JOURNALOFVIROLOGY, June, 1974,p. 1254-1262 Copyright0 1974 AmericanSocietyforMicrobiology
Vol.13,No. 6 Printed inU.S.A.
Base
Composition
and
Hybridization
Studies of the
Three
Double-Stranded RNA
Segments
of
Bacteriophage
061
J. L. VAN ETTEN, A. K. VIDAVER, R. K. KOSKI, ANDJ. P. BURNETT
Department ofPlantPathology, UniversityofNebraska, Lincoln,Nebraska68503, andLillyResearch
Laboratories, Indianapolis, Indiana 46206
Received forpublication3January 1974
The three double-stranded ribonucleic acid
(dsRNA)
segmentsof thebacterio-phage
46 were isolated and shown to have similar melting temperatures andbase compositions. RNA: RNA
hybridization experiments
with the isolatedsegments eliminate the
possibility
that thetwo smaller dsRNAsegments arisefrom a
cleavage
of thelarge
dsRNA segment. The two smaller RNA segments reannealrapidly
even at low temperatures; in contrast, thelarge
dsRNA reannealedonly
athigher
temperatures. Evidence is also presented whichsuggests that the dsRNAsmay contain ashort
single-stranded
RNAtail.Several
plant
and animal viruses contain adouble-stranded
ribonucleic acid(dsRNA)
ge-nome.
In mostcases,
thesedsRNAs have
aguanosine
plus cytosine
(G + C)
contentof 38 to 44%, and each virion contains several species of dsRNA with molecularweights ranging
from 0.2 x 106 to 2.9 x 106(11).
Previously,
wereported
the
isolation
andgeneral characteristics
of amorphologically unique
lipid-containing
bacte-riophage, 46,
ofPseudomonas
phaseolicola (10).
The phage genome was
composed
of at least three dsRNA segments withestimated
molecu-lar
weights of
2.2 x106,
2.8 x106, and 4.5
x 106(8). The
G+
C content of thetotal
dsRNA
was 56 to58%,
which ishigher
thanthatreported
forother dsRNA viruses except
forthe mycovirus
from Penicillium chrysogenum (7). The present
report
describes additional characteristics of
the
06
dsRNA segments.
MATERIALS AND METHODS
Virus culture, purification, and nucleic acid
extraction. The
host,
strain HB1OY of P. phaseoli-cola,wasgrowninasemisyntheticmedium(SSM)aspreviously reported (10). The phage was isolated from
7- or 25-liter lysates and was purified by CsCl
equilibrium sedimentation (10). The dsRNA was
isolatedfrom thepurified phage by the single-phase
phenol procedure of Diener and Schneider (4) as
described previously (8). Phage 06 32P-dsRNA was
prepared by adding 50 mCi ofH332PO4atthe start of a
7-liter fermentation in SSM modified by a 10-fold
reductioninthe phosphate concentration.
Centrifugationprocedures. Sedimentation coeffi-cients were estimated in linear-log sucrose density
gradients (2) equilibrated with0.3 M NaCl and 0.03
'Journal paper no. 3729. Nebraska Agricultural Experi-mentStation.
Msodiumcitrate, pH 7.0(2xSSC),andcentrifuged
in aSpinco SW41 rotor at 37,000 rpm at 14 C for 8to
12h. Thegradient columnswerescanned
photometri-cally at absorbancy at 254 nm
(A214)
withan ISCOdensity gradient fractionator (Instrumentation
Specialities Co.). Brome mosaic virus RNAs were
used as standards for determining sedimentation
coefficients.
Polyacrylamide gelelectrophoresis. Gel
electro-phoresiswas performed in2.4 or3% polyacrylamide
containing 0.5% agarose in 0.04 M Tris, 0.02 M
sodium acetate,0.001MsodiumEDTA,0.2%sodium
lauryl sulfatebuffer,pH7.2(1). Thegels(9cmlong,6
mm diameter) were pre-electrophoresed for4hat 5
mA/gel priortotheaddition of0.05- to0.1-mlsamples
in 6%sucrose.Thesampleswereelectrophoresedat5
mA/gel for4to8 h (analytical) orfor 16 h
(prepara-tive)at roomtemperature. Thegelswerescannedat
A260 with a Gilford model 2410 linear transport
system(GilfordInstruments), whichwascoupledtoa
Beckman DU spectrophotometer. When radioactive
profileswererequired, thegelswerefrozenwith solid
CO, andsliced into 1-mmsections. The radioactive
material waseluted from the gel slices (12) and the
samples werecountedasdescribed previously (9).
Strand separation and reannealing of the
dsRNA. Severalprocedureswereusedtoseparate the
06dsRNA intosinglestrands. These included heating
thedsRNAto 100Cfor 90 s in0.01MEDTA, pH 7.0,
followedby quickly coolingto -8C. In some
experi-mentssampleswereheatedin 3.7% (vol/vol) formal-dehyde and dialyzed overnight against 3.7% formalde-hyde in 0.001 M EDTA, pH 6.8, at 40 C (3). In
addition, separationofthedsRNA into single strands
was attempted in 50% formamide or 85%
dimethyl-sulfoxide (6) by heating at various temperatures and
for variousperiodsoftime.
Todetermine therateofreannealing of the dsRNA
segments, the sampleswereheated in0:01MEDTA,
quickly cooled, and then incubatedatvarious
temper-atures. Portionswereremoved at appropriate times,
1254
on November 10, 2019 by guest
http://jvi.asm.org/
DOUBLE-STRANDED RNA FROM PHAGE k6
quickly adjusted to 2 x SSC, and incubated with
RNase A (10
Ag/ml)
at 22 C for 15 min beforeelectrophoresing on polyacrylamide gels. The
percent-age ofreannealing was determined from planimeter
measurements of the
A,.0
profiles of thedsRNA andcompared to equivalent samples which had not been heat treated.
Isolationof the dsRNA segments. After
electro-phoresis of total06dsRNA on 2.4 or 3%
polyacrylam-ide gels for 16 h, the gels were scanned and the three
dsRNA regions were excised, frozen at -70 C, and then passed through a fine mesh screen. The dsRNA
was eluted from themeshed material by shaking in a
small volume of 2x SSC for 2 days at room
tempera-ture. After centrifugation to remove the residual
polyacrylamide, the RNA was precipitated with 2 volumes of ethanol. The precipitated RNA was dis-solved in 2x SSC, centrifuged on sucrose density gradients, and RNA regions were collected and dialyzed extensively against 0.01 x SSC containing
0.01 M EDTA, pH 7.0, followed by dialysis against
0.01x SSC, pH 7.0. RNA segments isolated in this
manner wereusedtoobtain themelting temperatures
(Tm) and base composition of each segment. The 06
dsRNA segments used for the hybridization
experi-ments wereisolatedby returning the excised
dsRNA-containinggelstothegel tubes and by
electrophores-ingthe dsRNA into dialysis sacks. The dsRNAswere
dialyzed and concentrated by ethanol precipitation.
Base composition analysis. RNA samples were
hydrolyzedbytreatmentwith0.3MKOH for 16 h at
37C. After neutralization with perchloric acid and
removal ofinsoluble potassium perchlorate, the pH
wasadjusted to8.8withNH4OH. Afterthe addition
of 0.05 M ammonium acetate (pH 8.8), alkaline
phosphatase was added and the sample was
incu-bated 3h at 37Cto removethephosphate residues
from the nucleotide mixture. The resulting
nucleo-sides were separated on a Bio-Rad A-26 column by
using ammonium formate buffer (0.4M, pH 4.7). The
eluate was monitored at
A2.4
and the quantity ofeach nucleosidewasdetermined from theareaofthe
respectivepeak. All analyseswereperformed in
tripli-cate.
Thermaldenaturation. Thermal transition
stud-ies were performed in a Cary 16 spectrophotometer
equippedwith anelectricallyheated5-position
auto-matic sample changer block. Cuvet temperatures
were monitored by athermistor inserted in a blank
cuvet containing buffer. Opticaldensities and
tem-peratures for eachsamplewererecordedcontinuously
withaheatingrateof 0.3to 0.6C/min.The sodium
ion concentration of all samples was adjusted by
dialysis againsttheappropriatedilution ofSSC prior
tomelting.
RNA: RNA hybridization experiments.
Dupli-cate samples, containing 0.05
A,..
of eachS2P-dsRNA (approximately 70,000 counts per min per
A
..0)
segment, were mixed with 0.5A260
ofun-fractionated, unlabeled
06
dsRNA and adjusted to0.01MEDTA, pH7.0. After a heat treatmentat100
C for 90 s,onesetofsampleswas incubatedat50C
for 60 min to allow reannealing. The samples were
then adjusted to 2x SSC and treated with 10
sg
ofRNase Aper mlat 22C for15min priorto
electro-phoresingfor15hon 3%polyacrylamidegels. Thegels
were scanned at
A260,
sliced, and counted. To provethat strand separation occurred during the original
heat treatment, the othersetofheat-treated samples
was immediately diluted fivefold with ice water,
centrifuged on linear-log sucrose density gradients,
scanned.and fractions werecollected and counted.
ssRNA tail structure. To determine if the
06
dsRNA containedsingle-stranded regions, 32P-dsRNA
(approximately 105 counts per min per A260) in 2x
SSC was incubated with RNase A (10
gg/ml)
andRNase T, (4,000 units/ml) at 22 Cfor 15min before
centrifuging on linear-log sucrose density gradient
columns. The amount of radioactivity that
sedi-mentedinthe top20%ofthegradientwasdetermined
after fractionation of the gradient columns. These
values were compared with untreated
06
dsRNAsamples. In addition, the dsRNA regions in sucrose
density gradient columns from RNase-treated
sam-ples were dialyzed, precipitated with ethanol,
sub-jectedtoasecond RNase treatment, andcentrifuged.
RESULTS
Strand
separation
of the dsRNA.
One
un-usual feature
of X6dsRNA noted previously
wasthe
disproportionate quantities
ofthe
threesingle-stranded
(ss)RNA
specieswhich
ap-peared after heating the dsRNA
to 100C
for 90 s in 0.01M EDTA, pH
7.0,quickly cooling
to -8C, and electrophoresing
onpolyacrylamide
gels
(8;Fig. 1). Almost all
ofthe
large dsRNA
segmentexisted
asssRNA, whereas only
asmall amountof the
twosmaller dsRNA
segmentselectrophoresed
asssRNA
species after this treatment.Variations
inthe
length
ofthe heat
treatmentand decreased
concentrations ofEDTA did
notalter this
pattem exceptthat
prolonged heating increased the amount
ofpolydisperse
low
molecular
weight RNA.
At-tempts toseparate
the
dsRNAs
intossRNAs
in50% formamide
or 85%dimethyl
sulfoxide
fol-lowed by electrophoresis
onpolyacrylamide gels
also
resulted
inprofiles similar
tothose shown
inFig.
1. Twopossible explanations
forthis
phenomenon
are(i)
the
large dsRNA
segmentcontained
alower G
+C
contentthan the
twosmaller
segmentsand thus
wasthe
only
seg-mentthat
separated
intosingle strands
or(ii)
all three
segments wereseparated
intosingle
strands but the
twosmaller
segmentsrean-nealed much faster than the
large
segment.The
following experiments indicate that (ii)
isprob-ably
theexplanation
for thisphenomenon.
(i) Thesharp
40% increase inthehyperchromicity
ofthe/6
dsRNA described
previously (8)
sug-gested
that all three segments hadseparated
into
single
strandsatthe sametemperature.(ii)
If thedsRNA
wasexposed
to 100C
for 90 sin 0.01 MEDTA,
andimmediately
incubated with RNase A in 2xSSC,
approximately
90%ofthe
RNA wasdegraded (0
timepoint
inFig. 2).
1255VOL.13,1974
on November 10, 2019 by guest
http://jvi.asm.org/
VAN
ETTEN
ET AL.E
c
0
~ii
J L
A B
I
l
RELATIVE MIGRATION
-4
FIG. 1. Electrophoresis of06RNAon2.4%polyacrylamide gelsfor4hat5mA/gel.(A) Untreated 46 RNA
showing the three dsRNA segments.(B)Phage
06RNA
whichwasheatedfor90sat100Cin 0.01MEDTA,
pH7.0,andquicklycooledto-8Cpriortoelectrophoresis. Thearrowsindicate the three ssRNAs
arising from
the three dsRNAs.Thus all three dsRNAs
must haveseparated
intosingle strands with the heat
treatment.(iii)
Ifthe melted RNAs
wereincubated
at24C,
the
small dsRNA
segmentreannealed
rapidly,
fol-lowed by the medium
sizedsRNA; essentially
noreannealing occurred with the
large dsRNA
at 24C (Fig. 2).
In contrast,all three dsRNAs
reannealed
rapidly
at 50C
(Fig. 2). Separate
experiments established that the
optimal
tem-peraturefor
reannealing
in 0.01 MEDTA
wasbetween
50and
60C.
(iv)
Ifthe dsRNA
washeated
to 100C
for 90 s in 0.01 MEDTA,
cooled
rapidly, and
immediately diluted fivefold with
ice waterbefore
centrifuging
on sucrosedensity
gradients, three
fast-sedimenting species
ofssRNA
wereobserved
which hadS values
of 24.7, 29.4, and 33.0 (Fig. 3). TheA2,4
peaks
for the threessRNA
species were in about the ratioexpected
ifallthreedsRNA
segments hadseparated
intosingle
strands.
(v)
Determina-tions of the Tm values and thebase
composi-tions ofthe three isolated dsRNA segments werenearly
identical(Table
2).Exposure
ofthe dsRNA to 100 C for 90 s in the presence of 0.01 M EDTA containing 3.7%formaldehyde,
followedby overnight dialysis
against
3.7%formaldehyde
at 40C,
resulted inthe
appearance of three ssRNA species whichsedimented
at 17.8, 20.1, and23.5S
(Fig. 3C).By using these S values and the
formula S
=0.083 M038 for
converting
sedimentationcoeffi-cients
offormaldehyde-treated ssRNA
to molec-ularweights (3),
one obtains values of 1.36 x106,
1.88 x106,
and 2.84 x 106 for the threessRNA
components.These values give
molecu-lar weights
of 2.72 x106,
3.76 x106, and
5.68 x 106for
the three dsRNAs and
atotal molecular
weight
of 12.16 x 106for the
phage
genome(Table 1). These values
areapproximately
15 to 30%larger than those calculated directly
fromthe
sedimentation coefficients
ofthe dsRNA
using
the
formula
M =2.24S5
16(ref. 5) orthosederived
previously from polyacrylamide gel
electrophoresis and by electron microscopy
(8; Table 1).Characteristics of
theisolated dsRNA
segments. The three dsRNA segments ofk6,
isolated
afterseparationby
polyacrylamide
gelelectrophoresis,
were free of cross-contamina-tion (Fig. 4). TheTm
values and the base com-positions werevirtually identical
for each ofthe
three separated segments (Table 2).Since
the sum of themolecular
weights
ofthe twosmaller
dsRNA segments areapproxi-mately
equal to the molecular weight of thelarge
dsRNA segment, it is possible that thej
1256 J.VIROL.
L")
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.115.402.63.314.2]DOUBLE-STRANDEDRNAFROM PHAGE
06
These
data eliminate the possibility that the twosmaller dsRNA segments arise from
a
cleavage
ofthe large
dsRNAsegment.a w < 60.
w z z w I-I z w 040
w a.
0 20 40 60
[image:4.495.50.243.51.381.2]TIME, MIN.
FIG. 2. The rate of reannealing of the three X6
dsRNAsegmentsat24and50C. Theprotocol forthe
experiment is described inMaterials andMethods.
The dashed lines andopensymbols wereobtainedat
24C and the solid lines and solidsymbols at50 C.
The symbols 0, *, and A represent the large,
medium, and small dsRNA segments, respectively.
smaller
segments arise from aspecific
cleavage
ofthe
large
dsRNA segment. Ifsuch
acleavage
occurred
wewould have
expected
morethan
the sixRNA
peaks observed
inFig.
1because
large
ssRNA would have reannealed with small
andmedium
ssRNA; such results have
neverbeen
observed. Additional evidence that
the twosmaller dsRNA
segmentswere notderived from
the
large dsRNA
segment ispresented
inFig.
5.Isolated
46
3P-dsRNA
segmentsallowed
toreanneal with
a10-fold
excess ofunlabeled,
unfractionated
06
dsRNA revealed that each segmentreannealed
only
with strands of the same size(Fig. 5A,
C,
andE). Figures 5B, D,
and F show that strandseparation
ofboth the radioactive andunlabeled
06
dsRNAactually
occurredduring
theoriginal
heat treatment; thus all threeunlabeled
strandshad theoppor-tunity
toreanneal witheach
32P-RNA
segment.E
CM
I' w Hr
B
RELATIVE DEPTH
FIG. 3. Rate zonal sedimentation of
06
dsRNA inlinear-log sucrose density gradients. (A) Untreated
dsRNA; (B) dsRNA heatedto 100Cfor90 sin0.01M
EDTA andquickly dilutedfivefold with icewater;(C)
dsRNA heated as in (B) butin the presenceof3.7%
(vol/vol)formaldehydeandthendialyzed for16h at
40C in the presence of 3.7% formaldehyde. The
sedimentation coefficients are indicated in the
fig-ures.
VOL.13,1974 1257
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.260.449.112.564.2]TABLE 1. Acomparisonof the molecular weightsof phage k6 dsRNAsasdetermined by four procedures
Molecularweightx106
Segment Electron
Polvacrvlamide
gel Formaldehyde- CalculatedfromtreatedssRNAY the S value Avg microscopy' electrophoresisab treated ssRNAc of thedsRNAd
Small ... 2.0 2.24 2.72 2.20 2.29
Medium ... 2.87 2.82 3.76 3.11 3.14
Large ... 4.78 4.52 5.68 5.00 5.00
Total ... 9.65 9.58 12.16 10.31 10.4
aValuereportedinreference8.
bDeterminedfrom melted
single-stranded
46
RNA(8).cDetermined from sedimentation
coefficients
of single-stranded06
RNA obtained afterformaldehyde
treatment.
dDetermined directlyfrom sedimentation coefficients of14.5, 15.5, and17.0ofthedsRNA(8)
by
usingtheformula ofM = 2.24S516(ref. 5).
TABLE 2. TheTm's of the three phage k6 dsRNA segments atseveral salt concentrations and the nucleoside
composition of these segments
T,, values(C) Basecompositiona
Segment
sodium
saltconcn0.0017 0.0099 0.0463 U G A C A/U G/C
Small ...70 81.7 93.0 23.0 28.7 20.9 27.4 0.91 1.05
Medium ... 70.3 82.9 94.0 21.6 30.0 20.2 28.3 0.94 1.06
Large... 70.2 82.2 94.0 22.9 29.2 20.4 27.6 0.89 1.06
Total... .70.0 83.0 94.2 22.1 28.9 20.4 28.6 0.92 1.01
aStandard deviationwas ±0.36calculatedontheassumptionthat the varianceforeachbase determination
was identical.
Evidence to suggest a ssRNA region in the
dsRNA. Previously,
wereported that phage
06
contained
a RNApolymerase
which mediatedthe
incorporation of ribonucleosidetriphos-phates
into dsRNA and we suggested that the enzymemight be completing
one ofthe
comple-mentarystrands
of thedsRNA
(9). If this suggestion was correct,06
dsRNA
might con-tain asmall
ssRNA"tail"
region. Phage06
32P-dsRNA
wasincubated with RNase
A andRNase
T1
in 2xSSC
prior tolayering
onlinear
log
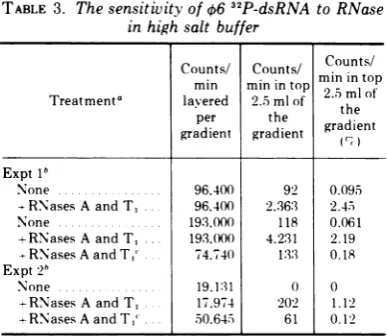
sucrosedensity gradients. As noted in Table 3, 1 to2.5%
ofthe
RNA layered on the gradient wasdegraded
so that it sedimented in the top 20% of the gradient. Less than0.1%
of the radioactivity in the nonenzyme-treated control was observed in this region of the gradient. If the dsRNA region from sucrose density gradi-ents ofRNase-treated
samples was collected, reprecipitated with ethanol, RNase treated a secondtime, and centrifuged on sucrose gradi-entcolumns,
less than 0.2% of the radioactivity layered on thegradient was degraded to slowlysedimenting
material. No obvious differences in the sucrosegradient
A254
profiles of any of thesamples
were observed.Therefore,
these dataare consistent
with
the
conceptthat
06
dsRNA
contains ashort ssRNA
"tail" region.
DISCUSSION
The
results
provide
additional
informationabout the three
segmentsof the
phage
06
dsRNA
genome.Previously,
we wereunable
toexplain the
disproportionate quantities
of
ssRNA
that
occurred
afterheating
the
dsRNA
to 100C
prior
toelectrophoresing
the RNA
onpolyacrylamide gels
(8). From the
experiments
presented,
it isclear that the
twosmaller RNA
segmentsreanneal much
morerapidly
atlow
temperaturesthan
thelarge
RNA
segment. Thisdisparity
inthe rate ofreannealing
occurs eventhough all
three RNA segments have similarbase compositions.
Therapid
reanneal-ing of the two smaller dsRNA segments also occurred after treatment with 85%dimethyl
sulfoxide
and 50% formamide sinceA2,,0
pro-files ofpolyacrylamide gels
ofdsRNA denatured
with these agents were similar to thosereported
inFig.
1.However,
three ssRNA segments wereobserved
ifthe
samples
werediluted fivefold
and
subjected
to sucrosedensity gradient
cen-trifugation.
1258 VAN ETYTEN ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
DOUBLE-STRANDEDRNA FROM PHAGE 06
1259
C
0
tion. ~ ~ c
w
TI;~REAIEMIRTO
[image:6.495.106.395.66.574.2]FI.
letrphreis-
fioae,6dRAsget
n24 oyarlmd esfr6ha Agl(,BFIG. 4.eEethropoessoisolatedcomdRNAsegentsolre
;mdu,
ndsal2.4 placryaiegsenilsy
fore6ea5f mA/gelonam(a,B
tion.
VOL.13,1974
on November 10, 2019 by guest
http://jvi.asm.org/
I c 0
6 m
w
z4
.14
)
ol
' m wE Is
2
2 8 2 4 w
6
EF
4- m s
Sm
2
20 40 60 80 20 40
RELATIVE MIGRATION , RELATIVE DEPTH
[image:7.495.118.394.41.584.2]-FRACTION NUMBER
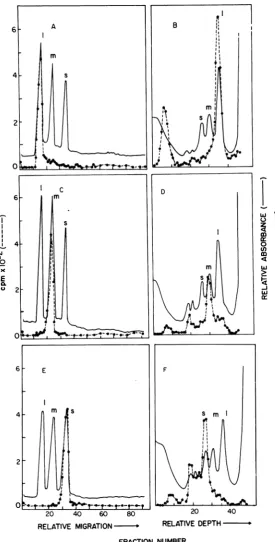
FIG. 5. Abilityof theisolated
06
32P-dsRNA segments to reanneal with a10-fold excess of unfractionated,unlabeled,6dsRNA (A,C,E).B, D,andFindicatethatboth the labeled and unlabeled dsRNAs had separated
intosinglestrands duringthe heat treatment. The protocol of the experiment is described in Materials and
Methods. The solid line represents the
A,,,
on gels (A, C, and E) and A2b4 on sucrose density gradientcolumns (B,D, andF); thedashed linerepresents the counts perminute. The experiments shown in A and B
were conducted with thelarge 32P-dsRNA segment; Cand D were conducted with themedium 32P-dsRNA
segment; and EandFwereconductedwith the small32P-dsRNA segment.
1260
on November 10, 2019 by guest
http://jvi.asm.org/
DOUBLE-STRANDED RNA FROM PHAGE
06
TABLE 3. The sensitivity of
06
32P-dsRNA to RNaseinhigh salt buffer
Counts/ Counts/ Counts/
mn minintop mi ntop
Treatmenta minlayered22.Smlof25mlof' theml
of'
per the gradientgradient gradient ()nt
ExptI'
None 96,400 92 0.095
--RNasesAandT, .. 96.400 2.36:3 2.45
None .. .... 193,000 118 0.061
+RNasesA and T. 193.,00() 4.231 2.19
-RNasesAandTl' 74,740 13:3 0.18 Expt 2b
None .. 19.131 0 0
+RNasesAand T. 17,974 202 1.12
+RNasesAandT," .50.645 61 0.12
a32P-RNAwasincubated with RNase'sA(10pg/ml)and
T, (4,000 units/ml) for 15 min at 22C in 2 SSC priorto layeringonlinearlogsucrosedensitygradients.The gradients were centrifuged in anSW41 rotor for 13 h at 6Cat:37.000 rpm andfractionated into 24 fractions(0.5ml per fraction). Thecountsreleased into the top five fractions (2.5 ml) were taken to indicate RNase released counts. 46 dsRNA sedi-ments in fractions 9through 16.
'Values determined f'rom 32P-dsRNA isolated from two different fermentations.
'06-dsRNAwhichwas collectedfromthe RNase's Aand
T,treatment. precipitatedwithethanol.andthensubjected totheRNasetreatment asecondtime asdescribedin footnote a.
Since the base
compositions of the threedsRNA
segments aresimilar and the
sum of themolecular weights
of the two smaller dsRNA segments is nearthat
ofthelarge
dsRNA, it waspossible that the
two smaller segments werederived
from a specificcleavage
of the largedsRNA
segment. Alternatively, the largedsRNA
segmentmight be
formed by the joining ofthe twosmaller dsRNA
segments. However,the absence
ofhybridization between
the vari-ousRNA segmentseliminates this
possibility.Previously, we reported that phage
t6
con-tained
anRNApolymerase which
mediated the incorporation ofribonucleoside
triphosphates intoall three dsRNA
components,
and wesuggested that the
enzyme was completing one ofthe
complementary strands
ofthe dsRNA
(9).The observation
that 1 to2.5%
ofX6dsRNA
isRNase
sensitive inhigh salt
(2xSSC)
supportsthis
concept.However,
theexperiments do
not provide conclusive evidence thati6
dsRNAactually
contains ashort ssRNA "tail" region. Forexample,
it ispossible
that a "contaminat-ing" ssRNA sediments in a sucrosedensity
gradient in the same region asi6
dsRNA and that this "contaminant" RNA is the source ofthe RNase-sensitive material. Another
possibil-ity is that a small percentage of thei6
virions containincomplete
dsRNA molecules and thatthe majority of the phage genomes are
com-pletely double stranded. Experimental
at-tempts to resolve these three possibilities have been inconclusive.
The average sum of the molecular weights of the three dsRNA segments as determined by fourdifferent procedures is 10.4 x 106(Table1).
Previously, we reported that the phage was
composed of about 13% RNA (10). Because of thesegmented properties of the virusgenomeit is possible that the virion might contain more
than one copy of identical dsRNA segments. However, a 60Co y-rayinactivation experiment indicated that this does not occur (J. Sands,
personal communication). If the phage contains onespecies of each of the dsRNAsegmentsper virion, this would indicate that the molecular weight of the i6particle is about 80 x 106.This value is consistent with the 60-to70-nm size of the phage observed with the electron micro-scope (10). Although it is possible that each of the three dsRNA segments consists of distinct populations of dsRNAs, it would seem to be
unlikely sincerepeatedattempts to separatethe
dsRNA into additional dsRNAsegments by gel electrophoresison 3to7%c polyacrylamidehave
been unsuccessful.
Inconclusion, the phagegenome iscomposed
of three unique species ofdsRNA whose base
compositions aresimilar.
ACKNOWLEDGMENTS
WeareindebtedtoMvron K. Brakke for the brome mosaic virus RNAs and hishelpful advice.
Thisinvestigationwassupportedinpartby PublicHealth Service Grant AI-10638 fromthe NationalInstituteofAllergy and Infectious Diseases. Research reported was conducted underProject no. 21-21,Nebraska Agricultural Experiment Station.
LITERATURE CITED
1. Bishop. D. H. L.. J. R. Claybrook, and S.Spiegelman. 1967. Electrophoreticseparationof viralnucleicacids
onpolyacrvlamide gels. J.Mol. Biol.26:373-387. 2. Brakke,M.K.,andN.VanPelt. 1970.Linear-logsucrose
gradients for estimating sedimentation coefficients of
plant viruses and nucleic acids. Anal. Biochem. 38:56-64.
3. Brakke, M. K., and N. Van Pelt. 1970. Properties of
infectious ribonucleic acid from wheat streakmosaic
virus.\'irology42:699-706.
4. Diener,T.O.,andI.R. Schneider. 1968. Virus
degrada-tion and nucleic acid release in single-phase phenol
systems.Arch. Biochem.Biophys. 124:401-412.
5. Franklin. R. M. 1967.Replicationofbacteriophage
ribo-nucleic acid:somephysical propertiesof single-strand-ed, double-strandsingle-strand-ed, and branched viral ribonucleic
acid. J. Virol. 1:64-75.
6. Katz,L., andS. Penman. 1966. The solvent denaturation
of double-stranded RNA from polio virus infected
HeLa cells. Biochem. Biophys. Res. Commun.
23:557-560.
7. Lemke, P. A., and T. M. Ness. 1970. Isolation and
characterizationofadouble-strandedribonucleic acid
VOL. 13, 1974 1261
on November 10, 2019 by guest
http://jvi.asm.org/
VAN ETTEN ET AL.
fromPenicillium chrvsogenum. J.Virol. 6:813-819. 8. Semancik, J. S., A. K. Vidaver. and J. L. Van Etten.
1973. Characterization of asegmented double-helical RNA frombacteriophage06.J. Mol. Biol. 78:617-625. 9. VanEtten, J. L.. A. K. Vidaver, R. K.Koski, andJ. S.
Semancik. 1973. RNApolvmerase activitvassociated withbacteriophage <6. J. Virol. 12:464-471. 10. Vidaver. A. K., R. K. Koski. and J. L. Van Etten. 1973.
Bacteriophage k6: alipid-containing virusof Pseudo-monasphaseolicola.J. Virol. 11:799-805.
11. Wood, H. A. 1973. Viruses with double-stranded RNA
genomes.J.Gen.Virol.20:61-85.
12. Zaitlin. M.. and V. Hariharasubramanian. 1970. An improvement ina procedure for counting tritium and carbon-14 in polvacrvlamide gels. Anal. Biochem. 35:296-297.