF HSV-2 733
B
,i. 1.
.i
1.~ ~ ~ ~ t
X.+e
#
Y,
*9..
'4
e
a
t
S
'4tsI. 'I
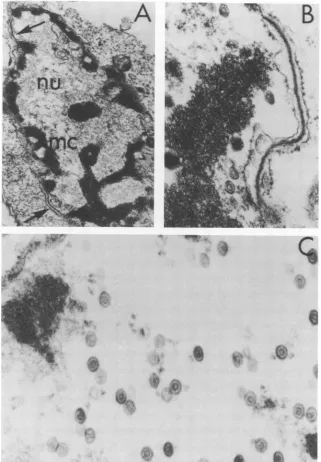
FIG. 4. (A) Human embryonic lung (HEL) cell infected with class A mutant tsC2 after 18 h at 38 C. Note
thepresenceofmarginated chromatin (mc) and thickened segments of nuclear membrane (arrows) and the
absenceof viralnucleocapsidsinthenucleus. x10,000.(B)HELcellinfectedwith class B mutant tsF3 after
18 h at38C showingextensive thickening of the nuclear membrane. x59,800. (C)HEL cell infected with
class BmutantstsF3after18hat38 C.Notenucleus containing relatively large amounts of nucleocapsids
containing partialcores. x59,800.
the cytoplasm of cellsexhibitingintactnuclear membranes (Fig. 5B and C).
A prominent feature of cells infected with DNA- mutantsinbothclass A and class B was the presence ofround, 50- to 60-nm
"perichro-matin-like" intranuclear granules (Table 4; Fig. 6).These were especially numerousincells infectedwithtsB5,aclass B mutant, andtsC2,
aclass A mutant. Nuclei of cells infectedwith
class B mutantstsAl, tsA8,and tsH9contained
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.89.410.71.533.2]734 CABRAL AND SCHAFFER
'A *~~~~~~~~~~~~~~~ti
-~~~~~-~r
'%- --' -
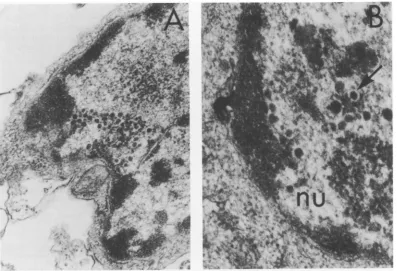
.-FIG. 5. (A) Humanembryonic lung (HEL) cell infectedwithmutanttsA8 after18 hat38 C.Nucleus (nu)
andcytoplasm (cy)ofinfected cell showapartialnucleocapsid budding from the nuclear membrane (long
arrow), whereasanenveloped partial nucleocapsid (shortarrow) is localized within the perinuclearspace.
Twoextranuclear enveloped naked particles arealsoseen(open arrow). (B)HEL cell infected withmutant
tsA8after18 hat38Cshowing envelopednucleocapsidcontainingapartialcorelocalizedincytoplasm.(C)
HEL cell infected with mutant tsAl after18 h at38Cshowingan envelopedempty nucleocapsidin the
cytoplasm. x59,800.
moderate numbers of granules. Granules were also present in small numbersinnucleiofcells
infected with one DNA+ mutant, tsD6, and
withthe WT virus. Granules were not observed
in nuclei of cells infected with three DNA+ mutants(i.e., tsE7,tsF3, ortsG4)orinnucleiof
mock-infected cells.
DISCUSSION
The sequence of events observed in
HSV-infectedcells has been reported in detail (6-9, 16). In cells infected in this study with strain 186 of HSV-2 and maintained at bothpT0and
npT0,
thickening and duplication of nuclear membrane were accompanied by the intranu-clearappearance of empty anddense-corednu-cleocapsids. Envelopment of the dense-cored
particles, and occasionally ofempty
particles,
occurred at the nuclear membrane. The
pres-ence ofintranuclear fibrillar bundles was also observed.
Although they resembled WT virus-infected
cellsatthepT', theHSV-2 mutants examined inthe presentinvestigation exhibited all forms of nuclear virus at the npT' except that of dense-cored particles (i.e., those containing DNA). Unlike the study with ts mutants of HSV-1 (13), neitherpreviously undescribed in-tranuclear forms nor aberrant nucleocapsids
were seenin cells infected with the HSV-2
mu-tants at the npT0. (It should be pointed out, however, that thin-section analysis was
per-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.100.426.66.411.2]TS MUTANTS OF HSV-2
735
-,
[image:9.503.51.448.73.344.2],,'a
FIG. 6. Humanembryonic lung cells infected with DNA- mutant tsB5 after 18 h at 38 C. (A)Infectedcell
containing numerous"perichromatin-like" granules within the nucleus (nu; arrow). x25,600. (B)Nucleusof
infected celldemonstratingthepresence ofa clear "halo" (arrow) separating thegranulefrom thesurrounding
nucleoplasm. x76,500.
formed at a later time in the HSV-1 study.)
Furthermore, all cellular changes and virus particle forms were alsoobserved in WT
virus-infected cells. These observationssuggest that
thevirusformsobserved atthe npT'represent
actualintermediates in the replicativecycle of HSV-2 orpossibly defective particles resulting
fromaberrantassembly.
No apparent correlation was observed
be-tween theviral DNAphenotype and the
pres-ence of physical particles in HSV-2
mutant-infected cells grown at 38 C. Although large numbers of empty and partial nucleocapsids
were foundinnuclei of cells infectedwithtwo
DNA+ mutants (tsE7 and tsF3), those of cells
infected withoneDNA+mutant(tsG4) and four
DNA- mutants (tsA1, tsA8, tsB5, and tsH9)
contained moderate numbers of particles. In
addition, the twoclass A mutants, one DNA+
and one DNA-, failed to produce observable
virus particles. All of the mutants, however,
induced thesynthesis ofviral-specific antigens at the npT° as determined by
immunofluores-cence (4) and polyacrylamide gel
electropho-reticanalysis (10).
Therefore, synthesis of viral proteins and
their subsequent assembly does not appear to
dependuponviral DNA synthesis. These
find-ings are inagreementwith thoseof Nii et al. (9)
and of Schafferetal. (13), forHSV-1, who
sug-gested that the information required for the synthesis of HSV
structural
proteins is readfrom the
parental
genome when viral DNA synthesisisinhibited.Dense-cored
particles were not observed inany ofthe
mutant-infected
cells at the npT'.The absence of dense-cored nucleocapsids in
DNA+
mutant-infected
cellssuggests that DNAencapsidation
didnot occurandthat thesemu-tants areblockedinanearlystepinmaturation preceding
encapsidation
of the viralgenome. Inaddition, envelopmentof emptyand
partial
nu-celocapsids,
observedpreviously
by Smith andDeHarven (15)andSchafferetal. (13),appears
not todependonDNA
encapsidation,
since en-veloped empty core particles were localized inthe cytoplasm of cells infected with tsF3, a
DNA+ mutant, andwith
tsA1
andtsA8, DNA-mutants.Moderate to large accumulations of
"peri-chromatin-like" granules
were present in nu-clei of cells infected with each of theDNA-VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
736 CABRAL AND SCHAFFER
mutants and grown at 38 C. These inclusions
were also present in small numbers in cells infected with WT virus and with one DNA+
mutant, tsD6. The granules have been re-garded by others as precursors to viral
nu-cleoids (14). Thepresenceofaclear "halo"
sur-rounding thesedensities and their50-to60-nm diameter have led some investigators to sug-gestthat theymaybeidenticaltothe
perichro-matin granules found in uninfected cells (16). That these inclusionswereespecially abundant
inDNA-mutant-infected cells and absent from mock-infected cells at the npT° suggests that they are not viral DNA and that they may
representaccumulations of viralorcellular
pro-teins, orextranuclear RNA. Since each of the
four DNA- mutants is defective in adifferent
cistron, theabsence of viral DNA itself andnot
adefect inaspecific viralgene mayaccountfor theappearanceof thesegranular clusters. The
nature and composition of the inclusions are
currently being investigated.
Similarities anddifferences between cells
in-fectedatthenpT°with ts mutantsof HSV-2in
these studiesand withtsmutantsof HSV-1(13)
arenoteworthy. Certainmutants of both types exhibitedthickening and duplicationofnuclear membranes and thepresence ofemptyand
par-tial nucleocapsids in nuclei. The relative
num-bers ofcapsidspernucleusprofilewere higher
in HSV-1 mutant-infected cells. Dense-cored capsids, however, were observed only in cells
infected with six DNA+ mutants of HSV-1. Consequently, all the HSV-2 mutantsused in thisstudywereapparently blockedat38C ina
stage ofassembly that precedes encapsidation
ofthe viral genome. In addition, aberrant
nu-cleocapsids present in cells infected with five different mutants ofHSV-1 werenot observed
in any ofthe HSV-2 WT- or mutant-infected cells. Althoughmutantsinfour HSV-2 comple-nentation groups possess thermolabile virion
structural components, these components are
either (i)not associated withthecapsidor (ii),
ifthey do represent capsid components, their assembly isnotvisibly defective at38 C.
Nucleocapsidswerepresentinnuclei of
HSV-1 DNA- mutant-infected cells, a finding in
agreement with the observations presented in
this communication. Thepresenceof capsidsin both HSV-1 and HSV-2 DNA- mutant-infected cells suggests that DNA synthesis is not
re-quired for nucleocapsid assembly. With regard to "perichromatin-like" granules, it was not
possible to determine accurately whether they were present in nuclei of HSV-1
mutant-in-fected cells since that investigation was
con-ducted using cells infected for 48 h at the npT0; inthepresent studythese granuleswere pres-ent only at earlier times p.i. Studies are
cur-rently being conducted to determine whether these granules are indeed characteristic of HSV-1 DNA- mutant-infected cells.
ACKNOWLEDGMENTS
This work was supported by Public Health Service re-search contract NO1 CP 53526within The Virus Cancer Programof the National Cancer Institute,research fellow-ship 1 F22 AI 847from the National Institute of Allergy and Infectious Diseases, and research grant CA 10, 893 fromthe NationalCancer Institute.
Weacknowledge the excellent technical assistanceof P. Palmer and S. Moore.
LITERATURE CITED
1. Chu, C.-T., and P. A. Schaffer. 1975. Qualitative com-plementation test for temperature-sensitivemutants ofherpes simplexvirus. J. Virol. 16:1131-1136. 2. Courtney, R. ., R. M. McCombs, and M.
Benyesh-Melnick. 1970. Antigensspecified by herpesviruses. I.Effect of argininedeprivation on antigensynthesis. Virology 40:379-386.
3. Dreesman, G. R.,and M.Benyesh-Melnick. 1967. Spec-trumof human cytomegalovirus complement-fixing antigens. J.Immunol. 99:1106-1114.
4. Esparza, J., D.J. M. Purifoy, P. A. Schaffer, and M. Benyesh-Melnick. 1974. Isolation, complementation andpreliminary phenotypic characterization of tem-perature-sensitive mutants ofherpessimplex virus type 2.Virology 57:554-565.
5. McCombs, R. M., M. Benyesh-Melnick, and J. P. Brunschwig. 1968. The use of Millipore filters in ul-trastructural studies of cell cultures and viruses. J. CellBiol. 36:231-243.
6. Nii, S. 1971. Electron microscopic observations onFL cells infected with herpes simplex virus. I. Viral forms. Biken J. 14:177-190.
7. Nii, S. 1971. Electron microscopic observations on FL cells infectedwithherpes simplex virus.II. Envelop-ment.Biken J.14:325-348.
8. Nii, S., C. Morgan, and H. M. Rose. 1968. Electron microscopy of herpessimplex virus. II. Sequence of development.J. Virol.2:517-536.
9. Nii, S.,H.S. Rosenkranz, C. Morgan, and H.M. Rose. 1968. Electron microscopy of herpes simplex virus. III.Effect ofhydroxyurea. J. Virol. 2:1163-1171. 10. Powell, K. L., D. J. M. Purifoy, and R. J.Courtney.
1975.The synthesis of herpes simplex virus proteins in the absence ofvirus DNA synthesis. Biochem. Biophys. Res. Commun. 66:262-271.
11. Rawls, W. E., D. Laurel, J. L. Melnick, J. M. Glicks-man, and R. H.Kaufman. 1968. A search for viruses insmegma, premalignant and earlymalignant cervi-cal tissues. The isolation of herpesviruses with dis-tinctantigenicproperties. Am. J. Epidemiol. 87:647-655.
12. Schaffer, P. A., G. M. Aron, N. Biswal, and M.
Ben-yesh-Melnick. 1973. Temperature-sensitive mutants ofherpes simplex virustype 1: isolation, complemen-tation andpartial characterization. Virology 52:57-71.
13. Schaffer, P.A., J. P. Brunschwig, R. M. McCombs, andM.Benyesh-Melnick. 1974. Electron microscopic
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
737
studies oftemperature-sensitive mutantsofherpes
simplex virustype1.Virology62:444-457.
14. Shipkey, F. H., R. A. Erlandson, R. B. Bailey, V. I.
Babcock, and C. M. Southam. 1967. Virus biogra-phies. II. Growth of herpes simplex virus intissue culture.Exp. Mol.Pathol.6:39-67.
15. Smith, J. D., and E. De Harven. 1973.Concentration of herpesviruses.J.Virol. 11:325-328.
16. Smith,J.D., andE.De Harven. 1973. Herpessimplex virusand human cytomegalovirus replicationin WI-38 cells. I. Sequence of viral replication. J. Virol.
12:919-930.
VOL. 18, 1976