Vol.65,No. 12
A
Second Neutralizing Epitope of
B19
Parvovirus Implicates the
Spike Region
in the Immune
Response
KOHJIYOSHIMOTO,1 STEPHEN ROSENFELD,1 NORBERTFRICKHOFEN,' DOUGLAS KENNEDY,2 ROBERTHILLS,3 SACHIKO KAJIGAYA,1AND NEAL S.
YOUNG'*
CellBiologySection, ClinicalHematology Branch, NationalHeart, Lung, and BloodInstitute, Bethesda, Maryland 20892,1 and Bureau ofBiologics, Drugs Directorate,2 and BureauofChemicalSafety,
Food Directorate,3 Ottawa, Ontario KJA OL2, Canada
Received12 July 1991/Accepted 16 September 1991
We used 18 monoclonal antibodies against B19 parvovirus to identify neutralizing epitopeson the viral
capsid. Of the 18 antibodies, 9 had in vitro neutralizing activityin abonemarrowcolony culture assay.The overlapping polypeptide fragments spanning the B19 structural proteins were produced in a pMAL-c
Escherichia coliexpressionsystemandusedtoinvestigatethebinding sitesof the neutralizing antibodies. One of thenine neutralizing antibodies reacted withboth VP1 andVP2capsid proteins and asingle polypeptide
fragmenton animmunoblot,identifyingalinearneutralizing epitope between amino acids 57 and 77 of the VP2
capsid protein. Eight of nineneutralizing antibodies failedtoreactwith eitherofthe capsidproteinsor any
polypeptide fragments, despite reactivities with intact virions in a radioimmunoassay, suggesting that
additionalconformationally dependent neutralizing epitopes exist.
Parvoviruses, the smallest DNA-containing animal
vi-ruses, are responsible for a wide variety of diseases in
vertebrates (18). In humans, diseases caused by B19
parvo-virusinclude erythema infectiosum (fifth disease) in children andapolyarthralgia syndrome in adults (20), transient
aplas-tic crisis inpersonswithunderlyinghemolysis (15),
sponta-neousabortion and hydrops fetalis inuteroinfection (1, 2, 9), andpersistent infection in immunocompromised hosts like patients with AIDS and those receiving chemotherapy for leukemia (11).
Theperiod of viremia is short in individuals with normal immune function and ends with formation of specific anti-bodies to the virus (16). In persistent infection, pooled humanimmunoglobulin is effective therapy (6). Despite the evident importance of antibody for viral clearance and resolutionofinfection, the humoralresponsetoB19 hasnot been well characterized and only a single neutralizing
epitope has been reported (17). We studied the reactivities of
a panel of anti-B19 neutralizing monoclonal antibodies
(MAbs) by using bacterium-expressed polypeptide frag-ments of the B19 capsid proteins. We identified a second
neutralizing epitope on the VP2 capsid protein. However, most of the MAbs appeared to recognize conformational ratherthanlinearepitopes.
MAbsweremadebyamodifiedcell fusionmethod(7, 10).
BALB/c mice wereimmunized intraperitoneally with 50 pLg
ofpurified B19 parvovirus in Freund's complete adjuvant and thenreceivedtwoboosterimmunizations of the antigen in Freund's incomplete adjuvant at 14-day intervals with a
trial bleeding atday 40. The mice subsequently received a
boosterintravenously 4 days before cell fusion. Spleen cells from the mice were fused with murine myeloma cell line Sp2/0-AG14 (AmericanType Culture Collection, Rockville,
Md.) bypolyethylene glycol 1500 (Sigma ChemicalCo., St.
Louis, Mo.). The hybridomas were cultured in Dulbecco's
modified Eagle medium containing 20% fetal bovine serum,
0.1 mM hypoxanthine, 0.4 ,uM aminopterin, 16 puM
thymi-*Corresponding author.
dine(HAT medium supplement; Sigma), and feeder layers of MEA(continuous BALB/ctissuegammairradiatedat12,000 rads), and 21 days after cell fusion, the supernatants of hybridomas were tested by radioimmunoassay against B19
parvovirus (3). For theradioimmunoassay, 6.4-mm-diameter polystyrene beads were coated by immersion ingoat anti-humanmuchain (Tago, Inc., Burlingame, Calif.) diluted in
0.05 M carbonate-bicarbonatebuffer, pH 9.6, atroom tem-perature for 3 h, and then washed withphosphate-buffered salinecontaining 0.05% Tween 20(PBST). The beads were
incubated for3 hat37°Cwith200 pul of humanserumknown tobeanti-B19parvovirus immunoglobulin M(IgM)antibody positive. Afterwashing with PBST, 200 p1ofpurifiedvirus,
diluted in PBST containing 10% fetal bovine serum to its working concentration, was added to the beads for 3 h at 37°Candtheywerewashedagain. The beadswereincubated with 200 ,ul of hybridoma supernatant for 18 h at room temperature and then incubated with
1251I-labeled
sheep anti-mouse immunoglobulin (Amersham Corp., Arlington Heights, Ill.)asthe secondantibodyatroomtemperaturefor 1 h. After washing of the beads, the radioactivity wasmeasured with a Gamma 4000 gamma radiation counter (BeckmanInstruments, Inc., Fullerton, Calif.).
Asummaryof the MAbs derivedfrom cell fusion is shown
in Table 1. Eighteenclones, named MAbs A, B, C, D, E, F, G, H, I, K, L, M, N,0,P, Q, R,S,reactive with B19 inthe
radioimmunoassay were isolated and cloned twice by the
limiting-dilution method. Established hybridoma clones
wereinjected into the peritoneal cavities ofPristane-treated
BALB/c mice, and ascites fluids were collected. Antibody
subclasses were determinedby enzyme-linked
immunosor-bentassayusingaHybridomaSubisotyping kit (Calbiochem
Co., SanDiego, Calif.).
In vitro neutralizing activity was assessed by measuring
the ability of the antibodytopreventvirus-induced toxicity
inaCFU-Eassay(2, 12). Serum obtained fromapatient with
sickle cell diseaseand transientaplastic crisisduringarecent B19 parvovirus epidemic was used as a virus source, and bone marrow mononuclear cells derived from normal
indi-vidualswereusedastargetcells. Asanegativecontrol, cells 7056
JOURNALOF VIROLOGY, Dec. 1991,p. 7056-7060 0022-538X/91/127056-05$02.00/0
CopyrightC 1991, AmericanSocietyforMicrobiology
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Summary of MAbs
MAb Subclass CFU-Ea
(%)Reactivity
in immunoblotA IgG2a 0 NDb
B IgG2a 92 None
C IgGl 0 ND
D IgGl 100 None
E IgGl 65 VPF6, SF3
F IgGl 82 None
G IgGl 0 ND
H IgGl 0 ND
I IgGl 0 ND
K IgGl 108 None
L IgG2a 0 ND
M IgGl 0 ND
N IgG2b 5 ND
0 IgGl 0 ND
P IgGl 81 None
Q IgGl 95 ND
R IgG2a 90 None
S IgGl 88 ND
aTheCFU-E assay results are means of each three experiments at an
antibody dilution of 1:75. % CFU-E was calculated with following formula % CFU-E = [colony number/(colony number of negative control - colony number ofpositivecontrol)] x 100.
bND, not done.
wereculturedwithout virus, andas apositivecontrol, cells werecultured with virus alone. With this assay, 9ofthe 18 clones (B, D, E, F, K, P, Q,R, and S)were neutralizingat titers between 1:150 and 1:3,750. Theneutralizing activities were between 65 and 108% at an antibody dilution of 1:75 (Table 1).
To determine the epitopes recognized by neutralizing antibodies, the recombinant VP1 and VP2 capsid proteins and the 11fusion proteins containing B19 parvovirus struc-tural protein polypeptide fragments were prepared and
immunoblotting
wasperformed. Preparation of the recombi-nantVP1 andVP2capsidproteinswasdescribedpreviously (8). The expression vectors were constructed as follows. DNA.segments encoding 11 recombinant proteins were amplified from pYT103c (a nearly full-length B19 cloned DNA) withadequateprimers by polymerase chain reaction. Polymerase chain reactions were performed byusing
astandard protocol in a total volume of 100 ,ul
containing
polymerase chain reaction buffer (10mMTris-HCl[pH 8.3], 50 mM KCI, 1.5 mM
MgCl2,
0.01%gelatin),
the four deoxynucleosidetriphosphates
at200,uMeach,2.5UofTaqpolymerase (Perkin
ElmerCetus, Norwalk,
Conn.),
ade-quate primers (21-mer), and pYT103c (a template). After purification of theamplified
DNAsfromanagarosegel, they
were inserted into a StuI site of maltose-binding protein vectorpMAL-c (Fig. 1A), which carriedatermination codon created
by
insertion ofanXbaI linker (NewEngland
Bio-labs, Inc., Beverly, Mass.) into a blunt-ended EcoRI site.TABLE 2. Amino acidsequences of shortfragmentsof VPF6
Fragment aminoNo. ofacids SequenceSqec
SF1 18 TFSRQFLIPYDPEHHYKV
SF2 19 YDPEHHYKVFSPAASSCHN
SF3 21 FSPAASSCHNASGKEAKVCTI
SF4 20 ASGKEAKVCTISPIMGYSTP
The sequencesof inserted portionswereconfirmedbyusing
a Sequenase kit (United States Biochemical). The fusion proteins ofB19 capsid sequences and maltose-binding pro-tein prepared for immunoblotting were designated VPF1, VPF2, VPF3, VPF4, VPF5, VPF6, VPF7, VPF8, VPF9, VPF10, and VPF11. Figure 1B shows the relationship of thesepolypeptides tothe amino acid sequences of the B19 capsid proteins. The B19-specific portion of each fusion protein ranged from 56 to 111 amino acids, and all 11 together spantheentire capsid protein sequence.
Thereactivity patternsofMAbs and VP1 and VP2capsid proteins and fusion proteins on immunoblot were analyzed byusing the materials andmethodsofthe Protein ImageKit (United States Biochemical). The isolated empty capsids wereboiled in sample loading buffer for 3 min. Escherichia colitransfected withpMAL-cfusion plasmidswascollected afterinduction with 0.3 mM
isopropyl-p-D-thiogalactopyran-oside. Bacteria were lysed in sample loading buffer and boiledfor 3min. Proteins were separated byelectrophoresis through asodium dodecyl sulfate(SDS)-8%
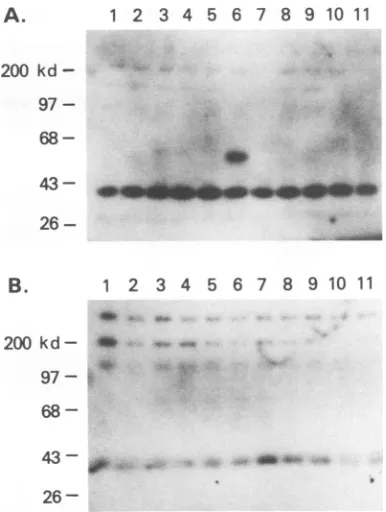
polyacrylamide gel and electrophoretically transferred to polyvinylidene difluoride membranes in transfer buffer containing 25 mM Tris and 192 mM glycine at 100 mA for 16 h. Membranes were blocked with 1% blocking solution, incubated with MAbs diluted 1:1,000, and incubated with alkaline phos-phatase-conjugated goat anti-mouse IgG (BRL). Antibody incubations were performed for 1 h at room temperature, andthe membranes were washedthree times between incu-bations with a salt buffer containing 0.1% Tween 20. Anti-body binding was detected with Lumiphos (United States Biochemical) chemiluminescent alkaline phosphate sub-strate.OnlyasingleneutralizingMAb, E, showed reactivity with bothVP1 and VP2 capsid proteins (Fig. 2). Since the entire sequenceof VP2is contained within VP1, MAb E recognizes a linear epitope of the VP2 capsid protein. MAb E reacted with fusion proteins of the expected sizes (53 to 54 kDa) on immunoblot, and thisreactivity was restricted to the VPF6 fusion protein (Fig. 3A and B). Todetermine in further detail the epitope recognized by MAb E, four short fragments of VPF6 were prepared by the same method and designated SF1, SF2, SF3, and SF4 (Table 2). When the reactivitiesof thesefour short fragments of VPF6 were tested by immuno-blot, only SF3 reacted withMAb E (Fig. 4), indicatingthat theepitope recognizedby MAb E waslocalizedto 21 amino acids lying between residues 57 and 77 ofVP2.
The genome of the B19 parvovirus isapproximately5.4kb long and contains two major open reading frames (5). The left open reading frame encodes the 71-kDa nonstructural protein (NS), and the right open readingframe encodes the two capsid proteins VP1 and VP2. The entire sequence of VP2 iscontained within VP1; the twoamino acid sequences are identical except for the amino-terminal 226 residues of VP1. Invirions, VP1representsonly about5% of the capsid proteins and theremainderis composed ofVP2 (3,4). While no detailed structural dataare available for B19 parvovirus, the structure of the related canine parvovirus (CPV) has been solved to the atomic level by X-ray crystallography (19).While thestructuralrelationshipof this virustoB19 can be extrapolated only tentatively, the amino acid sequences ofthecapsidproteins share41% homologyand 19%identity and mostmajor structural features appeartobe conserved in B19 parvovirus (Fig. 5). A single neutralizing epitope has been reported for B19 parvovirus and mapped to residues 328 to 344 of VP2 (17). This region
corresponds
to amino acids355 to373of CPV and contributes tothe spike ontheon November 10, 2019 by guest
http://jvi.asm.org/
7058 NOTES
polyl inker
A
&aIE
a 2.26
p tac
rrnD
ter inator r
Amp
lact
ori
Sac I kpn I Eag I haF I
I
11
1
I
I
I
Sft I
I
i
Polyt Inker: aUE...TCGAGC TCG GTA CCC GGCCGGGGA TCC ATC GAGCCT ACC CCT
hfa# I Xba I Sal I Pst I
I
I
1
II
1I
find Ill
I
I
GAATTC AGTAAA ACC CTCGATGGA TCC TCT AGA GTCGAC CTG CAGGCA AGC TTG...IacZa
B
781 a.s.
I
I
VP2
F
VPFI I- | VPF6
I-VPF2 I VPF7 |- I
[image:3.612.130.477.74.481.2]VPF3
|--VPF4
VPF5
VPF8
I-VPF9 I
VPFIO
I-VPFII I
FIG. 1. (A) B19-maltose-binding proteinfusionproteinswereprepared byinsertion of B19capsidgenefragmentsintoplasmid pMAL-c attheStuI restriction site.(B) Overlapping fragments ofthe B19 structuralproteingenesareshown with amino acid(a.a.)numbers and their positions within thesequencesofVP1 and VP2. From the aminoterminus,thepositionsof thepolypeptides in the amino acid sequence of the VP1 structuralproteinare asfollows: VPF1, 0to94; VPF2,52to142; VPF3,122to177; VPF4,158to227; VPF5, 187to265;VPF6,228 to334; VPF7, 314to423; VPF8, 403 to513; VPF9,493to603;VPF10,583to693;andVPF11,673to781.
threefoldaxis of thevirion. None ofourneutralizing MAbs
recognized this epitope, but MAb E recognized a region
from residues 57to77 of VP2 correspondingto71 to91 on CPV. Both neutralizing epitopes are also present on VP1.
These amino acids of CPV also fold to the spike. Two neutralizing epitopes mapping to residues Ala-300 and Asn-93 have been identified on CPV (14). Despite their
separationin the linearsequenceof thecapsid proteins, both of theseresidues alsomaptothespikeonthe threefold axis
and they may correspond structurally to the neutralizing regions identified on B19 parvovirus.
Of the nine neutralizing MAbs that we studied, only one
reacted with the capsid proteins and the peptide fragments
onanimmunoblot. These MAbswereoriginally screenedby
theradioimmunoassay, and under these conditions the
con-formationalstructureof the virion should bepreserved. It is
likely that the eight unreactive MAbs recognize
conforma-tionalepitopesthatarenolongerpresentonthe
membrane-transferred proteinin an immunoblot.
We havereportedthegenerationofneutralizingantiserum in animals immunized with B19 empty capsids containing
both VP1 and VP2 butnotVP2 alone(8). Theseresultsare
surprising given the identification of two neutralizing epitopes on VP2. It is suspected that the presence ofVP1 modulates theantigenicity of the viral particle and that the
antigenicityof the thoseepitopesistoolowtoberecognized
Eco RI
I
I
VPI
J. VIROL.
a
I
on November 10, 2019 by guest
http://jvi.asm.org/
1 2
200kd
-97
-68- -
83 kd
mp5-58
43
-1 2 3 4
200 kd
-97
-
68-43
-26
-FIG. 2. Thereactivitiesof MAb E and B19 capsid proteins were tested by immunoblotting. Ten microliters of purified empty capsids containingVP1and VP2capsid proteins were loaded on an SDS-polyacrylamide gel and blotted onto a polyvinylidene difluoride membrane. Lane 1 was probedwithMAb K, and lane 2 was probed with MAb E. kd, kilodaltons.
in theabsenceof VP1. The clinicalobservation that patients with persistent infection generate a specific antibody re-sponse that fails to neutralize the virus supports the idea that the neutralizing epitopes are of inherently low antigenicity. VP1may also directly affect theconformational structureof theneutralizing epitopes by its role in the formation of the structuralfeatures of B19, such as the spike on the threefold axis.
Determination of neutralizing epitopes is important for vaccinedevelopment. In some cases, linear epitopes recog-nized by neutralizing antibodies may be able to induce a
A.
200 kd
-97
-68
-
26-FIG. 4. The reactivity ofMAb E with shortfragmentsofVPF6 was analyzed by immunoblotting. Five microliters of bacterial lysates containing short fragments of VPF6 was loaded onto an
SDS-polyacrylamide gelandblottedonto apolyvinylidene difluoride membrane. Lanes: 1, SF1;2, SF2;3, SF3;4, SF4. Themembrane wastreated with MAb E.kd, kilodaltons.
1.MTSVNSAEASTGAGGGGSNSVKSMWSEGATFSA 33 1 MSDGAVQPDGGQPAVRNERATGSGNGSGGGGGGGSGGVG... ISTGTFNN 47
34 48
NSVTCTFSRQFL.IPYDPEHHPVSPIMG
QTEFKFLENGWVEITANSSRLVHLNHPESENYRRVVVNNMDKTAV|GNMA 8297
83 YS .. TPWRYLDFNALNLFFSPLEFQHLIENYGSIAPDALTVTISE 125
98 LDDIHAQIVTPWSLVDANAWGVWFNPGDWQLIVNTMSELHLVSFEQEIFN 147 126 IAVKDVTDK. TGGGVQV.TDSTTGRLCMLVDHEYKYPYVLGQGQDTLAPE 173 148 VVLKTVSESATQPPTKVYNNDLTASLMVALDSNNTMPFTPAAMRSETLGF 197
1 2 3 4 5 6 7 8 9 10 11
174 LPIWVYFPPQYAYL...TVGDVNTQGISGDSKKL.... ASEESAFYVL 214
198 YPWKPTIPTPWRYYFQWDRTLIPSHT.GTSGTPTNIYHGTDPDDVQFYTI 246
215 EHS.SFQLLGTGGTASMSYKFPPVPPENLEGCSQHFYEMYNPLYGSRLGV 263 247 ENSVPVHLLRTGDEFATGTFFFDCKPCRLTHTWQTNRALGLPPFLNSLPQ 296
0
43 -26
-264 PDTLGG...DPKFRSLTHEDHA...IQPQNFMPGPLVNSVSTKEGD 303
297 SEG4TNFGDIGVQQDKRRGVTQMGNTNYITEATIMRPAEVGYSAPYYSFE 346
304 SSNTGAGKALTGLSTGTSQNTRI6 EE YVTGINAIS 353 347 ASTQGPFKTPIAAGRGGAQTDENQAADGNPRYAFGRQHGQK..TTTTTGET 394
B. 1 2 3 4 5 6 7 8 9 10 11
200 kd-
_W
9
97-s
451 QPPPQIFLKILPQ.SGPIGGIKSMGITTLVQYAVGIMTVTMTFKLGPRKA 499
493 NCPGQLFVKVAPNLTNEYDPDASANMSRIVTYSDFWWKGKLVFK.AKLRA 541
68
-26
-FIG. 3. Antigen epitope recognized by MAb E detected by immunoblotting. Five microliters of bacterial lysates containing
each B19capsidprotein fragmentwasloadedonto
SDS-polyacryl-amide gels and blotted onto polyvinylidene difluoride membranes Lanes: 1, VPF1; 2, VPF2; 3, VPF3; 4, VPF4; 5, VPF5; 6, VPF6; 7, VPF7; 8, VPF8; 9, VPF9; 10, VPF10; 11,VPF11. Themembranes
weretreated with MAbsE(A)and K(B). kd, kilodaltons.
500 TGRWNPQPGVYPPHAAGHLPYVLYDPTATDAKQHHRHGYEKPEELWTAKS 549
[image:4.612.395.493.81.194.2]542 SHTWNPIQQM. SINVDNQFNYV..PSNIGGMKIVYEKSQLAPRKLY.... 584
FIG. 5. Comparisonof amino acidsequencesofthe VP2capsid proteinsof B19 and CPValignedformaximumhomology. Thelast
five amino acids of the B19 sequence do not contribute to the alignmentandarenotshown.Shadingin the upper line indicatesthe
binding sites of MAbs E and BEll. Shading in the lower line indicates neutralizing epitopesofCPV (residuesAla-300and
Asn-93). Theamino acids marked with stars arelocatedoutside ofthe CPV virion.
354 HGQTTYGNAEDKEYQQGVGRFPNEKEQLKQLQGLNMHTY..FPNKGTQQY 401
395 PERFTYIAHQDTGRYPEGDWIQNINFNLPVTNDNVLLPTDPIGGKTGINY 444
402 TDQIERPLMVGSVWNRRALHYESQLWSKIPNLDDSFKTQFAALGGW.GLH 450
445 TNIFNTYGPLTALNNVPPVYPNGQIWDK..EFDTDLKPRLHVNAPFVCQN 492
---a-mmmumm-mmb---Mm---maim
-_Mwmlq.__ lomppqmmw
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.110.267.83.198.2] [image:4.612.94.287.409.665.2]7060 NOTES
protective immune response in vivo. Unfortunately, the
neutralizing epitopes of many viruses are conformational,
and in thesecasespeptide vaccinesmaybe of limiteduse.In
addition, peptides are often poorly immunogenic. Despite
these drawbacks, the safety and simplicity of synthetic peptides make them attractive materials for vaccines. Fur-ther characterization of the neutralizing region we have
identified may allowits use as apeptide vaccine.
REFERENCES
1. Anand, A., E. S. Gray, T. Brown, J. P. Clewley, and B. J. Cohen. 1987. Human parvovirus infection in pregnancy and hydropsfetalis. N.Engl. J. Med. 316:183-186.
2. Brown, T., A. Anand, L.D.Ritchie, J. P.Clewley, and T. M. S. Reid. 1984. Intrauterine parvovirus infection associated with hydrops fetalis. Lancet ii:1033-1034.
3. Cohen, B. J., P. P. Mortimer, and M. S. Pereira. 1983. Diagnos-tic assays with monoclonal antibodies for the human serum
parvovirus-like virus (SPLV). J. Hyg. 91:113-130.
4. Cotmore, S.F., V. C. McKie, L. J. Anderson, C. R. Astell, and P. Tattersall. 1986. Identification of the major structural and nonstructural proteins encoded by human parvovirus B19 and mapping of theirgenes by procaryotic expression of isolated genomic fragments. J. Virol. 60:548-557.
5. Cotmore, S. F., and P. Tattersall. 1984. Characterization and molecular cloning of a human parvovirus genome. Science
226:1161-1165.
6. Frickhofen, N., J. L. Abkowitz, M. Safford, J. M. Berry, J.
Antunez-de-Mayolo, A. Astrow, R. Cohen,I.Halperin, L. King,
D.Mintzer, B. Cohen, and N. S. Young. 1990. Persistent B19 parvovirus infected with human immunodeficiency virustype 1
(HIV-1): a treatable cause of anemia in AIDS. Ann. Intern. Med. 113:926-933.
7. Galfre, G.,S. C. Howe, C. Milstein, C. W. Butcher, and J. C. Howard. 1977. Antibodiestomajor histocompatibility antigens produced by hybrid cell lines. Nature (London) 266:550-552. 8. Kajigaya, S., H. Fujii, A., Field, S. Anderson, S. Rosenfeld,L. J.
Anderson, T. Shimada, and N. S.Young. 1991. Self-assembled B19parvovirus capsids, produced inabaculovirus system,are
antigenically and immunogenically similar to native virions. Proc. Natl.Acad. Sci. USA 88:4646-4650.
9. Knott, P. D., G. A. C. Welply, and M. J. Anderson. 1984. Serologically proved intrauterine infection with parvovirus.Br.
Med.J. 289:1660.
10. Kohler, G., and C. Milstein. 1975. Continuous culture of fused cells secretingantibody ofpredefined specificity. Nature (Lon-don)256:495-497.
11. Kurtzman, G., B. Cohen, P. Meyers, A. Amanullah, and N. S. Young. 1988.Persistent B19parvovirusinfection as a cause of severe chronic anemia in children with acute lymphocytic leukemia. Lancetii:1159-1162.
12. Mortimer, P. P., R. K. Humphries, J. G. Moore, R. H.Purcell, and N.S. Young. 1983. A human parvovirus-like virus inhibits hematopoietic colony formation in vitro. Nature (London) 302: 426-429.
13. Ozawa, K., and N. Young. 1987. Characterization of capsid and noncapsid proteins of B19 parvovirus propagated in human erythroidbone marrowcell cultures.J. Virol.61:2627-2630. 14. Parrish, C. R., C. F. Aquadro, and L. E. Carmichel. 1988.
Canine hostrangeandaspecific epitopemapalongwith variant sequencesin thecapsid proteingeneof canineparvovirusand related feline, mink, andraccoon parvoviruses. Virology 166: 293-307.
15. Pattison, J. R., S. E. Jones, J. Hodgson, L. R. Davis, J. M. White, C. E.Stroud, and L. Murtaza. 1981. Parvovirus infec-tions and hypoplastic crises in sickle cell disease. Lancet i:664-665.
16. Saarinen, U. M., T. L. Chorba, P. Tattersall, and N. S. Young. 1986. Humanparvovirus B19-inducedepidemic red cellaplasia in patients with hereditary hemolytic anemia. Blood 67:1411-1417.
17. Sato, H., J. Hirata, M. Furukawa, N. Kuroda, H. Shiraki, Y. Maeda, and K.Okochi. 1991.Identificationof the region includ-ingtheepitope foramonoclonalantibody whichcanneutralize humanparvovirusB19. J.Virol.65:1667-1672.
18. Siegl, G. 1976. The parvoviruses, p. 1-109. Springer-Verlag, Vienna, Austria.
19. Tsao, J., M. S. Chapman, M.Agbandje, W. Keller, K. Smith, H. Wu, M. Luo, T.J.Smith,M. G.Rossmann, R. W. Compans, and C. R. Parrish. 1991. Thethree-dimensionalstructureofcanine parvovirus and its functional implications. Science 251:1456-1463.
20. Young, N. S. 1988. Hematologic and hematopoietic conse-quencesofB19parvovirus infection. Semin. Hematol. 25:159-172.
21. Young, N. S., P. P. Mortimer, J. G. Moore, and R. K. Humphries. 1984. Characterizationofavirusthat causes tran-sientaplastic crisis. J.Clin. Invest. 73:224-230.
J. VIROL.