0022-538X/78/0025-0824$02.00/0
Copyright01978 AmericanSocietyforMicrobiology Printed in U.S.A.
Characterization of the
5'-Terminal
Capped Structures of Late
Simian Virus 40-Specific
mRNA
G. HAEGEMAN AND W. FIERS*
LaboratoryofMolecular Biology, State University Ghent, 9000 Ghent, Belgium Received for publication25October 1977
32P-labeled,latesimian virus40-specificRNA was isolated frominfected CV1
cellsandcompletely degraded with RNaseT2 and bacterial alkaline phosphatase.
The RNase-resistant materialwas fractionated two dimensionally and further
characterized withPenicillium nuclease andnucleotide pyrophosphatase. Two
major 5' terminiwere identified in latesiirianvirus 40RNA, namely, 7-methyl
Gppp2',6-dimethyl ApUp and 7-methyl Gppp2',6-dimethylAp2'-methylUpUp.
Both 5' terminiarepresentinunfractionated viral RNA as well as in theseparated
16S and 19S species. Asboth caps differ onlyin secondary modification, it is
possiblethat theyarederived from the same site onthe DNA. The relatively
higher cap II content of the 16S mRNA may be related to its slowerrate of turnover.
Blocked5'terminiareageneralcharacteristic of eucaryotic cellular mRNA's (21, 24). Also,
mostof theeucaryoticviral mRNA's studied so
far are blocked attheir 5' end (24). The latter
contain exclusively a purine nucleotide in the
5'-penultimate position, whereas cellular mRNA'smay have eitherapurine or a
pyrimi-dine. This penultimate nucleotide can be
un-modified (cap 0structure), but in many cases
itappearstobemethylatedon theribose, and
in afew examplesanadditional base
methyla-tion alsooccursinthisposition (capIstructure).
Ribosemethylation of the second nucleotide is
oftenobserved in the mRNA's fromhigher
or-ganisms and the corresponding viruses, thus
leadingtothe formation of capII structures. It
is noteworthy that the methylation pattern of
the caps increases as one movesup the
evolu-tionaryladder.
Simian virus40 (SV40) isaDNA-containing
virus, whichproduces
virus-specific
RNA in thecytoplasm oflyticallyinfected monkeykidney cells(3, 6, 17, 27, 31).Studies on themethylation
pattern of SV40-specific RNA revealed
5'-ter-minal ends of the type7mGpppNm(13). Because
of the general interest in SV40, we have
char-acterized in more detail the 5' termini of the
predominantcytoplmmicviralRNAs, and their
structuresarereportedhere.
MATERIALS AND METHODS
Preparationof32P-labeledlate SV40RNA.CV1
monkey cells (Bio-Cult, Paisley, Scotland) were cul-tured inplastic petridishes(Falcon Plastics,Oxnard,
Calif.) andinfected with60PFU ofSV40(strain776)
percell. At24hafterinfection,thegrowingmedium
waschangedto alow-phosphate medium (containing
10'- M phosphate, i.e., 1% of the normal concentra-tion),and3mCi of3Pradioactivity (The Radiochem-ical Centre Ltd., Amersham, England) was added per 14-cm petri dish for alabeling period of 24h. The mediumwasthenremoved,and the ceUs were washed afew times withphosphate-bufferedsaline and finally lysedwith1% Nonidet P40. The nuclei were removed by centrifugation, and subsequently the lysate was subjected toseveral extractions with
phenol-chloro-form-isoamylalcohol.The extracted RNA was
precip-itatedatleast twice with ethanoltominimize contam-ination withco-precipitating[tP]phosphate, originat-ing from thepoolpresent inthe cellularcytoplasm. Isolation of polyadenylic acid [poly(A)]-containing RNAonoligodeoxythymidylicacid-cellulose (P-L
Bio-chemicals, Inc., Milwaukee, Wis.) was followed by
selection of theSV40-specific sequencesby hybridi-zationonSV40DNAlinkedtoSepharose4B (Phar-macia FineChemicals, Inc.,Piscataway, N.J.) (9). The SV40mRNA thuspurified usuallyrepresented around 0.2%of the totalcytoplasmicRNAfraction.
Preparative gel fractionation of 32P-labeled
RNA. Insome casesthe RNApreparation was frac-tionatedbypreparative electrophoresison anagarose slabgel (160 by120by3mm). This stepwas carried outeither before (seeFig. 4)orafter the selection by hybridization. A solution of 1.4% agarose (Sigma Chemical Co.,St.Louis, Mo.) in running buffer (0.02 MTris-acetateplus 1 mM EDTA, pH 7.5) was used. The RNAsamplesweredissolvedinrunning buffer, which had beenmixed with 50% deionized formamide, heated for 1 minat 70°C, and loadedonto the gel. The electrophoresis wasstarted immediately inthe coldat50 to 100V forabout2 h.Next, thegelwas
dismantled, and the 3P-labeled RNAwas localized by autoradiography. The bands were cut out, and theirradioactivitywasestimatedbyCerenkov count-ing. The material was recovered from squeezed gel bandsby several extractions withsmall quantities (i.e., 824
on November 10, 2019 by guest
http://jvi.asm.org/
100 to 200
pl
foragelvolume of 20by5by 3 mm) of 0.5MNaCl,containing5mMEDTA and 0.5% sodium dodecylsulfate. The addition ofafewdropsofphenol may facilitate the releaseof RNA from the agarose pieces.Finally, the 3P-labeled RNA was precipitated at-20°C togetherwith carrier yeastRNA,by adding two partsofethanol to the combined extracts. The recoveryof the elutionprocedureamounts to70%for RNA from the28Sregionandreachesmorethan 80% for material of smaller S values.Isolation of 5'-terminal structures. For degra-dation with RNase T2, the pelleted RNA (i.e., 32P-labeled viral orcellular mRNA and 40 ug of carrier yeastrRNA)wasdissolved inavolatile buffer (0.1 M formicacid-pyridine, pH 4.5) anddigestedovernight at37°Cwith 5 U of T2 enzyme(SigmaChemicalCo.). Next, thehydrolysatewasevaporatedtodryness, and the residuewasdissolved in 0.1 M ammonium hydro-gencarbonate containing0.5U of bacterial alkaline phosphatase(WorthingtonBiochemicals Corp., Free-hold, N.J.) forafurther incubation ofafewhoursat 37°C.Thereaction mixturewasevaporatedagain and washedseveral times with distilledwater toremove remainingtracesof salt.
Theseparation procedureinvolvedaclassical elec-trophoresisoncelluloseacetateinthe firstdimension (23), followed by ion-exchange chromatography on thin-layer plates in the second dimension. The T2 hydrolysate,dissolved inaminimalvolume of water, was applied as a thin line on prewetted cellulose acetatestrips(Schleicherand SchiillGMBH, Dassel, WestGermany) andsubjected toelectrophoresis at pH3.5in the presence of5Mureauntil thepink dye hadmigrated 20 to 25cm. The material was trans-ferredonto athin-layer chromatographic plateof pol-yethyleneimine (PEI)-cellulose (Machery-Nagel and Co., Duren,WestGermany)bythe inversedblotting technique(26)anddevelopedinthe seconddimension byascending chromatographywith avolatile buffer solution of1 M (or1.3M) formic acid-pyridine (pH 4.3).Radioactive spotswerevisualizedby autoradiog-raphy at -70°C, using preflashed X-ray films (RX Fuji X-Ray Film) (12) together with intensifying * screens (CAWO, Schrobenhaussen, West Germany) (R.Laskey and A.Mills, FEBS Lett.,in press). The nucleotide materialcanbe recoveredeasily from thin-layerchromatographyplates bythemicroelution tech-niquedescribedpreviously(28).
Analysisofcappedtermini. RNase T2-resistant material was further degraded by Penicillium (P1) nuclease (P-L Biochemicals, Inc.). The dry residue, after elution from thePEIplate,wasredissolved ina volatile buffer(0.1Mformicacid-pyridine, pH6), and 10AgofPI enzymewasadded. The reaction mixture wasincubated for 30 minat37°Cand thenevaporated to dryness. The materialwasthoroughly washed by the addition ofasmall volume ofwaterfollowedby dryingandthenappliedontominiplates of PEI-cellu-lose (6.6 by9.9cm) fortwo-dimensional chromatog-raphy: the first dimensionwasdeveloped witha20% formicacidsolution;inthe seconddirection,abuffer system of1 Mformicacid-pyridine(pH 4.3)wasused (27a).
Cap structures,resistanttoP1hydrolysis,were an-alyzed by digestionwith nucleotidepyrophosphatase (SigmaChemicalCo.).Thedriedmaterialwaswashed
withwater to removeremaining traces of triethyla-mine carbonate. After it was redissolved in the appro-priateenzymesolution(10U ofnucleotide pyrophos-phatase per ml of 25 mM Tris-chloride-2 mM
Mg2e,
pH 7.5), the pH was checked andeventuallyadjusted. The mixturewasincubated for 30 min at 37°C, and thedigestwastransferreddirectly onto miniplates of PEI-cellulose(6.6 by9.9cm) fortwo-dimensional sep-aration. A less concentratedsolution (e.g., 0.5 M formic acid-pyridine, pH 4.3, in the second dimension) was usedtoobtain agoodresolution between singly and doubly methylated nucleotides. Methylated nucleo-tides used as references were purchased from P-L Biochemicals,Inc.RESULTS
RNase T2 hydrolysis oftotal late SV40
RNA. 3P-labeled late SV40 mRNA was
pre-pared and completelydegradedwith RNase T2
andbacterial alkalinephosphatase,as described
above. A typical degradation pattern of SV40
RNA is shown in Fig. 1. Twospots are promi-nent, products 1 and 2, corresponding to two
different 5' termini in the late viral RNA.
Inor-ganic [32P]phosphate was run off the plate in
the firstdimension,andonlysometrailing
radio-activity from this giant spot isseen. The dark
spots at the topof the figure are cyclic
mono-nucleotides; these intermediates are due to
in-completeRNaseT2 hydrolysisandareresistant
tothephosphatase.
Theaforementionedproducts1and2contain
the7-methyl Gppp group, aswillbe shown
be-low;sugar(2') methylation ofthe following
res-idue(s) renders these resistant tothe action of
RNaseT2 and generatesthe longer 5'-terminal
oligonucleotides (1, 19, 32). That the caps are
derived from "late" SV40 mRNA follows froma
characterization of the cytoplasmic,
poly(A)-containing, SV40-specificRNAbyfingerprinting
[G. Haegeman and W. Fiers, Nature (London),
in press] and is in agreement with the
well-known abundance of late RNA relative to
"early" (3). Also, someminor spots maybe
ob-served in Fig. 1; they are probably not virus
specified, but they are derived from a small
contaminationwithcellularRNA.Indeed,in less
purified preparations ofSV40RNA, the entire
pattern of cellularcapstructures wasstillvisible,
but as the viral RNA was more rigorously
se-lectedby stringenthybridizationconditions,the
two majorspots becamemore andmore
promi-nent while the overall cellular contamination
disappeared. Thestrongestcontaminants
corre-spond tothe maincellular cap structures (e.g.,
products 3 and 4 in Fig. 5), but even in this
examplethetwoviralproducts1and2represent
65 to 70% of the total fraction of T2-resistant
material, whereas the latterspotsamount to at most 20 to 25%of the entire cap populationin
on November 10, 2019 by guest
http://jvi.asm.org/
826
U
I Q
o 0
14tmbP
Elec. pH
3.5
FIG. 1. Two-dimensional map on PEI-cellulose of a combined RNase T2 andphosphatase degradation of total late32P-labeledSV40 mRNA. The ion-exchangechromatographyin thesecond dimensionwascarried outwith1.3Mformic acid-pyridine (pH 4.3). Thedye marker, xylenecyanol FF,denotedby B, migrated10.5 cmstartingfromthefirst-dimensional originline.
thecaseofatotalcellularmRNAhydrolysate.
Furthermore, thesameRNAmaterial,inwhich
products 1 and2 are the majorcap spots, has
beenfingerprintedafter RNaseT1digestionand
correspondsindeedtothe viral RNA,asrevealed byanalysisof all of the T1oligonucleotides
pres-ent and correlation with the known DNA
se-quence.
Furthermore, combined P1 hydrolysis and
phosphatase treatment of total cellular RNA
yieldstwodifferentcap structures, anA-cap and
a G-cap, both of which are present in nearly
equimolaramounts(unpublished data). On the
otherhand, similaranalysisofthehighly
puri-fied SV40 RNA showed almost exclusively an
A-capstructure.
Themolar yield of the 5'-terminalgroupwas
calculated from an analogous experiment, in
which theradioactivitypresentinthe
Pa-resist-antcap structureandrepresenting exactly
three
phosphateresidueswascomparedwiththe totalradioactivity of the viral RNA. Assuming an
average chainlengthofapproximately1,600
nu-cleotides for the mRNA molecules, the
5'-ter-minalstructuresappear in 0.5 molar ratio only.
Atpresent, we do notknow whether this
phe-nomenonis due to aninefficientcapping mech-anism or is the result ofpreferentiallosses of the 5'terminusduringtheisolationprocedure; how-ever, T1 oligonucleotides originating from the spliced 5'-terminal leader sequence appear in molarquantity (Haegeman and Fiers, in press).
Analysis of T2-resistant cap structures.
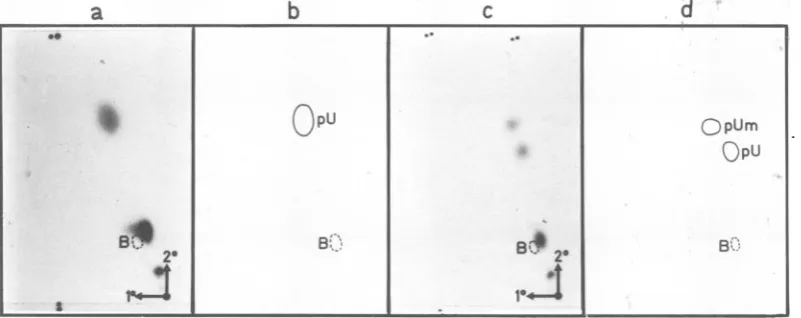
The T2-resistantproducts 1 and 2 were eluted
from the PEI plate and further characterized
by digestion with P1 nuclease (Fig. 2). Both
patternsshow a major componentcloseto the
bluedye andalesseramountofradioactivity in
the mononucleotide region; some undegraded
product remainsneartheapplication point. The
majorcompound,identical in both T2-generated
"caps," contains a
Pi-resistant
structure with the general formula 7mG(5')ppp(5')X. Asmen-tionedabove,complete
P1
hydrolysis of cellularRNA, followed by dephosphorylation, yields
mainlytwoproductsderived from the 5' termini,
namely,7-methyl GpppA and 7-methyl GpppG
(our unpublished data). The position of these
on the miniplates in the neighborhood of the
bluedyewasverifiedbyseveral control
experi-mentswith
32P-labeled
material and withoptical density (OD) references. In thecase ofthe twoSV40products (Fig. 2),the 5'terminus behaves
as anA-cap (G-caps are more delayed inboth
dimensions),buta morerefinedanalysis,aswell
asthe determination of the degree of
methyla-tion,isdescribed below.
Furthermore, enzymatic digestion of product
1 releasesa mononucleotide, which behaves in
this systemaspU. Indeed, carrier
mononucleo-tide pU, added to the reaction mixture as an
internalODmarker, chromatographsinexactly
thesameposition.Inthecaseofproduct2,two spots are visible in the mononucleotide region:
the slowestoneisagain pU,whereas the
faster-migratingspotmightbe the methylated analog
(it chromatographs like pU inthe first
dimen-sion, which separates mainly according to
charge) (27a). It actually coincides with
2'-O-methyluridine-5'-phosphate, added as an OD
If N
I %
I I
I
B
II
%11 -.1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.130.413.72.265.2]marker, but further confirmation was needed
since one cannotunambiguously determine from
*the mobility pattern in thissystemwhether the
methylationoccurs onthe base or onthe sugar
moiety ofthe nucleotide. Therefore, the
com-pound was rechromatographed on cellulose
platesin amixture ofisopropanol-anmonia-0.1
Mboric acid (7:1:2). Urnodifiedor
base-meth-ylatednucleoside-5'-phosphatesarestrongly
de-layedbecause of thecomplexing activityofboric
acid with thecis-diolgroups. The radioactivity
recovered was not retained at the origin, but
moved together with 2'-O-ribose-methylated
pU,thusproviding furtherproof for the
identi-fication(datanotshown). This analysis leadsto
thepartialstructure 7mGpppXmpUforproduct
1 and 7mGpppXmpUmpU for product 2, where
Xm indicates a ribose-methylated nucleotide,
since the residue to the 3' end of Xm is not releasedby RNaseT2 digestion.
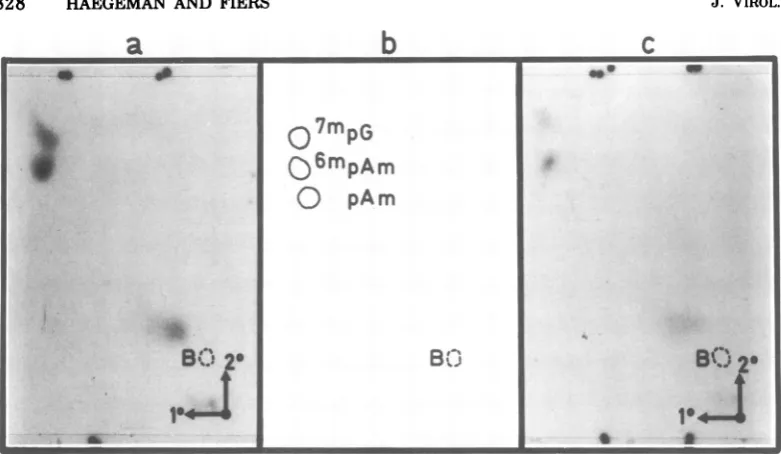
The identical Pi-resistant cap structures of
theT2 products 1 and2 (Fig. 2) were reeluted
from the PEIminiplate and independently
ana-lyzed bydigestion with nucleotide
pyrophospha-tase (Fig. 3). The experimentswere performed
in duplicate for each product, applying in one
2'-O-methyladenosine-5'-phosphate (pAm)as an
internal OD reference andinthe other
N6,2'-O-dimethyladenosmie-5'-phosphate (6mpAm).Both
sets offractionations showed an identical
pat-tern: inorganicphosphate (above the blue dye,
butmoving differentlyfrom undegraded P1-re-sistant caps), 7mpG (the fastest spot in both dimensions, also checked by an internal OD
reference), and the pA residue, in both cases
coincidingperfectlywith theinternal OD
refer-ence
'mpAm
andmovingprecisely onepositiona
b
fasterthan pAm (or6mpA),which inturnmoves
faster than pA. Although most of the capped
termini studied so far contain a
2'-O-ribose-methylated purinenucleotide, someviralRNAs
other than SV40 display a doubly methylated
A-residue in this position (11, 15, 16, 20, 25);
doubly methylated pG, however, has notbeen
observedsofar.
Thestructureof thetwoT2-resistant products (1and 2) isfully determinedby these analyses. They are 7mG(5')ppp(5')6mAmpU and
7mG-(5')ppp(5')6mAmpUmpU, respectively, and will
befurther indicatedby thetermscap Iandcap
II.Itisquite possible thatcapIIisderived from
cap Ibya secondmethylation step, a
phenom-enonthatalready hasbeenobserved in' thecase
of viral RNA (16, 20, 25) and appears rather
general even in cellular mRNA populations (4,
5, 7, 19, 29,32). However,asthelate viral SV40
RNA contains at least two RNA species (27,
31), we cannotexcludeapriorithat onetype of
cap structure maybe specifically derived from
one RNA species. This was checkedby
identi-fication of thecapsinfractionated SV40mRNA.
RNaseT2hydrolysisof fractionatedSV40
RNAspecies.Cytoplasmic, poly(A)-containing RNA wasisolatedfromSV40-infected cells and
fractionatedon a1.4%agarosegel. The
fraction-ation pattern was divided into a set of
banids,
from which the RNA material was eluted and
hybridized toSV40DNA. A distributionprofile
of the SV40-specific sequencesisshown inFig.
4a. Themajoritywaspresent in the 17Sto 18S
region andcorrespondstothemajor, late RNA
species,whichsedimentsin sucrosegradientsat
16S (30, 31). The 19S species is less prominent
and represents about25% of the 16S material.
[image:4.501.33.434.448.607.2]c
d
FIG. 2. Two-dimensionalchromatographyonminiplatesofPEI-celluloseofaP1 digest ofT2products 1
(a) and 2(c) and the corresponding diagrams, respectively, (b)and (d), showing the position of relevant referencemononucleotides(revealedunder UVlight).Theposition of theblue dye is indicatedbyadashed
circle.
so
* I
a)~pu
QpUm|BpU
a ~~~~* . 20 B'
on November 10, 2019 by guest
http://jvi.asm.org/
828 HAEGEMAN
a
1 ~ ~ ~b
cFIG. 3. Nucleotidepyrophosphatase analysis of Pi-resistantcapstructures,derived from products1and 2
(spotsnearthedyemarkerB inFig. 2). (a)and(c)showtherespective autoradiographs ofatwo-dimensional
separationonPEI-cellulose. (b)denotesthepositionofmethylatedreference nucleotides, whichwereadded asinternal markers indifferentcombinations and whichwerevisualized under UV light.Thetworadioactive spots, in (a)as well as in (c), correspond preciselytothe 7'pG and "'pAm internalreference compounds, respectively. Themonomethylated 6'pA and(2')pAm moveexactlythe samein thissystem. Bdenotes the position ofthe bluedye. ThespotthatmovesjustaheadofB in bothdimensionsisinorganic phosphate.
All of theSV40-specificfractionsweresubjected
tohydrolysis by RNase T2, followed by dephos-phorylation, and subsequently fractionated in two dimensions on PEI-cellulose plates. Both
cap I and cap II were present in all of the
fractionsanalyzed, and theyrepresented about 70%of thetotalamount ofT2-resistant products. As an example, the digestion patterns of the
18S and 19S SV40-specific material are
com-pared in Fig. 5. There seems to be relatively morecap IIin thematerial, whichruns at18S orfaster,ascompared with the 19SRNA. This is illustrated in Fig. 4b, in which the relative amountofcapIIisplotted for the different size classes; these results, however, are based on
countingverylowamountsofradioactivities and mustbe considered tentative.
DISCUSSION
The 5' termini of late poly(A)-containing
SV40-specificRNAhavebeenisolatedand
char-acterized in detail. It hasbeenclaimedthatPi RNasehydrolysisofSV40 mRNAreleases both 7mG(5')ppp 2'-O-methyladenosine and 7mG-(5')ppp2'-O-methylguanosine (13). Accordingto
ourdata, however,nosubstantialamountof
2'-O-methylguanosine containing capwaspresent in SV40 mRNA; furthermore, we have shown
thatthe modified adenosine residueis actually
NV,2'-O-dimethyladenosine.Infact,weconclude
thatonlyonemajorPI RNasecapispresentin late SV40mRNA, namely,7mG(5')ppp(5')SmAm.
Analysis with RNase T2 reveals two major types of cap structure, cap I and cap II, in cytoplasmic, late SV40 mRNA. Cap Iwas
iden-tifiedas7mG(5')ppp(5')6mAmpU,andcapIIwas
identified as7mG(5')ppp(5')(6mAmpUmpU. Both
types of 5' termini are also found among the wide variety of monkey cellular mRNA cap
structures (our unpublished data). It is quite possible that the 5' ends of both 16S and 19S
late mRNA'sarederived fromthesamesiteon
the genome, but differ only in the degree of secondarymodification. Themolecularbasis for suchahypothesiswasnotreadily evident until
veryrecently,asthe 16SRNA isbelievedtobe a processing product of the 19S molecule and
sharescommon sequences with the3'-terminal half of the 19Sspecies (2, 10, 30). Recent results, however, indicate a common5'-terminalleader
sequenceforthe two lateSV40mRNA's,which becomelinkedtotranscriptsfromanonadjacent
region on the genome (14, 22; Haegeman and
Fiers,inpress;S.Lavi andY.Groner,inpress). SimilartranslocationofRNAsegmentshasbeen showninadenovirusmRNA,and in thissystem thesharingofa commonleadersequenceiseven moreobvious (3a, 8,10a). Moreover,the locali-zationof theSV40capstructureintheHind C fragmentatposition0.72orcloser to the origin
s~~~~~
07mpG
*
Qo6mpAm
O
pAm
I
I*~::
8f
6 am U
-MF--m
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.64.455.60.287.2]0.4
[
0.3!-0.2
[
0.1 1
50
25
FIG. 4. (a) Separation of the total cytoplasmic poly(A)-containingRNAbyelectrophoresison a1.4% agarosegel,followedby selectionoftheSV40-specific sequencesby hybridization.Theposition ofthe rRNA markers is indicated. (b)The percent amountofcap II structure present in the totalpopulation (cap I plus capII)isplottedagainstthecorresponding gel fraction.
a
%
1I
B
I1
4
I 0 0
3
Elec. pH 3.5
1
do
2
wow
of replication (Haegeman and Fiers, in press;
Lavi and Groner, in press) suggests that the
putatively common 5'-terminal structure is
di-rectly derived from the 5' end of the primary
nucleartranscript. Indeed, aflow of the
meth-ylated cap structures from the heterogeneous
nuclear RNA tothe cytoplasmicmRNA
popu-lation has been observed inL-cells(18).
The relatively higher percentage of cap II
structure in the 16S mRNA mayberelated to
a difference inhalf-life,as aslower turnover of
the latter mRNA as compared with the 19S
species has been reported (2). In the case of
cellular mRNA's,it has in fact beenfound that
theformation ofcap IIstructures iscorrelated
with thelongevityof the mRNAmolecules (18).
Moreover, in the current view of "spliced"
mRNAmolecules, as is the caseforlate SV40 mRNA (see above), the relationship between the methylation of the cap structure and the
turnover of the RNA may perhaps be more complex. Indeed,ahigherrepresentation ofcap
IIin the 16S RNApopulationascompared with
the 19S may be either directly related to the
higherstability of the mRNA itselfor formed
byalongerexposuretomethylatingactivity of
the conserved 5'-terminal leader fragment,
which functions in the 19S mRNA and, after
cytoplasmic conversion (2), again in the 16S
mRNA. Alternatively, the 5'-leader sequence
containing thecapII structurecould be
prefer-entially used for transposition and linkage to
the 16Scoding portion.
V
b
p %
t A
11%% B J.;
1-'C,
0. 0
0i
4
3 ,0
I
0E
Elec. pH
3.5-1
2
FIG. 5. Two-dimensionalmap onPEI-celluloseofacombined RNaseT2andphosphatase degradation of 18S (a)and19S (b) SV40-specificRNA. The second dimensionwasdevelopedwith 1 Mformic acid-pyridine (pH 4.3). The bluemarker moved 7.5cmin the second dimension.
0
x
E
0.
u
a
28S
b
0
0 0
0.
(w
u
Kr)
I 0w
18S
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.46.237.63.323.2] [image:6.501.50.440.420.621.2]ACKNOWLEDGMENT1S
We thank J. Van derHeydenforculturingcells and virus
stocksand A. Van de Voorde for advice. Weareindebted to
M.Revel,in whoselaboratorypart of this workwasoriginally initiated.
G. H. holdsafellowshipfrom theNationaal Fondsvoor
Wetenschappelijk Onderzoek of Belgium. This project was
supportedbygrant 20.298 from theFondsvoorGeneeskundig
Wetenschappelijk Onderzoek.
LITERATURE CITED
1. Adams, J.,and S.Cory.1975.Modified nucleosidesand
bizarre 5'-termini inmouse myelomamRNA.Nature
(London) 255:28-33.
2. Aloni, Y.,M.Shani,and Y. Reuveni. 1975.RNAs of
simianvirus 40 inproductivelyinfectedmonkeycells:
kineticsof formation anddecayinenucleate cells. Proc.
Natl. Acad.Sci. U.S.A. 72:2587-2591.
3. Aloni, Y.,E.Winocour,andL. Sachs. 1968.
Character-izationof the simian virus 40-specific RNA in virus yielding and transformed cells. J. Mol. Biol. 31:415-429.
3a. Berget, S. M., C. Moore, and P. A. Sharp. 1977.
Splicedsegments atthe 5' terminus ofadenovirus 2late
mRNA. Proc.. Natl. Acad. Sci. U.S.A.74:3171-3175.
4. Cory, S.,and J. Adams. 1975. Themodified5'-terminal
sequencesinmessengerRNA ofmousemyeloma cells. J.Mol. Biol.99:519-547.
5. Desrosiers, R.,K.Friderici, andF. Rottman. 1975. Characterization of NovikoffhepatomamRNA
meth-ylation and heterogeneityinthemethylated
5'-termi-nus.Biochemistry14:4367-4374.
6. Fareed, G.,and D. Davoli. 1977. Molecularbiology of Papovaviruses. Annu. Rev. Biochem.46:471-522. 7.Furuichi, Y.,M.Morgan,A.Shatkin, W. Jelinek, M.
Salditt-Georgieff,and J.Darnell. 1975.Methylated,
blocked 5'-termini in HeLa cell mRNA. Proc. Natl. Acad.Sci. U.S.A. 72:1904-1908.
8. Gelinas, R., and R. Roberts. 1977. Onepredominant
5'-undecanucleotide in Adenovirus 2 late messenger
RNAs. Cell 11:533-544.
9. Gilboa, E.,C.Prives,and H. Aviv. 1975.Purification
of SV40 messenger RNA by hybridization to SV40
DNA covalently bound to Sepharose. Biochemistry 14:4215-4220.
10.Khoury, G.,B.Carter,F.Ferdinand,P.Howley,M.
Brown,and M. Martin. 1976. Genome localization of simian virus 40 RNAspecies.J. Virol.17:832-840.
10a. Klessig, D. F. 1977. Twoadenovirus mRNAshavea common5' terminal leadersequenceencoded atleast 10 kbupstream from their main coding regions. Cell 12:8-21.
11. Krug, R.,M.Morgan,and A.Shatkin. 1976.Influenza
viral mRNA contains internalN-methyladenosine and 5'-terminal7-methylguanosineincapstructures.J.
Vi-rol. 20:45-53.
12. Laskey, R.,and A.Mills. 1975.Quantitativefilm detec-tion of 3Hand14Cinpolyacrylamidegels by
fluorogra-phy.Eur. J.Biochem.56:335-341.
13. Lavi, S., andA.Shatkin.1975.Methylated simianvirus
40-specificRNAfromnuclei andcytoplasm of infected BSC-l cells. Proc. Natl.Acad.Sci.U.S.A. 72:2012-2016.
14. Marx,J. 1977. Viralmessengerstructure:somesurprising
newdevelopments. Science 197:853-855, 923. 15.Moss,B., and F. Koczot. 1976.Sequence of methylated
nucleotides at the 5'-terminus of adenovirus-specific RNA. J. Virol. 17:385-392.
16.Moyer, S., and A.Banerjee. 1976. In vivo methylation of Vesicular Stomatitis virus and its host-cell messenger RNAspecies.Virology 70:339-351.
17.Oda, K.,and R. Dulbecco.1968.Regulation of transcrip-tion oftheSV40 DNA in productively infected and in transformed cells. Proc. Natl. Acad. Sci. U.S.A. 60:525-532.
18.Perry,R.,and D. Kelley. 1976. Kinetics of formation of 5'-terminal caps in mRNA. Cell 8:433-442.
19.Perry,R., D. Keiley,K. Friderici,and F. Rottman. 1975.Themethylatedconstituents of L cell messenger RNA: evidence for an unusual clusteratthe 5'-terminus. Cell 4:387-394.
20. Rose, J. 1975.Heterogeneous 5'-terminal structures occur onVesicularStomatitis virus mRNAs. J. Biol. Chem. 250:8098-8104.
21. Rottman, F., A. Shatkin, and R. Perry. 1974. Se-quencescontaining methylated nucleotides at the 5'-termini of messenger RNAs:possible implications for processing. Cell 3:197-199.
22. Sambrook, J.1977.Adenovirusamazes atColdSpring Harbor, in "news and views." Nature (London) 268:101-104.
23. Sanger, F., and G. Brownlee.1967.Atwo-dimensional fractionation method forradioactive nucleotides. Meth-odsEnzymol.2:361-381.
24. Shatkin, A. 1977. Capping of eukaryotic mRNAs. Cell 9:645-653.
25. Sommer,S., M.Salditt-Georgieff,S.Bachenheimer, J.Darnell,Y.Furuichi,M.Morgan, and A. Shat-kin.1976.Themethylation ofadenovirus-specific nu-clear and cytoplasmic RNA. Nucleic Acids Res. 3:749-765.
26. Southern, E.1974.Animproved method for transferring nucleotides fromelectrophoresis strips to thinlayers ofion-exchangecellulose. Anal. Biochem. 62:317-318.
27. Tonegawa,S.,G.Walter,A.Bernardini,and R. Dul-becco. 1970. Transcription of the SV40-genome in transformed cells andduringlyticinfection. ColdSpring HarborSymp. Quant.Biol. 35:823-831.
27a. Volckaert, G.,and W.Fiers. 1977. Microthin-layer techniques forrapid sequenceanalysisof32P-labeled RNA: double digestion and pancreatic ribonuclease analyses.Anal. Biochem.83:228-239.
28. Volekaert,G.,W.MinJou, and W. Fiers.1976. Anal-ysis of32P-labelledbacteriophageMS2RNAbya mini-fingerprinting procedure. Anal. Biochem. 72:433-446.
29. Wei, C., A.Gershowitz,and B. Moss. 1975. Methylated nucleotides block 5'-terminusof HeLa cell messenger RNA. Cell4:379-386.
30.Weinberg, R., Z. Ben-Ishai, and J. Newbold. 1974.
Simian virus40transcription in productively infected and transformedcells. J. Virol. 13:1263-1273.
31. Weinberg, R.,0.Warnaar, and E. Winocour. 1972. Isolationand characterization ofsimianvirus 40
ribo-nucleic acid. J. Virol. 10:193-201.
32. Yang, N.,R.Manning, and L.Gage. 1976. The blocked andmethylated5'-terminalsequenceof aspecific cel-lularmessenger:themRNA for silk fibroinofBombyx
mori.Cell7:339-347.
on November 10, 2019 by guest
http://jvi.asm.org/