0022-538X/85/080489-05$02.00/0
Copyright © 1985, American Society for
Microbiology
Poliovirus Protease 3C (P3-7c) Does Not Cleave P220 of the
Eucaryotic
mRNA
Cap-Binding
Protein
Complex
KEVIN A. W. LEE,' ISAAC EDERY,' RONNIE HANECAK,2t ECKARD WIMMER,2 ANDNAHUM SONENBERGl 3*
DepartmentofBiochemistry' and McGill Cancer Center,3 McGill University, Montreal, Quebec, CanadaH3GIY6, and Department of Microbiology, School of Medicine, State University ofNew YorkatStonyBrook, Stony Brook,
New York J17942
Received24 January1985/Accepted 19April 1985
Infection of HeLa cells by poliovirus results in proteolysis of the large subunit(P220) of the cap-binding protein complex. This is believed to cause the rapid shut-off of host protein synthesis during poliovirus infection.Inthiscommunicationweexamined thepossible involvement ofpoliovirus proteins3C(aproteinase)
and 2C in cleavage of P220. Using antisera against these two viral polypeptides, we wereunable to inhibit
proteolysisof P220 inanin vitroassay. Theseresults indicate that viralproteins3Cand 2C arenotdirectly
involved incleaving P220 and hence do notcauseshut-off of cellular protein synthesis.
The mechanism by which poliovirus inhibits HeLa cell protein synthesis,asubject of intense study for severalyears
(4), has recently been clarified in some respects. In vivo, polioviruscauses arapid and extensive inhibition of cellular (capped) mRNA translation, whereas translation of the naturally uncapped poliovirus RNA proceeds with high efficiency (1). Many lines of evidence have demonstrated thatthe failureofcappedmRNAsto enterpolysomes is due to avirally induced defect in the translation initiation ma-chineryof the host cell (forarecentreviewseereference4). Thefact thatcellextractsprepared frompoliovirus-infected
cellsarealsospecifically deficient inanactivity requiredfor capped mRNA translation (3, 11, 12, 14), andthusfaithfully
mimicthe in vivosituation, providedanassayfor the factor
which isinactivated. Consequently, it has been shown that thecap-binding protein (CBP) complex (also termed eIF-4F or CBP II) can restore translation of capped mRNAs in extracts from poliovirus-infected cells (3, 14) or in a re-constituted translationsystemfrompoliovirus-infected cells (5), and thus it isthought, that poliovirus achieves inhibition ofcellular protein synthesis by somehow inactivating the CBPcomplex.
The CBP complex consists of three polypeptides, the
24,000-molecular-weight CBP (also termed CBP I or eIF-4E), eIF-4A, and an -220-kilodalton (kDa) polypeptide (2, 7). Etchisonet al. (6) havepresented evidence which indi-cates that the 220-kDa polypeptide is cleaved by a virus-dependent protease, yielding cleavage fragments of -130 kDa.Subsequently,weisolatedamodified CBP complex (by using m7GDP affinity chromatography) from poliovirus-infected HeLa cells, which contains proteolytic fragments of P220 withapparentmolecularmassesof 110to130kDa (10). Although it remains to be demonstrated directly that proteolysis of P220 results in loss of activity of the CBP complex, itappearsmostreasonable that proteolysis of P220 is thecause of inhibition of cellular protein synthesis.
It is currently not known whether the protease activity
*Corresponding author.
tPresent address: Department of Molecular Biology and Bio-chemistry, University of California, Irvine, CA 92717.
which cleaves P220is encoded by the poliovirusgenomeor whether it is an induced cellular activity. The poliovirus
genome encodes at least one protease activity (protein 3C [8]) and mayhaveprotease activities mapping elsewhere in
the genome. Protein 3C (formerly known as P3-7c [13]) is
knownto process the viral polyprotein toproduce mostof the viral polypeptides by cleavage between the Gln-Gly amino acid pairs (8). There are, however, other cleavage sites(one Asn-Ser andtwoTyr-Gly) thatarenotcleaved by protein 3C. The protease(s) responsible for these other cleavage events is unidentified. It has been shown (8), however, that the activity does not appear to reside in
protein 2C (formerly known as P2-X [13]), as had been previously claimed (9).
Several studies have shown that the virus-dependent activity which is responsible for inactivating the CBP com-plex and consequently inhibiting cellular translation can be assayed in vitro. Originally, Rose et al. (12) showed that translational restoring activity (i.e., the activity which can restore capped mRNA function in extractsfrom poliovirus-infected HeLacells)can beslowly inactivatedupon incuba-tion with a cell extract from poliovirus-infected cells. We have confirmed these results (11) and have also shown that crude initiation factorpreparations from infected cells have anactivity whichcan slowlyimpair thecap-binding activity of polypeptides present in crude initiation factors from uninfectedcells(11). These observationsareconsistent with thecontention that the mechanismby whichcellularprotein synthesis is inhibited in poliovirus infected cells is catalytic innatureand isnotaresultof steric hindrancebysomeviral
protein. Finally, Etchison et al. (6) have shown that the protease activity which cleaves P220 of the CBP complex can be detected in crude initiation factors from poliovirus-infectedcells.
Using polyclonal antisera against P220 of sheep erythro-cytes,weprobedextractsfromeitheruninfected(U-S10)or
poliovirus-infected (I-S10) cells and obtained the same re-sults as Etchison et al. (6). The assay and immunoblotting procedures were as follows. HeLa S3 cellswere grown in mediumsupplementedwith5% calfserum.Poliovirustype1
(Mahoney strain) infection of HeLacells was performedas
489
on November 10, 2019 by guest
http://jvi.asm.org/
490 NOTES
12 3 4 5 6 7 8
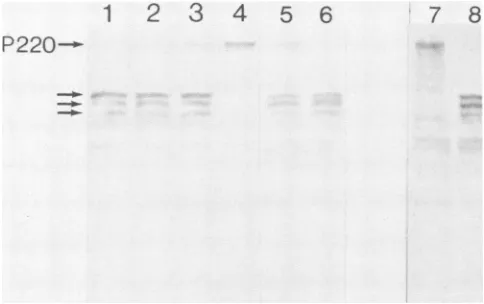
P220--FIG. 1. Invitroassayfortheproteasewhich cleaves P220 of the CBPcomplex. Lanes: 1, 10,ulofS10extractfrom uninfected cells
(U-S10) and 5 of S10 extract from poliovirus-infected cells (I-S10); 2, 10 ,ul of U-Si0 and 2.5 ,u ofI-S10;3, 10 pu1 of U-Sl0 and 1 pIofI-S10; 4, 10 ,ul of U-Sl0 only; 5,10 1.IofU-Sl0and 5 pu1 of I-S10 which were not preincubated; 6, a sample of the reaction
mixture usedinlane5, after 30 min ofincubationat37°C;7 and 8, U-Sl0andI-S10, respectively, whichwerenotincubated.
previously described with 10 to 20 PFU per cell, and preparation of cellextractswas aspreviously described (11, 12). Extractsweremixedas indicated below and incubated for30 min at 37°C. Reactions were stopped by addition of electrophoresis sample buffer and resolved on 10% poly-acrylamide gels containing sodium dodecyl sulfate. After electrophoresis, polypeptides were transferred to nitro-cellulose paper according to the method of Towbin et al., (15). Nitrocellulose blotswere presaturated with 1% bovine serumalbumin inTBS(10 mM Tris [pH 7.5], 150 mM NaCl) for 30minatroom temperature. Blots were incubatedwith anti-P220 antiserum in 1% bovineserumalbumin in TBSfor 3 hatroomtemperature. The antiserawereraised in rabbits against sheet CBP complex injected intradermally (16) as describedelsewhere (10) and werediluted 2,000-fold in 1% bovineserumalbumin in TBSbeforeuse. Blotswere subse-quently washed with six changes of TBSover aperiod of 30
min, followed by incubation with peroxidase-conjugated
goat anti-rabbit immunoglobulin G (Boehringer Mannhein Biochemicals) diluted 1,000-fold in 1% bovine serum albu-min in TBS for 1 h. Immunoreactive species were then visualized by staining with diaminobenzidene as described elsewhere (15).
Figure 1 is an immunoblot showing that the anti-P220
serumreactedmainly witha220-kDapolypeptidepresentin
the U-Sl0 extract (lane 7), whereas this polypeptide was absent in the I-S10 extract (lane 8). Instead, the antisera recognized in I-SlO polypeptides that were presumably cleavage products of P220 with molecular weights between 110,000 and 130,000 (lane 8). In an attempt to assay the protease activity in vitro, we mixed U-Sl0 and I-S10 and monitoredproteolysis of P220 by probing with anti-P220. As a control we incubated U-Sl0 alone for 30 min and found thatP220 is stable under these conditions (lane 4). We have repeated this experiment with many different cell extracts
and have never detected degradation of P220, even after
longer incubation times (data not shown), an observation suggesting that P220 is not intrinsically unstable. Lanes 1
through 3 show mixtures ofU-Sl0withdecreasingamounts
ofI-S10. The results show thataratio ofU-Sl0toI-S10of
2(10 ofU-Sl0 and 5 ,ul ofI-S10, in which bothextracts
contained
equal protein
concentrations as determined byA280/A260
readings) was sufficient to completely proteolyze P220 after30minofincubation(lane 1).
The samebands of 110 to 130 kDa in lane 8arealsoseeninlanes 1through
3. Itis reasonableto
assumne
that thedisappearance
ofP220fromU-S1O upon
mixing
withI-S10
occurred because the P220was cleaved to yield the smaller products. In accord with
thisassumptionwehaveperformedmorequantitativeassays
using 125I-labeled protein
Aand have detectedappearance ofthe
putative cleavage products (data
notshown).
With a U-S1O-to-I-S1O ratio of4,
there was almostcomplete
proteolysis
ofP220aftera30-minincubation(lane
2). Witha U-S1O-to-I-S1O ratio of 10, there wasclearly
some P220 remaining after 30min(lane 3). Thus,aU-S1O-to-I-S1O ratio of 4wasapproximately
theendpoint
for titration of theI-SlO extractagainst
theproteaseactivity
underourassaycondi-tions. Lanes 5 and 6 show that the loss of P220 is time
dependent.
Lane 5 shows P220-related antigens after asimple mixing
of U-S1O and I-S10 withoutincubation,
and lane 6 shows aportion
ofthe samesample
after30 minof incubation. Theintensity
ofthe P220antigen
in lane 5 isweaker than thatofthe
P220
cleavageproducts, although
theopposite
might
beexpected
based ontheamountsofU-Sl0
and I-S10 added. Thisapparentcontradiction most
probably
resulted from the fact that P220 was less
efficiently
trans-ferred to nitrocellulose paper than thecleavage products
under our transfer conditions. It is alsopossible
that thecleavage
products
reacted better with theantibody
on nitro-cellulose paper than P220. It should be noted that theincubations shown in lanes 5 and 6 are from a different
experiment
than those in lanes 1through
4.Thus,
theabsolute
amountofP220detectedwasless inlane 5 thaninlane4dueto variationin
staining intensity
betweenexperi-ments. Insummary, the results
presented
inFig.
1 confirmprevious
reports(6)anddemonstrate that there isaproteaseactivity
in extracts frompoliovirus-infected
HeLa cells which candegrade P220.Wenextwantedto determinewhether
poliovirus proteins
3C or2C are involved in P220proteolysis.
Protein 3C is alikely
candidate for such a protease, since it is the viralprotein
involved in mostof thecleavages
of viral precursorpolypeptides
toyield
both structural andnonstructuralpro-teins (8). Ithas been
reported
thatprotein
2Chas proteaseactivity
involved inprocessing
ofpoliovirus protein
pre-cursors
(9),
butthiswasnotverified ina more recentstudy
(8).
Initially
we tested theactivity
of ourpreparations
of antibodiesagainst
proteins
3C and 2C. Anti-3Cis knowntoinhibit
cleavages
atGln-Gly
pairs
which occurwhenproc-essing
of viralprecursors isassayed during
invitro transla-tion ofendogenous
poliovirus
RNA in extracts frompoliovirus-infected
cells(8).
Consequently
we used thisassayto testthe
activity
of anti-3C. Translation incubations with extractsfrompoliovirus-infected
HeLacells were car-riedoutaccording
tothe methodofLeeandSonenberg (11),
except that theextracts werenotnuclease treated. Thecell extract waspreincubated
for 60 minat4°C
withantibody
buffer or thedesired
antibody.
The translation incubations werethenperformed.
Reaction mixtures contained(in
atotal volume of 25,ul)
9RI
of cell extract, 130 mMpotassium
acetate, 0.4 mM
magnesium
acetate, 20 mM HEPES[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid,pH
7.5],
1 mMATP,
200,M
GTP,9 mMcreatinephosphate,
22mgof creatinephosphokinase
perml,
2.5 mMdithiothreitol,
0.2mM
spermidine,
19amino acids(10 ,uM
each,
minusmethio-nine),
20,uCi
of[35S]methionine
(>1,000
Ci/mmol;
New J.VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.43.285.57.209.2]1 2 3
I
4
5
6
7
8
-v
1ABCD-
3ABCD-3CD---- _
} a
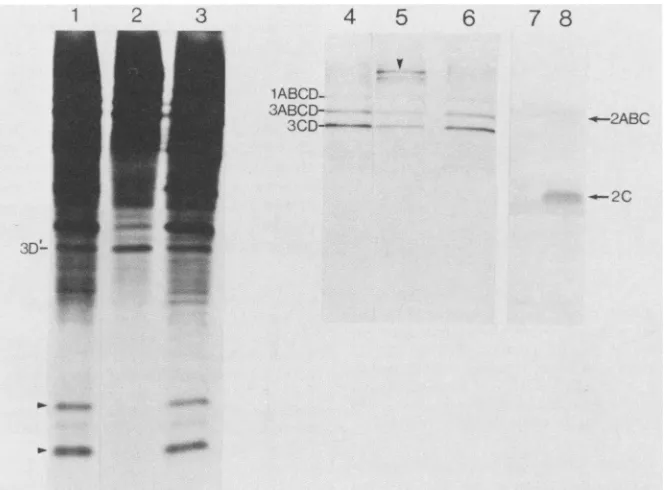
FIG. 2. Effects of anti-3C and anti-2Conprocessingofpoliovirusprecursorproteins. Lanes 1through3 and 4through6 showdifferent
exposuresof thesameautoradiograph (exposuretimeswere10 h and 5min, respectively).Lanes: 1 and4,translationproductsintheabsence
ofantibody;2 and5,translationproductsin thepresenceof 60,ugofanti-3C;3 and6,translationproductsin thepresenceof 60pLgofanti-2C; 7 and 8, animmunoblot ofextractsfrom uninfected (lane 7)and infected (lane 8)cells probedwithanti-2C. Blotting conditions were as
describedinthetext.
England Nuclear Corp.), and 6 ,ul of antibody buffer (10mM
Tris[pH 8.0], 10 mMKCI)orimmunoglobulin G fractions of the antibodies indicated below. Antisera topoliovirus pro-teins 3C and 2C and purification of immunoglobulin G
fractionwas asdescribed elsewhere (8). After incubation for 60 min at 37°C, samples were resolved on 10% sodium dodecyl sulfate-polyacrylamide gels, followed by autoradi-ography.
Figure 2 (lanes 1 through 6) shows the [35S]methionine-labeled proteins produced in an extract from poliovirus-infectedHeLa cells. Lanes 1through 3 and4through 6 show different exposures of the samegel. Lanes 1 through 3 are overexposed to show the lower-molecular-weight polypep-tides. Lanes 4through 6 areunderexposed to showclearly thehigher-molecular-weightprecursors.Lanes 1 and 4 show endogenous translation products in the absence of antibody. Major bands which correspond tothe three precursor pro-teins,asdetermined from theirmigration relativeto molecu-lar weight markers (1ABCD, 3ABCD, and 3CD; formerly P1-la, P3-ib, and P3-2, respectively), are indicated to the left oflane 4. Inaddition thereare several lower-molecular-weight bands which correspondtothe various viral proteins derivedfrom thehigher-molecular-weightprecursors. Addi-tion of anti-3C to the translation incubation resulted in inhibition ofprocessing,asindicatedby the disappearance of thelower-molecular-weight bands and the build-up of high-er-molecular-weight polypeptides (lanes 2 and 5). The high-molecular-weight polypeptide in lane 5 (indicated by an arrowhead) migratedas an -150-kDa polypeptide and con-sisted of the combined amino acid sequences of 2ABC and 3ABCD as previously shown (8). Inhibition of 3C activity was specific. This is indicated by the fact that the Tyr-Gly cleavage that produces protein 3D' (formerly known as P3-6b) and which is notcarried outby protein 3C was also
not inhibitedby anti-3C, as previously shown (8; compare lanes1 and2). Densitometry of the lower-molecular-weight bands(indicated by arrowheadstothe leftof lane 1)showed
that under the conditions ofourassay,morethan90% of 3C activitywas blocked by anti-3C. Additionofanti-3Ctothe
translation incubation had no effect onthe total incorpora-tion of[35S]methionine into trichloroacetic acid-precipitable material(datanotshown). Lanes 3 and 6 showtheeffectsof
anti-2Conpoliovirus proteinprocessing. Itcanbeseenthat
anti-2C had no effect on processing of poliovirus polypeptides,aspreviously shown (8).To ascertain that the anti-2C antibodywasactive, weprobedHeLa cellextracts from uninfected and poliovirus-infected HeLa cells. The immunoblot is shown in lanes 7 and 8, representing the uninfected and infected cell extracts, respectively. The
antibody reacted with protein 2C in extractsfrom infected cells(lane 8) andgavenoreaction witha similar-molecular-weightpolypeptide inextractsfromuninfected cells (lane 7). There was also a weak reaction with a higher-molecular-weight band in extracts from infected cells which most probably correspondedtotheprecursorpolypeptide 2ABC. Thus, weconclude that the antibodies weusedwere active in inhibiting the activity or recognizing their cognate anti-gens. However,itshouldbeemphasized that the onlyassay
wehavefor the anti-2Cantibodywasimmunoreactivityona nitrocellulose blot, and it is possible that anti-2C cannot
inhibit theenzymatic activity of protein 2C.
Wenextasked whetheranti-3Coranti-2C could inhibit the proteolysis of P220. The protease assay was performed
under the conditions used for Fig. 1, lane 1, toensure that efficient proteolysis was achieved but that the protease activity was not in vast excess. Figure 3 (lanes 1 and 2) shows the U-Sl0 extract and a mixture of the U-Sl0 and I-S10extracts, respectively, incubated for 30 min.In lanes3
30---2ABC
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.142.474.70.315.2]492 NOTES
12 3 4 5 6 7 8 Ehrenfeld (Proc.
Natl.
Acad. Sci. U.S.A., inpress),
whichdemonstrated that 3C activity can be separated from the P220 proteolyzing activity and that antibodies against
pro-tein 3C donotinhibit P220proteolytic cleavage.
If proteins 3C and 2C are not directly responsible for cleavage of P220, then the question of the identity of this protease remains unanswered. It is possible that a hitherto-uncharacterizedpoliovirus-encoded protease is involved, or alternatively that poliovirus infection induces a cellular activity that cleaves P220. If the latter is true, it would be
interestingto know whether such anactivity plays a role in
[image:4.612.72.285.73.219.2]regulationof protein synthesis in situations other than during poliovirus infection of HeLa cells.
FIG. 3. Effects of anti-3Cananti-2Conproteolysis of P220. The
protease assay was carried out and samples were processed for
immunoblottingasdescribed in thetext.Lanes: 1, 4,ul of U-S10; 2
through8, 4,ul of U-Sl0 and2,ul ofI-S10;3through 5, 2, 10, and 20
,ug of anti-3C, respectively;6through8, 2, 10, and 20,ug of anti-2C, respectively. Theextractswerepreincubated for60minat4°C with the desired antiseraasdescribed fortheexperimentshownin Fig.2.
through 5, increasing amountsof anti-3C were addedto the incubation under the same conditions as for the in vitro translation experiments. That is, the infected extract was preincubated with anti-3C for 60 min at 4°C. The highest amount of antibody added (expressed as micrograms of antibodypermicroliter of I-S10)wasinexcessof theamount which resulted inmorethan90% inhibition of the 3C activity (lane 5). It is clear fromourdata that anti-3C hadnoeffecton the protease activity whichcleaved P220, as evidenced by the absenceof P220 in lanes 3 through 5. Lanes 6 through 8 show that anti-2C (added in the same amounts as anti-3C) also hadnoeffecton proteolysis of P220. We conclude that the activity which cleaves P220 is not the same as that (protein 3C) which cleaves poliovirus precursor polypep-tides. The data alsosuggests thatprotein 2C is not directly involved in proteolysis of P220.
The resultspresented here and in previousreports estab-lishthat P220of the CBP complex is proteolytically cleaved in poliovirus-infected cells. However, it remains to be proven that the cleavage is indeed the cause for loss of activity of the CBP complex. The identification of a viral
proteaseresponsible for the degradation of P220 would lend support tothe proposedmechanism of inhibition of host cell protein synthesis. The results shown here indicate that the poliovirus proteinase 3C is notinvolved in the cleavage of P220, because anti-3C antibody didnotinhibit P220cleavage under the same conditions as itinhibited poliovirus protein cleavage. A similar conclusion canbe made forpolypeptide 2C, but with the reservationpointedoutabove.
It remainspossible that 3C amino acid sequences harbor different proteaseactivities dependingon whetherthey are partofhigher-molecular-weight forms (3CDor3C')orof the
matureform of protein 3C. If thiswerethecase,then anti-3C might not block both putative activities, thus raising the
possibility that protein 3C might still be involved in
proteolysis of P220. However, the fact that the polyclonal antisera used in our experiments recognized all forms of
protein 3C (8) argues against this possibility. In anyevent,
the main conclusion from our data is that the activity required for processing ofpoliovirusprecursors at the Gln-Gly amino acid pairs and the activity which proteolyses P220
are not the same. Our conclusion is in accord with recent
results obtained by R. E. Lloyd, D. Etchison, and E.
We thank Ellie Ehrenfeld forcommunicating her results to us
beforepublicationand Alice Newman forvaluablehelpin someof
theexperiments.
This research was supported by agrant from the Medical Re-search Council of CanadatoN.S. and PublicHealth Servicegrants AI-15122 and CA-28146 from the NationalInstitutes of Healthto E.W. N.S. isarecipientofaTerryFox CancerResearch Award
fromtheNational CancerInstitute ofCanada. K.A.W.L.andI.E.
arerecipientsofpredoctoralresearchfellowships fromtheCancer
ResearchSociety (Montreal).
LITERATURECITED
1. Baltimore, D. 1969. The replication of picornaviruses, p.
101-176. In H. B. Levy (ed.), The biochemistry of viruses.
Marcel Dekker, Inc.NewYork.
2. Edery, I., M.Humbelin,A.Darveau, K. A. W. Lee,S. Milburn, J. W. B. Hershey, H. Trachsel, and N. Sonenberg. 1983. In-volvementof eukaryotic initiation factor4Ain thecap
recogni-tionprocess.J. Biol. Chem.258:11398-11403.
3. Edery, I., K. A. W. Lee, and N. Sonenberg. 1984. Functional characterizationofeukaryoticmRNAcapbinding protein
com-plex: effects on translation ofcapped andnaturally uncapped
RNAs.Biochemistry23:2456-2462.
4. Ehrenfeld, E. 1982. Poliovirus-induced inhibition of host cell
protein synthesis.Cell 28:435-436.
5. Etchison, D., J. Hansen, E.Ehrenfeld,I.Edery, N.Sonenberg,S.
Milburn, and J. W. B. Hershey. 1984. Demonstration in vitro thateucaryoticinitiation factor 3 is activebut thatacap-binding protein complexis inactive inpoliovirus-infected HeLa cells. J.
Virol. 51:832-837.
6. Etchison, D., S. C. Milburn, I. Edery, N. Sonenberg, and
J. W. B.Hershey. 1982. Inhibition ofHeLacellprotein
synthe-sisfollowing poliovirusinfection correlateswith theproteolysis
of a 220,000-dalton polypeptide associated with eucaryotic
initiation factor 3 andacapbinding protein complex.J. Biol. Chem. 257:14806-14810.
7. Grifo,J. A.,S. M.Tahara,M. A. Morgan, A.J.Shatkin,and
W. C. Merrick. 1983. New initiator factoractivity requiredfor
globinmRNA translation. J. Biol. Chem.258:5804-5810. 8. Hanecak, R.,B. L.Semler, C. W.Anderson,and E. Wimmer.
1982. Proteolytic processing of poliovirus polypeptides: anti-bodies to polypeptide P3-7c inhibit cleavage at glutamine-glycine pairs. Proc.Natl. Acad. Sci. U.S.A. 79:3973-3977. 9. Korant, B., N.Chow, M. Lively,and J. Powers. 1979.
Virus-specifiedprotease inpoliovirus-infectedHeLa cells. Proc.Natl.
Acad. Sci. U.S.A. 76:2992-2995.
10. Lee, K. A.W.,I.Edery,and N.Sonenberg.1985. Isolation and
structural characterization of cap-binding proteins from
poliovirus-infectedHeLa cells. J. Virol.54:515-524.
11. Lee, K. A. W., and N. Sonenberg. 1982. Inactivation of
cap-binding proteins accompanies theshut-offof hostprotein syn-thesis by poliovirus. Proc. Natl. Acad. Sci. U.S.A. 79: 3447-3451.
12. Rose, J. K., H. Trachsel, K. Leong, and D. Baltimore. 1978. Inhibition of translationby poliovirus:inactivation ofaspecific
initiation factor. Proc. Natl.Acad. Sci. U.S.A. 75:2732-2736. 13. Rueckert, R.R.,and E. Wimmer. 1984. Systematic
nomencla-
P220--J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
tureofpicornaviral proteins. J. Virol. 50:957-959.
14. Tahara, S., M.A. Morgan, and A. J. Shatkin. 1981. Two forms ofpurified m7G-cap binding protein with different effects on capped mRNA translation in extracts of uninfected and poliovirus-infected HeLa cells. J. Biol. Chem. 256:7691-7694. 15. Towbin, H.,T.Staehelin,andJ. Gordon. 1979. Electrophoretic
transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedures and some applications. Proc. Natl. Acad. Sci. U.S.A. 76:4350-4354.
16. Vaitukaitis, J.L. 1981. Production of antisera with small doses of immunogen: multiple intradermal injections. Methods Enzymol. 73:46-52.