Copyright( 1971 American SocietyforMicrobiology Prinited inU.S.A.
Separation and Structure
of
Components
of
Nuclear
Polyhedrosis Virus of the Silkworm
TOSHIYUKI KHOSAKA, MICHIO HIMENO, AND KONOSHIN ONODERA
Laboratory ofBiochemistry, Department of Agricultural Chemistry, Kyoto Untiversity, Kyoto, Japan Received for publication 21 September 1970
Morphology of structural components of nuclear polyhedrosis virus (NPV) particles of the silkworm (BombyxmoriLinne) wasstudiedbyelectronmicroscope using negative staining. NPVparticles isolated from polyhedra could be separated into five structuralcomponentsby centrifugation in sucrosedensity gradients. The
lowest band (band I) was found to consist of thick rod-shaped particles (330 by
80nm) with knobby surfaces and with occasional protrusion at one end. The
second band from the bottom (band II)wasshowntoconsistmainlyof slender
rod-shaped particles (360 by 60 nm), in which internalstructureswerevisibleas adense mass. Regular striations werealso seen on the surface of theseparticles. By treat-ment with mercaptoethanol, these particles weredrastically damaged,and in some cases the internal substances werepartially released, producing empty inner
mem-branes ofvarious degrees of disintegration.In bands III andIV,both empty
spheri-cal and empty rod-shaped membranes were present. Band III was rich in empty
spherical membranes which wereshownto be theouter membranes of thick
rod-shaped particles. The empty rod-shaped membranes, the inner membranes, were
mainly located in band IV and havecrossstriationsonthe surface. It is remarkable
that the uppermost band (band V) consisted purely of small spherical particles,
somewhat heterogeneous in size and shape (around20 to 25nm in diameter),
in-dicating the particles to be the degradation product of the virus particles. Similar
particlescouldalsobeobservedwithin the empty inner membranes.
Although the structure of nuclear polyhedrosis
virus (NPV) ofinsectshas been investigated by a number of workers, and was found toconsist of
complex rod-shaped particles, the fine structure of theparticles still remains obscure. Earlier
ob-servationsontheshadowedpreparations ofvirus particles isolated from polyhedra by treatment with weakalkali(1,2,3) haveledtotheworking modelthat severalspherical subunits with a
diam-eterof about40 nm wereenveloped witha setof
membranes (outer and inner membranes) to
form rod-shaped particles (4). On the contrary, Krieg (8) observed that the rod-shaped particles haveahelicalstructuresimilartothatof tobacco mosaic virus and concluded thatthesubunits do
not represent viral developmental stages but
artifactsproducedby alkaline degradationof rod-shaped particles. Such structural components
were also found innegatively stained virus prep-arations but were interpreted as discs or groups ofdiscs of the innermembranes (5).
Examination ofultrathin sections ofpolyhedra
(3, 9,10), however, gave no evidence for support-ing the existence of a subunit structure in rod-shaped particles. More recently, Kozlov and
Alexeenko (7) observed the central helical core in virusrods treated withchloroformorureaand
suggested that the structure of the inner core is
similartothreadlikebacteriophage
fd.
In aprevious report (6), it wasshownby thin sectioning of tissue infected with NPV of the silk-wormthat,attheearlier stages of the maturation process, theflexuous rod particleslooselypacked with inner membrane first appeared in the cell
nucleus and have laterallystriatedinternal
struc-tures. These particles were, then, envelopedwith
the outer membrane to form a compact rigid
structurebeforeembeddinginpolyhedralprotein.
Inthe presentstudy, the finestructure of virus
particles isolatedfrompolyhedrawasinvestigated by separating viral structural components by means ofsucrose density gradient centrifugation and byexamination of each component by
elec-tronmicroscopy.
MATERIALS AND METHODS
Preparation of polyhedra. The sources for
prepa-ration of polyhedra were the hemolymph, collected
shortly
before
death, of experimentally diseased fifthinstar larvae of the silkworm. The polyhedra were
267
on November 11, 2019 by guest
http://jvi.asm.org/
collected from the hemolymph by settling out for a
few days in the cold. Thesediment containing
poly-hedra was placed onandcentrifuged through a 50%c;
sucroselayerat8,000 X gfor15 min at 2 C. As a
re-sult, the polyhedra were recovered as a pellet at the
bottom of the centrifuge tube, whereas the majority
of the impurities remained over the sucrose layer.
After washing several times by cycles ofsuspending
in distilled water and centrifugation (3,000 X g,
15 min, 2 C), the polyhedra were further treated
with massive doses of lipase and trypsin for 2 hr at
room temperature to remove any residual impurities.
Finally, thepolyhedra wereagainwashedextensively
with distilled waterand stored as dry powder in the
cold.
Isolation of virus. Virus particles were isolated
from polyhedra by treatment with Na2CO3 solution
of various concentrations (from 0.005M to 0.05M)
containing 0.05M NaCl for 1 to 3 hr at 5 C. After
removal of impurities by low-speed centrifugation,
centrifuge. Thepelleted virus particlesweresuspended
thesuspension wascentrifugedforI hrat 105,000 X g
at 5 C in the RP 30 rotor ofaHitachi 55P-2
ultra-in -0.067 M phosphate buffer (PB), pH 7.8, and
purified by another cycle of low- and high-speed
centrifugation.
Sucrose density gradient centrifugation. The
puri-fiedviruswassuspended inasmallamountof -0.067
M PB (pH 7.8), and Tween 80 was added to give a
final concentration of 0.05% The suspension was
layered onto 30 to 50%X, gradient of sucrose in the
same buffer, andcentrifuged for 2 hr at 25,000rev/
min, 5C, in aswinging bucket rotor. After the run,
fractions were collected through a needle by
punc-turing the bottom of the tube, and measured as
absorbance at260nm.
Electron microscopy. The structural components
thusseparated were removed from sucrose either by
high-speed centrifugation or by filtration through a
shortcolumn (I by 20 cm) ofSeplhadex G-25,
equi-librated with -0.067M PB, pH 7.2. The pellets
ob-tained in the former case were resuspended in a
small volume of the same buffer. Each drop ofthe
samples was mixed ondental wax with one drop of
2%' phosphotungstic acid adjusted to pH 6.8, and
the specimens were examined immediately in a
JEM 7Aelectron microscope.
RESULTS
Asdescribed byBergold (4), NPV of insects is
easily purified, practicallyfree of host materials,
because polyhedra can readily be obtained in a
very pureform forthefollowingreasons.(i) They
are highly characteristic macromolecules several
microns indiameter withhighdensity (2), and (ii)
although proteinaceousin nature, theyare
resist-ant totheusualproteolytic enzymes. In addition
polyhedra consist only of virus particles besides
polyhedralproteinmatrix(3, 10).
Examination ofisolated virus particles in the
electron microscope revealed the presence of
several structural components. To separate these
E 0
cs 0.1 5 Band
II
Band VuX0.10 m \ BandIVI
c / \ Band I>/
1i
0.05
0. 0
10 20 30 40
Tube Number
FIG. 1. SUcrose delnsity gradienit profile of} NPV
stru(ctlurlal componzenits of B. mori. Virutsparticles were
isolated from polyhedra by treatment with 0.01 f
Na2C&O soluitionl conztaininiig 0.05m NaCl for I h1rat
SC. A 0.3-ml anmounlitofvirues suspenisionlin 0.067,ii
PB (pH 7.8) vith 0.05(%, Tween 80 waslayeredo0ito
4.4 mloJ30 to
50%(i,
sucrosegradientanidcentriJifigedat25,000 rev,lmiiifor 2 hr- at 5C in a Hitachi SW 40
Rotor.Fractioniswere collectedasdescribedin the text.
SedimnentationiisfromrighttoleJt.
components, sucrosedensity gradient
centrifuga-tion wasfoundtobeuseful. Undertheconditions
used, virus particles, isolated by dissolving
poly-hedra with 0.01 M Na2CO3 solution containing
0.05 MNaCIfor1 hr,could beseparatedintofive
bandsin 30 to 50% sucrosedensity gradients by
centrifugation in the SW 40 rotor at25,000 rev,
minfor2hr at 5C(Fig. 1).
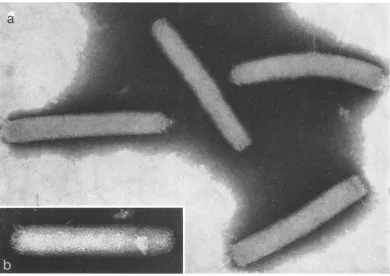
ItisshowninFig.2that thelowest band(band
I) is foundtoconsist ofthick rodparticles,
meas-uring330by 80nminaveragesize, which havean irregular knobby surface structure and occasion-ally a terminal protrusion at one end. When
en-larged, however, further details could notbe ob-tained, since the staining merely outlined the particles.
BandII (Fig. 3a) consistsmainly of slender rod particles,360 nmlong and60 nmwide.Asseenin
Fig. 3b, this particle appears to have transverse
regular bandingonthe surface, in whicha dense
mass ofinternal structure can also be seen. Of
theseparticles,some wereobservedtobedistorted
inshapeand some werepartially degraded.When
treated with 1 (j' mercaptoethanol, the slender
rodsweredrasticallydamaged,resultinginpartial release oftheinternalsubstance in someparticles
and producing empty rod membranes with
vari-ousdegrees of disintegration(Fig. 4).
BandsIIIand IV werefairly close together so
that the two bands could not be easily distin-guished. For furtherseparation, thesetwobands
were combined and recentrifuged in 20 to 40%c
sucrosegradientat25,000rev/minfor2 hr at 5C.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:2.498.266.456.78.212.2]MORPHOLOGY OF NUCLEAR POLYHEDROSIS VIRUS
FIG. 2. Virusparticles, negativelystainedwith 2%cphosphotunigstic acid,from thelowestband(ban1d1) in the'
gradient.Kniobby surfaced rod-shaped particles, 330by 80tim inaverage size, areseen. A termitalprotrusion is
observedat oneendofsomeparticles. X 50,000.
[image:3.498.117.374.79.284.2]a
FIG. 3. (a) Virusparticlesfoundinband 11. Theseslenderrod-shaped particles,about 360 nim longand 60 tinm
wide, arepartially penetrated by the stain andare somewhat elongated longitudinally anddistorted in shape.
X160,000. (b) Slender rod-shapedparticles. On thesurface, regularbanding is visible. Note that an internfal
substatice canalsobeseen inside the particle. X 160,000.
269
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.498.55.445.328.604.2]Afterthe run,however, only single a diffuse band
appeared in the gradient. Whenfractionated and
measured by opticaldensityat280nm, twowidely diffused, overlapping bands could be resolved, indicating a considerable size variation of these components. Examination of these bands in the electron microscope revealed that both empty sphericalandemptyrod-shaped membranes were present.
Band III is rich in empty spherical and frag-mented membranes (Fig. 5). Even at higher magnification, further details could not be
re-solved in these membranes. During examination
of band I, such membranes could frequently be
seenfree from the thick rodparticles to give the
appearance of slender rod-shaped particles (Fig. 6). Theemptysphericalmembranes are thus
[image:4.498.78.450.177.409.2]con-sidered to be the outer membrane of the thick
FIG. 4. Slenider rod-shaped particles treatedWith 1%O
mercaptoethaniol.
Drasticdegradationz
isapparenit.
Olne
particleis releasin7gitsintternialsuibstance, anidaiiotherontehas partiallyreleased. X 200,000.
FIG. 5.
Menmbranleolis
materials foundinban7d Ill. Emptysphericalaldfragme,itary
membranies
areobservel.X 50,000.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.498.129.401.447.628.2]rods. Similar findings have been reported in the
isolated virus preparations from some species of NPV polyhedra (2, 7). The empty rod-shaped membranes were mainly located in band IV. On
the surface, regular striations are clearly visible (Fig. 7) as can be seen with the slender rod-shaped particles (Fig. 3b). It is noted that the same ob-servation has been made by Kozlov and Alexeenko(7) with NPV of the silkworm. It seems obvious that these membranes are the inner mem-braneof the thick rod-shaped particles.
The uppermostband (band V) appeared atthe
sucrose-suspensioninterfaceas asharp band and was found to consist purely of small spherical particles with a diameter of about 20 to 25 nm
(Fig. 8). It must be noted that the particles are somewhat heterogeneous in size and shape.When
analyzed in a10to 40c% sucrose density gradient bycentrifugation for3 hrat25,000rev min, these particles behaved as a broad diffuse band, indi-cating size heterogeneity. It is also noteworthy that withinthe emptyinner membranes found in
band IV, similarparticlesarefrequentlyobserved
(Fig.
9).
DISCUSSION
NPV of the silkworm was shown to be
de-graded in a progressive fashion from thick rod
particlestosubviralstructuralcomponents by the
action ofalkali.
The thick rod particles observed in the lowest
band (Fig. 2) are apparently identical to those
found inultrathin section ofpolyhedra (3, 9, 10)
andwereelucidatedto consist of an internal
sub-stance enveloped with two layers of
membrane,
outer and inner membranes (Fig. 3a, 3b, 5, and
7). The outer membrane may serve to retain the
compact rigid structure of virus rods, since the
slenderrods, which had lostthe outer
membrane,
wereelongated longitudinallyand frequently were
distorted in shape during the process ofelectron
microscopy (Fig. 3a). This is supported by the
results obtained by thin sectioning of the tissue infected with NPV of the silkworm (6).
It is evident that when slender rods degrade,
furtherbreakages may occur preferentially at the end of the particles to release internal substances (Fig. 4) because the structure of the empty inner
~~~~~~~~~~~,.
lip/A
FIG. 6. Outermembranies arefreeing from thick rod-shaped particles as menitedmembranles. Thissamplewastakeni frombalndI. X 50,000.
empty sphericalmembrcanies or
fraig-FIG. 7. Emptyrod,iinnier memnbranesfrombauid IV.Reglalarstriationsareclearly seeuionithe suirfcace. X 150,000.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.498.103.395.331.529.2] [image:5.498.89.414.563.631.2]membrane was well preserved exceptatthe ends
ofthe membrane (Fig. 7). Although the internal
substance which appearedas a dense mass (Fig. 4) mayrepresent ahighly organized structureand
is possibly helical in configuration, as suggested
previously (5, 7),no direct evidence couldbe ob-tained.
The facts that the small spherical particles
which seemed heterogeneous in size and shape (Fig. 8) were consistently found and shown to
::k.
:
F'"4
I
a,
P..
i.
FIG. 8. Smallsphericalparticlesfrom theuppermost band(band V). It is remarkable that these particlesar.
[image:6.498.63.454.137.400.2]somewhatheterogeneouis insizeandshape. X 186,000.
FIG. 9. Membraneouis materialsfoundinband IV. Itmust be notedthat within the emptyinnermembranes
particles similar to thoseshownIinFig. 8 are observed. X 192,000.
n
.4,,.i '.
v...t,f:.'
11
AM
Av.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.498.66.453.441.609.2]behaveas abroadlydiffused band insucrose
gra-dients indicatethat the component doesnot
rep-resent any structural unit but is a degradation
product of virus particles. Similar particles are
frequently observedwithin theemptyinner
mem-braneafter variousdegreesofdisintegration. This
strongly suggests that the component (Fig. 8) might be thedegradation product of the internal
substance.
The small spherical viral subunits observed earlier in the shadowed preparation of virus particles (1,8) maybeinterpretedas
correspond-ingto those shown in Fig. 8. It is probablethat
metalshadowingcaused misunderstanding ofthe
componentas aviral subunit.
Theinfectivity ofeach fractionwas examined,
and it has been found that bands I andIIshow the
activity ofNPV. The detailed accounts will be
reportedelsewhere.
LITERATURE CITED
1. Bergold, G. H. 1953. Themultiplication of insect viruses,
p. 276-283.In P. Fildes andW. E. vanHeyningen (ed.),
Thenatureofvirus multiplication, 2nd Symposium Soc. Gen. Microbiol. Cambridge University Press, London and New York.
2. Bergold, G. H. 1958. Viruses ofinsect, p. 60-142. In C. Hallauer and K. F. Meyer (ed.), Handbuch der Virus-forschung, vol. 4. Springer, Vienna.
3. Bergold, G. H. 1963. Finestructureofsomeinsectviruse3.
J.InsectPathol.5:111-128.
4. Bergold, G. H. 1963. The natureof nuclear-polyhedrosis. viruses,p.413-456.InE. A.Steinhaus (ed.), Insect pathol-ogy, an advanced treatise, vol. 1. Academic Press Inic.,
NewYork.
5. Harrap, K. A., and B. E.Juniper. 1966.The internal struc-tureofan insect virus. Virology 29:175-178.
6. Himeno, M., S. Yasuda, T. Khosaka, and K. Onodera. 1968. The finestructureofanuclear-polyhedrosisvirus of
thesilkworm. J. Invertebr. Pathol. 11:516-519.
7. Kozlov, E. A., and I.P. Alexeenko. 1967.
Electron-micro-scope investigation of the structure of the nuclear-poly-hedrosis virus of the silkworm, Bombyxmori. J.Invertebr.
Pathol. 9:413-419.
8. Krieg, A. 1961. Uber den Aufbau und Vermehrungsmoglich-keitenvonstabchenformigen Insekten-Viren.II. Z.
Natur-forsch.16b:1 15-117.
9. Morgan, C., G. H. Bergold, D. H. Moore, and H. M.Rose.
1955. The macromolecular paracrystalline latticeof insect
viralpolyhedral bodies demonstrated in ultrathinsections examined intheelectronmicroscope. J. Biophys. Biochem. Cytol.1:187-190.
10. Morgan, C., G. H. Bergold, and H. M. Rose. 1956. Use of serial sections to delineate the structure of Porthetris dispar virus in the electron microscope. J. Biophys. Bio-chem.Cytol. 2:23-28.