0022-538X/81/070255-08$02.00/0
Cessation
of
Reentry of Simian Virus
40
DNA into Replication
and Its
Simultaneous
Appearance in
Nucleoprotein
Complexes
of the
Maturation Pathway
HWA-TANG WANG AND ANN ROMAN*
Departmentof Microbiology and Immunology, Indiana University School of Medicine, Indianapolis,
Indiana46223
Received 11 February1981/Accepted 15 April 1981
Newly synthesized SV40DNA isused as a template for further DNA synthesis
(reenters replication) or as a substrate in the assembly of virions (maturation
pathway). The timecoursesofreentry intoreplicationandprogressionalongthe
maturationpathwaywereboth determined onidentical samples. DNA,
synthe-sizedduringa20-minpulse, reentered replicationover aperiod of several hours
and thenwasremovedfromthepoolofmolecules availableforreplication. The
cessation ofreentrycoincided with the maturation of this DNA from the
chro-matinformtoprevirion and virion forms. Morereentryand less maturation was
observedat 24hpostinfection thanat42hpostinfection.Thedataareconsistent
withthe hypothesis that the factor(s) responsibleforcessation of reentry is also
responsible for initiation ofthematuration pathway.
The stepsin the maturation (encapsidation)
pathway of simian virus40
(SV40)
arecurrentlyunderinvestigation in several laboratories (1,3,
4, 6-8, 11). New extraction procedures have
re-vealed that (i) the majority of newly synthesized
SV40
DNA isencapsidated and that (ii) severalspecies of nucleoprotein complexes (NPCs)can
be isolated (1,3,6-8). Pulse-chase experiments
suggest that labeled SV40 DNA is converted
from chromatin
(NPC-I)
toprevirions(NPC-II)tomaturevirions(1, 3, 6-8). NPC-I iscomposed ofDNA andcellular histones
Hi,
H2a, H2b, H3,and H4 (3, 7). It is used for biosynthetic
pro-cessesand has associated with it SV40Tantigen
(13, 19, 23), the DNA-binding
protein
involvedin initiation of viralDNAsynthesis (14, 24, 25,
27) and
regulation
ofviraltranscription
(18, 20,26), and cellular
proteins
involved in thesepro-cesses
(for
examples,
seereferences 5,15).
NPC-IIis aheterogeneousset of
complexes
grouped
together because they sediment more
rapidly
than NPC-I under neutral conditions and yet
are salt labile. NPC-II is
composed
ofDNA,
histones withahigher
degree
ofacetylation
thanNPC-I, and viral structuralproteins (3, 4, 11).
Maturevirionsaresaltstable and containDNA, allhistones except
Hi,
and viralstructuralpro-teins (3, 12, 16). The order of addition ofviral
proteinstoanNPChasnotbeendetermined.
Althoughthese studies shed
light
onthe stepsinvolvedin virus
assembly,
littleis known of thefactor(s)
determining
the fate ofnewly
synthe-sized DNA. Romanand Dulbecco
(22)
showedthatnewly synthesizedpolyomaDNA can serve
as atemplate forfurtherDNAsynthesis(reenter
replication)overaperiodof2to 3 h and then is
removed from the pool of molecules available
for replication. Green and Brooks (9) reported
similarkinetics ofreentryofnewly synthesized
SV40 DNA into
replication.
Recently, Romanreported that polyoma DNA labeled at 20 h
postinfection (p.i.) reentered replication at a
greater ratethanmolecules labeledat 28hp.i.
and that agreater proportion of molecules
la-beled at 20h
p.i.
reenteredreplication
than at28hp.i. (21). These datasupportthehypothesis
of Roman and Dulbecco that therateofreentry
is a reflection of the availability of
proteins
required for initiation of DNA synthesis (22).
Regardless
of thetimep.i.
examined,however,
progenymolecules
appeared
tobe removed fromthe
pool
available forreplication
about3haftertheir initial
synthesis.
A role for maturationproteins was
postulated
by Roman andDul-becco(22) and Roman(21).
Similarly,
Garberetal.(8) and
Baumgartner
etal.(1)compared
theirdeterminations on the rate of
encapsidation
ofDNAwith the
published
dataonthe kinetics ofreentryof DNA into
replication
(9, 21, 22),andpostulatedanassociationbetween thetwo
path-ways.
Theresearchreportedherewasconductedto
determine, onidentical
samples,
thekinetics ofreentryofnewly
synthesized
DNAintoreplica-tionand the kinetics ofmaturationof thissame
DNAto establish the
temporal
relationship
of255
on November 10, 2019 by guest
http://jvi.asm.org/
256 WANG AND ROMAN
the twopathways toeach other. Since Roman had previously reported that the kinetics of
reen-try were dependent upon the time in the
infec-tiouscycle examined(21), the kinetics of thetwo
pathways were also determined at24 and 42h p.i. Cessation ofreentryintoreplication of DNA
labeledduringa 20-minpulsewasfound to
co-incide with the maturation of this DNA from
chromatin to previrions and virions. The rate
and extent ofreentry decreased with time p.i.
whereas the rate ofencapsidation increased.
MATERIALS AND METHODS
Cellsand virus. BSC-1cells,agiftfrom M.Singer,
weregrowninDulbecco modifiedEaglemedium
con-taining 10%calfserum and50,ugofgentamycinper
ml. SV40,agift from W.Eckhart,wasplaque purified
three times andpropagatedat alow(0.05)multiplicity
of infection. Virus stockswerepreparedfrom infected
cells by threecyclesoffreezingandthawingfollowed
by low speedcentrifugation andtheywere storedat
-200C.
Labeling and extraction of viral DNA and
NPCs.ConfluentBSC-1 cellswereinfected withSV40
at multiplicity of infection of 10 PFU per cell. At
different times after infection, the cells wererinsed
twotimes with TDbuffer(150mMNaCl,5mMKCI,
0.7 mM Na3HPO4, and 20 mM Tris, pH 7.4), and
labeled for20minwith75or150,uCiof[3H]thymidine
(specificactivity, 40 to 60Ci/mmol) per ml in
Dul-beccomodified Eagle medium containing 2% serum
and25mMHEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid) (pH7.0). Cellswerethenrinsed
twicewith TDbuffer,covered with Dulbeccomodified
Eaglemediumcontaining106 Mthymidine,2%serum
and25mMHEPES for30min,and then chasedwith
bromodeoxyuridine(BUdR) chasemedium[5x 10-'
M5-bromodeoxyuridine, 2 x 10-5 M
5-fluorodeoxyu-ridine(28),9x10-6Mdeoxycytidine (2, 17),2% serum,
and25mMHEPES in Dulbecco modified Eagle
me-dium].
After variouslengthsofchase,DNAwasextracted
from half of theplatesby the method of Hirt (10). The
Hirt supernatantwasusedto determine thekinetics
of reentry.
NPCs were extracted from infected cells by the
method of Fernandez-Munoz et al. (7). Cells were
rinsed twice with coldTBS buffer (TD buffer
contain-ing 0.9 mMCaCl and 1mMMgCl2)and scraped into
cold hypotonic buffer (25 mM Tris, pH 7.9, 1 mM
MgCl2,and 0.4 mMCaCl2).Thecellswere broken by
homogenization (30 strokes) inatightly fitting glass
Douncehomogenizer. The lysates were centrifuged at
2,000 rpmfor5min.The NPC-containing supernatant
was used to determine the kinetics of maturation.
Similar results were obtainedifthecellswererinsed
with TD buffer or extracted with hypotonic buffer
containing0.5 Mdithiothreitol and
phenylmethylsul-fonylfluoride.
Theefficiency of extraction of3H-labeledDNA in
the form of NPCswasdetermined by comparing the
total 3H countsper minute (cpm) in unfractionated
NPCstothe total3Hcpm inDNAextracted by the
J. VIROL.
method of Hirt. Theefficiency of extraction of NPCs
wasapproximately 70% that of DNA. During the 6-h
chase,nodecrease in the 3H cpmwasobserved,
sug-gesting that essentially all of the prelabeled DNA
remained cell associated.
Analysis of viral DNA. Form I DNA was
sepa-rated fromform II andreplicative-intermediate DNAs
bylayering Hirtsupernatantsonto5to20%alkaline
sucrose gradients containing 0.75 M NaCl, 1 mM
EDTA, and 0.3 M NaOH(pH12.6). Thesampleswere
centrifuged inanSW41 Tirotor at39,000 rpm for 2.6
h. Fractions were dripped from the bottom of the
centrifuge tubes, and theradioactivityinasamplewas
determined afterprecipitationwith 5% trichloroacetic
acid. Fractions containingform I were pooled,
neu-tralized, and mixed with CsCl in TE buffer (10 mM
Tris,pH 7.4,1 mMEDTA)togiveafinaldensityof
1.72g/ml. Thesampleswerecentrifugedinatype40
rotor at33,000 rpmfor48htoseparateform I DNA
with BUdR substitution inonestrand(HL DNA)from
unsubstituted form I DNA(LL DNA).The percentage
of 3H-labeled HLformIDNA[(HLDNA cpm+ HL
DNAcpm+LLDNAcpm)x 100]wasdetermined.
In someexperiments, thesameprotocolwasused
to determine the percentageof labeled HL DNA in
theslowlysedimenting DNA (formII, RI,cellular)of
theinitial alkalinesucrosegradient. Whenmentioned,
the percentage of labeled HL DNA in the Hirt
super-natantwasalso determined inCsCl.
Analysis ofSV40 NPCs. The NPC supernatants
werelayeredonto 10 to40%sucrosegradients
contain-ing 2 mM Tris(pH7.4) and spun inanSW41 Tirotor
at37,000 rpm for90min. Fractionsweredrippedfrom
the bottom and processed as described above. The
percentagesofNPC-I and of NPC-II+Vwere
deter-minedby the formulas %(NPC-I) = (NPC-I cpm +
total NPCscpm) x 100and%(NPC-II+V) =
(NPC-II+V cpm + total NPCs cpm) x 100, respectively.
Fractions from theNPC-II+V regionofthegradient
were pooled, and salt-labile NPC-II was separated
from salt-stable virions by density gradient
equilib-riumcentrifugation inCsCl (p =1.34g/ml in TBS).
Thesampleswere spun in atype40rotor at 33,000
rpmfor40h. Fractions were dripped from the bottom
andprocessedasdescribed above. The percentage of
virionswascalculatedby theformula(V cpm+
NPC-II+Vcpm)x%(NPC-II+V).Similarly,thepercentage
ofNPC-II was equal to (NPC-II cpm + NPC-II+V
cpm) x %(NPC-II+V). Where noted, the different
NPCs werepooled, and the DNA was extracted by the
method of Hirt (10) andsubjected to density gradient
equilibrium centrifugation as described above to de-termine the percentage of HL DNA in each NPC.
RESULTS
Location of NPCs.Several laboratories have
reported the
sedimentation
and buoyant densitycharacteristicsof chromatin (NPC-I), previrions
(NPC-II), andvirions (1, 3, 6-8). Initial
experi-mentswereconducted to locate these complexes in sucrose and CsCl gradients. SV40-infected
BSC-1 cellswere labeled for 20minwith
[3H]-thymidine at 33 h p.i. NPCs wereextracted, at
on November 10, 2019 by guest
http://jvi.asm.org/
the end ofthe pulse or after a 6-h chase with
10'
Mthymidine, andseparated on 10 to 40%sucrose gradients (Figure 1A and 1B,
respec-tively). With a 12-min pulse, only the slower
sedimenting species of NPC was labeled (data
not shown). Thelabeling kinetics and location
of thisspecies in the gradientidentify it as
NPC-I (1, 3,6-8). The more rapidlysedimenting
spe-cies contained NPC-II+V.This was verified by
poolingthe fractions (Fig. 1A and B) and
sub-jecting the complexestoequilibriumdensity
gra-dient centrifugation in CsCl(p = 1.34g/ml) (Fig.
1C andD). Whereas the NPCs located in the
601
50-40
f
E
a
0u
0~
3020
10
0 80
70
60
50
E
v.
40 30
20
10
NPC-II+V region aftera20-minpulsewere
pre-dominantly salt-labile NPC-II, approximately
50% were salt stable after a 6-h chase. The
sucrose gradients were used to quantitate the
distribution oflabelbetweenNPC-I and
NPC-II+V. When necessary, NPC-II andvirionswere
further separated on CsCl. It should be noted
that, on the CsCl gradients, a new NPC was
detected which had a greater buoyant density
thanmaturevirions. Thiscomplexwasseparable
from bothNPC-II and mature virions and
ap-pearedtobeanintermediate in the maturation
pathway andnot abreakdownproduct of virions
5 10 15 20 25 5 10 15 20 25
FRACTION NUMBER
FIG. 1. Separation of NPCsonsucroseandCsClgradients.SV40-infectedBSC-1 cellswerelabeledfor20
minat33hp.i.with75
gCi
of[3HJthymidineperml(ca.40to60Ci/mmol). NPCswereextractedfromsomecellsafterthepulse (AandC), fromothersaftera6-hchasein10-6 M thymidine(BandD).NPC-containing
supernatants weresedimented on 10to40%sucrosegradients containing2 mMTris (pH 7.4) (A andB).
Gradientswerecentrifugedfor90minat37,000rpm inanSW41Tirotor.Sedimentation isfrom righttoleft.
Fractionswerepooledasshown andbandedtoequilibriuminCsCI(p=1.34g/ml)inatype40rotorat33,000
rpmfor40h(CandD).Thearrowrepresentstheposition ofthemajority ofcomplexeslabeled ina12-hpulse. Thedensity ofthismaterialis 1.34g/ml.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.128.370.208.569.2]258 WANG AND ROMAN
(data notshown);it was namedNPC-III. Inthe
quantitation ofthe percentage ofnewly
synthe-sized DNA inchromatin, previrions, and virions,
thecpminNPC-IIIwereaddedtoNPC-II, since
bothareprevirions.
Effect of BUdR substitution on
encapsi-dation. The protocol for following reentry of
newlysynthesized DNA into replication involves
the use of BUdR. Thus, it was necessary to
determinethe effect of BUdRontheprocessof
encapsidation. At 36 h p.i., 4 plates of
SV40-infected BSC-1 cells were labeled for 20 min
with [3H]thymidine and chased with 10-6 M
thymidine for30min.NPCswereextracted from
twoplates.Athirdwasrinsed andcovered with
Dulbecco modified Eagle medium containing
BUdRchase medium.Theremaining platewas
rinsed, and Dulbecco modified Eagle medium
with 106Mthymidine was added. NPCswere
extracted from the latter two
plates
6 h later.Thepercentageof NPC-II+V extracted from all
fourplateswasdeterminedon10 to40%sucrose
gradients. BUdRsubstitution didnotaffect the
extentof accumulation ofNPC-II+Vduring the
6-h chase
(Table
1). Thesetwospecies
ofNPCswere further separated on CsCl. Virions
accu-mulatedto asimilarextentunder thetwochase
conditions(Table 1).
Comparison of the kinetics of
maturation
and kinetics ofreentry. Todirectlycompare
thetimecourseof maturationtothat ofreentry, at 33 h p.i., SV40-infected BSC-1 cells were
pulse-labeled
for20minwith[3H]thymidine
(75,uCi/ml),
chased for30minwith 106M thymi-dine, and chased again with the BUdR chasemedium for different lengths of time.The
kinet-ics of maturation of this pulse-labeledDNA was
determined by plotting the percentages of
la-TABLE 1. Effect ofBUdRontheextentof
encapsidationof viral chromatin
%(NPC- %Wt
Chase medium
II+V)'
at: %(V)bat:Oc 6 0 6
BUdR 42 89 7 45
10-6 Mthymidine 43 91 8 46
aThe percent ofNPC-II+V wasdetermined from
separationon 10to40%sucroseby using the formula
(NPC-II+V cpm + total cpm of NPCs)x 100.
bThe percent of virions (V) wasdetermined after
the separation of NPC-II from V by CsCl density
equilibrium centrifugation by using the formula
%(NPC-II+V) x(V cpm + NPC-II+V cpm).
cChase time in hours. SV40-infected BSC-1 cells
were labeled for 20 min with [3H]thymidine and
chasedfor30minwith10-6 M thymidine. NPCs were
either extractedatthis time(0)oraftera6-hchasein
the mediumshown(6).
beledNPC-I,NPC-II, andvirionsversusBUdR
chase time (Fig. 2).At theend ofthe
pulse
and30-min chase with 106 M thymidine (T = 0),
the majority of
[3H]DNA
was in NPC-I; verylittle was in virions. As the BUdR chase time
increased,thepercentage of[3H]DNAin
NPC-I decreased, with the rate of decrease being
greatestduring the first2 to 3h after initiation
of the BUdR chase. Therewas a concomitant
increase in the fraction of
[3H]DNA
appearingin virions. The fractionof
[3H]DNA
in NPC-IIremained
relatively
constant during the chaseperiod (Fig. 2). The level of the NPC-II pool
varied fromexperimenttoexperiment (compare
Fig. 2A and B; similar variations in levels of
NPCshavebeenseenby others [6]). Withinany
one experiment, the level of NPC-II was the
samein thepresenceorabsence ofdithiothreitol
andphenylmethylsulfonylfluoride. Therefore, it
seems
unlikely
that this variation is due tobreakdown of virions.
Todetermine the reentry kinetics for newly
synthesized
DNAduring
the BUdRchase,DNAwasextractedby theHirtmethod, formIDNA
was isolated onalkalinesucrosegradients, and
HL DNAandLL DNAwereseparated by CsCl
densityequilibrium gradient centrifugation.
3H-labeled HL DNA represented DNAthat
repli-catedduring the short
3H
pulse andwassubse-quentlyused astemplateforfurther DNA
syn-thesis(reentered replication).Thepercentage of
HLDNAplottedversuschasetime with BUdR
showed the kinetics of reentry of the
pulse-la-beledDNAinto
replication (Fig.
3). It should benoted that only one round ofreentry was
de-tectedby the
protocol
since form I DNA withBUdRsubstitution in both strands (HH DNA)
wouldnotbelabeled. LL DNAreentered
repli-cation(wasconvertedtoHLDNA)over aperiod
of2 to3h. After thislengthofchase,either the
pulse-labeledDNAceasedtoreenterreplication
or the rate at which it reentered greatly de-creased.
Also plotted in Fig. 3, for direct comparison
with the reentry
kinetics,
is the rate atwhich[3H]DNA left the NPC-I pool, calculated from
the data in Fig. 2. A good correlation may be
seenbetween the time thatpulse-labeledDNA
wasremoved from thepool of molecules
avail-able forreplicationand thetimeit left the
NPC-Ipool.
The NPC data presented in Fig. 2 represent thefate of all pulse-labeledDNA, whereas the reentry data in Fig. 3represent only the fate of
forn
I DNA (containing 85 to 89% of the3H
cpm) in a Hirt
supernatant.
The reentry kineticswas,therefore, determined directly on a sample
ofaHirt
supernatant
and inparallelonthe formJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.53.244.465.528.2]z
a I
LL
0
0 2 4 6 0 2 4
LENGTH BUDR CHASE-HRS
FIG. 2. Relationship between the length of BUdR chase and percentage of
[3H]DNA
inNPC-I, NPC-II,and virions. (A and B) Results of representative experiments. NPCs were isolated as described in the text.
Symbols:0,percentageof NPC-I; A, percentage ofNPC-II; A, percentage of virions.
<: zo 00
I
0n
cL IUU
0a.
zz
u>
=u-i
:L-0 2 4 6
*1UO
z 0
-j
20 m
0 2 4 6
[image:5.493.104.397.64.296.2]LENGTH BUDR CHASE- HRS
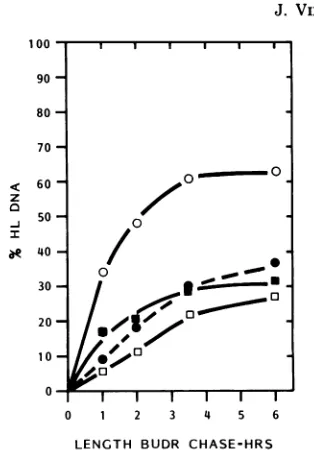
FIG. 3. Relationship between the fraction of the initial[3H]DNAinNPC-I leaving the NPC-Ipool and the
percentageof 3H-labeledHL DNA (kinetics of reentry) with increasingchasetime in BUdR. (A and B) Data
from thesameexperiments showninFig.2A and B.To calculate thefraction of[3H]DNA leaving the
NPC-Ipoolduring the B UdR chase, thetotalpercentageof NPC-Iatthe timeBUdR was added (e.g., 51% atT=
0;Fig. 2A)was setequalto1.0.Thefraction of[3H]DNAleaving the NPC-Ipool is determined bytheformula
1-[o(NPC-IT=
x+
%(NPC-IT=-q],in which X and0 arethelengthsof the BUdR chase. The percentage of3H-labeled HL DNAwasdeterminedafterseparation offormIHL and LL DNA moleculesby CsCl density
equilibriumcentrifugationasdescribed in thetext.
I DNAisolated from thatsupernatant. The
ki-netics ofreentrywasqualitativelysimilartothat
determined for form I DNA (Table 2). The
percentage of HL DNA, however, was lower
than that found when only form I DNA was
analyzed.Thiswasexpected,since thenon-form
I DNAisolatedonalkalinesucroseandanalyzed
on CsCl contained only
3H-labeled
LL DNA(datanotshown).
Distribution of 3H-labeled HL DNA in
NPC-I, NPC-II, and virions. To determine
more precisely the relationship of reentry to
maturation, the percentages of 3H-labeled HL
DNA in NPC-I and in NPC-II+V were deter-259
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.53.448.345.485.2]260 WANG AND ROMAN
mined. SV40-infected cells were pulse-labeled and chased for various lengths of time as de-scribed above. NPC-I and NPC-II+Vwere
sep-arated on 10 to40% sucrose gradients, and the percentage of HL DNAwasdetermined on each pool. In parallel, the percentage of HL DNA in form I isolated from a Hirt supernatant was
determined. Figure4showstheaccumulation of HL DNA inNPC-I, NPC-II+V, and form I. 3H-labeled HL DNA accumulated in NPC-I for a period of 2 to 3 h then ceased toaccumulate, a pattern qualitatively similar to the kinetics of reentrydetermined on form I DNA. 3H-labeled HL DNA accumulated more slowly and to a lesser extent inNPC-II+V. It is possible to cal-culate the reentry kinetics curve for total NPCs
bytakinginto accountthe relative contribution
of each NPC pool tothe totalNPCs extracted at each time. This curve isvery similar to that foundby analyzing form I DNA (Fig. 4).Thus, the kinetics ofreentry establishedbyanalysisof form I DNA isdirectlycomparabletothe kinet-ics ofmaturation.
Comparisonof thekineticsofreentry and
maturation at 24 hp.i.versus 42 hp.i.
Ro-man(21)demonstratedthat thekineticsof
reen-try ofpolyoma DNA was dependent upon the time in the infectious cyclethat wasexamined.
Earlyin thereplicationcycle, reentry occurred
more rapidly and to a greater extent than at
latertimes. If the cessation ofreentry is related to astep in the maturationpathway,thekinetics
ofmaturationshould alsobedifferentat
differ-enttimes.Therefore,thekineticsofreentry and
maturationweredetermined at 24and 42 hp.i.
(Table 3). More extensive reentry was seen at
24 h p.i. than at 42 h p.i. When cells were
harvested aftera20-minpulseand30-min chase
(T=0),thepercentageoftheNPC-II+V at42 hp.i.wastwice that seen at 24 h p.i.Therefore,
pulse-labeledDNAproceedsalongthe
matura-tion pathway more rapidly later in infection.
TABLE 2. Kineticsof reentry determinedonisolated
formIDNAorunfractionated Hirtsupernatantsa
% of HL DNA at: HL DNA source
ob 2 6
Form I 0 17 26
Hirt supernatant 0 15 24
aAt36h
p.i.,
SV40-infectedcells
werepulse-labeledwith [3H]thymidine, chased with 10' M thymidine,
and chasedagain for different lengths of time with a BUdR chase mixture. DNA was isolated by the Hirt
method (10). Eitherasamplefrom the Hirt
superna-tantwasdirectly analyzed by CsCldensity equilibrium
centrifugation,orformI DNAisolatedfrom the Hirt
supernatantwasanalyzed.
bChase time in hours.
100
z
6
bU -o- 50- o
40 /
302g
20-10~~~~~~~
1-
0-0 1 2 3 4 5 6
LENGTH BUDR CHASE-HRS
FIG. 4. Percentage of 3H-labeled HL DNA in
NPC-I, NPC-II+V, total NPC, and form I DNA.
Poolsof NPC-I andNPC-II+V,asshown inFig. 1,
weredeproteinized, put into CsCl gradients (p=1.72
g/ml) andcentrifuged for48hat33,000 rpm inatype
40 rotor to separate 3H-labeledHL DNA from
3H-labeled LL DNA.Symbols:0, Percentage ofHL DNA
inNPC-I; O, percentageof HL DNA inNPC-II+V;
0, percentage of HL DNA inform I;E, calculated
percentageof HL DNA in total NPC. The percentage
of HL DNA in total NPC at each timepointwas
calculatedby the formula[%(HL DNA inNPC-I)x
(NPC-I cpm . total NPC cpm)]+ [%(HL DNA in
NPC-II+V)x (NPC-II+Vcpm +totalNPC)J.
TABLE 3. Kinetics of reentry and maturation of
DNApulse-labeledat 24and42hp.i.
Timep.i. %of HL DNAat: % ofNPC-II+Vat:
(h) oa 2 6 0 2 6
24 0 28 45 25 51 80
42 0 16 31 50 64 80
aChase time in hours.
The percentage of NPC-II+V was the same (80%) after a 6-h BUdR chase at the two times; the20% of[3H]DNAwhichremained in NPC-I may be eitherunavailable for packaging or
in-efficiently packaged.
DISCUSSION
Experiments were undertaken to determine
whether the cessation ofreentry of newly syn-thesized DNA into replication coincided with theprogression of that DNAalong the matura-tion pathway. SV40-infected BSC-1 cells were labeled for a short period with [3H]thymidine
andthen chased with BUdR. Theuse ofnewly
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.277.434.49.274.2] [image:6.493.259.451.437.500.2]synthesized DNA as the template for further DNA synthesis (reentry into
replication)
wasmeasured by the conversion of H-labeled LL
DNA to3H-labeled HL DNA (Fig. 3, Table 3). In parallel, its progression along the maturation pathwaywasmonitored by determining the per-centageof 3H cpm in different NPCs (Fig. 2 and 3; Table 3). The time course of appearance of
[3H]DNAinNPC-I,NPC-II, and virions (Fig. 2)
wasconsistentwith previous reports that NPC-Iprogressed to virions through the intermediate NPC-II (1, 3, 6-8).
Two facts emerge from this study. First, ces-sation of reentry and maturation are closely linked intimne.There do not appear to be factors that terminate reentry for anysignificant period oftime before maturation. Cessation of reentry
coincides with the maturation of DNA from
NPC-ItoNPC-IIorvirions (Fig.3).Thetime of
cessation of reentry and of the conversion of
NPC-I toNPC-II+V varies somewhat from
ex-perimenttoexperiment, butthe two arealways
coincident (Fig. 3). Second, newly synthesized DNA becomes encapsidated more rapidly late in the infectious cycle. This is consistent with
theobservationthat therateand extent of
reen-trydecreaseswith timep.i. (Table3).After a
6-hBUdR chase, 80% of the pulse-labeled DNA hasmaturedtoNPC-II+V,whether the experi-ment wasinitiated at24 or42 hp.i. (Table 3). However, when thepool of NPC-II+V was
sep-aratedonCsCl,24and 59%ofthe
[3H]DNA
wasin the virionfractionat 24and 42 hp.i.,
respec-tively (datanotshown). This suggests that not
only isthere a slower progressionofNPC-I to
NPC-II+V early in infection but also that the
progressionfromNPC-IItovirions isslower.
We propose that the fate of a newly
synthe-sized SV40 moleculeis determined by whether
it isboundfirstbyaproteinorproteins involved
inreplication (or transcription) or byaprotein
or proteins involved in maturation. This
com-petition,between thebindingoflargeTantigen
(andperhapsotherproteinsinvolved in DNAor
RNA synthesis) and a structural protein(s),
could beaffected
by
therelative abundance ofearlyversuslate
proteins.
Theaffinity
ofdiffer-entproteins for the NPC-I
template
would alsoaffect the outcome of the
competition.
Coca-Pradosetal. (4)
recently proposed
that whethera DNA molecule was used for replication or
maturationmight dependontheextent of
ace-tylation of histones, with structural proteins
having a higher affinity for histones with a
greaterlevel ofacetylation.
In summary, the data presented suggest a
close correlation betweenremoval of DNA from the pool ofmolecules available for replication
and progression of this same DNA
along
thematurationpathway. Thesimultaneousanalysis
of the kinetics of reentry and maturation, using
severalmutantsofSV40 defectivein known viral
proteins,should result in a detennination of viral proteins involved in one or both of these pro-cessesandreveal essential details of their
inter-dependence.
ACKNOWLEDGMENTS
WethankRichard Bockrath, Howard Edenberg,andSteve
Larsenfor helpful suggestions during thepreparation of the
manuscript.
Thisinvestigationwassupported by PublicHealthService
grants CA-19708 and CA-29318 from the National Cancer InstituteandbyaProgramDevelopmentProject grant from the IndianaUniversitySchool of Medicine.
LITERATURE CITED
1. Baumgartner, I., C. Kuhn, and E. Fanning. 1979.
Identification and characterizationoffast sedimenting SV40 nucleoproteincomplexes. Virology96:54-63. 2. Ben Porat, T. C.,C. Coto,and A.S. Kaplan. 1966.
Unstable DNAsynthesized by polyoma virus-infected cells. Virology30:74-81.
3. Coca-Prados, M., and M.-T. Hsu. 1979. Intracellular
formsof simian virus40nucleoprotein complexes. II.
Biochemical and electron microscopic analysis of simian
virus 40virionassembly.J.Virol. 31:199-208.
4. Coca-Prados, M., G.Vidali, and M.-T. Hsu. 1980.
In-tracellular forms of simian virus40nucleoprotein
com-plexes. III.Study of histonemodifications.J. Virol.36: 353-360.
5. Edenberg, H., S. Anderson, and M. DePamphilis. 1978.Involvement of DNA polymeraseinsimianvirus 40DNAreplication. J.Biol. Chem.253:3273-3280.
6. Fanning,E., and I. Baumgartner. 1980.Role of fast-sedimenting SV40nucleoprotein complexesinvirus
as-sembly. Virology102:1-12.
7. Fernandez-Munoz, R., M. Coca-Prados, and M.-T. Hsu. 1979.Intracellular forms of simianvirus 40
nu-cleoprotein complexes.I.Methods of isolation and char-acterization ofCV-1 cells.J.Virol.29:612-623. 8. Garber,E.A.,M. M.Seidman,and A. J. Levine.1978.
The detectionand characterizationofmultiple formsof
SV40nucleoprotein complexes. Virology90:305-316. 9. Green,M.H., andT.L. Brooks. 1978.Recently
repli-catedsimian virus 40 DNA is apreferential template fortranscription andreplication.J.Virol.26:325-334. 10.Hirt,B. 1967.Selective extractionofpolyoma DNA from
infectedmousecell cultures. J.Mol.Biol.26:365-369. 11.LaBella, F.,andC.Vesco. 1980. Latemodifications of
simianvirus 40 chromatinduringthelytic cycleoccur
inanimmatureform of virion. J. Virol.33:1138-1150. 12.Lake, R. S., S. Barban, and N. P. Salzman. 1973.
Resolutionandidentificationofthe core deoxynucleo-proteins of the simian virus40.Biochem.Biophys.Res.
Commun.54:640-647.
13.Mann, K., and T. Hunter. 1979.Association of simian
virus40Tantigen with simianvirus 40nucleoprotein complexes.J.Virol. 29:232-241.
14.Myers, R. M., and R. Tjian. 1980. Construction and
analysisof simian virus 40originsdefective intumor
antigenbindingand DNAreplication.Proc.Natl. Acad. Sci. U.S.A. 77:6491-6495.
15. Otto,B., andE.Fanning. i978. DNApolymeraseais associated with replicating SV40 nucleoprotein
com-plexes.Nucleic Acids Res. 5:1715-1728.
16. Pett, D. M., M. K. Esters,and J. S. Pagano. 1975. Structuralproteinsofsimian virus 40. I. Histone char-acteristics oflow-molecular-weightpolypeptides.J.
on November 10, 2019 by guest
http://jvi.asm.org/
262 WANG AND ROMAN
rol.15:379-385.
17. Reichard, P., Z. N.Canellakis, andE.S. Canellakis.
1961.Studieson apossible regulatorymechanism for
thebiosynthesis ofdeoxyribonucleicacid. J. Biol. Chem. 336:2514-2519.
18. Reed, S. I., G. R. Stark, and J. C. Alwine. 1976.
Autoregulation ofsimian virus 40geneAbyT antigen. Proc.Natl. Acad. Sci. U.S.A. 73:3083-3087.
19. Reiser, J.,J.Renart, L.V.Crawford, and G.R.Stark.
1980.Specific association of simianvirus 40tumor
an-tigen with simianvirus 40 chromatin.J. Virol. 33:78-87.
20. Rio, D., A.Robbins, A. Myers, and R. Tjian. 1980.
Regulationof simian virus 40early transcriptioninvitro by a purified tumor antigen. Proc. Natl. Acad. Sci. U.S.A.77:5706-5710.
21. Roman,A. 1979. Kinetics ofre-entryofpolyomaprogeny
form I DNA intoreplicationas afunction oftime
post-infection.Virology96:660-663.
22. Roman, A., and R. Dulbecco. 1975. Fate ofpolyoma
form I DNAduring replication.J. Virol. 16:70-74. 23. Segawa, M.,S.Sumio,and N.Yamaguchi.1980.
As-sociationof simian virus 40 Tantigenwithreplicating nucleoprotein complexesofsimian virus40.J. Virol.35: 320-330.
24. Shortle,D. R., R.F.Margolskee, andD. Nathans.
1979.Mutationalanalysis ofthesimian virus40
repli-con:pseudorevertantsofmutantswithadefective
rep-licationorigin. Proc.Natl.Acad. Sci. U.S.A. 76:6128-6131.
25. Tegtmeyer, P. 1972. Simian virus 40deoxyribonucleic acidsynthesis:the viralreplicon. J. Virol.10:541-598. 26. Tegtmeyer, P.,M. Schwartz,J. K. Collins, andK.
Rundell. 1975.Regulationoftumorantigen synthesis by simian virus40geneA. J. Virol.16:168-178. 27. Tjian,R. 1978.Thebinding siteonSV40DNAforaT
antigen related protein.Cell13:165-179.
28. Weil, R., M. R.Michel, and G. K.Ruschman. 1965.
Induction of cellularDNAsynthesis by polyoma virus.
Proc.Natl.Acad. Sci. U.S.A.53:1468-1475.
J. VIROL.


![FIG. 3.percentagefrom Relationship between the fraction of the initial [3H]DNA in NPC-I leaving the NPC-Ipool and the of 3H-labeled HL DNA (kinetics of reentry) with increasing chase time in BUdR](https://thumb-us.123doks.com/thumbv2/123dok_us/1477198.100352/5.493.104.397.64.296/percentagefrom-relationship-fraction-initial-leaving-kinetics-reentry-increasing.webp)