0022-538X/80/07-0184/10$02.00/0
Integration of the Simian Virus
40
Genome into
Cellular
DNA
in Temperature-Sensitive
(N) and
Temperature-Insensitive
(A) Transformants of 3T3
Rat
and Chinese Hamster
Lung
Cells
ANA BERTA CHEPELINSKY,* ROLAND SEIF, AND ROBERT G. MARTIN
Laboratory of Molecular Biology,National Institute ofArthritis,Metabolism and DigestiveDiseases,
NationalInstitutesofHealth, Bethesda, Maryland20205
We studied thepattern ofintegration of the simian virus 40 (SV40) genome into the cellular DNA of N-transformants (temperature sensitive) and
A-trans-formants (temperature insensitive) derived from 3T3-Fisher rat and Chinese hamster lungcells.The SV40DNAwascovalently linkedtothecellular DNA in
both typesof transformants. In therat cells, most N-transformants contained
SV40sequencesintegratedatasingle site;mostA-transformants contained SV40
sequences integrated at two to five sites. In the Chinese hamster cells, no significant correlation between the number of integration sites and the phenotype of the transformantwasfound; onetothree integration siteswereobserved for both the N- and A-transformants. Single copies and tandem repeats of SV40
sequences were observed in A- and N-transfornants derived from rat cells. A-transformants arise neither by amplification of the SV40genome norby integra-tionataunique site.
Infection of mammalian cells by tsA mutants of simian virus 40 (SV40) and polyoma virus
gives risetotransformantsthat areeither tem-perature sensitive(N) or temperature insensitive
(A) (7-10, 12, 13, 17, 20-23, 26-28, 32). The
relative proportions of the two types of trans-formants seem todependupon several parame-ters, including thegrowth state ofthe cells for
thefirst few
days postinfection
(23, 26, 27) and the ability of the virus to express the small-tantigen (28).Although several models have been proposed (12, 14,23,26-28), the molecular
mech-anism(s)
responsible
forthe difference betweenthe two types of transformants has not been
found yet. In an attempt to gainsome insight
into these mechanisms, we examined the
inte-grationpatternsoftheSV40genome into
cellu-lar DNAin anumberof N- andA-transformed
rat and hamstercell lines.
Ithas beenreportedthat the numberof
inte-grationsitesandnumber ofcopies of theSV40
genome integrated per site varied from one
transformed cell line to another (5, 16). This
studywasundertakentodetermine
what,
ifany,correlations existbetween the temperature sen-sitivityof the tsAmutant-transformedcelllines
andthe number ofcopies of the SV40genome
integrated. Wefoundthat the SV40genome is
integratedinto host sequences in N-transform-ants aswell as in A-transformants and thatno specialpattem ofintegrationisassociated with the difference intemperaturesensitivity.
MATERIALS AND METHODS
Transformed cells. The N- and A-type tsA209,
SV40-transformed rat cells and the tsA209 and tsA209 viable deletion double-mutant-transformed Chinese hamsterlung (CHL) cellshave been described
previ-ously (20, 27, 28).
ViralDNA. TheSV40form I DNA used for nick translation and also toobtain form II and III DNAs thatwereusedasstandardswaspurchasedfrom Be-thesda Research Laboratories.
Isolation of high-molecular-weight cellular DNA.Ratcells orCHL cellsweregrowntoconfluence
in Dulbecco-Vogt modified Eagle medium
supple-mented with 10% fetal calf serum. After cells were
washed with phosphate-buffered saline, they were
treated with lysisbuffer(10mM Tris-hydrochloride,
pH 7.9,1 mM, EDTA,0.5%sodiumdodecylsulfate),
incubated with 0.5mgofproteinase K (Boehringer)
perml overnight at 370C, and extracted twice with
phenol andoncewithchloroform-isoamylalcohol(24:
1,vol/vol).Theextractswerethendialyzed,incubated
with 0.1 mgof RNase A (Boehringer) per ml, incu-bated again with proteinase K (0.1 mg/ml),
reex-tracted, anddialyzed,asdescribedbyBotchanet al.
(4). The molecularweightof the DNAwaschecked byagarosegel electrophoresis.
Restriction enzymes, digestion, and agarose
gels.Cellular DNA(25 ,Lg)wasdigestedfor 2to5 h
with restrictionenzymesby usingtheincubation
me-dia specified by New England Biolabsor Bethesda
ResearchLaboratories, dependingonthesourceofthe enzyme. Afterincubation, 20mM EDTA and 0.1to
0.2M NaCl (final concentrations) were added. The
samples were extracted once with phenol and once
withchloroform-isoamylalcohol (24:1). Sodium
ace-184
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATION OF SV40 GENOME INTO CELLULAR DNA 185
tate(pH 6.0;finalconcentration,0.2M)and 2volumes
of absoluteethanolwereadded,and the solutionswere
stored overnightat-20°Cand then centrifugedina
Brinkmancentrifugefor15min.Theprecipitateswere
suspendedinasolutioncontaining10 mM
Tris-hydro-chloride(pH 7.8),6 mMEDTA,0.2mgof bromophe-nol blue per ml, and 7% glycerol and resolved by
electrophoresis in a 0.7% agarose (Marine Coiloids)
horizontal slab gel in a solution containing 40 mM
Tris-acetate (pH 7.8), 5 mM sodiumacetate, and 1 mM EDTA (31) for 16 to 18 h at 40 V at room
temperature. LambdaphageDNAHindIIIfragments
labeledwith32P attheir 5' ends(BethesdaResearch
Laboratories) were used as DNA molecular weight
standards.
Blotting.Transfer of DNA from thegelsto
nitro-cellulosepaperwasperformedbythe Southern
tech-nique(30)withsomemodifications (5, 16).Gelswere
stained with 2.5 pgof ethidium bromideperml,and
picturesweretaken underUVillumination. Thegels
werethen treated with 0.2 M NaOH-0.6MNaCl for 45 to 60min, neutalizedwith 1 MTris-hydrochloride
(pH7.3)-0.6MNaCl for 45min,andequilibratedwith
6xSSC (lxSSC is 0.15 M NaClplus0.015M sodium
citrate)for 30 to 45minwithgentlerotation.Blotting
onnitroceliulose membrane filterpaper(0.45pm;type
HAWP; Millipore Corp.) was performed downward
(gelatthetop)forapproximately3h andupwardfor
approximately20h, soakingthe 3 MMpaperbridgein
6x SSC. The nitrocellulose paper wasrinsedfor 15
minin 6x SSC and air dried. Itwasbaked for2 h at
800Cunderavacuumandkeptatroomtemperature until itwastobe used.
RadioactivelabelingofSV40DNA. Nick trans-lationwasperformedbythe method of Maniatisetal.
(18),with modificationssuggested byShoshanaSegall
(personal communication). The incubation mixture
contained [a-3P]dATP [tetra(triethylammonium)
salt]and[a-32P]dCTP[tetra(triethylammonium) salt]
(300to500Ci/mmol [New EnglandNuclearCorp.]or
approximately 2,000 to 3,000 Ci/mmol [Amersham
Corp.] previouslydried),each ataconcentration of 3.6
pM; the mixture also contained 3.6 pM unlabeled
dGTP, 3.6 pMTTP,50mMTris-hydrochloride (pH
7.8),3 mMMgCl2,10 mM,B-mercaptoethanol,50 pgof
bovineserumalbuminperml,and 10 pgof SV40 form I DNA(BethesdaResearchLaboratories)perml. The mixturewaspreincubatedfor 15minat150C.
Esche-richia coli DNApolymeraseI(40U/ml;Boehringer)
and 1.2 ng of DNase I (Worthington Biochemicals
Corp.) per ml were thenadded, andthe incubation
wascontinued for 110to120min (untilaplateauwas
reached in theincorporationof3p into trichloroacetic
acid-precipitable material). A solutioncontaining 10
mMTris-hydrochloride (pH 7.5),0.1MNaCl,10mM
EDTA, and 0.5% sodiumdodecylsulfate wasadded;
the sample was extracted with phenol and passed
through a Sephadex G-50 fine column equilibrated
with 10 mMTris-hydrochloride (pH 7.5)-0.1M
NaCl-1 mM EDTA, and the fractions that contained the
labeled DNAwerepooled.Anapproximate yield of1.5 x 10Wto6x108cpm/pugofDNAwasobtained.
Hybridization.Blottednitrocellulosepapers were
soaked in 3xSSC forapproximately45minatroom
temperature.Theywerethenpreincubatedfor 3 h in
3xSSC-10x Denhardt solution (lx Denhardt
solu-tion contains 0.02% bovine serum albumin, 0.02%
Fi-coll, and 0.02%polyvinylpyrrolidone)at68°C and later
with a solution containing 3x SSC, 1OX Denhardt
solution, 0.1% sodiumdodecyl sulfate, 10 pg of
poly-adenylic acid per ml, and 100pg of sheared and
dena-turedsalmon sperm DNA per ml for1hat680C. The
hybridization mixture contained 3x SSC, lOx
Den-hardt solution, 0.1% sodium dodecyl sulfate,10 pg of
polyadenylic acid per ml, 100 pig of sheared and
dena-tured salmon sperm DNA perml,and 0.9x 107to1.2
X 107cpm of denatured32P-labeledSV40 DNA
(ob-tainedby nicktranslation; concentration, 20 to 90 ng/
ml) per ml in a total volume of 7 to 14 ml; the
hybridizationwasinsealedbagsat680C for
approxi-mately 40 h. Afterhybridization, therewere four to
fivewashings withasolutioncontaining3xSSC,1Ox
Denhardtsolution, 0.1% sodium dodecylsulfate, and
50pg of sheared and denatured salmon sperm DNA
per ml and onewashing with a solution containing
0.lx SSC, 50 pg of denatured and sheared salmon
sperm DNAperml,and 0.1% sodiumdodecylsulfate
at680C; the filter paperwasthen rinsedoncewith3x
SSCat roomtemperature(15). Milliporefilterpapers
wereairdried andexposedtoKodak X-Omat R film
with Max IPickerscreens at-70°Cfor2 to 9days.
RESULTS
Integration of theSV40genomein N- and A-type tsA209 and tsA2O9 viable deletion
double-mutant-transformed CHL celis. High-molecular-weight cellular DNA from CHL cellstransformed bythe tsA209and tsA209 vi-able deletion double mutants ofSV40 was di-gested with restriction enzymes that cut the cellular DNA but not SV40 DNA. One of the limitations of the Southern technique is that the transfer efficiency of pieces of DNA
approxi-mately20,000 base pairs long or longer is lower than that of smaller pieces (1, 5) and may go undetected. Therefore, several restriction en-zymes were used, and the highest number of bandsobtainedwith any of them was considered the minimumnumberofintegration sites. The enzymes used were BglII,SacI,SstI (isoschizo-merofSacI),and XbaI.
Figure 1shows the patterns of integration of SV40 into the cellular DNA of several N- and
A-type CHL cell lines transformed either by
tsA209 or by one of several double mutants. Sincehamstercells are semipermissive forSV40, the cells were grown at
410C
in orderto avoid the presence of free SV40 DNA thatoccurs at330C (19). No form I or II SV40 DNA was observed.
Table 1shows theminiimnum number of
inte-grationsites and the sizes of thedifferent bands
obtained with each restriction enzyme for the tsA209 N-andA-transformants. Fromthe sizes oftherestrictionfragments itispossible to
cal-culate the maximum number of copies of the SV40 genome possible per integration site; i.e.,
VOL. 35,1980
on November 10, 2019 by guest
http://jvi.asm.org/
186 CHEPELINSKY, SEIF, AND MARTIN
A 2 3 4 5 6 7 8 9
lo
11} 3~~~~~~~~~~~~~~~~~~
B | 2 3 4 .5
_ W*W
* .:.*
FS-... p>. ^
..:.eF...
.... ^ * .. r.: .;. ,¢
X,
..
*.
...
i.. i z :..6..*, ..:.:f~~~~~~~~~~~~:
I ! -Oo
[image:3.508.63.460.62.333.2]
-s4-0
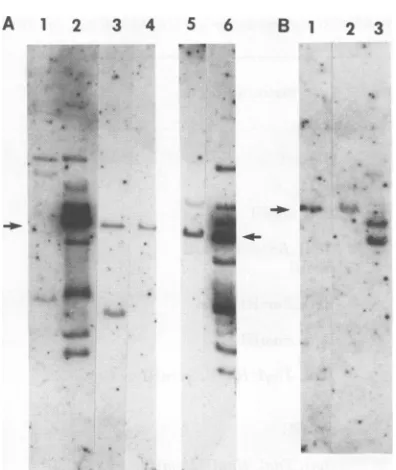
FIG. 1. HybridizationpatternsofCHLtransformed ceUlineswithSV40DNAno-cutenzymes.(A)Cellular
DNAwasdigested withBglII,runingels,and hybridizedasindicated in thetext.Lane1,32P-labeledADNA
HindIII fragments (numbers ofbasepairsperbandareindicatedbyarrows;I, II, and III,SV40 form I, II,
andIII DNA);lane2, CHLA209L4;lane 3,CHLA209L5; lane4,CHLA209L38; lane5,CHLA209L49;lane6,
CHLA209L62; lane 7, CHLA209L51; lane 8, CHLA209A289L5; lane 9, CHLA209A289L4; lane 10,
CHLA209A88L3;lane11,SV40forn II,III,andIDNAs. (B) XbaI digestion ofCHL cellular DNA. Lane1,
CHLA209L51; lane2, CHLA209L38; lane3, CHLA209A29OL4; lane4, standardsasin (A), lane 1; lane5,
CHLA209A292L14.
CHLA209L4andCHLA209L5must have fewer
than two full-size SV40 DNA copies each,
whereasCHLA209L38 andCHLA209L62 must
have fewerthanthreecopiesandCHLA209L49 musthave fewer than fourcopies.One to three
integration siteswereobserved in the different N-and A-transformed celllines, andthepatterns varied both between the N- and A-types and amongthe differentN-and A-transformants.
N-and A-transformantsofCHLcells induced
by tsA209 viable deletion double mutants (12)
werealso examined.Thenumber ofintegration
sitesvariedfromonetothree,and themaXimum number ofcopies of the SV40genome per
mte-gration site varied from fewer than two to
fewer than six (Table 2). In cell lines
CHLA209,6292L14,
CHLA209A287L5, andCHLA209A292L19 bands of similar molecular
weights(insidethelimits oferror)wereobserved after digestion with XbaI. When these results
werecombinedwith the results withthe tsA and
double-mutant transformants, 4 of 12
N-trans-formants appearedto have arisen from the
in-sertion of SV40 ata single site, and 2 of10
A-transformantsarosesimilarly. Itmaybe
signifi-cantthatof11celllinesestablishedat multiplic-itiesofinfection from 0.065to0.48(Tables1and 2), 7 appeared to have SV40 integrated at a single site, whereas of12celllinesestablishedat
multiplicities of infection from 0.7to3,nonehad SV40integratedatonlyasingle site.
In some of thecelllinesthatappeartohave SV40 DNAintegratedatonlyonesite, the cel-lular DNAwastreated with restrictionenzymes
that cut the SV40 genome once (e.g., BglI,
TaqI, BamHI, EcoRI). Thepresence oflinear, full-length SV40 DNA (formn III) after this treat-mentwould indicatearepetition of the relevant site. The flanking sequences bound to cellular DNAmay ormaynotbedetected accordingto
their sizes and/or the size of the SV40 region attached (Table 3). No evidence formorethan asingle full-length insertionwasfoundfor sev-eral N-type cell lines (e.g., CHLA209L5 and CHLA209L62),whereasrepetition of the TaqI, BamHI, and EcoRI sequences wasobserved in CHLA209L49,anA-typeline.
IntegrationoftheSV40genomein N-and
J. VIROL.
-4-0
J:io
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATION OF SV40 GENOME INTO CELLULAR DNA 187
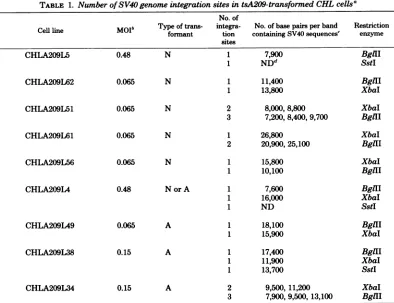
TABLE 1. Number of SV40 genome integration sites intsA209-transformedCHLcellsa
No. of
Cellline Molb Type offormanttrans- integra-tion containing SV40No. of base pairs persequences'band Restrictionenzyme sites
CHLA209L5 0.48 N 1 7,900 BglII
1 NDd SstI
CHLA209L62 0.065 N 1 11,400 BgllI
1 13,800 XbaI
CHLA209L51 0.065 N 2 8,000,8,800 XbaI
3 7,200, 8,400, 9,700 BglII
CHLA209L61 0.065 N 1 26,800 XbaI
2 20,900, 25,100 BgIII
CHLA209L56 0.065 N 1 15,800 XbaI
1 10,100 BgllI
CHLA209L4 0.48 N or A 1 7,600 BgII
1 16,000 XbaI
1 ND SstI
CHLA209L49 0.065 A 1 18,100 BglII
1 15,900 XbaI
CHLA209L38 0.15 A 1 17,400 BglII
1 11,900 XbaI
1 13,700 SstI
CHLA209L34 0.15 A 2 9,500, 11,200 XbaI
3 7,900,9,500, 13,100 BglII
aDNA fromCHLtsA209-transformed
cell lines
wasincubated with restriction enzymesasdescribed in thetext.
bMOI,Multiplicityofinfection(see reference 20).
'The number of basepairswasdeterminedby using3P-labeled A DNA
HindIII
fragmentsasstandards(seeFig. 1). Theerrorinthedetermination of basepairs,whichwasbasedondataobtained in differentexperiments,
wasapproximately200basepairs forfragments smaller than 9,500 base pairs; however, forfragments10,000to
30,000 basepairslong itwasonthe order of 1,000 basepairs insome casesduetothehigherslopeof thecurve
ofmolecularweightversusmobilityin thatregion(2, 5).
d ND, Not
determined.
A-type tsA209-transformed rat celis. The
integrationof theSV40genomeintothecellular
DNAs of N- and A-transformed rat cells was alsostudied. Someof the patterns ofintegration obtainedwiththe rattransformantsare shown in Fig. 2. Asexpected, nofreeSV40DNAwas observed since rat cells are nonpermissive for SV40.Three of theA-typecell lines had similar
integration patterns withtwo different
restric-tionendonucleasesthatdo not cutSV40 DNA.
Cell lines A209-FR3T3-A30 and
A209-FR3T3-A37wereobtainedfromthesameflask;it cannot
berigorously excluded theyarenotderivedfrom
the same transformed cell. However,
A209-FR3T3-A43 was isolated in a separate
experi-ment and must therefore have arisen by an independentevent.
Table4 showstheresults obtained with
sev-eral restriction enzymes that do not cut SV40 DNA.Amongthe
N-transformants,
one to threeintegrationsites were observed, and among the
A-transformants one to five integration sites were observed. For the N-transformants there was a predominance of single-integration-site
patterns (5 of8), whereasfor the A-transform-antsmultipleintegration sites predominated (9
of10).
Insertions of sizes smaller than full-length
SV40 DNA have been observed, as in
A209-FR3T3-A99. In a long fihn exposure of A209-FR3T3-A43 digested with anSV40DNA no-cut enzyme, a light band of 4,000 base pairs was
observed,whichmay have been due tothe pres-ence of a small piece of SV40 DNA in that
fragment.
A number ofthecell lineswere examined after
VOL. 35,1980
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Numberof SV40 genome integration sites in viable deletion tsA209double-mutant-transformed
CHL cells
Cellline CHLA209A288L3
CHLA209A289L14
CHLA209A29OL5
CHLA209A29OL12
CHLA209A291L4
CHLA209A292L14
CHLA209A292L16
CHLA209A287L5
CHLA209A287L6
CHLA209A289L5
CHLA209A290L4
CHLA209A291L2
CHLA209A291L13
CHLA209A292L19
Type of No.of in-MOja transform- tegration ant sites
2.3 N 1
2 2
3 N 2
2
1 N 2
3
0.14 N 1
1
1.6 N 1
1 2
2.9 N 2
3
2.9 N 2
2
2 A 2
3
0.9 A 2
2
2 A 2
0.7 A 3
3
2.4 A 2
3
0.3 A 1
2
No. of base pairs per band con-tainingSV40sequencesb
16,100 19,500, 23,000 12,900, 16,500
6,600, 13,200 9,400, 18,000
NDC
7,900, 20,000, 28,100
19,000 19,200
12,200 28,600 10,700, 17,000
ND
10,400, 15,500, 21,000
15,300, 19,700 17,300, 25,000
19,800, 23,000 10,800, 15,500, 23,700
19,500, 24,500 17,000, 24,000
9,100, 11,600
7,100, 14,500, 23,700 10,500, 13,800, 17,000
11,300, 21,500 ND
8,000 ND
0.9 A 2 ND
3 10,800,15,200, 22,900
aMOI, Multiplicity of infection (see reference20).
'See
Table 1,footnotec.'ND, Notdetermined.
digestionwithrestrictionenzymesthatcutSV40
DNAonce.Figure3shows thepatternsobtained
after digestion ofcellular DNA from somecell
lines with two of these enzymes, and Table 5
summarizes all of theseresults. Among the
N-transformants withoneintegration site,the
fol-lowingobservations weremade. (i) One (A209-FR3T3-N94)contained fewerthan 1.5SV40
ge-nomic equivalents (sequence repetitions of the
BglI,BamHI,andTaqIsiteswerenotpresent).
(ii) Three transformants contained partial
re-peats of theSV40genome,asdemonstratedby
thefactthatsomerestriction enzymesthat cut
SV40DNAoncegaverise tofull-lengthformIII
DNA,whereas others did not. And(iii)no
trans-formant contained a complete duplication. In
theonlyA-transformantcontainingone
integra-tion site(A209-FR3T3-A12)fewerthan 1.4
cop-ies of SV40 DNA were present (BglI, TaqI, HpaII, and BamHI did not release form III
SV40 DNA). In the cell lines withmore than
oneintegration site,sevenof ninecontained at Restriction
enzyme
SstI XbaI BglII
BglIl XbaI
Bgll
SstI
XbaI
BgllI
BglII
XbaI SstI
BgllI
XbaI
BglII
XbaI
BglII XbaI
XbaI
BglII
BglII
XbaI
BglII
XbaI
BglII
XbaI
BglII
BglII
XbaI
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATION OF SV40 GENOME INTO CELLULAR DNA 189
leastsomepartial repetition.
notdistinguish from which
DNAwasderived.
DISCUSSII
Integration of SV40 D genome of transformed CHL and 3T3 rat transforn
TABLE 3. Presence ofsingi
repeats ofSV40 sequencesii genome oftransforrm
Type of Forn III Cellline trans- SV40
formant DNA" CHLA209L5 N
CHLA209L62 N _
CHLA209L4 Nor A
CHLA209LA9 A +
'Presence (+) or absence (-) released after digestion with the e table.
A I 2 3 4 B I
VW
i _ ...p
FIG. 2. Hybridizationpattei celllines digested with SV40 (A) XbaIdigestion of 3T3rat tr
1,A209-FR3T3-A30; lane2,A2
A209-FR3T3-A43; lane 4, SV4 DNAs. (B) SacI digestion of cells. Lane1,A209-FR3T3-A37
A30; lane 3,A209-FR3T3-A43; larweight standards (see Fig.
However,wecould contained fromone tofiveintegration sites. For
of the sitesform III the CHL transformed lines, most contained SV40 DNA integrated atmultiple sites.
How-ON ever, among those lines generated at initial mul-tiplicities of infection of <0.48 PFU/cell, the
)NA into the
cell
majority had SV40 sequences integrated at acells.
Among the single site (7 of 11), whereasallofthosegener-nants examined, all ated at multiplicitiesofinfectionof>0.7 PFU/
cellhadSV40 sequencesatmore than one site lecopies or tandem (12 of12). For the 3T3 rat transformants, which
ztegrated
inthecell weregenerated at initial multiplicities ofinfec-ed CHLcells tionof from 5 to100PFU/cell
(i.e.,
at multiplic-ities of infection higherthanusedwith the ham-Restrictionenzymes ster cells), most contained SV40 integrated atmultiplesites (12of18),but asignificantnumber
TaqI,BamHI, EcoRI, hadSV40 integratedatasingle site. (Botchanet BglI al. [5] alsoobserved apredominanceofmultiple
TaqI,BamHI, EcoRI
integration
sites inSV40
rattransformants.)
TaqI,BgtIBamHI, EcoRI, Thus,it appears that in ratcells
single-site in-TaqI,BamHI,EcoRI
tegrations (6 of 18) are more common at highofSV40
forn
III DNA multiplicities of infection than they are inham--nzymesindicated on the stercells(0 of
12).
This differencemayrelatetothe fact that hamster cells are
semipermissive
andratcells arenonpermissive.
Among thecell 2 3 4 linesthatappeared
to contain SV40integrated,, at asingle site,somemaycontain onlya
single
copy of the SV40 genome
(CHLA209L4,
CHLA209L5,
CHLA209L62, A209-FR3T3-N94,A209-FR3T3-A12), whereas others clearly
con-s-
_ >? tainedpartial repetition
of the SV40 genometi7
(A209-FR3T3-N11,
A209-FR3T3-N53,
A209-WE
wJ *FR3T3-N30) and still others maycontain
mul-'AM
9-
500 tiple repetitions of the SV40 genome*^0*
(CHLA209L49).
0
'f Some
explanation
isrequired
for the fact that
4-_, transformed celllinesthatcontain
papovavirus
-l sequences at asingle site frequentlyhavepartialv
4o
300repetition
of thepapovavirus
genome.Tandemrepeats of viral sequences have beenobserved before in
polyoma
virus-transformedratcells(a
iS _~-
semipermissive
system) (2)
and inSV40-trans-formed rat cells (a
nonpermissive system) (5).
Weobserved tandemrepeatsofSV40sequences *4- 330
in oneof four CHL lines (asemipermissive
sys-.4-'°0°tem)
andinthreeof fiveratlines(anonpermis-sive system). At least one mechanism for the generation of tandemrepeatsisvia
rolling
circlereplication. Rolling
circlereplication
inpermis-sive cells has been
demonstrated
forpolyoma
virus (3) and SV40 (R. G. Martin and V. P.
rns
of
transformed
rat Setlow,Cell, in press) andsuggestedinnonper-DNA
no-cut
enzymes-
missive cells forSV40
(W.
Chia andP.
Rigby,
ansformedcells. Lane personalcommunication).We
speculate
thatthe0 form II
III7
and
Igeneration
oflnear
moleculesisprerequisite to3T3 rat
transformed
integration
within
the host genome and that l-lane2, A209-FR3T3-rolling
circlereplication
is thepreferred
mannerlane4, DNAmolecu- for
generating
linear molecules. Othermecha-1). nisms forgenerating tandemrepeats have been
VOL. 35,1980
on November 10, 2019 by guest
http://jvi.asm.org/
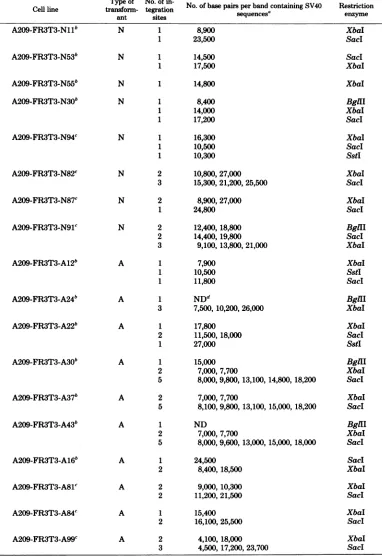
[image:6.508.54.249.153.568.2]T Cellline tra A209-FR3T3-Nllb
A209-FR3T3-N53b
A209-FR3T3-N55b
A209-FR3T3-N30b
A209-FR3T3-N94C
A209-FR3T3-N82C
A209-FR3T3-N87c
A209-FR3T3-N91c
A209-FR3T3-A12b
A209-FR3T3-A24b
A209-FR3T3-A22b
A209-FR3T3-A30b
A209-FR3T3-A37b
A209-FR3T3-A43b
A209-FR3T3-A16b
A209-FR3T3-A81C
A209-FR3T3-A84c
A209-FR3T3-A99C
Iype of No. of in-nsform- tegration ant sites
N 1
1
N 1
1
N 1
N 1
1 1
N 1
1 1
N 2
3
N 2
1
N 2
2 3
A 1
1 1
A 1
3
A 1
2 1
A 1
2 5
A 2
5
A 1
2 5
A 1
2
A 2
2
A 1
2
A 2
3
No. of base pairs per band containingSV40 sequences'
8,900 23,500
14,500 17,500
14,800
8,400 14,000 17,200
16,300 10,500 10,300
10,800,27,000 15,300,21,200,25,500
8,900,27,000 24,800
12,400,18,800 14,400,19,800
9,100,13,800,21,000
7,900 10,500 11,800
NDd
7,500,10,200,26,000
17,800 11,500,18,000 27,000
15,000 7,000, 7,700
8,000, 9,800, 13,100, 14,800, 18,200
7,000, 7,700
8,100, 9,800, 13,100, 15,000, 18,200
ND 7,000, 7,700
8,000, 9,600, 13,000, 15,000, 18,000
24,500 8,400,18,500
9,000,10,300 11,200,21,500
15,400 16,100,25,500
4,100,18,000 4,500, 17,200, 23,700
Restriction enzyme
XbaI SacI
SacI XbaI
XbaI
BglHl XbaI SacI
XbaI Sacl SstI
XbaI SacI
XbaI SacI
BglII SacI XbaI
XbaI SstI SacI
BglII XbaI
XbaI Sacl SstI
BglII XbaI SacI
XbaI Sacl
BglII XbaI Sacl
SacI XbaI
XbaI SacI
XbaI SacI
XbaI Sacl
aSee Table 1,footnotec.
bThese
cell lines
arethesame asthosedescribedinreference27,but withthe letterL,M,orHinstead ofNorAbefore the lasttwodigits.
cThese celllineswereisolatedasdescribed in reference28.
dND,Notdetermined.
190
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.508.77.459.52.608.2]INTEGRATION OF SV40 GENOME INTO CELLULAR DNA 191
A 1 2 3 4
Y :::::
s:::A;'
*.;s...
... ,,l *: .. pffl Z ..Ak..W, S p.
w4 *4hy.7r;fs >
w
-*:b.
4
F
* _F *A,. W,2..
!I_
_ b
* .4,,.5
..:
i-.
**.:.o::.#
_*
_
5 6r1
-:
0
B 1
FIG. 3. Hybridizationpatternsof
transft
cell linesdigestedwithSV40 DNAone-cut
(A) BglIandTaqI digestions of3T3ratcell
A209-FR3T3-N53TaqI digestion; lane2,.
lecularweight standards(see Fig. 1); lan4 FR3T3-A22TaqIdigestion;lane4,A209-Fi
BglI digestion; lane5, A209-FR3T3-A22 B
tion; lane 6,DNAmolecular weightstanc
Lane 1, SV40form IIIDNA;lane 2, A20
N30 TaqI digestion; lane3, A209-FR3T3-,
digestion.Arrows indicateposition of form
DNA.
proposed (2, 6).
Theresultsobtained with theviablE doublemutantsindicate thatsmall-te
notrequired for integration of the SV4i into thecellularDNA. Furthermore,sI
tigen does not seem to affect the pa integration; i.e., both single-site and siteintegrationswerefoundwhether c viruswas capable of expressing thesxi
tigen.
The conclusion thatSV40 DNA do(
tegrate atspecific sites in the cell gen
basedontheobservationof differentpi integration in different cell lines(5).Hz is important to point out that integ
specific sequences cannot be ruled possibletogetdifferent restrictionpal
spite integration in thesame geneif (i)
amounts ofhost DNA are deleted ur
integration, (ii) differentamountsofSI areintegrated in thesamegene, or(iii
tionoccursatdifferent portions(restric
ments) of thesamegene. Furthermor
2 3 DNAcontainshighly repeatedsequenceswhich exist repeated in tandem or interspersed with unrelatedsequences(29). TheSV40genome can recombine with thesehighly repeatedsequences (24, 25).Differentpatternsof integration of the
viral DNA in different cell lines may be
ex-plained by integration in a specific highly
re-peatedsequencethatisinterspersed with unre-latedsequences.
Alternatively,
ifthe viral DNAintegrated in a sequence repeated in tandem,
j* identical bands would be observedin different
* W cell
lines,
even whenintegration
hadactually
occurred at different sites. Three rat
trans-formed
cell
lines (at least two of which wereobtainedindependently) presented thesame
in-*
tegration
pattern withtwodifferent SV40no-cutenzymes. It remains to be established whether
multiple
bands in asingle
cell line ariseby
independent
events.Integration
of the SV40 genomeinto thecellular
DNA ofN- and A-transformants. There was some suggestion thatN-transform-ants were more
likely
thanA-transformnants
tocontain SV40 at a single insertion site. Taken
ornmed
rat together,9of20N-transformants
and3of20A-IenLanmes.
transformants contained SV40 DNAintegrated DNA mo- at asingle
site. For the CHL cells 4 of 12N-e3,
A209.
transformants
and 2 of 10A-transformants
con-R3T3-N53 tained
single-site insertions;
for theratcells 5 of gilI diges- 8N-transformants and 1of10A-transformants lards. (B) contained single-site insertions.)9-FR3T3- Among the lines with only one integration site
A30
TaqI
we foundA-transformants
with fewer than 1.4III
SV40
copies (A209-FR3T3-A12) andwithmore than1.5copies
(CHLA209L49)
andN-transformants
with fewer than 1.5 copies (CHLA209L5, CHLA209L62, A209-FR3T3-N94) and withadeletion more than 1.7
copies
but fewer than 2copies
antigen is(A209-FR3T3-N30).
The maximum andmini-0genome mum numbersarebased on the
assumption
of mall-tan- tandem repetition of the integrated sequencesLtterns
of andcomefrom theability
orfailure of theSV40
multiple- DNAone-cutenzymetorelease
full-length SV40
)r notthe DNA. No correlationbetweencopynumber and nall-tan-
phenotype
isapparent, since of those testedtwo CHL N-transformants have fewer thantwo cop-es notin- iesandoneA-transformantmayhavemorethaniomewas two
copies,
whereas for the rat cells threeN-atterns
oftransformants
have fewer thantwocopies
andowever, it one may have a
single
copy, but the oneA-ration at transformantalso may havea
single
copy. out. It is Several conclusionscanbe drawnconcerning
tternsde- the
relationship
between SV40 genome copy different number and the temperaturesensitivity
ofthe2on SV40
resulting
transformants.(i) N-transformants
doV40 DNA not arise from the failure of
SV40
to become)integra-
integrated.
(ii)
A-transformantsdonotariseby
tionfrag- gene
amplification
orby
asimple
genedosage
e,cellular effect. And
(iii)
neither A-norN-transformantsVOL. 35,1980
4" ft"
on November 10, 2019 by guest
http://jvi.asm.org/
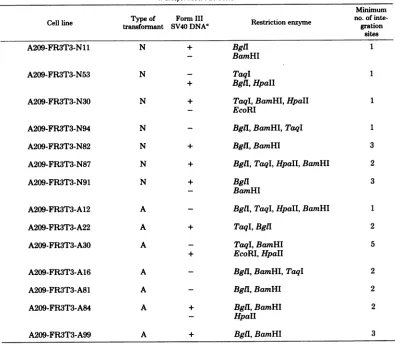
[image:8.508.47.245.74.309.2]TABLE 5. Presenceof singlecopiesortandem repeatsofSV40 sequences integrated in the cell genome of transformed rat cells
Minimum
CeRline Typeof Form III Retito
nyeno.
ofinte-Cell line transformant SV40DNA0 Restriction enzyme gration sites
A209-FR3T3-N11 N + BglI 1
- BamHI
A209-FR3T3-N53 N - TaqI 1
+
BglI,
HpaII
A209-FR3T3-N30 N + TaqI, BamHI,HpaII 1
-
EcoRI
A209-FR3T3-N94 N - BglI, BamHI, TaqI 1
A209-FR3T3-N82 N + Bgll, BamHI 3
A209-FR3T3-N87 N + BglI, TaqI, HpaII, BamHI 2
A209-FR3T3-N91 N + BglI 3
- BamHI
A209-FR3T3-A12 A - BglI, TaqI, HpaII, BamHI 1
A209-FR3T3-A22 A + TaqI, BglI 2
A209-FR3T3-A30 A - TaqI, BamHI 5
+
EcoRI,
HpaII
A209-FR3T3-A16 A - Bgll, BamHI, TaqI 2
A209-FR3T3-A81 A - Bgll,BamHI 2
A209-FR3T3-A84 A + Bgll, BamHI 2
- HpaII
A209-FR3T3-A99 A + Bgll, BamHI 3
aSeeTable3andFig.3.
haveunique integration patterns.
One
possible
mechanism to explain theN-phenotypewasthatSV40
replicated
as aplasmidin N-transformants. This possibility is
elimi-nated by the finding that all N-transformants containedintegrated SV40sequences.
Asecondpossible explanationfor the
temper-ature-insensitivephenotype ofthe
A-transform-antsis thatT-antigenisrequired for the main-tenance of transformation in all transformed cells but that high levelsof thethermolabile
T-antigen resultin"leaky"expression of the
trans-formed phenotypeattherestrictivetemperature
(7,11,33).Itisstillpossible thatin A-transform-ants T-antigen accumulates more than in N-transformants. However, we haveeliminatedthe
possibilitythatsuchleakinessinvariablyresults
from the integration of multiple copies of the
SV40genome. It cannot, forexample,beargued
that all
A-transformants
contain multiplere-peatsoftheearly regionofSV40 andthateven
those A-transformants with SV40 sequences at
asingle site containmultiple repeats of theSV40
genome. Cell line A209-FR3T3-A12 contains
fewer thantwo copies of theSV40early region,
and cell lines A209-FR3T3-A16 and
A209-FR3T3-A81 contain a maximum of two
func-tional early regions. Except for CHLA209L49,
wehave not found anA-transformant that
re-leasedunit-lengthformIII SV40DNA withall
of the SV40 DNA one-cut restriction enzymes
assayed. Thisallows us to rule out the possibility of more than five or six copies of the SV40
genome tandemly integrated (gone undetected
by SV40DNA no-cutrestrictionenzymes due to thelimitations of the Southern technique). We conclude that gene dosage alone cannot explain theA-transformants.
Another
possible explanation
for the A-trans-formants isthattheyall result from mutationofagrowth controlgeneby
integration
of the viral genome into it.T-antigenwould not berequiredfor the maintenance of transformationin these
cells, in contrast to N-transfornants, inwhich
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.508.66.461.99.444.2]INTEGRATION OF SV40 GENOME INTO CELLULAR DNA 193
therole ofintegrationof theSV40genomeis to
permanently provide large T-antigen (27, 28).
Certainly with thisapproach we could discern nouniquepatternofintegrationin
A-transform-ants which distinguished them from N-trans-formants. Furthermore, there was no readily
discemable common element to either the A-typeorN-typeintegrationpattern.We have not eliminatedthepossibilitythatA-transformants
result from integration of the viral genome at
any of a small number of control genes. Rat transformants withasingleintegrationsitewere
frequent among N-transformants but rare amongA-transformants. Ifgrowthcontrolgenes
representaminority of the cellulargenome, (i)
A-transformants would arisepreferentiallyfrom
cells in which the viral genome was able to
integrate atseveralsites, and (ii)
N-transform-antswould arise more frequentlyfrom cellsin
which the SV40genomewasabletointegrateat onlyonesite.
LITERATURE CITED
1. Basilico, C., S. Gattoni, D. Zouzias, and G. Della Valle.1979.Loss ofintegrated viral DNAsequencesin polyoma-transformedcellsisassociatedwithanactive viralAfunction. Cell 17:645-659.
2. Birg, F., R. Dulbecco, MLFried,andR. Kamen.1979. State and organization ofpolyoma virus DNA se-quencesintransformedratcelllines.J.Virol.
29:633-648.
3. Bjursell, G.1978. Effectsof2'-deoxy-2'-azidocytidineon
polyomavirus DNAreplication: evidence for rolling circle-type mechanism. J. Virol.26:136-142.
4. Botchan, M., G. McKenna, and P. A. Sharp. 1973. Cleavage ofmouseDNAbyarestrictionenzyme as a
cluetothearrangementofgenes.ColdSpring Harbor Symp. Quant.Biol. 38:383-395.
5. Botchan,M.,W.Topp,and J. Sambrook. 1976.The arrangementof simianvirus40sequencesin the DNA
oftransformedcells.Cell9:269-287.
6. Botchan, M., W. Topp, and J. Sambrook. 1978. Studies
onsimianvirus 40excision fromcellular chromosomes. ColdSpring Harbor Symp. Quant. Biol. 43:709-719. 7. Brockman,W. W. 1978. Transformation ofBALB/c-3T3
cellsby tsAmutantsofsimian virus40- temperature sensitivity of the transformed phenotype and
retran-formationby wild-type virus. J.Virol. 25:860-870. 8. Brugge, J. S., and J. S.Butel.1975.Role of simianvirus
40geneAfunction inmaintenance of transformation.
J.Virol.15:619-635.
9. DiMayorca,G., J. Callender, G.Marin, and R. Gior-dano. 1969.Temperature-sensitivemutantsofpolyoma virus.Virology 38:126-133.
10. Eckhart, W.1969.Complementation and transformation by temperature-sensitive mutants of polyoma virus. Virology38:120-125.
11. Edwards, C. A. F., G. Khoury, and R. G. Martin.1979. Phosphorylation of T-antigen and control of T-antigen expression in cellstransformed bywild-type andtsA mutantsof simian virus40.J. Virol.29:753-762. 12.Fluck, M.M., and T. L Benjamin.1979.Comparisons
oftwoearlygenefunctions essential fortransformation inpolyomavirusandSV40. Virology 96:205-228. 13.Fried, M.1965.Celltransformation ability ofa
tempera-ture-sensitive mutant of polyoma virus. Proc. Natl. Acad.Sci. U.S.A.53:486-491.
14. Gaudray, P.,M.Rassoulzadegan, and F. Cuzin.1978.
Expressionof simian virus 40earlygenes intransformed ratcells is correlated with maintenance of the trans-formedphenotype. Proc. Natl. Acad. Sci. U.S.A. 75: 4987-4991.
15.Jeffreys,A.J., and R. A. Flavell.1977.Aphysicalmap of the DNAregionsflankingthe rabbit,B-globingene. Cell 12:429-439.
16.Ketner,G., and T. J.Kelly,Jr.1976.Integratedsimian virus 40 sequences intransformed cell DNA:analysis usingrestriction endonucleases. Proc. Natl. Acad. Sci. U.S.A. 73:1102-1106.
17.Kimura,G.,and A.Itagaki.1975.Initiation and main-tenance of celltransformationbysimian virus 40:.aviral genetic property. Proc. Natl.Acad.Sci. U.S.A. 72:673-677.
18.Maniatis,T., S. G. Kee, A. Efstratiadis, and F. C. Kafatos.1976.Amplificationand characterization ofa
,8-globingenesynthesizedinvitro. Cell8:163-182.
19.Martin,R.G., andJ. Y.Chou.1975.Simian virus40 functionsrequired for the establishment and mainte-nanceofmalignanttrnsformation.J. Virol. 15:599-612.
20.Martin, R.G.,V. P.Setlow, and C. A. F. Edwards. 1979.Rolesof simianvirus 40 tumor antigens in trans-formationof Chinesehamsterlungcells: studies with simianvinrs40double mutants. J.Virol.31:596-07. 21. Osborn, M.,and K.Weber. 1975. Simian virus 40 gene
Afunction and maintenance oftrnsformation.J.Virol. 15:636-644.
22. Rassoulzadegan,M., B.Perbal, and F.Cuzin.1978. Growthcontrol in simian virus 40-transformedratcells: temperature-independentexpression of the transformed phenotype intsAtransformants derived by agar selec-tion. J. Virol. 28:1-5.
23. Rassoulzadegan,M., R.Seif,and F.Cuzin.1978. Con-ditionsleadingtotheestablishment of the N (a gene-dependent) and A (agene-independent) transformed statesafterpolyoma virus infection of rat fibroblasts. J. Virol. 28:421-426.
24. Rosenberg, M., S.Segal,E.ILKuff, and M. F. Singer. 1977. The nucleotide sequence of repetitive monkey DNAfound in defectivesimian virus 40. Cell 11:845-857.
25. Rosenberg, H.,M. Singer, and M. Rosenberg. 1978. Highlyreiteratedsequencesof .sim
-iansimian.Science200:394-402.
26. Seif, R., and F. Cuzin. 1977. Temperature-sensitive
growthregulation in one type oftransformedratcells inducedby the tsa mutant of polyomavius.J.Virol. 24:721-728.
27. Seif,R.,and R.G.Martin. 1979. Growth state of thecell
early after infection with simian virus 40 determines whether the maintenance oftransformation willbe A-genedependentorindependent. J. Virol. 31:350-359.
28. Seif,R., and R. G.Martin. 1979.Simian virus 40small
t-antigen is notrequired for themaintenanceof trans-formation but may act as apromoter (cocarcinogen) duringestablishment of transformation inresting rat cells.J. Virol. 32:979-988.
29. Singer,D. S. 1979.Arrangement of a highly repeated DNAsequence in the genome and chromatin of the African greenmonkey. J. Biol. Chem. 264:5506-5514. 30. Southern, E. M. 1975. Detection of specific sequences
among DNAfragmentsseparated by gel electrophore-sis.J.Mol. Biol. 98:503-517.
31. Sugden,B., B. Detroy, R. J. Roberta,and J. Sam-brook. 1975. Agaroseslab-gel electrophoresis equip-ment.Anal.Biochem.68:36-46.
32. Tegtmeyer, P. 1975. Functions of simian virus 40 gene A intransforming infection. J. Virol.15:613-618. 33. Tenen, D. G., R. G.Martin,J.Anderson, and D. M.
Livingston. 1977.Biologicaland biochemical studies of cells transformed by simian virus40 temperature-sensitivegeneAmutants andAmutant revertants.J. Virol.22:210-218.
VOL. 35,1980