Copyright@ 1972 American Society for Microbiology
Proteins Specified
by Herpes Simplex
Virus
VIII.
Characterization
and
Composition
of Multiple
Capsid
Forms
of
Subtypes
1
and
2
WADE GIBSON AND BERNARD ROIZMAN
Department of Microbiology, The UniversityofChicago, Chicago,Illinois60637
Received for publication7August1972
Two classes of herpesviruscapsids, designated A and B, wereisolated from the
nuclei of humancells infected with herpes simplex virus (HSV). A andB capsids
share incommonfour structural proteins, i.e., no. 5, 19,23, and24. Bcapsids
con-tain 7.7 to 9.7 times moredeoxyribonucleic acid than A capsids; moreover, they
contain proteins no. 21 and 22a in addition. All of the proteins contained in the
capsid except no. 22aare present in the enveloped nucleocapsids (virions) in
ap-proximatelythesamemolar ratios.The capsid proteins ofHSV-1 cannotbe differ-entiatedfromtheirHSV-2counterpartswithrespecttoelectrophoretic mobility. A
thirdclass of capsids, designatedC capsids, wasisolated fromvirionscontainedin the cytoplasm of infected cells by the same procedure used to obtain A and B
capsids.The Ccapsidscontain allof the proteinspresentin A capsids plus proteins
1to3and 21.
A preceding paper (25) inthis series reported on the composition, electrophoretic mobility,
and molecular weight of proteins comprising the herpes simplex virus (HSV) virion.
Subse-quently (13) it was shown that a number of the
glycosylated proteins present in the virion were
also present in purified membranes of infected
cells. The conclusion that these glycoproteins
were constituents of the envelope of the virion wasreinforcedbythe observation that NP40, a
mild nonionic detergent, partially stripped them fromthesurfaceof thevirion (25). The question
still remained astowhich proteinsare the
struc-turalcomponentsof the viral capsid.
Pertinent to the studies of the composition of
viralnucleocapsids reportedinthispaper arethe
following observations and reports. (i) The
nucleocapsid assembles in the nucleus, and in
cellsinfected withmost herpesvirusesit acquires
anenvelopeasit budsthrough the innerlamella of the nuclear membrane (3, 22). (ii) Electron
microscopy studies (18-20, 23, 24) have shown that at least two types of capsids are present
within the nucleus of HSV-infected cells, and thattheyappeartodiffer, atleast withregardto
staining properties, from the capsid contained within virions accumulating in the perinuclear
space orcysternaeoftheendoplasmicreticulum. (iii) The threemost currentdescriptionsof
herpes-virus nucleocapsids (1, 14, 21) have been based
on structures obtained by widely differing
tech-niquesand,notsurprisingly,arenotinagreement
about the exact protein composition.
Further-more, two of thethreedescriptions (14, 21) are
based on radioisotopic profiles obtained by
sectioning the analytical acrylamide gels-a technique that does not afford ashigh a
resolu-tion as absorbance profiles from either stained
gelsorautoradiograms.
Weare presentingevidence here insupport of
thefollowing: (i)two classesofcapsids,differing
withrespect tobothdeoxyribonucleicacid(DNA)
content and protein composition, are present in
the nucleiof HSV-infectedcellsand (ii) the
pro-tein composition of capsids derivedby stripping
of theenvelope from virions differs from that of eithertypeofintranuclear
capsid.
MATERIALS AND METHODS
Virus and cells. ThepropertiesofHSVsubtypes 1
and 2used in these studies have been described
else-where (7, 15, 25). Enveloped virions were produced
in HEp-2 cells, and the intranuclear capsids were
produced in either HEp-2 or Vero (simian) cells.
Pertinentinformation aboutthegrowth,maintenance,
infection, andradioisotopic labelingofcells hasbeen
presented in detail by Spear and Roizman (25).
Deviations from these general techniques are
indi-cated in thetext.
Isotopes and reagents. The isotopes and reagents
were reconstituted 3H-protein hydrolysate (mixture
no. 3130-08, 1 mCi/ml) and thymidine-2-14C (no.
2523-96, 10 /ACi/mi) from Schwarz/Mann,
Orange-burg, N.Y.; uniformly labeled 14C-L-amino acid
mixture (mixture no. NEC-445, 100 ,Ci/ml), and
1044
PrintedinU.S.A.
on November 10, 2019 by guest
http://jvi.asm.org/
D-glucosamine-1-14C (NEC-193X, 100,Ci/ml) from New EnglandNuclear Corp., Boston, Mass.;
acryl-amide from K & K Laboratories, Plainview, N.Y.;
N,N'-methylenebisacrylamide and N,N,N' ,N'-tetra-methylethylenediamine (TEMED) from Eastman Chemicals, Rochester, N.Y.; dextran 10 from
Phar-macia, Uppsala, Sweden; Coomassie Brilliant Blue
fromColabLaboratories, Inc., Glenwood, Ill.;
ultra-pure urea from Mann Research Laboratories, New
York, N.Y.;NonidetP-40, agiftfrom ShellOilCo.,
New York, N.Y.; sodium desoxycholate from Schwarz/Mann, New York, N.Y.; BRIJ 58 (poly-oxyethylene [20] cetyl ether) from Atlas Chemical Industries, Inc., Wilmington, Del.; herring sperm DNA, typeIV,fromSigmaChemicalCo.,St. Louis,
Mo.; bovine serum albumin (crystallized A-grade)
from Calbiochem, Los Angeles, Calif.; and deoxy-ribonuclease I (electrophoretically purified) from Worthington Biochemical Corp., Freehold,N.J.
Isolation of intranuclear virus particles. Cells were
harvested between 18 and 24 hr after infection;
col-lected by centrifugation at 1,500 X g for 10 min (low-speed centrifugation); and resuspended (5 X 107 cells/ml) in 0.15 M NaCl, 0.01 M tris(hydroxy-methyl)aminomethane (Tris), pH 7.2, and 0.002 M MgCl2containing
1%c
NP40.After incubationfor 30min at 0 C, the nuclei were sedimented from the
suspension bylow-speedcentrifugation andlysedby
adding sodium deoxycholate (DOC) to a final
con-centrationof 0.5%. Thelysatewasgentlywarmed in
the presence of deoxyribonuclease (50 ,ug/ml) to
decrease the viscosity. BRIJ-58 and urea were then
added to final concentrations of 0.5% and 0.5 M,
respectively, and the extract was clarified by
low-speed centrifugation. The supernatant fluid was
layered on top of linear, 10 to 40% (w/w) sucrose density gradients,prepared in 0.15 M NaCl and 0.01 M sodium phosphate buffer (pH 7.2), and centrifuged
for 60 min at 23,000 rev/min and 4 C in a Spinco
SW27 rotor. Two light-scatteringbandslocated near
themiddleof the tube wereresolvedduring the course ofcentrifugation. Each was recovered mechanically
fromthetopof the gradient and eitheranalyzed
im-mediately,orfrozen and stored at -20 C.
Isolation of enveloped virions. Enveloped virions
were isolated essentially as described by Spear and
Roizman (25). Briefly, infected cells were swollen in
hypotonic buffer and disrupted by several strokes
with a Dounce homogenizer, and the cytoplasmic
fraction was layered onto 3 to 30% dextran-10
gradients and centrifuged at 23,000 rev/min for 60
min in a Spinco SW25.3 rotor at 4 C. Enveloped
virions sedimentedto apositionjust above the middle
of the tube, and formed alight-scattering zone that
wascollected mechanicallyfrom the top of the tube.
Thismaterialwas not furtherpurified.
Preparation ofdetergent-treated cytoplasmicvirions.
(i) NP-40-extracted cytoplasmic virions were
pre-pared by suspendingapproximately 108infected cells
in 1 ml of 0.15 M NaCl, 0.01 M Tris (pH 7.2), and
0.002 M MgCl containing 1% NP-40. After
incuba-tion for 20min at 0 C the nuclei were removed from
the lysate by low-speed centrifugation. The lysate wasfurtherclarified by a secondlow-speed centrifuga-tion.
(ii) NP-40/DOC-extractedcytoplasmicvirionswere
prepared by further processing a clarified NP-40 lysate,obtained as above, with DOC (0.5%),
deoxy-ribonuclease (50 Mug/ml), BRIJ-58 (0.5%), and urea
(0.5M) in thesamefashionasdescribed above for the
isolation of intranuclear virus particles. The lysate
wasclarifiedby low-speedcentrifugation.
Both NP40 and NP-40/DOC lysates were
sub-jected to rate-velocity sedimentation on 10 to 40%
(w/w), linear sucrose density gradients prepared in
0.15 M NaCl and 0.01 M sodiumphosphate (pH7.2)
andcentrifugedat 23,000rev/min for 60 min at4 C inaSpincoSW27or SW25.3rotor.One visible band
wasresolvedfrom eachlysate atabout themid-point
of the tube; itwas collected mechanically from the
top ofthegradientand stored at -20C.
Analytical gel procedures. The electrophoretic,
staining, and autoradiographic techniques used in
these studies have beendescribed in detail bySpear
and Roizman (25). The important features are as
follows. (i) Samples to be analyzed were solubilized
by heatingat 100C for 2 minin the presenceof2%
sodium dodecyl sulfate (SDS), 5%
beta-mercapto-ethanol, and 50 mm Tris (pH7.0). Solubilized
prep-arations wereapplied to thegelsin 50- to 150-Militer
volumes generally containing between 50 and 100,ug
ofprotein. (ii) Discontinuous SDS gels (4, 16) were
formed in 12-cm glass tubes cleaned by soaking
overnight inasolution of concentrated sulfuric
acid-dichromic acid and rinsed thoroughly with water,
rather thansiliconizedasrecommendedin theoriginal
procedure. The main gel (8.5 cm in length) contained
8.5% acrylamide; the stacker gel (2.0 cm inlength)
contained 3% acrylamide. (iii) Proteins in the gel
were fixed and stained with 0.03% Coomassie
Bril-liant Blue in 10% acetic acid and 25% 2-propanol
according tothe procedureofFairbankset al. (10). Destainingwas in 10% acetic acid, 10% 2-propanol. (iv) Autoradiograms were made from gels sliced longitudinallyanddried asdescribed byFairbanks(9).
A mechanical press was used to hold the dried gel
firmly against the X-ray film during exposure. (v)
Absorbance profiles were made from stainedgelsand
autoradiograms, both scanned at 550 nm, with the Gilford linear transport attachment and Gilford recordingspectrophotometer.
Protein and DNA analyses. Protein concentrations
were determined by the Lowry procedure (17) as
modified by Eggstein and Kreutz (6). Bovine serum
albuminwasused as the standard.
DNA concentrations were determined by the
di-phenylamine technique described by Dische (5) as modified by Burton (2). Herring sperm DNA was
used as thestandard.
Electron microscopy. Thin sections were prepared
forelectronmicroscopyexamination from pellets of
A and Bcapsids.Briefly, capsidsuspensionsrecovered
from nuclear lysates as described above were
sub-jected to centrifugation at 25,000 rev/min for 2 hrat
4 Cin aSpinco SW27 rotor. Thepelletsthat were
ob-tained were fixed in gluteraldehyde, postfixed in
osmium tetroxide, embedded in Epon, and stained
withuranylacetateandleadcitrateprior to
examina-tion in anAEI-EM6Bmicroscope. A detailed
descrip-tion of this procedure and of the technique used to
1045
VOL. 10, 1972
on November 10, 2019 by guest
http://jvi.asm.org/
prepare wholemountsnegatively stained with sodium phosphotungstic acid has been published elsewhere (25).
RESULTS
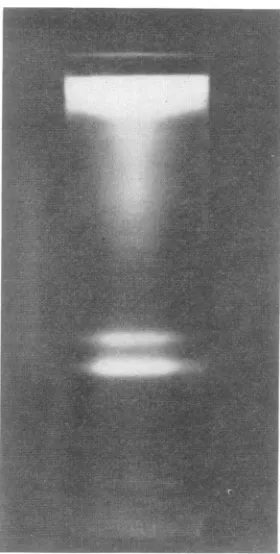
Centrifugation of nuclear lysates. Nuclear ex-tractspreparedasdescribed above, on centrifuga-tionthroughadensity gradient, formedtwobands designated astop and bottom (Fig. 1). A third
band occasionally appeared near the bottom of
the tube; its occurrence generally indicated in-complete solubilization of cytoplasmic
mem-branes with NP-40. The bands shown in Fig. 1 were obtained from approximately 109 cells
infected withHSV-2.
Somevariability has been noted in theamount
ofmaterial in the top and bottom bandsrecovered from different
preparations;
however, severallines of evidence indicate that it does not result
from the
extraction
procedureperse.Specifically, (i) extending NP-40 extraction to 24 hr at 0C or substituting Triton X-100 for NP-40,(ii)
changing theNP-40 extractionto15minat37C, and (iii)
allowing
thefinalextracttostandfor24 hr at 0 Cprior
to centrifugation do not visibly affect theyield
ofthe material in either the toporbottom bands.
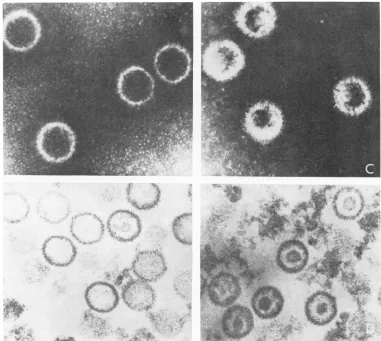
Electron microscopy. Electron
microscopy
studies revealed the presence ofcapsids in both
top and bottom bands. Enveloped or
partially
enveloped
particles
were absent. In general thecapsids
in each band were uniform in size andappearance. In
negatively
stained(sodium
phos-photungstic acid) preparations,
thecapsids
in thetop band werefilled with
stain,
whereas thosein the bottom band were filled onlypartially,
delineating an internal structure
(Fig. 2A, C).
Examination of thin sections oftop-and bottom-band material
(Fig.
2B,D)
stained withuranyl
acetate and lead citrate showed that
top-band
capsidsappeartobe empty, whereasbottom-band
capsids
containacentrally located, densely
stain-ing structure. Cross-contamination ofthe bands
was estimated by
particle
counts to be lessthan 10%.Forthe sakeof
brevity
anduniformity,
weshall designate the particles in the top and bottombands asA andB
capsids,
respectively,
withoutdifferentiating between the terms
capsid
andnucleocapsid.
DNA-protein
composition
of top- and bottom-band capsids. Two series ofexperiments
weredone. In the first
experiment
DNA andprotein
composition were determined on an unlabeled preparation of A and B
capsids
obtained from cellsinfected with HSV-2.Following
their isola-tionasdescribedabove,thecapsids
werepelleted
byadditional
centrifugation.
Sequential
DNAand proteindeterminationsweredoneon eachpellet.
FIG. 1. Top and bottom bands in asucrose density
gradienit
obtained by centrifugation of aniuclear
lysatefrom HEp-2 cells
inzfected
withHSV-2. The lysatewaslayeredon top of a 10 to 40% (w/w) sucrose density
gradient prepared in 0.15M NaCIand0.01 .fsodium
phosphate (pH 7.2) and centrifuged at 23,000
rev/min
at4Cfor60miii ina
Spinico
S W27rotor.In the second experiment, approximately 4 X
108
cells were infected withhSV-1
and labeledwith 3H-amino acids
(1
,Ci/ml) and'4C-thymi-dine (0.4
,Ci/ml)
from 1 to 24 hrpcstinfection.
The nuclear lysate prepared from these cells wascentrifuged on sucrose density gradients. After
centrifugation, the top and bottom bands were
collected; an equal volume (50
Mliters)
of each was precipitated with cold,7%sY-
trichloroaceticacid, collected by vacuum filtration on 0.45-,m HA nitrocellulose discs, dried, and counted in a
PackardTri-Carb spectrometer.
Theresultssummarized in Table 1 indicate that Acapsids contain7.7to 9.7times less DNA than B capsids. Analyses of the size, structure, and
density of the B capsid DNA by rate velocity
sedimentation in neutral and
alkaline
sucrose density gradients, and by isopycniccentrifuga-tioninCsCl solutions, respectively, indicate that it has the same characteristics as that isolated
from mature virions reported previously from
ourlaboratory (11, 12, 15).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.293.433.82.359.2]HSV-SPECIFIED PROTEINS
I
/1
.is
FIG. 2. Electrontmicrographsof top (A andB)and bottom(CandD) band,HS V-2capsids. Wholemountswere
niegativelystained with2%7osodiumphosphotungsticacid containing 0.01% bovineserumalbumin(Aan2dC). Tkin-sectiontedpreparationiswerestained withuranylacetateandleadcitrate(BandD). Thecapsidsareapproximately
100 imn indiameter.
TABLE1. DNA and protein content ofA andBcapsids
DNA/protein of
Material analyzed DNA Protein DNA/protein DNA/protein of
Acapsids
Expt 1 (HSV-2)
Acapsids 9.27,ug 671
,/g
0.014 9.7Bcapsids 40.43JAg 297 ,Ag 0.136
Expt2 (HSV-1)
Acapsids 58 counts'mina 1,194counts,/minb 0.048 7.7
Bcapsids 282counts/min 766counts/min 0.368
a
14C-thymidine
was used to label HSV-DNA.bH-amino acid mixture was used tolabel HSV proteins.
V.,
Structural proteins of A and B capsids. The protein components of theAand Bcapsidswere identified on the basis of three szries of
experi-ments. The first series was a direct comparison
ofAand Bcapsid proteins and an estimation of theirmolarproportionality. For the comparison, 3 x
109
cells were infected with HSV-2 and labeled with '4C-amino acids from 4 to 20 hrVOL.10, 1972
.,a't:J,.0,. "-9, ..
;ILTAON'PLI";O.- .%,.
I
AlI.-z.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.49.430.82.423.2]postinfection. The nuclei were then isolated and
the extract was centrifuged in sucrose density gradients. The top and bottom bands were col-lected, and the capsids were solubilized and
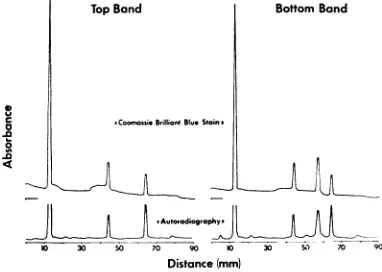
sub-jectedtoelectrophoresis in8.5% polyacrylamide gels. Absorbance profiles of the stained gels and
of their respective autoradiographic images are shown in Fig. 3. Estimations of the molar
pro-portionality of the capsid proteinswere based on
staining analyses of three different preparations
of B capsids, solubilized and separated
electro-phoretically in 8.5 % polyacrylamide gels as
described above.Asshownin Table 2,the molar
proportions of B capsid proteins 5, 19, 21, 22a, 23, and 24 are approximately 12:4:1:16:8:1,
respectively.
Thepurpose of the secondseries was to
estab-lish the identityof the A and B capsid proteins.
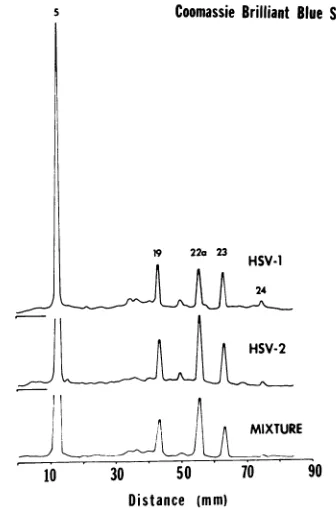
This was accomplished by subjecting HSV-1, Bcapsid proteinsto electrophoresis alone and in an artificial mixture with partially purified,
en-veloped HSV-1 virion proteins. Absorbance profiles of the stainedgelsareshown in Fig. 4.
Theobjectiveof the thirdseries was to compare the proteins of HSV-1 and HSV-2 intranuclear
capsids. Electropherograms of the HSV-1 and
HSV-2,Bcapsid proteinsare shown in Fig. 5.
The results of these analyses can be
sum-marized as follows. (i) B capsids contain four
major proteins and several mino detectable by autoradiography (F
band). On the basis of
co-electrn
virionproteins (Fig. 4), we have i
of the
major
proteins asbeingno. The fourth major protein is notvirion and has been designated a
a
.0
.,0
TopBand
*Corossi B;l11irntBlue Stain*
X 30 50 70 90 10 3
Distance(mm)
FIG. 3. Electropherograms ofthe p)
in the top (A capsids) andbottom (E
isolatedfrom HEp-2 cells infected u
labeled beginning 4 hr postinfkction
acids. Absorbance profiles of both
BrilliantBlue-stainedgelsandoftheir.
[image:5.493.263.456.82.195.2]imnagesareshow,z.
TABLE 2. Proteincompositionzof HSV-2 B Capsids
Protein Percent total Molecular Molar
no. protein in weightb ratio'4-SD
capsidaai SD (Xi10-3)
5 62.5 i 2.8 155 12.59 i 0.56
19 6.8 i 1.1 53 4.00 i 0.66
21 1.5 i 0.6 44 1.06 i 0.44
22a 20.5 i 2.8 38.8 16.50 4: 2.25
23 8.1 4 0.5 33 7.66 4 0.47
24 0.8 i 0.1 25 1.00 i 0.12
aAverage + standard deviation of four
de-terminations: two were made from gels stained with Coomassie Brilliant Blue stain and scanned
at 550nm, asdescribed in Materials and Methods;
the third was made from a gel fixed overnight in
25% isopropanol-10% acetic acid, stained with
Fast Greenstain (1% in water, pH 3.0) for 12hr,
destained overnight, and scanned at 620 nm; and
a fourth was made from one of the Coomassie
Brilliant Blue-stained gels scanned at 500 nm.
IDetermined as described in reference 25 on the
basis of migration in 6, 7, 8.5, and9%
polyacryl-amidegels byusing proteins of known molecular
weight as standards.
cCalculated by dividing the percent of the total
protein by the molecular weight and normalizing
with respect to the protein present in lowest
amount, no. 24.
ir components Based on its
electrophoretic mobility
in thezig.
3, bottom "mixture"gel
(Fig. 4),
we estimate thatprotein
ophoresis
with 22a hasamolecularweight
of38,800.
dentified three Onlytwo of the minor Bcapsid proteinsoccur
5, 19, and 23.
reproducibly
in the samerelative amounts from present in the preparation to preparation; one of these cor-ts protein 22a.responds
to virion protein 24 (Fig. 4). Thesecond has been tentatively identified as protein
21 because it iselectrophoretically indistinguish-Botom Band able from protein 21 and the amounts of protein in band 24 and putative band 21 are about the same, as shown both by staining (Fig. 4and 5) and autoradiography (Fig. 3). A similar
pro-portionalitywas found between viralproteins 21 and 24 inhighly purified preparations of virions
(reference 25, Fig. 6,
"4C-amino
acidautoradio-gram).
(ii)
Acapsids
lackprotein
22a and the minor component tentatively identified above as pro-tein 21, but are otherwise indistinguishablefrom Bcapsidsinprotein composition.
(iii)
Threeprincipal
lines of evidence indicatethat the
proteins
listed above are structuralroteins contained
components
of the A and Bcapsids
andarenotI capsids) banids
impurities.
Briefly,
proteins
5, 19, 23,
and 24 arevith HSV-2 and presentin
approximately
thesamemolarratios inwith '4C-amino A and B
capsids
and in virions(Fig.
3 and4)
the Coomassie asthose shown in Table 2 for B capsids. Secondly,
autoradiographic in one
experiment
the bottom-band material was layered on top of a discontinuousdensity
VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.62.253.470.606.2]HSV-SPECIFIED PROTEINS
CoomassieBrilliantBlue stain
10 30 50 70 90
[image:6.493.42.227.72.391.2]Distance (mm)
FIG. 4. Electropherograms of B capsid and virion
proteinsalonean1dinanartificial mixture. Three
com-panion gelswere subjectedtoelectrophoresis, onze
coiz-taimzingpartially purified virions (HSV-J virionzs),
an-other containingHSV-1 Bcapsidproteins, anda third
containingapproximately equalamounts of virionanld
B-capsidproteins(mixture). Absorbance profiles ofthe
CoomassieBrilliantBlue-stainedgelsareshown.
gradient, formed by layering equal volumes of CsClsolutions
prepared
in0.005 M sodium phos-phate (pH 7.1) with densities of 1.25, 1.35, and1.9 g/cm3, respectively. The gradient was then
centrifuged for 14 hrat30,000 rev/min and20 C in aSpincoSW41 rotor. Adiscretebandformed below the initial 1.25 to 1.35 g/cm3 interphase and contained all ofthe proteins in the relative
amountspresent in the sucrosedensity gradient. Thirdly, the electrophoretic mobility ofthe pro-teinsisindependent of the host, HEp-2 or Vero
cells,in whichthey are produced.
(iv) The electrophoretic mobilities of HSV-1 andHSV-2, Bcapsidproteins are thesame (Fig.
5).
Analysis of the capsids produced by stripping virons of theirenvelope. These experiments were
designed tocompare the protein composition of
capsids produced by stripping virions of their
envelope with that ofA and B
capsids
isolated
from nuclear lysates.
Approximately 4 x 108 HEp-2 cells were
in-fected with HSV-1 and labeled beginning 2 hr
after infectionwith
'4C-glucosamine.
Three typesofparticleswereisolated fromthecells 24 hrafter infection, as described above: partially purified
virions,
NP-40-extractedcytoplasmic
virions,and
NP-40/DOC-extracted cytoplasmic
virions.Briefly, the partially purified virions were
iso-lated fromhalf ofthe infected cells. The remain-ing cells were treated with 1 %c NP-40 in 0.15 M
NaCl, 0.01 M Tris
(pH
7.2), and 0.002 MMgCI2
for20minat 0 C. Thenuclei whichwere notlysed
by thisprocedure werepelleted bycentrifugation
at 1,500 x g for 10 min. Half of the clarified
NP-40lysate was notfurtherprocessed; the other halfwas treated
sequentially
with DOC(0.5%c),
deoxyribonuclease (50
ug/ml),
BRIJ-58(0.5%),
and urea (0.5 M) in exactly the same fashion as
nucleiwerelysed for the preparation ofA and B
capsids. The NP-40 and the NP-40/DOC-treated lysates were then banded by centrifuga-tion on sucrose density gradients as described
Coomassie Brilliant Blue Stain
9
223HSV-1
_Oj
MIXTURE
10 30 50 70 90
Distance (mm)
FIG. 5. Electropherograms ofHS
V-J
andHS V-2 Bcapsidproteins. Companiongels
con2taining,
respectively,HSV-J,
B capsidproteins (HSV-i);
HSV-2, B capsidproteins (HS V-2); and a mixture
of
approximatelyequalamountsof
HSV-i
and HSV-2, Bcapsid proteins(mixture) weresubjectedtoelectrophoresis.Shownhere
areabsorbaiiceprofiles oftheCoomassie Brilliant Blue-stainedgels.
VOL. 10,1972
on November 10, 2019 by guest
http://jvi.asm.org/
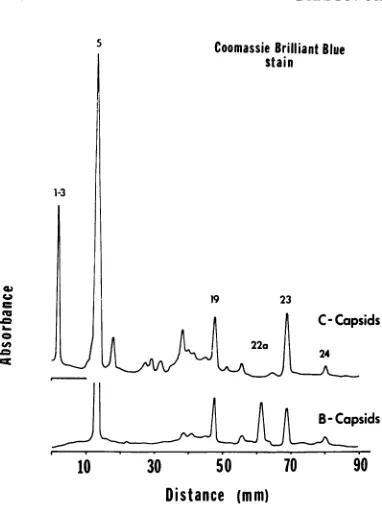
[image:6.493.263.431.342.597.2]1-3
Coomassie BrilliantBlue
stain
19 23
C-Capsids
II ~~~~~~~~22a
B-Capsids
10 30 50 70 90
[image:7.493.62.253.67.321.2]Distance (mm)
FIG. 6. Electropherograms ofB andCcapsids
iso-latedfrom HSV-J-infected HEp-2 cells. Absorbance
profiles oftheCoomassieBrilliant Blue-stainedgelsare
shown.
above. One band formedineachgradient andwas
collected and storedat -20C.
Analyses of these three types of particles
re-vealed the following. (i) NP-40 treatment alone reduced the specific activity (counts per minute
of"C-glucosamine permicrogram of protein) of the NP-40particleby 90% ascompared with the partially purified virions. The electrophoretic profile of the particles obtained after treatment ofthecytoplasmwith NP-40alonewasessentially thesame as that ofparticles produced by NP-40
treatment of highly purified virions previously
described (25). (ii) NP-40treatment followed by DOC extraction further reduced the specific activitytolessthan 2% thatofthevirion. Fig.6 shows the electrophoretic profile of the proteins
presentintheseparticles, designatedasCcapsids.
The data are noteworthy from two points of view. First, the C capsids contain all of the B capsid proteins except protein 22a. Secondly, they contain several proteins presentinthevirion (proteins1to 3) absentfrom the A and Bcapsids
and purified membranes. They lack, however, thenonglycosylated proteins 4 and22,aswellas
allof theglycoproteinspresent inpurifiedvirions and in the purified membranes. We should add that the electropherograms of C capsid proteins exhibited a number ofless well-defined and less consistently reproduciblebands betweenproteins
5and 19. Parenthetically, increasing the detergent concentrations from 0.5% to 1 % had little noticeable effect on the protein composition of the C capsid.
DISCUSSION
Inthis paper we are reporting the isolation and composition of three classes of HSV capsids. The salient features of the data, particularly as they relatetothe structure of the virion and its assembly, may be summarized asfollows.
(i) Nucleocapsids class A and B are derived from the nucleus of the infected cells. As discussed below, they had not been enveloped at the time of
isolation. The A and B capsids differ in four respects. First, they differ in hydrodynamic properties in that A capsids sediment more slowly than Bcapsids. Second, they differ
morphologic-allyinthat Acapsids lack thecoreassociated with
viral DNA (reference 8, Fig. 2B, D). Third, as would be predicted from the electron micro-graphs, they differ in DNA content. B capsid
contains 10 times more DNA than A capsids;
this DNA is hydrodynamically and structurally
thesame asthatisolatedfrom virions and previ-ously described (11, 12, 15). Lastly, A and B
capsids differ in protein composition. Although
A andBcapsids ofHSV-1 cannot bedifferentiated from their HSV-2 counterparts, with respect to the proteins present or their electrophoretic
mobilities,the Bcapsids ofboth subtypes have a
major protein (no. 22a) not present in the A
capsids. It is pertinent that protein 22a is not a
contaminant of the B capsid but rather a
tena-ciously bound, structural component of that
particle. This conclusion is basedon twofindings. First, itisassociated inthe sucrosedensity
gradi-entsonlywiththe bottom band; it isabsent both from the gradient fractions above and below the
bottom band. Moreover, it sedimentsthrough a 2Murea-20% sucrose cushion and bands in CsCl
solutions inassociation with the B capsid.Second,
protein 22a is synthesized and becomes a struc-tural component of B capsids in both HEp-2 and Vero cellslong afterhostsynthesisisshutoff.
(ii) The C capsids have been derived by extrac-tion of virions with the same reagents as those used to lyse the nuclei ofinfected cells. As evi-denced bya98% decrease in thespecificactivity
of
"IC-glucosamine,
this procedure removesessentiallyallof theglycoproteins associatedwith the virions. The C capsids differ substantially
fromboth A and B capsidswith respect to pro-tein composition. This, basically, is the datum which led us to conclude that A and B
capsids
do not represent degradation products of
en-veloped cytoplasmic virions.The most
interesting
featureofthe Ccapsid is its retention ofspecific nonglycosylatedproteins throughoutthe
rigorous
5VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
detergent-isolation procedure. Thus, whereas proteins 4 and 22 were completely removed, protein band 1 to 3 was nearly quantitatively
retained, None of these are found as structural
components of A or B capsids. Preliminary
studies based on pulse-chase experiments and some propertiesof the proteins suggest that
pro-tein 22a isa precursor toprotein 22. The nature
and function of proteins 1 to 3 are not certain.
As previously reported (N. Frenkel and B.
Roizman,Proc. Nat. Acad. Sci. U.S.A., inpress;
Proc.25th Annu.Symp.Fund. CancerRes. M. D. Anderson Hosp. Tumor Inst., in press), these proteins are synthesizedlaterthan theremaining proteins ofthevirion andcorrespond, in time of synthesis andamountofDNAtemplate required
for their synthesis, to species ofribonucleic acid
transcribed from thesame amount of DNA late
ininfection.
One hypothesis, which reconciles the data re-ported in thispaper, thetranscriptional program
of the virus (N. Frenkel and B. Roizman, Proc.
Nat. Acad. Sci. U.S.A., in press; Proc. 25th
Annu.Symp.Fund. CancerRes. M. D. Anderson
Hosp. TumorInst., in press),the time of
synthe-sis of viral proteins (Gibson and Roizman,
manuscript in preparation), and the observation
that in HSV-infected cells empty capsidsdo not,
as a rule, become enveloped (22), is as follows. Protein 22a is found in association only with capsids containing DNA; B capsids, containing thisprotein,arethen able to bindproteins 1 to 3
and subsequently become enveloped. In the processprotein 22a ismodified, perhaps cleaved,
such that it is subsequently removable fromthe
enveloped particles by detergent treatment. Ac-cording to this scheme, protein 22a is on or ex-posed to the surface of the capsid and, as previ-ouslysuggested (N.Frenkel andB. Roizman,see
in press references cited above), proteins 1 to 3
regulateand enableenvelopment.
(iii) We do not know at present whether A capsids are by-products or the precursors to B capsids. Themajor differences between A and B capsidsarethepresenceinBcapsids ofa"core,"
more DNA, and two additional proteins, no.
21 and 22a. If protein 22a, present in higher
abundance than protein 21,is indeed onthe
sur-face ofthe capsid, as we suspected based on its
disappearance from virions, it wouldfollow that the only protein which conceivably might be
associated with the DNA core of the capsid is
protein 21.
(iv) Thedatapresented inthispaperpermit us
to order the virion proteins reported in
preced-ing papers of this series into four groups as shown inTable 3. They are as follows: group 1
r.
.c
r.
._
- +
± +
C14~~~~~~~~~~
C1 + +
n-~++++ev
+ ++
+
+ -H+
-+ 4i
+~-n + +
<14-+ -H++
+ +-+
+
x + +
t + '.+
1 +
-I
-< -+
Qo o
ua._
i o Q
_E _
-0
-Y . *t
$L
Q
ECA e
Cono
u E ~'o
00v~~
U05,.<
a, o'
X
NO,X
Y 0sr
-ej - 0% -) e
VC.) C
°°C X~
-_ t_3
D,, OU :)
_s .o
gO
Wnn =:/: -N
~
1051
H +
! lll
u
In
2
w
3-1 Cd
14
.z
0
.-d
.E
V)
on November 10, 2019 by guest
http://jvi.asm.org/
comprises proteins associated with membranes, i.e., proteins 7, 8, and 11 through 18 (13, 25;
T. Savage, B. Roizman, and J. W. Neine, J.
Gen. Virol., inpress). We areledto suspect that bandno. 19 containstwoproteins of whichoneis glycosylated and associated with membranes whereas the other isnot glycosylated and associ-ated with capsids. As indicassoci-ated in the table, membranes isolated from infected cells contain
traceamountsof proteins 22, 23, and 24. Itisnot
presentlyclearwhether thesearespecific structural
components or adventitious contaminants of
purified membrane preparations. Group 2 com-prises proteins present in A and B capsids, i.e., proteins5, 19, 21, 22a, 23, and 24. Proteinno. 5
accounts for more than 60%' (w/w) of the pro-teinconstituents of thecapsids (Table 2). Group 3comprises proteins 1 to4 and protein22which
we suspect bind to the capsids and intercalate
between the capsid and the envelope. Finally, group4comprises several minorbutreproducible
componentsof the virion whichwehavenotbeen
abletoplace.Theseareproteins 6, 9, 10,and 20.
ACKNOWLEDG MENTS
These studieswereaidedbyU.S. Public Health Service grant CA 08494 fromthe National CancerInstitute,grantNP15Gfrom theAmerican CancerSociety, and grantGB 27356X from the National Science Foundation. W. G. is a U.S. Public Health Service predoctoraltrainee(Al 00238 from the Nattional Institute of Allergy andInfectious Diseases).
We thank J. Heine,D.Furlong,and P.Wicdncrfor assistance with the electronmicroscopy.
LITERATURECITI'ED
1. Abodeely,R.A.,E.Palmer,L. A.Lawson,anidC.C.Randall.
1971.Theproteinsofenveloped and de-enveloped equine abortion(herpes)virus and theseparateenvelope. Virology. 44:146-152.
2. Burton, K. 1956. Astudyoftheconditions andmechanism of thediphenylaminereaction for thecolorimetric estima-tion ofdeoxyribonucleicacid. Biochem. J.62:315-323. 3. Darlington, R.W., and L. H. Moss. 1968. Herpesvirus
en-velopment.J. Virol. 2:48-55.
4. Dimmock,N.J.,and D. H.Watson. 1969.Proteinsspecified byinfluenza virusininfectedcells:analysis by polyacryla-midegelelectrophoresisofantigensniotpresent in thevirus particles.J. Gen. Virol. 5:499-509.
5. Dische, Z. 1930.Ubereinigeneuecharaikteristische Farbreak-tionen derThymonukleinsaureundcine Mikromethodezur
bestimmung derselbenintierischenOrganmit hilfedieser Reaktionen. Mikrochemie. 8:4-32.
6. Eggstein, M.,andF. H. Kreutz. 1955.Vcrgleichende
Unter-suchungenzurquantitativen EiweissbestimllmliungimLiquor
und eiweissarmen Losungun. Klin. Wochenschr. 33:879-884.
7. Ejercito, P. M., E. D. Kieff, and B.Roizman. 1968. Charac-terization of herpes simplex virusstrains differing in their effect on social behavior of infected cells. J. Gen. Virol. 3 :357-364.
8.Epstein, M. A. 1962. Observations on the fine structure of
matureherpes simplex virus andonthe composition ofits nucleoid. J. Exp. Med. 115:1-12.
9. Fairbanks, G., Jr., C. Levinthal, and R. H. Reeder. 1965. Analysis of '1C-labeled proteins by disc electrophoresis. Biochem. Biophys. Res. Commun. 20:393-399.
10. Fairbanks, G., T. L. Steck, and D. F. H. Wallach. 1971. Electrophoretic analysis of the major polypeptides ofthe human erythrocyte membrane. Biochemistry 10:2606-2617. 11. Frenkel, N., and B. Roizman. 1971. Herpes simplex virus:
genome size and redundancy studied by renaturation kinetics. J. Virol. 8:591-593.
12. Frenkel, N., and B. Roizman. 1972. Separation of the herpes-virus deoxyribonucleic acid duplex into unique fragments and intact strandonsedimentationin alkalinegradients. J. Virol. 10:565-572.
13. Heine, J. W., P. G. Spear, andB.Roizman. 1972. Proteins specified by herpes simplex virus.VI.Viralproteins in the plasma membrane. J. Virol.9:431-439.
14. Kaplan, A. S., and T. Ben-Porat. 1970. Synthesis of proteins incells infected with herpesvirus. VI. Characterization of the proteins of the viral membrane. Proc. Nat. Acad. Sci. U.S.A. 66:799-806.
15. Kieff, E. D., S. L. Bachenheimer,and B. Roizman. 1971. Size, composition,andstructureof thedeoxyribonucleicacid of herpes simplexvirussubtypes1and 2.J. Virol. 8:125-132.
16. Laemmli,U. K. 1970.Cleavage of structural proteins during theassemblyof the head ofbacteriophageT4.Nature (Lon-don)227:680-684.
17. Lowry,0.H., N. J. Rosebrough, A. L. Farr, andR.J. Ran-dall. 1951. Proteinmeasurementwiththe Folinphenol
re-agent.J.Biol. Chem. 193:265-275.
18. Morgan, C., H. M. Rose, M. Holden, and E. P. Jones. 1959. Electronmicroscopic observations onthe development of herpes simplex virus. J. Exp. Med. 110:643-656.
19. Nii, S., H. S. Rosenkranz, C. Morgan, and H.M.Rose.1968. Electronmicroscopyofherpes simplexvirus. II[.Effect of hydroxyurea.J.Virol.2:1163-1171.
20. Nii, S.,C.Morgan,H. M.Rose, andK.C. Hsu. 1968.
Elec-tronmicroscopyofherpes simplexvirus. IV. Studies with ferritin-conjugatedantibodies. J.Virol. 2:1172-1184. 21. Olshevsky, U., and Y. Becker 1970. Herpes simplex virus
structuralproteins. Virology40:948-960.
22. Roizman, B. 1969. Theherpesviruses-a biochemical defini-tion of thegroup.Curr.Top.Microbiol. Immunol. 49:1-79 (Heidelberg).
23. Roizman, B., S. B. Spring,andJ. Schwartz. 1969. The herpes-virion and its precursors made in productively and in abortivelyinfected cells. Fed. Proc. 28:1890-1898. 24. Schwartz, J., andB. Roizman. 1969.Similarities and
differ-encesinthedevelopmentoflaboratorystrains andfreshly isolated strains of herpes simplex virus in HEp-2 cells: electronmicroscopy.J. Virol. 4:879-889.
25. Spear, P. G. and B. Roizman. 1971. Proteins specified by-herpes simplex virus. V. Purification and structural
pro-teins of theherpesvirion.J. Virol. 9:143-159.