JOURNAL OFVIROLOGY, July 1995, p. 4500–4505 Vol. 69, No. 7 0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
Processing and Evolution of the N-Terminal Region of the
Arterivirus Replicase ORF1a Protein: Identification
of Two Papainlike Cysteine Proteases
JOHAN A.DENBOON,1KAY S. FAABERG,2JANNEKE J. M. MEULENBERG,3
ALFRED L. M. WASSENAAR,1PETER G. W. PLAGEMANN,2
ALEXANDER E. GORBALENYA,4,5
ANDERIC J. SNIJDER1*
Department of Virology, Institute of Medical Microbiology, Faculty of Medicine, Leiden University, Leiden,1
and Department of Virology, Institute for Animal Science and Health (ID-DLO), Lelystad,3
The Netherlands; M. P. Chumakov Institute of Poliomyelitis and Viral Encephalitides, Academy of Medical Sciences, Moscow, Russia4; Department of Biological
Sciences, Purdue University, West Lafayette, Indiana5; and Department
of Microbiology, Medical School, University of Minnesota, Minneapolis, Minnesota2
Received 17 November 1994/Accepted 4 April 1995
Two adjacent papainlike cysteine protease (PCP) domains, PCPa and PCPb, were identified in the
N-terminal region of the open reading frame 1a replicase proteins of the arteriviruses porcine reproductive and respiratory syndrome virus and lactate dehydrogenase-elevating virus. The replicase of the related virus equine arteritis virus contains only one active PCP in the corresponding region. Sequence comparison revealed that
the equine arteritis virus PCPacounterpart probably was inactivated by loss of its catalytic Cys residue. For
both porcine reproductive and respiratory syndrome virus and lactate dehydrogenase-elevating virus, the
generation of two processing products, nsp1a and nsp1b, was demonstrated by in vitro translation.
Site-directed mutagenesis and sequence comparison were used to identify the putative active-site residues of the
PCPa and PCPb protease domains and to show that they mediate the nsp1a/1b and nsp1b/2 cleavages,
respectively.
Equine arteritis virus (EAV), lactate dehydrogenase-elevat-ing virus (LDV), porcine reproductive and respiratory syn-drome virus (PRRSV), and simian hemorrhagic fever virus constitute a recently recognized group of enveloped positive-strand RNA viruses, the arteriviruses (see reference 25 for a review). The genomic sequences of the EAV Bucyrus strain (5), the LDV C strain (8), the LDV P strain (24), and the Lelystad strain of PRRSV (Lelystad virus; 22) have been de-termined. Comparative sequence analysis and the presence of striking similarities in genome organization and expression previously revealed an evolutionary relationship between ar-teriviruses and coronaviruses (5, 28).
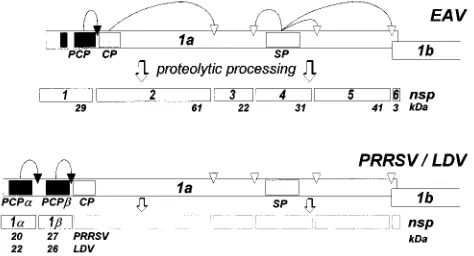
The arterivirus replicase gene is composed of two open reading frames (ORFs). ORF1a is translated directly from the genomic RNA, whereas ORF1b can be expressed only by ri-bosomal frameshifting, yielding a 1ab fusion protein (5). Both replicase gene products are multidomain precursor proteins which are proteolytically processed. The currently available EAV ORF1a protein processing scheme comprises the gener-ation of nonstructural protein 1 (nsp1) to nsp6 (30; see Fig. 5). In contrast to the C-terminal three-fourths of the arterivirus ORF1ab protein, its N-terminal part is highly variable, both in size and in sequence. A papainlike cysteine protease (PCP) in EAV nsp1 has been shown to autoproteolytically cleave the nsp1/2 site (29, 30). Sequence alignments and mutagenesis
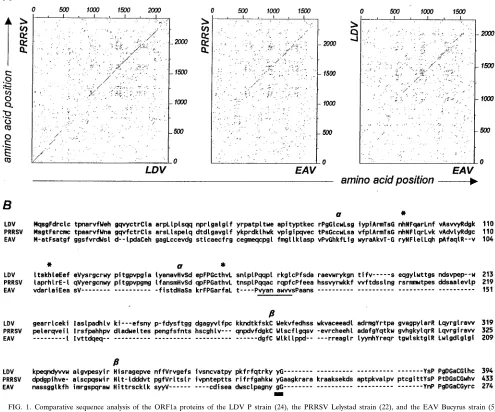
studies identified Cys-164 and His-230 as its catalytic residues (29). Typical for most PCPs, the catalytic residue Cys-164 is flanked at the C-terminal side by Trp, a bulky, hydrophobic amino acid (12, 26). The PRRSV and LDV ORF1a proteins were previously predicted to contain two PCP domains in their N-terminal region (3, 8, 22; Fig. 1), to which we will refer as PCPa(N terminal) and PCPb(C terminal). Godeny et al. (8) have suggested that PCPais the equivalent of the EAV nsp1 autoprotease. In their alignment, the PRRSV-LDV PCPb do-main has no equivalent in EAV. However, we have performed an independent comparative analysis of the EAV, PRRSV, and LDV ORF1a proteins and propose an essentially similar nsp1 organization for each of the three arteriviruses. The PCPadomain of PRRSV and LDV was found to be similar to an inactivated PCP motif in the N-terminal domain of EAV nsp1, and their PCPb domain could be aligned with the pre-viously described EAV PCP domain (Fig. 1).
PRRSV Cys-76 is part of a conserved Cys-Trp dipeptide and was predicted to be the PCPacatalytic Cys residue (Fig. 1B). Four conserved His residues were identified which could be the second member of the PCPa catalytic dyad: 92, His-115, His-146, and His-157. However, His-92 and His-115 are extremely close to the putative catalytic residue Cys-76. The distance between Cys-76 and His-146 or His-157 is much more similar to those reported for other viral PCPs (12, 29). The putative PCPacatalytic Cys has not been conserved in EAV, and this has probably rendered the protease inactive. However, sequence similarity to the region immediately downstream of PRRSV Cys-76 is still present (Fig. 1B). Furthermore, PRRSV His-146 is part of a small domain (His-X-Ser-X-X-X-Phe-Pro-Gly) which is also conserved in EAV (His-122; Fig. 1B).
Iden-* Corresponding author. Mailing address: Department of Virology, Institute of Medical Microbiology, Leiden University, Postbus 320, 2300 AH Leiden, The Netherlands. Phone: 71261657. Fax: 31-71263645. Electronic mail address: [email protected].
4500
on November 9, 2019 by guest
http://jvi.asm.org/
tification of putative active-site residues in PRRSV PCPbwas facilitated by alignment with the EAV nsp1 protease and the LDV sequence (Fig. 1B): Cys-276 and His-345 are conserved and align very well with the EAV PCP catalytic residues (29). Like EAV Cys-164, PRRSV Cys-276 is flanked by a Trp resi-due. His-345 is the only conserved His located between Cys-276 and the predicted nsp1b/2 cleavage site (Fig. 1B). The N-terminal 385 amino acids of the LDV and PRRSV ORF1a proteins are very similar (48% amino acid identity; Fig. 1), and all putative PRRSV catalytic residues were found to be con-served in LDV. We therefore assumed that LDV also contains two PCP domains and that the active-site residues are Cys-76 and His-147 for PCPaand Cys-269 and His-340 for PCPb. In conclusion, we expected the N-terminal regions of the PRRSV and LDV ORF1a polyproteins to be cleaved twice by two
[image:2.612.58.553.81.493.2]separate proteases, in contrast to the single autoproteolytic cleavage which occurs in the homologous region of EAV (29). To test whether the N-terminal parts of the ORF1a proteins of PRRSV and LDV are indeed processed in more than one place, two sets of 39-truncated ORF1a expression constructs were created (Fig. 2 and 3). Plasmid pIRES, a pBS2 (Strat-agene) derivative carrying a T7 promoter, a copy of the en-cephalomyocarditis virus internal ribosomal entry site (16), and a number of downstream restriction sites, served as a vector. Viral sequences were fused to the encephalomyocardi-tis virus internal ribosomal entry site by a 12-nucleotide linker encoding the N-terminal extension Met-Ala-Thr-Thr. Termi-nation codons for translation and appropriate linearization sites were introduced downstream of the viral sequences. The PRRSV and LDV expression vectors were used in in vitro
FIG. 1. Comparative sequence analysis of the ORF1a proteins of the LDV P strain (24), the PRRSV Lelystad strain (22), and the EAV Bucyrus strain (5) (EMBL-GenBank database accession numbers U15146, M96262, and X53549, respectively). (A) Proportional dot matrix comparison generated with the COMPARE option of the University of Wisconsin Genetics Computer Group software (6) with a window size of 21 and a stringency of 11. (B) Multiple sequence alignment of the N-terminal region of arterivirus ORF1a proteins, generated with the OPTAL program (9) with the PAM250 scoring table (4) and the CLUSTALV program (15) with the PAM250 and BLOSSUM62 tables (13). Tentative borders of conserved domains were determined through pairwise sequence comparisons in the dot plot fashion, by utilizing the high-resolution DotHelix program (21) in conjunction with the PAM250 and BLOSSUM62 tables. Conserved amino acids are capitalized. Gaps introduced during alignment are indicated by dashes. Active-site residues of PCPaand PCPbare indicated by the lettersaandb, respectively. Alternative His residues which were tested during PCPamutagenesis are marked with asterisks. The region estimated to contain the PRRSV and LDV nsp1a/1bcleavage site is underlined, and the EAV nsp1/2 cleavage site (29) and the predicted PRRSV and LDV nsp1b/2 cleavage sites are marked with a bar.
on November 9, 2019 by guest
http://jvi.asm.org/
translation assays to express increasing N-terminal segments of the ORF1a protein (Fig. 2 and 3).
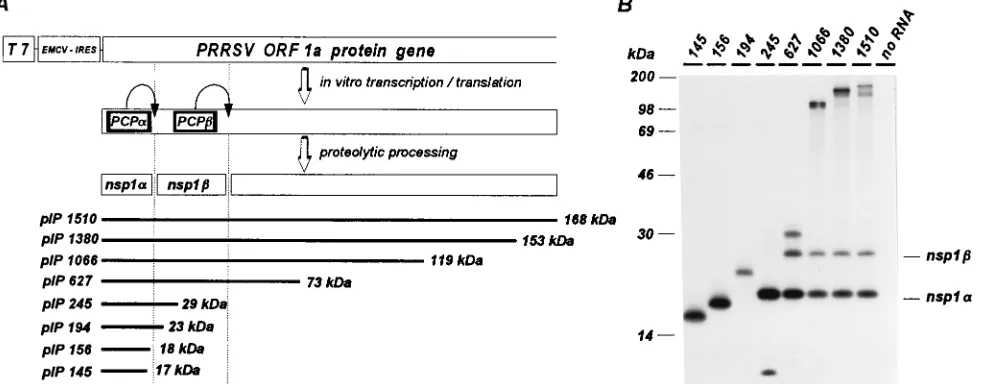
For PRRSV, proteolytic processing was not observed for the translation products derived from the three shortest constructs (Fig. 2), but an approximately 20-kDa N-terminal cleavage product was generated from all larger plasmids and was named nsp1a. Since construct pIP245 (Fig. 2A) did not contain the PCPbdomain, we assumed that this cleavage was mediated by the proteolytic activity of PCPa. The translation products of pIP627 and all larger plasmids contained both predicted pro-tease domains. As expected, a second cleavage product of approximately 27 kDa (nsp1b) was observed (Fig. 2B). This demonstrated that the N-terminal ORF1a domain of PRRSV, indeed, contains an additional cleavage site, in comparison with EAV. Similar results were obtained with the LDV con-structs (Fig. 3). Most of the pIL212 product, which contained only the PCPa domain, remained unprocessed. However, a small amount of an approximately 22-kDa N-terminal polypep-tide (nsp1a) was produced. This protein was generated much more efficiently after translation of pIL414 and pIL549. The latter two constructs, which contained both the PCPa and PCPbdomains, produced an additional 26-kDa cleavage prod-uct which was concluded to be LDV nsp1b. The C-terminal product of pIL414 (estimated size, only 4 kDa) could not be observed because of the absence of methionine residues, but that of pIL549 was seen as a protein with the predicted size of 19 kDa.
To test whether the two cleavages are mediated by separate proteolytic activities and to probe the predicted active-site residues, amino acid substitutions were introduced into PRRSV construct pIP627 (Fig. 4). When Cys-76 was changed for either Gly or Ser, the nsp1a/1b site was no longer pro-cessed: only a 45-kDa nsp1ab precursor and the 30-kDa C-terminal pIP627 product were seen. Replacement of His-92 with Phe or Tyr did not affect cleavage. This was also true for the His-1153Ala substitution, whereas the nsp1a/1bsite of a His-1153Tyr mutant was found to be cleaved with approxi-mately 50% efficiency. Five substitutions (Asp, Phe, Ile, Asn, and Tyr) were tested at the His-146 position, and they all completely abolished cleavage at the nsp1a/1bsite.
Replace-ment of His-157 produced variable results. The Tyr-157 mu-tant cleaved with wild-type efficiency, but the His-1573Ile mutation was only 10% active. Finally, the Asn-157 and Asp-157 mutants were also partially functional proteases (80 and 50% cleavage, respectively). Thus, of all of the mutations of conserved His residues, only the five His-146 substitutions completely inactivated the nsp1a/1bcleavage, although some of the other replacements affected it. To analyze the putative PRRSV PCPb active-site residues, Cys-276 was replaced by Ile, Leu, Arg, or Ser and His-345 was replaced by Asp or Tyr. All of these substitutions abolished processing of the nsp1b/2 site, and only nsp1a and a 53-kDa precursor (consisting of nsp1band the 30-kDa C-terminal pIP627 product) were ob-served (Fig. 4). To confirm that no other proteolytic activity was present in the pIP627 translation product, a double mutant containing two inactivated PCP domains was shown to produce a single precursor protein of the expected size. Our experimen-tal data on PRRSV show that (i) PCPaand PCPbare papa-inlike cysteine proteases mediating cleavage of the nsp1a/1b and nsp1b/2 junctions, respectively, and (ii) Cys-76–His-146 and Cys-276–His-345 most likely form the catalytic dyads of PCPaand PCPb, respectively.
For the LDV PCPs, the predicted catalytic Cys residues were also tested experimentally. Two mutations were introduced into construct pIL549 (Fig. 3). A Cys-763Ser substitution completely inhibited cleavage of the nsp1a/1bsite, whereas the nsp1b/2 junction was no longer processed when Cys-269 was replaced with Ala. In view of the high percentage of sequence identity between LDV and PRRSV (Fig. 2), the catalytic His residues of the two LDV proteases (His-147 and His-340, re-spectively) can be predicted confidently.
[image:3.612.65.557.75.267.2]The EAV PCP does not cleave in trans, a conclusion based on the results obtained with mixed in vitro translation reactions in which the proteases and substrates required for a trans-cleavage reaction were produced simultaneously (29). By using a similar approach, we were unable to detect any trans-cleav-age activity of PRRSV PCPaand PCPb(data not shown). On the basis of the size of the nsp1aprotein, the PRRSV nsp1a/1b cleavage site should be located near residues 164 to 168 (Pro-Leu-Pro-Gln-Gln), which are absolutely conserved in PRRSV
FIG. 2. Identification of two proteolytic cleavages in the N-terminal domain of the PRRSV ORF1a protein. (A) Schematic representation of the N-terminal part of the ORF1a protein and the set of truncated cDNA constructs. (B) In vitro translation of transcripts derived from the cDNA constructs shown in panel A. Translation products were generated in a rabbit reticulocyte lysate in the presence of [35S]methionine and analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(19). ECMV-IRES, encephalomyocarditis virus internal ribosomal entry site.
4502 NOTES J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
and LDV (Fig. 1B). However, the observed sodium dodecyl sulfate-polyacrylamide gel electrophoresis migration difference between the two nsp1a species indicates that their cleavage sites may not have been aligned in our sequence comparison. The EAV nsp1 PCP cleaves between two Gly residues at po-sitions 260 and 261 (29). Since the N-terminal domain of nsp2 is highly conserved (Fig. 1), the regions which should contain the PRRSV-LDV nsp1b/2 junctions could easily be aligned, the only complication being a 35-amino-acid insertion in the PRRSV sequence. The sizes observed for the PRRSV and LDV nsp1a, nsp1b, and nsp1abproteins (Fig. 2–4) and our sequence alignment are most consistent with cleavage between Tyr and Gly (positions 384 and 385 for PRRSV and 380 and 381 for LDV).
The arterivirus PCPs belong to a rapidly expanding group of
[image:4.612.78.343.70.471.2]viral proteases assumed to be related to the cellular papainlike thiol proteases (7, 12). Most viral PCPs, including the five arterivirus PCPs which have been identified, are so-called leader proteases which direct a single cleavage in the N-termi-nal region of a precursor (10, 12). Papainlike leader proteases
FIG. 3. Identification of two proteolytic cleavages in the N-terminal domain of the LDV ORF1a protein. (A) Schematic representation of the N-terminal part of the LDV ORF1a protein and the set of truncated cDNA constructs used for in vitro transcription and translation studies. (B) In vitro translation of tran-scripts derived from the cDNA constructs shown in panel A. In addition, pIL549 protein mutants containing a Cys-763Ser or a Cys-2693Ala substitution and a double mutant protein were expressed. ECMV-IRES, encephalomyocarditis vi-rus internal ribosomal entry site. The positions of the nsp1aand nsp1bcleavage products, the C-terminal residual cleavage product (C-term.), and the nsp1ab precursor protein are indicated on the right.
FIG. 4. Mutagenesis of putative PRRSV PCPaand PCPbactive-site resi-dues. Nucleotide changes were introduced by oligonucleotide-directed mutagen-esis as described by Kunkel et al. (18). (A) Schematic representation of cDNA construct pIP627, which was used to test the effect of amino acid substitutions on the proteolytic activity of PCPa(B) and PCPb(C). (B) Expression of PCPa mutants. Translation of the wild-type pIP627 protein is shown in the leftmost lane. The amino acid substitution tested is indicated at the top of each lane. The positions of the nsp1a and nsp1bcleavage products, the C-terminal residual cleavage product (C-term.), and the nsp1abprecursor protein are indicated on the right. (C) Expression of PCPbmutants. A PCPa-PCPb double mutant containing the Cys-763Ser and Cys-2763Ile mutations was also expressed. ECMV-IRES, encephalomyocarditis virus internal ribosomal entry site.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.300.549.80.552.2]have been characterized and/or predicted for (among others) potyviruses (23), closteroviruses (1), aphthoviruses (12, 17), and the hypovirulence-associated virus of the chestnut blight fungus (27). The presence of one or two PCPs in the N-terminal half of the ORF1a protein has also been predicted for coronaviruses (2, 14, 20).
Our combined information allows us to present a prelimi-nary processing scheme for the ORF1a proteins of EAV, PRRSV, and LDV (Fig. 5). Like the EAV nsp1 protease, the PCPaand PCPb domains of PRRSV and LDV are likely to mediate an autoproteolytic cleavage at their own C terminus only. Two additional proteolytic activities have recently been identified in the EAV ORF1a protein: a cysteine autoprotease in the N-terminal domain of nsp2 (31) and a chymotrypsinlike serine protease in nsp4 (32). Highly conserved PRRSV-LDV counterparts of both of these EAV proteases were identified by sequence comparison, suggesting that the PRRSV and LDV ORF1a proteins contain four proteolytic activities (Fig. 5).
Both arteriviruses and coronaviruses contain the same set of conserved replicase domains: papainlike protease(s)–chymot-rypsinlike protease–polymerase–putative metal-binding do-main–helicase–conserved C-terminal ORF1b domain (2, 5, 8, 11, 14, 22, 28). The C-terminal three-fourths of the ORF1ab polyprotein appears to form the core of this coronaviruslike replicase (28), which is likely to be processed by the chymot-rypsinlike protease. The N-terminal half of the ORF1a protein is variable, suggesting that species-specific, rather than group-specific, functions have been coupled to the set of basic func-tions provided by the core replicase. The papainlike protease-directed processing of the N-terminal replicase domain into relatively unique nonstructural proteins appears to be a com-mon property of viruses that belong to the coronaviruslike superfamily.
A.E.G. is most grateful to Michael Rossmann for hospitality during his stay at Purdue University. We thank Willy Spaan for helpful dis-cussions and reading of the manuscript.
The work of A.E.G. was supported by a travel grant from the Netherlands Organisation for Scientific Research (NWO), a Cooper-ation in Applied Science and Technology award to A.E.G. and Michael Rossmann, a grant from the National Science Foundation to Michael Rossmann, and grants from the International Science Foundation. K.S.F. was supported by USPHS training grant 09138.
REFERENCES
1. Agranovsky, A. A., E. V. Koonin, V. P. Boyko, E. Maiss, R. Frotschl, N. A. Lunina, and J. G. Atabekov. 1994. Beet yellows closterovirus—complete
genome structure and identification of a leader papainlike thiol protease. Virology 198:311–324.
2. Baker, S. C., K. Yokomori, S. Dong, R. Carlisle, A. E. Gorbalenya, E. V. Koonin, and M. M. C. Lai.1993. Identification of the catalytic sites of a papainlike cysteine proteinase of murine coronavirus. J. Virol. 67:6056–6063. 3. Chen, Z., L. Kuo, R. R. R. Rowland, C. Even, K. S. Faaberg, and P. G. W. Plagemann.1993. Sequence of 39-end of genome and of 59-end of ORF 1a of lactate dehydrogenase-elevating virus (LDV) and common junction mo-tifs between 59-leader and bodies of seven subgenomic mRNAs. J. Gen. Virol. 74:643–660.
4. Dayhoff, M. O., R. M. Schwartz, and B. C. Orcutt. 1978. A model of evolu-tionary change in proteins, p. 345–352. In M. O. Dayhoff (ed.), Atlas of protein sequence and structure, vol. 5. National Biomedical Research Foun-dation, Washington, D.C.
5. Den Boon, J. A., E. J. Snijder, E. D. Chirnside, A. A. F. de Vries, M. C. Horzinek, and W. J. M. Spaan.1991. Equine arteritis virus is not a togavirus but belongs to the coronaviruslike superfamily. J. Virol. 65:2910–2920. 6. Devereux, J., P. Haeberli, and O. Smithies. 1984. A comprehensive set of
sequence analysis programs for the VAX. Nucleic Acids Res. 12:387–395. 7. Dougherty, W. G., and B. L. Semler. 1993. Expression of virus-encoded
proteinases: functional and structural similarities with cellular enzymes. Mi-crobiol. Rev. 57:781–822.
8. Godeny, E. K., L. Chen, S. N. Kumar, S. L. Methven, E. V. Koonin, and M. A. Brinton.1993. Complete genomic sequence and phylogenetic analysis of the lactate dehydrogenase-elevating virus. Virology 194:585–596.
9. Gorbalenya, A. E., V. M. Blinov, A. P. Donchenko, and E. V. Koonin. 1989. An NTP-binding motif is the most conserved sequence in a highly diverged monophyletic group of proteins involved in positive strand RNA viral rep-lication. J. Mol. Evol. 28:256–268.
10. Gorbalenya, A. E., and E. V. Koonin. 1993. Comparative analysis of the amino acid sequences of the key enzymes of the replication and expression of positive-strand RNA viruses. Validity of the approach and functional and evolutionary implications. Sov. Sci. Rev. Sect. D Biol. Rev. 11:1–84. 11. Gorbalenya, A. E., E. V. Koonin, A. P. Donchenko, and V. M. Blinov. 1989.
Coronavirus genome: prediction of putative functional domains in the non-structural polyprotein by comparative amino acid sequence analysis. Nucleic Acids Res. 17:4847–4861.
12. Gorbalenya, A. E., E. V. Koonin, and M. M. C. Lai. 1991. Putative papain-related thiol proteases of positive-strand RNA viruses. FEBS Lett. 288:201– 205.
13. Henikoff, S., and J. G. Henikoff. 1992. Amino acid substitution matrices from protein blocks. Proc. Natl. Acad. Sci. USA 89:10915–10919.
14. Herold, J., T. Raabe, B. Schelleprinz, and S. G. Siddell. 1993. Nucleotide sequence of the human coronavirus 229E RNA polymerase locus. Virology 195:680–691.
15. Higgins, D. G., A. J. Bleasby, and R. Fuchs. 1992. CLUSTAL V: improved software for multiple sequence alignment. Comput. Appl. Biosci. 8:189–191. 16. Jang, S. K., H.-G. Kra¨usslich, M. J. H. Nicklin, G. M. Duke, A. C. Palmen-berg, and E. Wimmer.1988. A segment of the 59nontranslated region of encephalomyocarditis virus RNA directs internal entry of ribosomes during in vitro translation. J. Virol. 62:2636–2643.
17. Kleina, L. G., and M. J. Grubman. 1992. Antiviral effects of a thiol protease inhibitor on foot-and-mouth disease virus. J. Virol. 66:7168–7175. 18. Kunkel, T. A., J. D. Roberts, and R. Zakour. 1987. Rapid and efficient
site-directed mutagenesis without phenotypic selection. Methods Enzymol. 154:367–382.
19. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680–685.
20. Lee, H. J., C. K. Shieh, A. E. Gorbalenya, E. V. Koonin, N. la Monica, J. Tuler, A. Bagdzhadzhyan, and M. M. C. Lai.1991. The complete sequence (22 kilobases) of murine coronavirus gene 1 encoding the putative protease and RNA polymerase. Virology 180:567–582.
21. Leontovich, A. M., L. I. Brodsky, and A. E. Gorbalenya. 1993. Construction of the full local similarity map for two biopolymers. BioSystems 30:57–63. 22. Meulenberg, J. J. M., M. M. Hulst, E. J. de Meijer, P. L. J. M. Moonen, A.
den Besten, E. P. de Kluyver, G. Wensvoort, and R. J. M. Moormann.1993. Lelystad virus, the causative agent of porcine epidemic abortion and respi-ratory syndrome (PEARS), is related to LDV and EAV. Virology 192:62– 72.
23. Oh, C.-S., and J. C. Carrington. 1989. Identification of essential residues in potyviral proteinase HC-Pro by site-directed mutagenesis. Virology 173:692– 699.
24. Palmer, G. A., L. Kuo, Z. Chen, K. S. Faaberg, and P. G. W. Plagemann. Sequence of the genome of lactate dehydrogenase-elevating virus: hetero-genicity between strains P and C. Virology, in press.
25. Plagemann, P. G. W., and V. Moennig. 1991. Lactate dehydrogenase-elevating virus, equine arteritis virus, and simian haemorrhagic fever virus: a new group of positive-stranded RNA viruses. Adv. Virus Res. 41:99–192.
26. Polgar, L., and P. Halasz. 1982. Current problems in mechanistic studies of serine and cysteine proteinase. Biochem. J. 207:1–10.
27. Shapira, R., and D. L. Nuss. 1991. Gene expression by a hypovirulence-FIG. 5. Preliminary proteolytic processing schemes of the EAV, LDV, and
PRRSV replicase ORF1a proteins. Indicated are the positions of the protease domains described in this report and elsewhere (29–31) and the approximate locations of their cleavage sites. The end products of proteolytic processing and their sizes are indicated. CP, cysteine protease. SP, serine protease.
4504 NOTES J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.61.298.73.201.2]associated virus of the chestnut blight fungus involves two papainlike pro-tease activities. Essential residues and cleavage site requirements for p48 autoproteolysis. J. Biol. Chem. 266:19419–19425.
28. Snijder, E. J., and M. C. Horzinek. 1993. Toroviruses: replication, evolution and comparison with other members of the coronaviruslike superfamily. J. Gen. Virol. 74:2305–2316.
29. Snijder, E. J., A. L. M. Wassenaar, and W. J. M. Spaan. 1992. The 59end of the equine arteritis virus replicase gene encodes a papainlike cysteine pro-tease. J. Virol. 66:7040–7048.
30. Snijder, E. J., A. L. M. Wassenaar, and W. J. M. Spaan. 1994. Proteolytic processing of the replicase ORF1a protein of equine arteritis virus. J. Virol. 68:5755–5764.
31. Snijder, E. J., A. L. M. Wassenaar, W. J. M. Spaan, and A. E. Gorbalenya. The arterivirus nsp2 protease: an unusual cysteine protease with primary structure similarities to both papainlike and chymotrypsinlike proteases. J. Biol. Chem., in press.
32. Snijder, E. J., A. L. M. Wassenaar, L. C. van Dinten, A. E. Gorbalenya, and W. J. M. Spaan.Unpublished data.