Copyright©1968 American SocietyforMicrobiology Printed inU.S.A.
Interference
Among Group A
Arboviruses
EUGENE ZEBOVITZ AND ARTHUR BROWN
BiologicalSciences Laboratories, Fort Detrick, Frederick, Maryland 21701
Received for
publication
29 July 1968Interference among group
Aarboviruses
isdescribed
whichdoes
not involve themediation
ofinterferon. Interference was observed
only if theinterfering
virushad
an
advantage over the
challenge virus, either in time or in multiplicity of infection.
Adsorption,
penetration, and uncoating of challenge virus did not appear to be
in-hibited,
but thesynthesis of infectious viral ribonucleic acid
of thechallenge
viruswas
significantly retarded. It
wasshown with
temperature-sensitive viruses or mutants
that
the
replication
of viral ribonucleic acid by the interfering virus was required to
establish
interference. A mechanism of interference based on a competition for
replication sites or substrates is compared with other possible explanations.
The practical and theoretical
implications
ofviral
interference have stimulatedconsiderable
interest in this research
area.Many reports have
been
published
since thediscovery
ofinterference inplant
viruses(14)
and inanimal viruses
(11).
Those papers that appeared
prior
tothediscovery
of interferon
(13)
have been reviewedextensively
by
Henle(10)
andSchlesinger
(18).
The role ofinterferon
as animportant
factor inthedevelop-ment of
nonspecific
resistanceof the host cellstosuperinfection
with a second related or unrelatedvirus is now
firmly
established(6).
It ispresently
difficult
todeterminewhich of theearly reports
onviral interferencewereconcerned with interferon
production
by
the hostorby
otherfactors. It isapparent,
however,
that there areseveraltypes
ofviral interference
that do not involvethe
media-tion of interferon(5, 7,
12, 15-17).
This report
describes anoninterferon-mediatedinterference
among
differentarboviruses;
thisinterference
requires
that theinterfering
virus be able toreplicate
viral ribonucleic acid(RNA)
inthe host cell beforeit caninterfere withthe
growth
of the
challenge
virus.Further,
interference of thechallenge
virusappears
to occurafteritsuncoat-ing
but beforesynthesis
of infectious viralRNA.MATERIALS AND METHODS
Virus strains. In mostof the experiments
reported
here, the virus used to induce the interference was either the Trinidad strain of Venezuelanequine
encephalitis (VEE) virus or strain T, which is a
small-plaque,
temperature-sensitive
variant of this virus. In a fewexperiments,
theinterfering
viruses were: a large-plaque revertant ofT,designated
V5,whosemaximum
growth
temperaturewasunchanged
(S. Halle, personal communication); an attenuated variant(A)
of VEE virusoriginally
describedby
Berge et al. (1); and atemperature-sensitive mutant (Ets-4) of the Louisiana strain of eastern equine
encephalitis virus (EEE). EEE virus served as the
challenge virus in most experiments. Properties of
these viruses, except for Ets-4 and V5, have been
describedby Brown (3).
Cellcultures. Cell cultures were prepared from
10-day-oldchickembryos. Thechick embryo (CE)
mono-layersweregrown,inlactalbuminhydrolysate medium
containing 10%calf serum, for24hr at 37 Cbefore infection. Details of the preparation ofmonolayers,
medium, and growth conditions were described in an earlier paper(22).
Virus growth. Except wherenoted, CE cell
mono-layersin 60-mmplasticpetri disheswereinfected with a virus seed preparedas a 10% CE seed, or as un-diluted tissue culture fluids obtained from infected
CEmonolayers.Theviruswasallowedtoadsorbfor 15 min at room temperature; thecells were washed twice withphosphate-buffered saline (PBS)atpH 7.4 to remove excess unattached virus, and were over-layeredwith5mloflactalbuminhydrolysatemedium
containing 10% calf serum. The cultures were
in-cubatedat37 Cina5%
C02-95%
airincubator.When the
interfering
and challenge virus weresimultaneously added to CE monolayers, the virus
seedsweremixed beforeinfectionof the cells. When the challenge virus was added at some time after
infection of the cells with the
interfering
virus, the culture medium was removed, the challenge viruswasadded, and,aftera 15-min adsorption periodat room temperature, the infected cells were washed twice withPBSandoverlayeredwith thelactalbumin hydrolysate medium. The cultures were reincubated at 37 C andsamplesof the culture mediumwere col-lectedatvarious intervals.In mostoftheexperiments, actinomycin D (1 or 2
,Ag/ml)
was incubated with cells for30min before infection andwasinthe culture medium at the same concentrationthroughout
theexperiment.
Virusassay. Virusassayswereperformedon24-hr 1283
on November 11, 2019 by guest
http://jvi.asm.org/
CE monolayers prepared from 10-day-old chick embryos. The overlay medium was lactalbumin
hydrolysate-agar (21). In those studies involving mixed infection with VEE and EEEviruses, titers in the growthmedium were determined in thepresence
ofa 1:100dilution of anti-VEEserum (whose plaque
neutralization titer exceeded 1:10,000) added to the
agaroverlaymedium.PlaqueformationbyVEE virus
was inhibited but that of EEE virus was not. This permittedassay ofEEE virusgrowth in thepresence
ofa large excess of VEE virus. When strain T was
used, it was not necessary to add antiserum to the overlaybecause this virus formed very smallplaques and waseasily distinguishable from EEE virus when
assaysweremade onsamplesfrom mixed infections.
In reconstruction experiments involving mixtures
andcontrols,itwasfoundthat 100-to200-foldexcess
of T over EEE did not inhibit plaque formation of EEE, thusjustifying the procedure ofplaquing and countingEEEinthepresenceofexcessT.
Extractioni and assay ofinifectious RNA (IRNA). InfectedCEmonolayers wereremovedwitharubber
policemanand suspended in 0.02 M phosphate-0.001 M ethylenediaminetetraacetic acid buffer (pH 7.4).
Thecellswereextracted twice withcoldphenol (4 C), and theviralRNAwasprecipitatedfrom theaqueous
phase with three volumes of9570 ethyl alcohol
con-taining 2.0%// potassium acetate. The precipitate was
dissolvedin PBS, andthe IRNA wasassayed onCE monolayers treated with 1.0NI NaCl in a 0.1MI
tris-(hydroxymethyl)aminomethane-chloride buffer (pH 8.3) bythemethod of Colon and Idoine (4).
RESULTS
Demiionstration of interference. Interference of thechallengevirus could be demonstrated intwo
ways:(i) by infectingcells withVEEvirus several hours beforesuperinfectingthecultureswithEEE virusatmultiplicities equaltothose used forVEE
virus, and (ii) by infecting the CE cells
simul-taneously with two viruses at different
multi-plicities; the interfering virus was added at an
input multiplicity of about 10 plaque-forming
units (PFU) percell, whereas thechallengevirus
wasusedatamultiplicityof 0.1 PFUpercell. When equal multiplicities of the two viruses were employed, the degree of interference was
dependent upon the time of superinfection with thechallengevirus(Table 1).Thedegreeof inter-ference increased with the time that elapsed
be-fore superinfection with the second virus. Maxi-mal inhibition of the growth of EEE virus was observed when it was used to superinfect cells 5
to6 hr after infectionwith T virus.
The effect ofinfecting CEcells simultaneously with strainT and EEEvirus isshown inTable 2.
Strain T was addedata constant multiplicity of infection (MOI) of 10 andEEE viruswasadded
at an input MOI ranging from 1.0 to 0.01. The previous experiment and others showed that simultaneous infection at equal input
multiplici-TABLE 1. Effect of timne of superinfection on initer-ference with challenzge virius
Growth response ofchallenge Timeofsuperinifection virus(EEE)
with 11EEv-irusa
I'lFUml
Logioinhibition
hr-0 7.7 X 109 0
1 3.1 X 109 0.4
2 1.9 X 109 0.6
3 4.5 X 108 1.2
4 1.9 X 108 1.6
5 2.5 X 107 I 2.5
6 9.0 X 106 2.9
7 1.1 X 107i 2.9
aCultures were infected with strain T virus at
an input MOI of 10. Cultures were then washed two times with PBS and overlayered with culture medium. At the indicated times, themediumi was removed and the cultures were superinfected with EEE virus at the same multiplicity; the cultures were then washed twice and overlayered with
growth medium.
ICultures were held at 37Cfor 24 hr after
addi-tion of the challenge virus before virus assays were made.
cThis 24-hr titer was approximately the same
for all theEEE singly infected control cultures.
TABLE 2. Effect ofiunput muiiltiplicity ofthe
challenige
vi-rusoln
thein tee
fierence
wvit/i challen1ge
viruis
Inputmultiplicity PFU/m (2- hr)
Lo-io
irlhibition ofchallengexvirus'i E/i(4ucioControlb 1.9 X 10] 0
1.0 4.0 X 108 0.7
0.1 4.7 X 107 1.6
0.01 5.1 X 10- 2.6
Multiplicity of the interfering virus (strain T) was heldconstant at 10 PFU/cell.
bThe 24-hr control value was the growth re-sponse of EEE virus in the absence of strain T; this was approximately the same for each of the multiplicities tested.
ties of 10 resulted in no interference of either
virus;
multiplicities
higher
than 1 were therefore omitted.Theresults showedaprogressive
increase in the degree ofinterference as the multiplicity ofEEE virus decreased. Maximal
interference
was observedwhen thelowestmultiplicity
ofchallenge virus was used. The effect ofmultiplicity
in thisexperiment
wasprobablyonlyanother manifesta-tion oftheeffect oftime, since,
atthelower multi-plicities, more than one cycle ofchallenge virusmultiplication
would be required to reach the levels obtained by the interfering virus in oneon November 11, 2019 by guest
http://jvi.asm.org/
[image:2.484.252.443.80.222.2] [image:2.484.253.441.374.473.2]cycle. Inthe absence of strain T, EEE virusgrew
normally andto high titer; although the dataare notshown, therewas nointerference with T virus
growth in such doubly infected cells.
Therewas astronginhibitionof the growth of thechallenge viruswhen theinterfering viruswas
givenagrowthadvantage inthe CEcells either by
being inoculated several hours earlierorata
sig-nificantly higher MOI than the challenge virus (Tables 1 and 2). If the interfering virus was
treated with specific neutralizing antiserum just before infecting CE cells, interferenceto
superin-fectionwiththechallengeviruswasnotobserved. Controls consisting ofneutralizing antiserum to EEEvirus,ornormalserum,when incubated with
VEE as interfering virus prior to infection, did not preventtheinterference. These results showed
that infection bythe virusparticlewasnecessary
toestablish interference and that the interference
was probably not dueto interferon in the virus
suspension. The latter conclusion was supported
by the fact that virus that was sedimented and washed twice had the same interfering capacity
as crude virus.
Interference inducedbydifferentstrainsofVEE virus. Interference of the growth of EEE virus could also be demonstrated when other strains of VEE virus wereused. In addition, EEEvirus
could be used as the interferingvirus and could
inhibit the growth ofany strain of VEE virus.
However, there seemed to be some variation
amongvirus strains in theircapacity to serve as interfering viruses (Table 3). Strain A was
con-sistentlymoreeffectiveasaninterferingvirus than wasthe Trinidadstrainortheothers;theTrinidad strain was more effective than either T or V5;
therewasnosignificantdifference betweenTand
V5. The differences in the degree of inhibition induced by different strains may possiblybe ex-plained bydifferences in thecapacityof the
[image:3.484.251.444.82.216.2]differ-ent virus genomes to attach to replication sites
TABLE 3. Capacity of differentt straintsofVEE virus tointerfere with thegrowth ofEEE
virus inchick embryo cell culture
VrEE virusstramna Degree of EEE virusinterferenceb
A 2.4
Trinidad 1.6
T 1.4
VS 1.1
aInput MOI, 100PFU/cell.
bInput MOIofEEEvirus, 1 PFU/cell.Thedegree
[image:3.484.48.239.513.616.2]of interferenceisexpressedaslog,0units ofEEEvirus at20 hr. Valuesareaveragesof fourexperiments.
TABLE 4.
Effect
ofactiniomycinz D oniinterferenice
with EEE virus multiplication by strain Ta
EEE virus (PFU/ml)
Time Sri
Strain T+ Stai T+ EEEvirus
EEE virusb EEEvirus alone
EE actinomycinD~' aln
hr
0 1.2 X 105 1.2 X 10' 1.2 X 105 6.5 4.1 X 106 6.4 X 106 6.0 X 107 22 4.7 X 107 4.6 X 107 2.4 X 109
aActinomycin D (1.0
gg/ml)
added at 2 hrbefore infection.
bInfection by strain T followed by EEE 3 hr laterat equal multiplicities (100).
within
the hostcellorbydifferences
in their rates or extentofviral
RNAreplication.
Is the interference of virus growth mediated by interferon?
Actinomycin
D wasused to helpob-tain evidence
onwhetherinterferon
was a factorin
theinterference
that was observed. This drug isknown
toinhibit both
theformation
and actionof
interferon in virus-infected
cells, yet it does not interfere with the synthesis of many RNA viruses (8, 20). Ifinterferon
wereinvolved
in the inter-ference observed here, then the challenge virusshould be able
tomultiply
normally in
thepres-ence of
actinomycin
D. Whenactinomycin
D(1
Ag/ml)
wasadded
2hrprior
toinfection,
it hadnoeffectonthe
interference
ofEEEvirus
in cells that had beenpreviously infected
with a highmultiplicity
of
strain
Tvirus (Table
4). In boththe presence and absence of
actinomycin
D, the growth of thechallenge virus
wasinhibited
to thesame extent, about 1.6
log10
less
than thatob-tained for
thecontrol culture.
Thattheactinomy-cin D was
active
was shownby
thefact
that growth ofvaccinia virus
wasinhibited
by more than99%in
CEcells
in
thesameexperiment.
Inaddition,
thedrug abolished
theinterference
re-sulting
fromadded
chick interferon (50
plaque-inhibiting units) in
aSindbis virus-CE cell
test system.Fromtheseresults, it
seemslikely
that theinterference observed
does not result from theformation
oraction
ofinterferon
by the host cell. Subsequentexperiments
werecarried
out inthe presence of
actinomycin
D,andby infectingwith
equal
multiplicities
of theviruses
3 to 4 hrapart.
RNAsynthesis by the challengevirus.The inter-ference ofEEE virus growth that resulted when
strainT wasinoculatedontoCE cells4 hr
earlier
is shown in
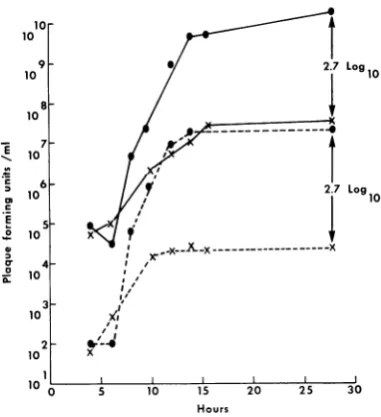
Fig.
1.TheEEEvirus titer
wasreduced2.7
log1o
below
that observed forthe controlcul-ture. In
doubly
infectedcells,
the synthesis ofon November 11, 2019 by guest
http://jvi.asm.org/
vitz and Brown, unpublished data). EEE virus, on the other hand, replicated normally at this
tem-perature. It waspossible, therefore, to inhibit the 7 ~~~~2-7Loglo growth of
strain
Tselectively
by
incubating
in-I fected CEcultures
at 42 Cprior
toand after thecultures were superinfected with EEE virus.
Figure
2shows
theeffect
ofincubating doubly
J Z j K
infected cultures
at 42 C. Inthis
experiment, T/o/x,
viruswasadsorbed
tocells
atroomtemperature
2.7
Log10
washed
asusual,
and
incubated
at 42Cfor
3 hr- ,,
before superinfecting with
EEEvirus
andincubat-X
f'
fl ting
further at 42C.
The interference normallyrx--X-x--- ' observed was largely abolished;i.e., the maximum /
.',,'
titer
of EEEvirus
was notinhibited.
These resultssuggestthat, as a
minimum condition
toestablish
interference,
theinterfering
virus RNA must beable
toreplicate in
thehostcell in order
toinhibit
thegrowthof the challenge
virus effectively.
25 The
results
discussedabove
support
the notion5 10 15 20 25 30 that a
competition
forreplication
sites or sub-Hours stratesaccounts for the interference observed. To 1.Interference
of challenge virus (EEE) explore this idea further, a recently isolated tem-andinfectious RNA synthesis by strain T virus. perature-sensitive mutantofEEE
virus wasem-infected at equal multiplicity (10 PFU/ml).
ployed.
At 37C, but not at 30 or 42C,
Ets-4virus ige virus was addedto the culture3hrafter infec- exhibited anunusually
high rate and extent ofith strain T virus. Symbols:
(@-*)
singly viral RNAsynthesis;
itinduced the formation of IEEEvirustiter; (X-X)doubly
infectedEEE approximately three times the amount of viral iter; (0--0)
singly infected EEE IRNA; RNAcompared to the parent, but produced90%
()doublyinfected EEE IRNA. lessinfectious virus and at least50%7
lesscomple-ment-fixing
antigen
(Zebovitz
and
Brown,
unpub-IRNA
wasreduced
tothe
sameproportion asthat
of the maturevirus.
Theseresults suggest that theinterference
phenomenoninvolves
a veryearly
step
in
thesynthesis
of thechallenge
virus,
prob-ably
before
thevirus
hashad
theopportunity
tosynthesize
its
IRNA. On thebasis ofourlimiteddata
(Fig. 1),
there appears tobe
noobvious
preferential interference between
IRNAsynthesis
and
viral
structural
protein
synthesis; otherwise,
the curvesof
virus interference and
IRNAsynthe-sis
would
not beexpected
toparallel
each other. However,since
nodirect
measureofviral
protein
synthesis
wasmade
independent
ofinfectious
virus,
this conclusion is tentative.
Apparently,
theearly
step of thechallenge
virus
infection
that was inhibited was not adsorption,penetration,
oruncoating
of thevirus
genome, because interference was ofthe samemagnitude
whenIRNA of the
challenge virus
wasusedin theplace
ofinfectious virus.
Thisconclusion
receives
additional
support fromsomeincidental evidence
obtained
in anexperiment
(described
below) with
the T strain as
interfering virus
at 42C.In
previous
studies(22),
itwasestablished
that the RNA of VEEvirus
enters thecell andis
main-tained
in aviable
state, even forprolonged
periods
at 44C,
although
nonewRNAis
synthesized.
The sameproved
true forstrainTvirus
at42C(Zebo-10 .
9 10
E
"I
c c
r-8 10
7 10
6
10
105[
104
3
10 2
0 ~~~~~~~0
.1,
11
00s
X"
0 5 10 15 20 25 30
Hours
FIG. 2. Effect of incubationat42 Con thecapacity
of strain T virus to interfere with EEEvirusgrowth.
Challenge virus (EEE) wasadded3hrafter infection of CE cells with strain T. Symbols: (0) EEE virus
growthinabsence ofstrain T; (X) EEE virus growth
onCEcells infected with strain T. (0) StrainT virus
growth inabsenceof EEE virus. 10
10 9 10
8-10
7-7
- 10
°
106
10
.t
5.2 10
10 2
3:10
10
FIG. growth Viruses
Challen
tion wi infected virus ti (X-->
Yr%'ITA
1286
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.484.50.241.61.270.2] [image:4.484.255.443.385.572.2]TABLE5. Comparison of Ets-4andEEE as interfering virus
Time aftersuper- Control VEE EEE+VEE Logioinhibition Ets-4 + VEE Logioinhibition
infection' (PFU/ml) (PFU/ml) (PFU/ml)
hr
0 7.1 X 104 6.0 X 104 5.1 X 104
5 1.5 X 106 4.0 X 105 0.6 1.0 X 105 1.1
8 2.0 X 107 3.0 X 106 0.8 1.9 X 105 2.0
10 2.0 X 109 7.2 X 107 1.6 3.5 X 105 3.8
a Cellswereinfectedwith eachvirusat anMOIof 10, 3 hr apart.
lisheddata). Because thismutantproduces larger
amountsof viral RNA than does theparent, one
might predictthatmorereplicationsiteswouldbe
occupied and therefore agreater degreeof
inter-ference should result when it, instead of the parent, is usedastheinterferingvirus. Ets-4was indeed a much better interfering virus than its parentwhenVEE was used as a challenge virus
(Table 5). From doublyinfected cells,VEEvirus
wascounted in thepresenceofa 1:100 dilution of anti-VEEserum(titer,1:100,000).Theinterfering
virusinthis experimentwasincubatedat37 C for 3 hr before challenge virus was added. If the
initial 3-hr incubation was carried out at 30 or 42 C,at whichtherate andextent ofviral RNA
synthesisof Ets-4were depressed to levels closer to those found in the parentvirus,the degreeof
interference was likewise reduced (Table 6). It was notreducedto the level inducedbyEEE
be-cause, when incubation was resumed at 37C afteronly3hrateither of the other temperatures,
Ets-4 stillhadanadvantagein theextentofviral RNAsynthesis.
DISCUSSION
Several types of interference among viruses have been described (2) that do not involve the
mediationof interferon. Except forafew reports
(16, 19), all of the interference phenomena de-scribedrequiredthat theinterferingvirus be able to initiate certain synthetic processes in thehost cell. To demonstrate the interference between
arboviruses, itwas necessaryto givethe
interfer-ing virus a growth advantage either by previous
infectionof the host cells orby using high multi-plicities of the interfering virus relative to the
challengevirus. Thegreater thegrowth advantage given to theinterfering virus, the greater the
de-gree of interference. The interference, however,
didnotresult fromageneraldeterioration ofcell
metabolism due to infection by the interfering
virus. At least 40 hr ofinfection (at multiplicities
greater than 1) with EEE or VEE viruses was
requiredfor the cellsto showa cytopathic effect, even though peak titer was attained between 10 and 12 hr. Furthermore, in our hands these
TABLE 6. Effect of initialtemperature of
intcubation
on interference
intduced
by Ets-4virusaInitialtempofincubationb
Time after superinfection
37C 30C 42C
5 1.4 0.9 1.0
8 2.2 1.2 1.4
20 3.8 2.8 2.9
aExpressed as the
log,0
decrease of VEE virusin doubly infected cells compared to controls incubated with VEE virus alone.
bCultures infected with Ets4 virus were incu-bated at 37, 30, or 42 C for 3 hr prior to
super-infection with VEE virus. Incubation was then resumed at 37 C for all cultures.
viruses
causelittle
or noinhibition
oftotal RNAor
protein synthesis
inmonolayer
cultures of CE cells foratleast
12 hrafter
infection.
In contrast to the
interference described
byPohjanpelto and
Cooper(16),
the presence of thevirus
genomein
thecell
without
accompanying IRNAreplication
inoursystem was notsufficient
to
induce interference of
challenge
virus growth. When thegrowth
ofstrain
T was prevented byincubation
at 42 C, the growth of the challengevirus (EEE)
was notinhibited.
Itis known
that theblock in
thegrowth of
Tvirus
occursbecause
synthesis
of IRNAis inhibited
at 42 C. These data thereforeindicate
that theinterfering virus
genome must
be able
toreplicate
IRNAin
the cellin
order to prevent thegrowth
of thechal-lenge
virus.
Thedata
above
appeartosupportthehypothe-sis
of Cordsand Holland
(5)
thatinterference of
challenge virus
occursbecause ofcompetition
for replication sites or substrate necessary for viral replication. Thishypothesis
wasfurther strength-enedby
showing
that Ets-4was abetterinterfering
virus
than was EEE at 37C, a temperature atwhich it
produces
threetimesasmuchviral RNAas does theparent EEE
virus.
Atinitial
tempera-turesof
incubation
of 30 and42C,atwhich
Ets-4virus
begins
toproduce
viral RNA at rateson November 11, 2019 by guest
http://jvi.asm.org/
[image:5.484.50.442.65.148.2] [image:5.484.252.444.191.294.2]ZEBOVITZ AND BROWN
proaching
that of EEEvirus, Ets-4 interferes toa lesserextentwith VEE virus growth thanat37 C.Interference of challenge viruswasobservedin
one experiment in whichsuperinfection was car-riedoutwith the IRNA of challenge virus instead of thevirion, and inasecondexperiment in which
the inhibition of IRNA synthesis of challenge virusaftersuperinfection with the virion was evi-dent. Itseems reasonable toconclude, therefore, that interferenceprobably occurredatsomepoint after theentryanduncoatingof theviralnucleic
acid but beforetheinitiation of IRNAsynthesis. This conclusion is similar to that reported by Cords andHolland (5) forenteroviruses.
TheevidenceontherequirementforviralRNA
synthesistoestablishinterference, and the
inhibi-tionofsynthesisofIRNA ofchallenge virus,
sup-portthe interference mechanismbasedon a
com-petition for replication sitesormetabolites. Inter-ference of this type, although less directly stated
or supported, has beendescribed forarboviruses by Henderson and Taylor (9) andbyP.T. Allen
(unpublished data). Recent preliminary
experi-mentsindicate that various arbovirusescan
inter-fere with Newcastle disease virus and vesicular stomatitus virus, and vice versa, in the presence ofactinomycin D. We have not,however,
elimi-nated adsorption, penetration, or uncoating of
challenge virus in these combinations, nor have weyetfollowed viralRNA ofchallenge virus in
these systems. If, infact, the actinomycin
D-re-sistant interference isasbroadassuggested above, the major hypotheses reviewed by Bratt and Rubin (2) as alternatives to the competition hy-potheses would be eliminated because of the rela-tivespecificity required. Further experiments are needed, not only to determine how broad is the
interference
described here, but also to provide direct proof where possible of the competition hypotheses.If the kind of interference described in the
present paper is found to extend nonspecifically
tounrelatedviruses, then its role in vivoas a
non-specific mechanism ofresistance to virus disease would have to beevaluated, particularly in rela-tion to interferon-mediated interference. The
latter isusuallyassumed toplayadominant role
inmostofthein vivo interference that hasbeen described before and after thediscovery of inter-feron.
ACKNOWLEDGMENTS
ThispaperisdedicatedtoStewartA. Koser,onthe
occasion of his 75th birthday, in recognition ofhis distinguishedcareerinteaching andresearch.
Weexpressappreciation fortheexcellent technical assistance of Virginia Elsner, Delbert Davisson, Rosa Bell, and Cordia Harris.
LITERATURE CITED
1. Berge, T. B., I. S. Banks, and W. D. Tigertt. 1961. Attenuation of Venezuelan equine encephalomyelitis virus by in vitro cultivation in guinea pig heart cells. Am. J. Hyg. 73:209-218.
2. Bratt, M. A., and H. Rubin. 1967.Specific inter-ference among strains of Newcastle disease
virus. I. Demonstration and measurement of theintereference.Virology 33:598-608. 3. Brown, A. 1963. Differences in maximum and
minimumplaque-forming temperatures among selected group A arboviruses. Virology 21: 362-372.
4. Colon, J. I.,and J. B. Idoine. 1964. Factors affect-ing plaque formation by the infectious ribo-nucleic acid of the equine encephalitis viruses. J. Infect. Diseases 114:61-68.
5. Cords, C. E., and J. J. Holland. 1964. Interference between enteroviruses and conditions affecting its reversal. Virology 22:226-234.
6. Finter, N. B. 1966. Interferons, p. 340. W. B. Saunders, Philadelphia.
7. Hackett,A.J., F. L.Schaeffer,and S. H.Madin. 1967. The separation of infectious and auto-interfering particles in vesicular stomatitis virus preparations. Virology 31:114-119. 8. Heller, E. 1963. Enhancement of Chikungunya
virus replication and inhibition of interferon
productionbyactinomycin D. Virology 21:652-656.
9. Henderson, J. R., and R. M. Taylor. 1961. Studies on mechanisms of arthropod-borne virus interference in tissue culture. Virology 13:477-484.
10. Henle, W. 1950. Interference phenomena between animal viruses: areview. J. Immunol. 64:203-236.
11. Hoskins, M. 1935. Aprotectiveaction of
neuro-tropic against viscerotropic yellow fever virus in Macacus Rhesus. Am. J. Trop. Med. 15: 675-680.
12. Huang, A. S.,andR.R.Wagner. 1966. Defective T particles of vesicular stomatitis virus. II. Biologic role in homologous interference. Virology 30:173-181.
13. Isaacs, A., and J. Lindemann. 1957. Virus inter-ference. I. The interferon. Proc. Roy. Soc.
(London)Ser.B147:258-267.
14. McKinney, H. H. 1929. Mosaic disease in the Canary Islands, West Africa, and Gibraltar. J.Agr.Res.39:557-578.
15. Marcus, P. I., and D. H. Carver. 1967. Intrinsic interference: a new type of viral interference. J.Virol. 1:334-343.
16. Pohjanpelto, R., and R. D. Cooper. 1956. Inter-ference betweenpolioviruses induced by strains thatcannotmultiply. Virology 25:350-357. 17. Roizman, B. 1965. Abortive infection of canine
cells by Herpes simplex virus.
III.
The inter-ference of conditional lethal virus with an extended host range mutant. Virology27:113-117.
1288
on November 11, 2019 by guest
http://jvi.asm.org/
INTERFERENCE AMONG GROUP A ARBOVIRUSES
18. Schlesinger, R. W. 1959. Interference between animal viruses, p. 157-189. In F. M. Burnet
and W. M. Stanley (ed.), The viruses, vol. 3. AcademicPress,Inc.,NewYork.
19. Steck, F. T., and H. Rubin. 1966.Themechanism ofinterference betweenanavian leukosis virus
and Rous sarcoma virus. I. Establishment of
interference. Virology 29:628-641.
20. Taylor, J. 1964. Inhibition of interferonaction by
actinomycin. Biochem. Biophys. Res.
Com-mun.14:447-451.
21. Zebovitz, E. 1965. A defined maintenance medium forsupporting chick fibroblastmonolayersand for plaque formation by Venezuelan and eastern equine encephalitis viruses. J. Infect. Diseases115:77-82.
22. Zebovitz, E., and A. Brown. 1967. Temperature-sensitivestepsinthebiosynthesisofVenezuelan equine encephalitis virus.J.Virol.1:128-134.