Copyright© 1977 AmericanSocietyforMicrobiology Printed inU.S.A.
Replication Process of the Parvovirus H-1
VIII.
Partial
Denaturation Mapping and Localization of the
Replication
Origin of H-1 Replicative-Form DNA with Electron
Microscopy
IRWIN I. SINGER AND SOLON L. RHODE III*
PutnamMemorialHospital Institute for MedicalResearch,Bennington, Vermont05201 Receivedforpublication 11June 1976
Partial denaturation mapping, restriction endonuclease digestion, and elec-tronmicroscopy were used to determinewhichendof the linearduplex replica-tive-form (RF) DNA molecule contains the origin of RF replication for the parvovirusH-1. Thisoriginwaslocalized withinapproximately 300basepairs of
the arbitrarily designated right end of the RF DNA, inthe EcoRI orHaeII-A
fragment. Based ondenaturationbehaviorinformamide,therightendwas also found to have a relatively high guanine plus cytosine content, whereas the regionadjacenttotheleft terminus oftheRFDNAmolecule was adenineplus thymine rich.
In the previous paper ofthis series (10), we The tsl mutant of the H-1 parvovirus was grownat presented electron microscopic evidence indi- the restrictive temperature (39.5°C) in serum-syn-catingthatreplication ofreplicative form (RF) chronized hamsterembryofibroblasts; RF DNA
rep-DNA, inthe nondefective parvovirus H-1 (tsl lication is enhanced, and progeny DNA synthesis
mutant), occursvia adouble-stranded (ds) lin- shows a temperature-dependent inhibition in this
muant),
s t.
.
mutant. Infected cells were labeled withear Y-Shape rep
licatiove
ntermed
ate(RIl,
[3H]bromodeoxyuridine
(Q3H]BUdR),
and the viral1.55
.tm
inlength. Replicationappeared tohi-
DNA was extracted by the Hirt method. LabeledH-1
tiate near (within 15%o of the genome length) RF and RI DNA were purified by velocity sedimen-one end of the RF DNA molecule; a single tation in a sucrose gradient followed by isopycnic replication fork proceeded toward the other end centrifugation in a second gradient ofCs2SO4. The
ata uniformrate. Thepurpose ofthisstudy is yield of viral DNA fromonepreparationwas
suffi-to determine which end of the H-1 RF DNA cient for all of theexperimentsdescribed below
(ex-moleculecontainsthe initiation siteof RF repli- cept forone in which the label was [3H]thymidine
cation,viapartial denaturationmapping of in- instead of [3H]BUdR). RF DNA wasdigested with
tactRFDNA, RIDNA, andpurifiedEcoRIor EcoRI orHaeII (from
Haemophilus aegyptius)
re-tact
NaeII
RFndonua,eRI
endonuclease-generatedenA,and
purified Eo
fragmentsofRRFo strictioncompletion of the digestionendonuclease (9; unpublished data); the wasverified byanalyti-DNA. cal agarose-gel electrophoresis, and the fragments
Wefoundthat the origin of RF replication is were separated via velocity sedimentation through a
localized nearthe external end of the EcoRI or sucrose gradient. Partial denaturation of the viral
HaeII "A" RFfragment, which constitutes ap- DNA was achieved using high concentrations of proximately 80% of the H-1 genome length (9). formamide (Matheson Scientific Co.) in the
DNA-In addition, this terminus of the A fragment spreadingsolution;82%wasthe formamide
concen-hereafter termedthe right end of the RF mole- tration expected to initiate partial denaturation,
cule, has a relatively highguanine plus
cyto-whereas
90% formamide was anticipated tocom-cule, has a
relatively
whgh
guanine
plus cyto- pletely melt the DNA (13). Three microliters ofasine (G+C) content, whereas the small EcoRI
0.5-mg/ml
cytochrome c solution (horseheart,
orHaeII"B" RF fragment (representing about Sigma, type III) in 0.5 M Tris(pH 8.5)-0.05 M EDTA 20% of H-1 genome), at the left end of the RF was thoroughly mixed with 1 ,lof purified (10)H-1
DNA, has an elevated adenine plus thymine DNA (in 0.01 M
TrisipH
8.5]-0.001 M EDTA), 41 to(A+T) composition. 45 ,ul offormamide, and 1 to 5
Al
of water to produce50 ,ulofspreading solution possessing the desired MATERIALS AND METHODS formamide concentration. This was immediately spreadonto afreshly preparedhypophase of 1 mM The methodsof H-1 viruspropagation, RFDNA Tris(pH 8.5)-0.1 mM EDTA containing from 52 to labeling andpurification, and electron microscopy 60% formamide; a30%difference between the form-have already been describedindetail in the previous amideconcentrations ofthe spreading solution and papers ofthis series (9, 10);therefore, only a brief hypophase was alwaysmaintained toapproximate methodological summary will be presented here. isodenaturing conditions in both "phases" (3). All
724
on November 10, 2019 by guest
http://jvi.asm.org/
solutions were adjusted to 23°C before spreading, . . and were also membrane-filtered (Millipore Corp.)
(except for the DNA and formamide).Preparation of Eco Rl A
grids, DNA spreading, uranyl acetate staining, Pt-Pd rotary shadowing, electron microscopy, and measurement of the DNA contour length were per- 20 formed as previously detailed (10; 4).
RESULTS x
Evaluation of restriction endonuclease E
EcoRIB
digestions and purification of H-1 RF DNA u
fragments. Analytical gel electrophoresis was 10 _
used to determine whether all ofthe H-1 RF
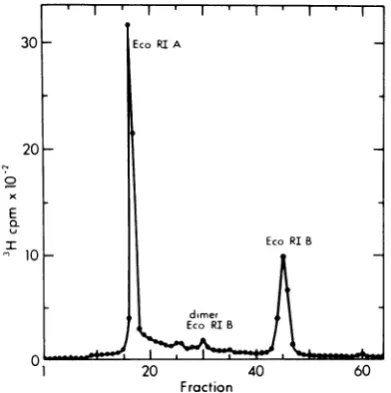
DNA had been cleaved byEcoRI orHaeII. Fig-ure 1shows an agarose-gel profile for theEcoRI
digestion used in this study. All of the labeled A
H-1 RFwas cleaved into fragment A, represent- 0 _
ingapproximately 78% of the label, fragment B 1 10 20 30
representing about 22% of the label, and a Tube Number
small amount ofdimer-length B fragment, pre- FIG. 2. Preparative sucrose gradient
centrifuga-sumably derived from the digestion of dimer RF tion of restriction endonuclease EcoRI fragments A
DNAswhichare linked "tail-to-tail" at the left and B of tsl H-1RF DNA. The EcoRIdigestofH-1
ends of their component monomers (9). Similar RF DNAdescribed inFig.1 wassubjectedtovelocity
results were obtained with endonuclease HaeII sedimentation in agradientof5 to 20% sucrose in 50
(unpublished data).
The A andB H-1 RF frag- mMTris-hydrochloride
(pH 8.0), 1 MNaCl,
1 mM mentswereseparated
usingvelocity
sedimen- EDTA, and0.2%Sarkosyl,
for 7 h at 42,000 rpm in atationthroughasucrosegradient(Fig. 2),orb SW50rotor at4°C.Fractionsof150 ,lwerecollected
tationtrgauory through the bottom of the tube, and
10-pi
aliquots
preparative gel electrophoresis. Electron micro- were used to determine the position of
radioactivity.
scopic
examination ofRF fragments prepared Fractions containing the EcoRI-A and EcoRI-Bfrag-by either method showed slight interpeak con- ments were pooled as illustrated, and the DNA was
precipitatedwith 2.5 volumesofethanolat-20°C for
I 16h. Theprecipitates werecollected by
centrifuga-tion, washed once with 70% ethanol-30% 0.3 M
30 - EcoRIA NaCl-50 mM Tris(pH 7.5)-i mM EDTA,
recentri-fuged,andfinallydissolvedin10mMTris(pH8.5)-i mM EDTAfor electron microscopy. The directionof sedimentation isfromrighttoleft.
tamination, but thiswas of little consequence
20 since these fragments are readily
distinguish-ableonthe basis of
length.
Thesepreparationsx of H-1 RF A and B fragmentswereutilized in
E the experimentsoutlined below.
EcoRIB Partial denaturation mapping of
H-1
RF,n10_ 2 _
RI,
andrestriction endonucleasefragments
ofRF DNA. A highlypurifiedmixture ofH-1 ds
RF and RIDNA, combined with either82, 84,
dimer or 86% formamide in the presence of
cyto-Eco RI l chrome c and
immediately spread
onto aniso-01 20 4060-
denaturing
hypophase
at23°C,
exhibited a120 40 60 unique partial denaturation pattern. One to
Fraction two loops, presumably formed by the preferen-FIG. 1. EcoRIdigestion ofthe[3H]BUdR-labeled tial melting of A+T-rich regions ofDNA
(5-7),
H-1 RF DNA preparation used throughout this were found at an end ofthe RF molecules(Fig.
study. tsl H-1 RFDNA waspreparedaspreviously 3A-H). This A+T-containing region extended described(9). Analiquotwasdigestedwith EcoRI in fromaDNAend(often
forming
aterminalloop)
a reaction mixtureof50pi for4h at37C,and a4-Pi al of e
genomlngth
to
loop)portionwassubjectedtoelectrophoresisin acylindri- along 20% of the genome length into the
RF
calgel (0.6by15cm) ofl.4% agaroseat 25Vfor17h molecule in 76% of the partially denatured RFs;
at23°C (9).Thegelorigin is locatedtotheleft,and the looptraversed 20 to 50% of the RF length in
the anode is located totheright. the
remaining
DNAs(Fig.
4A-C).The extent ofon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.251.448.48.249.2] [image:2.501.48.244.369.566.2]_ . , f s1,
A F
1K
:~~~~~~~~~~~:
FIG. 3. Electronmicrographs oftslH-i RF DNA, RIDNA, HaeII-B,and EcoRI-B fragments,partially
denatured with formamide. Bar = 0.5 pm. (A) RF DNA partially denatured with 82%formamide; the
hypophasecontained 52%formamide. (B-H)As(A),butpartiallymelted withaspreadingsolution
contain-ing84%formamideandahypophasewith54%formamide.(I-L) Uncleaved RI DNAincompletelymeltedas in(B-H).Thepercentageofgenomereplicationincreasesfrom lefttoright:I=12%, J=14%,K =36%,andL = 78%. (M-S)HaeII-Bfragments labeled with[3H]thymidineandpartially denaturedas in (B-H). (T-Z) [3H]BUdR-substitutedEcoRI-Bfragmentspartiallymeltedinan86%formamidespreadingsolution,usinga
hypophase containing56%formamide.
intramolecular
melting
increased withrising
(Fig.
4F). DNAfragments
isolated from restric-formamide concentrations above 82%: 14% of tion endonuclease HaeIIdigests
of the H-i RF thepartially
denatured DNAs showed more described above were alsopartially
denatured than 20%melting
with 82% formamide, 19% with 84% formamide.Incomplete
melting waswith 84% formamide, and 44% inthe presence observedinthe small B
fragments (Fig.
3M-S) of 86% formamide.Partially
denatured RIs andwasconspicuously
absent from thelarger
A(Fig.
31-L) were also observed,constituting
fragments.
Approximately
60% of these par-24% of the total number ofpartially
denaturedtially
denatured Bfragments
apparently
weremolecules; a similar
frequency
of RIs was seen melted closetotheleftend of the RFDNA,thuswhen this DNA was
prepared
under nondena-exhibiting
terminalloops,
whereas othersap-turing conditions with 50% formamide.
Only
pearedtohave meltedthrough
theHaeIIcleav-the
unreplicated
regionsof theseRIDNAmole- age site,forming Y-shaped fragments (Fig.
cules exhibited evidence of
partial melting.
4D). Similar results were obtained usingre-Thus, terminal denaturation
loops
were local- striction endonuclease EcoRI, whosecleavage
ized atthe endof the molecule
opposite
tothat site islocated 0.016 of thegenomelength
tothecontaining
theorigin
of RF DNAreplication
right
of the HaeII siteonthephysical
mapofon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.69.456.64.403.2]___________________ 1, and thus produces B fragments equal to
_________________ ~ 21.6% of the RF DNA length (9; unpublished
A_
_ data). EcoRI-B fragmentsalso exhibitedeither_______________ terminal
loops
or forked ends(Fig.
3T-Z,
Fig.
--____________
4E), whereas the A fragments appeared un-melted upon exposure to 86% formamide. The initiation site for H-1 RF DNA replication is________________
therefore located at the right end of themole--______________ ~ cule, within adomain comprised of 6% of the
- _______________ genome (the smallest fork size), whereas a
_ ______________ unique region of elevated A+T content,
local-E ized
by
reference to theEcoRIIHaeII
cleavage
________________
sites, is atthe left end of theRFDNAmolecule.-____________
=Localization of a G+C-rich region near the--______________ initiation site of H-1 RF DNA
replication.
-________
During experiments conducted to verify that___
_D
our preparation ofH-1 ds RF DNA was fully --=________
melted by90%formamide under the samecon-________
-ditions employed for partial denaturation, we____,
____
observed a dramatic increase in RF contour-
__________
- length(loops were no longerfound). ThemeanC _ E
lengths
ofH-1 RFDNAdecreasedwithincreas--E
ing concentrations of formamide: 1.33 + 0.12,um
(n = 55. +99% confidence interval)at82%.9
formamide,
1.25 + 0.04,um
(n = 154) for 84%,HaI Ecofa and 1.13 ±0.08 ,um (n = 66) at 86%. We
there-FRACTIONAL LENGTH OF GENOME fore expected the
RF
length to decreasefurther whenfully
melted in 90%formamide,
espe-ciallysincethelength ofsingle-stranded (ss)
H-___________________ 1 viralDNAwas33%shorter thanthatofds
H-___________________ 1 RF DNA in 50% formamide (10). However,
________________ the mean contour length ofH-1 RF spread in
______________________ 90%formamide was 1.40 + 0.02 ,um (n = 131), ____________________ whereas that of monomeric ss H-1 DNA
puri-=___________________
fied from virions was nearly halfthat valueF whenpreparedunder the same conditions: X =
0.71 ± 0.04 Am (n = 141) (Fig. 5A). These
____________________ resultssuggested that90% formamide had
en--__________________
tirely converted dsH-1RFto assDNA molecule__-_- ofdimer length. Sincetreatment of dsH-1 RF
6di 63 0:i 07 dI
FRACTKNAL LENGTH OFGENOME [3H]thymidine-labeled HaeII-B fragments, 84% FIG. 4. Formamidepartialdenaturation maps of formamide;and(E) [3H]BUdR-substitutedEcoRI-B tsl H-1 DNA. (A-C) Intact RF DNA, (D) HaeII fragments, 86% formamide. In the case ofthe B endonuclease B fragments ofRFDNA, (E) EcoRI fragments, denaturation sitesrepresented byclosed endonuclease Bfragments ofRFDNA, (F)RIsofH-1 loopswereplacedattheleftendofthe map, andthose RFDNA.Contourlengths ofthe measured molecules melted regions that appeared forked (presumably
werenormalizedto1.0forthe RFs andRIs,or to0.2, meltedthroughthenucleasecleavage sites)were ori-the approximatefractional sizeoftheH-1 chromo- ented to the right. (F) Partially denaturedH-1 RI someforthe RF Bfragmentsgenerated byboth endo- DNAs obtained using 82, 84, or86% formamide. nucleases;arrowsindicate thepositionsoftheHaeII Branched (replicating) molecules whose lengths andEcoRIcleavage sites on theH-1 genome map. (sumoflengths of unreplicatedbaseplusonebranch) The location of denatured regions (indicated by werewithin thelengthrangeofRF DNAsprepared heavy lines)wasorientedtotheleftendofthe RF and with corresponding formamide concentrations, and RImolecules,andexpressedas afraction oftheH-1 which also contained denaturation loops, were
se-genome length. The lengths ofthese denatured ss lectedforthis map. Thelengthswerenormalized and regions were notcorrectedforincreasedshrinkage, thepositionsofthereplication forkswereplottedasa
relativetods areas, whichoccurswith theformamide fraction ofthe total chromosomal length; denatura-concentrations used here. (A)82%Formamide; (B) tionloopswerepositionedasin(A-C).These RIsare 84% formamide; (C) 86% formamide; (D) displayedinorderoftheir extentof replication.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.72.212.60.489.2]I I, 1 I I I II I I5II lI I I 1I 'I I I I 'I ' I '
A.90% C.90%
X.0.19
dimer 1
qds Eco RI-B
40
U~~~~~~~~~~~s.40r
40
~~~monomer
,
x.0.71 dsRF
dimer
E~~~~~~~~~~~~~~~~~~~d Ecofr RI_1
30
_monom
Viral RF.w6
20
|
RFnom~
RFbU5R
R
EcoRI4) I I . 8 RI.A
0
~~~~~~~~I
I0l
QS
1.I.
.
.
.
.
o0
B.96%
D.
96%
length
E
of1.40,um,corremonomeri-.8S6
z
40
monomer ia0.64
30
20
10 RF
~~~~~~~~~~~~~~~~Eco
RI-A10R
00
05
1.0
1.5
2.0
0
0.5
1.0
1.5
Length
(m)
FIG. 5. Histograms ofthe contour lengthsofintact tslH-i RF DNA moleculesand theirendonuclease
EcoRI-generated fragments spread after "fully denaturing" (90%formamide,230C), or fhyperdenaturing"
(96%formamide,s300C)incubations.SinglesarrowsidepictmeanlengthsforssDNA,anddoublearrowsdenote
the meanlengthsofdsDNA. (A)H-iRE DNA meltedwith 90%formamideat230C (solidlines)hasamean
length of1.40 pm, correspondingtodimer-lengthssDNA, determinedby measuringssDNA extractedfrom
purifiedvirions (brokenlines; =(0.71 pm), underthesame denaturation andspreadingconditions. The meanlength ofdsREDNAmonomersmeasuredunder partiallymelting conditions (86% formamide,230C)
was1.13 pm(histogram notshown). (B) DistributionofH-i Re DNAlengthsobtainedbytreatmentunder
whypermeltingconditions"(96%formamide,300C)andimmediatespreadingin90%formamideat23mC.The
meanlengthis0.86 pm,and themajor histogrampeakcorrespondstothatofssDNAmonomers(ssviralDNA in[ADspread in thesamemanner.(C) Endonuclease EcoRI-A and-Bfragments ofH-i REDNApreparedin 90% formamide233C.atssBfragmentshadameanlengthof(0.19 m,which isshorterthanthatobservedfor monomerdsBfragmentsprepared underpartiallydenaturingconditions(86%formamidet,23tC;s 0f24=
pm,histogramnotshown).Afragmentsexhibitedtwomajorpeaks in90%23[C (solidformamideat lines),
withmeanscorrespondingtossdimerlength(~=1.15pmn),andssmonomerlength (* =0.61 pm).Themean
length of monomer ds EcoRI-A fragments prepared in 86% formamide at 239C (partially denaturing
conditions)was0.86 pm(histogramnotshown).(D)HistogramofEcoRI-Afragments ofH-iRE
'"hyperdena-tured" with 96%formamide at30"C, andspread as in (B). The DNA was completelydissociated into ss monomerAfragments (~=0.64 pm), whosecontourlength distribution coincides with thessAfragment
monomersobservedafterdenaturation with 90%formamide (shownin[C]).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.64.460.51.465.2]REPLICATION 729
withanalkalinesucrose
gradient
doesconvertafter
similar treatment of ds H-1 RF (Fig. 5A).90% of it into monomer ss H-1 DNA
(9),
weIncubation
of EcoRI-Afragments
of H-1 RFdecided
tofurtherinvestigate
thenatureof the DNA in 96%formamide
at 30°C for 30 minlinkagebetween the two ss monomers. ds H-1 converted them intossmonomers (£ = 0.64 +
RF DNA (in 10mM
Tris[pH
8.5]-1 mMEDTA)
0.03,um;
n = 107) (Fig. 5D). These datacollec-was placed in 96% formamide at
30°C
for 30tively demonstrate
thatthe
regionofhigh
G+ Cmin,followed
by
theaddition of sufficient cyto- content islocated close
tothe
right
endofthe
chromecsolution (in0.5M
Tris[pH
8.5]-0.05
M RF DNAmap.EDTA)tolower the formamideconcentrationof
theresulting solutionto
90%,
andimmediately
DISCUSSIONspread onto an
isodenaturing
hypophase
con- Features oftheH-1
RF DNA monomer, astaining 60%
formamide
at23°C.
Weestimated determinedinthis and the twoprecedingstud-that the
melting
temperature(Tm)
of the 90% ies in this series, are summarizeddiagramati-formamide spreading solution was
approxi-
cally inFig. 6. Our results show that aregionmately
300
below theT,,
ofpoly(deoxyguani-
susceptibletomeltingunder partialdenaturingdylic
acid:deoxycytidylic
acid)[poly(dG
dC),
conditions existsadjacenttothe endo R-EcoRIand that the Tm of the96%formamidemixture and HaeII cleavage sites, which are located
was
10.40C
above that of thishomopolymer.
approximately 20% of the genome lengthfrom These calculations are based on a Tm of780C
the arbitrarily defined left end of theH-1
RFfor dG dC in a solution with acation concen- DNA molecule (9). Partially denatured
inter-trationof6mM(3)andapH of7.0 (12), aswell mediates in
H-1
RFDNAreplication(RIs)
simi-as a 0.6°C Tm reduction for each 1% increase larlyexhibited melting near the left end of theinformamide concentration (1). After
exposing
molecule, whereas the origin ofreplication,de-ds H-1 RFDNA tothe96%formamide
solution,
termined by the position of early replicationthe contour
length
decreasedto that ofssH-1
forks, was localized close to the right end of thisDNAmonomer size (Fig.
5B):
X =0.86
±0.04
DNA. Based on the size of the smallestdaugh-um, n = 197,
indicating
that theregion
com- ter arms observed, the region containing theprising theintermonomer
linkage
probably
has replication origin is located within 700 basea
high
G+C contentandmorestablehydrogen
pairs (10), but more probably within 300 basebonding, as
opposed
to acovalent"turnaround" pairs(this study) of the right end of theH-1
RFofribonucleotides.
We performed the above experiments on Left
Right
EcoRI restriction
endonuclease-generated
A A-TRich
G-CRich
and B fragments of dsH-1 RF DNA to deter- R c
mine which end of this molecule is G+C rich
C
and
responsible
forlinking
the termini of the'E
monomers inthess
dimers
generated
with90%HaeEL
EcoRI V Originformamide. Themean
length
of the ds Afrag-
t I Iment in 86% formamide
(distribution
not 0 1 2 3 4 5shown)
was 0.86 ±0.05 am(n
=72), represent-
Kilobase Pairs
ing78.2% of the genome, and that of the ds B
fragment
spread
under thesameconditionswas FIG. 6.Schematic
representation
ofH-i
RFDNA 0.24 + 0.02 ,um (n =75),
equal
to 21.8%of themonomer in the
foldback
configuration
(9).The
mol-RF length. These size measurements are in ecule istion endonuclease HaeII and EcoRI sites4,900 base pairs in length, with the restric-(arrows)good agreement with data obtained from ana- located 900 and 1,000 base pairs,respectively, from
lytical
gel
electrophoresis
ofEcoRI-generated
the endcontainingthe covalent turnaround (basedH-1 RF DNA
fragments (9). When
the Bfrag-
on molecular weights given in[9D;
the large andments were
spread
inthepresenceof90%form- small RFfragmentsgenerated bytheseendonuclea-amide, their
meanlength
decreasedto 0.19 ± ses arereferred
to asA and B(respectively).
V,
Viral 0.01,tm
(n =85),
indicating
that thisfragment
strand; C,complementary
strand. Thezonein whichdoes not contain the
G+C-rich
linkage
group the initiation site forRF DNA replication is located isunder
study (Fig.
50).Preparation
of the A delineated by the open bar and is composed of basefragmentin 90%formamide
generated
twosub- pairs 4,600 through 4,900(6%
of genome). The Bpopultn(e
banalysis,
region isrelatively
richin A +T content(indicatedbypuion (revea led
byhiAStor
a 3X
crosshatchedbar),and theright
end hasanelevated(Fig. 50) with meanlengthsOf 0.61 ± 0.03 zm G+C composition (asterisk). More recent
experi-(n = 94) and 1.15 + 0.03 ,um
(n
=108),
repre- ments have shown that the structure oftherightendsenting the ss A
fragment
monomer andss A is morecomplicated thanindicatedinthisdiagram
fragment
dimer,
respectively,
as was observed(manuscript
inpreparation).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.254.441.386.472.2]DNAmolecule. The right terminus of the H-1 cause many of the undigested RF molecules chromosome also exhibits a region that is re- treated with 86% formamide exhibited
melting
fractory to melting in a solution containing 90% at thesecleavage sites. Furthermore,
complete
formamide [withan effectivemelting tempera- melting of the left terminus of RI molecules ture about 3°C below that of poly(dG dC)], re- subjected to partial denaturation would
sulting in the formation of ss dimers under have generated double Y-shaped molecules
these conditions. No knobs or branches were (> <), which were not observed. However,
detected at the midpoint of these ss dimers after we could not definitely exclude thepossibility
electron microscopic study of more than 200 thatmeltinghad occurredthroughthenatural molecules, indicating that this presumably ds left RF end because the resultant Y-shaped
linkage region isrelatively small.The observa- molecules would have been easily confused tions that it remains intact under conditions with unmelted early RIs and therefore would that completely melt the rest of the RF mole- not be included in oursample.
cule, and that exposing this DNA to an esti- It is alsopossible that the branchedportions
mated
T,,,
of10.4°C above the melting point of of the RIscontaining denaturationloopsin thedG dC homopolymers converts the ss dimers B region could have been formed viamelting of
into ssmonomers, strongly suggested that this the right end the molecule instead of being linkage is due to a G+C-rich terminus rather intermediates of semiconservative RF DNA than tocovalentbonding withribonucleotides. replication. We believe this to be unlikely since
The renaturation properties of the EcoRI-A branched structures were never observed fragment, which contains the rightend, have amongmorethan 200EcoRI-Afragments mea-also ruledoutthepossibility ofacovalent link- sured with the electronmicroscope afterpartial
age at the right endof H-1 RF DNA (9). Cur- denaturation. Also, the number and
distribu-rentexperimentsindicate that the structure of tion of replication forks along the contour the rightendis morecomplex than indicated in lengths of incompletely melted H-1 RI mole-Fig. 6. Inaddition, theHaeII or EcoRI endonu- cules is very similar to that observed after clease B fragment, containing the left end of spreadinginnondenaturing conditions (10). the RF DNA, forms denaturation loops under The electron microscopic localization of the
partially denaturing conditions (82, 84, or86% initiation site forH-1RFDNA replication close
formamide), and therefore is A+T rich (5-7). to the right terminus of the RF molecule is
Most of the RF molecules observed had par- consistent with radiochemical experiments in
tially melted so close to the left end that a the accompanying manuscript (9), monitoring
terminal loopwasgenerated. Sinceonly 10% of the incorporation of [3H]thymidine intoEcoRI
H-1 RFmoleculesexhibited evidence of a cova- fragmentsof H-1 RF DNA after short labeling lently closed turnaround at the left end (9), it periods, which indicated that the origin of RF seems unlikely that covalent bonds were re- DNA synthesis is located somewhere within
sponsible forstabilizing the apex of this end in theEcoRI-A fragment. The latter experiments
the other90% of themolecules under partially have also shown that the initiation point for
denaturingconditions. It is more probable that progeny ssDNA synthesis probably has a
simi-this B apex isstabilizedby adG dC enrichment lar location. These data collectively indicate similar to that present atthe right terminus, that the H-1 RF DNA is a structurally and
but exhibiting a lower melting temperature, metabolically polarized molecule, with RF and
since EcoRI-B fragments (with the left end) probably progeny DNA replication initiating
werecompletely dissociatedby90%formamide, nearthe dG+dC-rich right end, and then
pro-whereas the A fragments (with the right end) ceedingtothe leftterminus(10). This left end is were not. Theexpected10%of the B fragments, adjacent to a region with a high dA+dT con-which should have been dimer length in 90% tent, as demonstrated by partial denaturation formamide due to the presence of a covalently mapping.Italso exhibitsatleast two structural closedturnaround, were not demonstrated with forms thatdifferby50to70base pairs in length anycertainty usingelectron microscopic meth- (9).Theself-cohesiveproperty of this left termi-ods. We also observed that incomplete melting nus implies that it contains an inverted self-conditions gave rise to Y-shaped B fragments in complementary sequence, which forms a cova-addition to loops located at either the natural
lently
closed turnaround in some molecules. left terminus, or near the middle of this frag- This structural model for the left end of H-1 RF ment. We postulate that these Y-shaped frag- DNAissimilartothat recently propounded for mentsresulted from melting through theEcoRI both DNA ends of adenovirus-associated virusand HaeII cleavage sites, rather than from
(AAV),
a defective parvovirus, only capable ofmelting of the natural left end of the RF, be-
replication
in cells coinfected with adenoviruson November 10, 2019 by guest
http://jvi.asm.org/
VOL.
(2). AAV ss viral DNA molecules are either adenovirus for adenovirus-associatedvirus
multipli-plus or minus strands containing self-comple- cation.Nature(London)New Biol. 244:71-73.
mentarytemnlsep 3. Davis, R. W., and R. W. Hyman. 1971. A study in
mentary terminal sequences capable ofintra- evolution: the DNAbasesequence homology between
molecular annealing with theconsequentgen- coliphages T7 and T3. J. Mol. Biol. 62:287-301.
eration ofss circles orconcatamers (8). It has 4. Davis,R. W., M.Simon, and N. Davidson. 1971. Elec-beenproposed that both ends of duplex AAV RF tronmicroscope heteroduplex methods for mapping
DNAcontainaself-complementaryfoldbackse1 regionsofbasesequencehomologyinnucleicacids,p.
D)NA
containaself-complementary foldback se-413-428.
In L.Grossman and K.Moldave (ed.),Meth-quencewhose 3' hydroxyl terminus may serve ods in enzymology, vol. XXI. Academic Press Inc.,
as a primer for both RF and progeny DNA NewYork.
replication at either end of the RF molecule 5. Goldstein,L., M.Thomas, and R. W.Davis. 1975.Eco
(1 Since the initiation site
of.
H.1 RF and T TRIendonuclease cleavagemap ofbacteriophageP4-(11). Since the
initiation site of H-i
RFand
DNA. Virology 66:420-427.presumably progeny DNA synthesis is at or 6. Inman, R. B., andM. Schnos. 1970. Partial
denatura-near the G+C-rich right end of the H-1 RF tion of thymidine- and 5-bromouracil-containing A
molecule,
andacovalentturnaroundislocated DNA in alkali. J. Mol. Biol. 49:93-98.ontheleft,
end,(9,
this invertedself-comple,
7. Jacob, R. J., J. Lebowitz, and A. K. Kleinschmidt.On the left end (9), this inverted self-comple- 1974. Locating interrupted hydrogen bonding in the
mentary terminus is notinvolvedin theinitia- secondary structure of PM2 circular DNA by compar-tion of H-1 DNA replication. It is of further ativedenaturation mapping. J. Virol. 13:1176-1185. interest thatunlike
AAV,
H-1 ss progeny DNA 8. Koczot, F., B. J.Carter, C. F. Garon, and J. A.Rose.consists
ofonlonefthetwocmplemnta1973.
Self-complementarity of terminal sequencesconsists of only one of the two complementary within plus or minus strands of
adenovirus-associ-strands, whichmaybe relatedtotheasymmet- ated virus DNA. Proc. Natl. Acad. Sci. U.S.A.
ricinitiationof DNAsynthesis atonlytheright 70:215-219.
end ofthe H-1 RF DNAmolecule. 9. Rhode,S. L. 1977.Replication process oftheparvovirus
H-1. VI.Characterization of a replication terminus of H-1 replicative-form DNA. J. Virol. 21:694-712. ACKNOWLEDGMENTS 10. Singer, I. I., and S. L. Rhode III. 1977. Replication This work wassupportedby PublicHealthService grant processof theparvovirusH-1.VII. Electron micros-* CA-07826-11 from the National Cancer Institute, and a copy of replicative-form DNA synthesis. J. Virol.
generousgiftfromThe Given Foundation. 21:713-723.
Wegratefully appreciate the excellent technical assist- 11. Strauss, S. E., E. D. Sebring, and J. A. Rose. 1976. anceofRobertCostantino and JessicaBratton,and thank Concatemers of alternating plus and minus strands Kay A. 0.EllemandHelene Toolan forcritically reading are intermediates in adenovirus-associated virus thismanuscript, andVirginiaHaas and Janeen Pratt for DNA synthesis. Proc. Natl. Acad. Sci. U.S.A.
73:742-secretarialduties. 746.
12. Szybalski,W.1967.Effectsofelevatedtemperatures on LITERATURE CITED DNA andon somepolynucleotides:denaturation,
re-naturationand cleavage of glycosidic and phosphate 1. Bluthmann, H.,D.Bruck,L.Hubner,and A.Schoff- esterbonds, p. 73-122. In A. H. Rose (ed.),
Thermo-ski. 1973. Reassociation of nucleicacidsinsolutions biology. Academic Press Inc., London.
containingformamide. Biochem. Biophys. Res. Com- 13. Wolfson, J., D.Dressier, and M. Magazin. 1971. Bacte-mun.50:91-97. riophage T7 DNA replication: a linear replicating 2. Carter,B.J., F. J.Koczot,J.Garrison,R.Dolin,and J. intermediate. Proc. Natl. Acad. Sci. U.S.A.
69:499-A.Rose. 1973. Separatehelper functionsprovided by 504.
![FIG.3.[3H]BUdR-substituteddenaturedhypophaseinghypophasein= 78%. (B Electron micrographs of tsl H-i RF DNA, RI DNA, HaeII-B, and EcoRI-B fragments, partially with formamide](https://thumb-us.123doks.com/thumbv2/123dok_us/1550049.107536/3.501.69.456.64.403/substituteddenaturedhypophaseinghypophasein-electron-micrographs-haeii-ecori-fragments-partially-formamide.webp)



