0022-538X/81/110440-10$02.00/0
Structures of the Oligosaccharides of the Glycoprotein Coded
by Early Region
E3
of Adenovirus
2
ROSALIND KORNFELD' ANDWILLIAM S. M. WOLD2*
Division ofHematologyandOncology, Washington University School of Medicine,' and Institute for
Molecular
Virology,
St.LouisUniversity
MedicalCenter,2
St.Louis,
Missouri63110Received27April 1981/Accepted23June1981
Early region E3 of adenovirus 2 encodes a glycoprotein, E3-gp25K, that is a
good model with whichto study structure-function relationships in
transmem-brane glycoproteins. We havedetermined thestructuresof theoligosaccharides linked to E3-gp25K. The oligosaccharideswere labeled with [2-3H]mannose in
adenovirus 2-early infected KBcellsfor 5.5 h (pulse)orfor 5.5 h followedbya
3-h c3-hase (pulse-chase). E3-gp25Kwasextracted andpurified bychromatography onDEAE-Sephacel in 7 Murea,followedby gel filtrationonacolumn of Bio-Gel
A-1.5m in 6 M guanidine hydrochloride. An analysis of the purified proteinby sodiumdodecylsulfate-polyacrylamidegelelectrophoresis indicated that itwas
>95%pure.Theoligosaccharides wereisolatedbypronasedigestion followedby
gel filtration on a column of Bio-Gel P-6, then by digestion with
endo-,8-N-acetylglucosaminidaseH, followed by gel filtrationonBio-GelP-6, and finallyby
paper chromatography. The pulse sample contained equal amounts of Man9GlcNAc and Man8GlcNAc and small amounts of Man7GlcNAc and
Man6GlcNAc. The pulse-chase sample had predominantly Man8GlcNAc and much less Man9GlcNAc, indicating that processing of the Man9GlcNAc to
Man8GlcNAc had occurred during the chase period. Thus, Man8GlcNAc is the major oligosaccharide onmatureE3-gp25K. Thestructuresofthese oligosaccha-rides were established by digestion with a-mannosidase, methylation analysis,
andacetolysis. The oligosaccharides found had typical high-mannosestructures
thathave been observed in other membrane and solubleglycoproteins, and the branching patternsandlinkages of the mannoseresidues of Man9GlcNAcwere
identical tothose of the lipid-linkedGlc3Man9GlcNAc2 donor. Thus, adenovirus 2 infection (early stages) apparently does not affect the usual cellular
high-mannose glycosylation pathways, and despite being virus coded, E3-gp25K is
glycosylated in thesame manner as atypical mammaliancell-codedglycoprotein.
Two glycoproteins, which we will designate E3-gp25K and E3-gp22K,aresynthesizedduring
earlystagesof infectionof human cellsby ade-novirus 2 (Ad2) (9, 10, 20). (gp25K and E3-gp22K have been referred to as E19K and
E17.5K by other workers [19-21, 24-26]. We have used the designations gp25K and gp22K because theyareglycoproteins thatmigrate in our sodium dodecyl sulfate [SDS]-gel system
withapparentmolecularweightsof25,000 [25K]
and22K,asjudgedbytherelative mobilities of
protein standards. Since glycoproteinsgenerally
migrate more slowly in SDS-gels than do
non-glycosylated proteins of comparable molecular weight, thetruemolecularweights of E3-gp25K
andE3-gp22Kwillbelessthan 25K and22K.) E3-gp25K and E3-gp22K have indistinguish-ablepeptide mapsand, thus, arehighly related
in their primary structures (19, 25; Q. Kapoor and W. Wold, unpublished data). Cells contain much more gp25K than gp22K.
E3-gp25K is localizedontheplasma membrane (4,
19, 20) and is complexed with the major
trans-plantation antigen of the host cell (15).
E3-gp25K has been purified to homogeneity, and antisera have been prepared (20; Kapoor and
Wold, unpublished data).
Four lines of evidenceprovethatE3-gp25Kis
coded by early region E3 (map position 76 to
86). First, it isimmunoprecipitated byantisera
against Ad2-transformed cell lines that retain E3(1, 10, 11, 23,27). Second,it isnotsynthesized incellsinfectedbyAd2+ND1, which hasa
dele-tionbetweenmappositions80.3 and 85.5(19, 24,
25).Third,it is translated in vitro from
r-strand-specificandE3-specificmRNA (18,19). Fourth,
440
on November 10, 2019 by guest
http://jvi.asm.org/
the partial N-terminal amino acidsequencehas been aligned with the DNAsequence in E3 (8, 19).
Aspects of the structure of gp25K can be
predicted from the DNAsequence (8, 18). The primarytranslation product is 159amino acids
long, and it containsaputative N-terminal
hy-drophobic signal sequence of about 17 or 18
amino acids (18). The C-terminus contains a
hydrophilic tail of15amino acids,which is
pre-ceded by a hydrophobic domain of 23 amino acids. Thereare twopotential sites for N-glyco-sylation, Asn-Val-Thr, located in the N-terminal portion of the molecule. By analogy with other membrane glycoproteins that have a similar
type of primary structure (21, 22), E3-gp25K
maybe oriented in themembrane such that the glycosylated N-terminal portion of the protein protrudesfrom thecell, the hydrophobic domain
traverses the membrane, and the hydrophilic tailextends into the
cytoplasm
and anchors the protein in the membrane (18).E3-gp25Kwas first showntobe glycosylated by
labeling
with[3H]glucosamine
(9, 10, 19,20) or[2-3H]mannose
and by binding toconcana-valin AorLens
culinaris
lectins (10, 20).Gly-cosylation
isinhibited by tunicamycin (19)orby2-deoxyglucose
orglucosamine
(25). When translated invitro, glycosylation
occursonly in thepresenceofamicrosomal fraction (18, 19).Since much is known about E3-gp25K and since it isaviralgeneproduct that issynthesized in fairly large amountsin infected cells, it isa
good
model with whichtostudy
structure-func-tion relationships in transmembrane glycopro-teins.Inthiscommunication,wereportthecom-pletestructuresof theoligosaccharides linkedto
E3-gp25K. The
oligosaccharides,
labeled with[2-3H]mannose,
were examined after a 5.5-hla-beling
period(pulse)
and also after a5.5-h la-belingperiod
followed by a 3-h chase (pulse-chase). At both times, all of the oligosaccha-rides were of the high-mannose type, withMan9GlcNAc2
andMansGlcNAc2
predominating
inthepulse sample and
ManmGlcNAc2
predom-inating in thepulse-chase sample.MATERIALS AND METHODS Virusinfection ofcellsandglycoprotein
label-ing. Methods used for thepurificationand
quantita-tionof Ad2 stocks have been describedpreviously (6,
28). Suspension cultures of KB cellswere grown at
37°C at aconcentration of3.5 X 105cells per ml in
Eagle minimal essential medium (MEM) containing
5% horseserum. Cells from2literswerecollectedby
centrifugation,suspendedin100ml of MEM without
horse serum, and infected with500PFU of Ad2(strain
38-2) per cell. After1h,cellsweredilutedto2liters
with MEMcontaining5%horseserum.At 2.5 h
post-infection, cells were collected by centrifugation and
werewashed in warm MEM containing l/lo the normal
glucose (i.e., 200 mg/liter) and 5% dialyzed horse
se-rum. Thewashing step wasrepeated, and the cells
weresuspended in1liter of warm MEM containing 'Ao
the normalglucose, 5%dialyzed horse serum, and20
Lg of1-/-D-arabinofuranosylcytosine per ml. The
1-fi-D-arabinofuranosylcytosine wasusedtoinhibit the
transition from earlyto late stages of infection, the
resultantshutoff of E3-gp25K synthesis (20), and the
possible modification of the host cell glycosylating
machinery. At 3.5 h postinfection, 10 mCi of
D-[2-3H]mannose (New England Nuclear Corp.; 14.5 Ci/
mmol) was added. Samplestaken immediately after
theaddition of the label and at 1-h intervals indicated
that incorporation of[2-3H]mannose into
trichloro-aceticacid-insoluble materialwaslinear overthe
la-belingperiod (datanotshown).
At 9 hpostinfection, the cellsweredivided intotwo
500-mllots,pulse andpulse-chase, andthe cellswere
collected by centrifugation. Thepulse-chase cellswere
washedonce in warm MEM containing the normal
amountofglucose, 5% horse serum, and20,ugof
1-ft-D-arabinofuranosylcytosine and thenwerefurther
in-cubated in500mlof thesamemedium. Thepulse cells
were washed twice at 4°C with cold
phosphate-buffered salinelackingCa2" and
Mg2"
and thenwerekeptonice. At12hpostinfection,thepulse-chase cells
werecollected andsimilarly washed with cold
phos-phate-buffered saline. Thus, both lots of cells were
labeled for5.5hwith[2-3H]mannose, but the
pulse-chasecellswerechasedfor3h withglucose.
Extraction andpurificationof
[2-3H]mannose-labeledE3-gp25K.Cellpelletsweresuspended in9
ml of isotonic high pHbuffer (0.14 M NaCl, 1 mM
MgCl2,10 mMTris-hydrochloride, pH8.5)containing
1mM phenylmethylsulfonyl fluoride (a protease
in-hibitor)at 1 to2°C andwerelysed by the addition of
Nonidet P-40to aconcentration of 0.5%. After5min
ofgentle mixingwith a pipette, the nucleiwere
re-movedby centrifugationat200xgfor10minat4°C.
The supernatant was removed, and the pellet was
washedoncewithlysis buffer. The supernatantswere
pooledandclarifiedbycentrifugationat12,000xgfor
10 min at4°C in aSorvall SS-34 rotorand then at
82,000xgfor1hat4°Cina50Tirotor.Triton X-100
was added to a final concentration of 1%, and the
samplesweredialyzedfor2daysat4°C againstDEAE
buffer (6 M urea, 10mM NaCl, 1%Triton X-100, 1
mMdithiothreitol, 1 mMphenylmethylsulfonyl
fluo-ride,20mMTris-hydrochloride, pH8.5). Theyieldat
this stagewas 1.38x 108and8.27x 107cpmfor the
pulseandpulse-chase samples, respectively.
TwoDEAE-Sephacel columns (1.1 by 7cm) were
poured,equilibrated againstDEAEbuffer, and loaded
with the protein samples at a rate of3 ml/h. The
columns were washed with DEAE buffer until the
counts being eluted were negligible and then were
eluted with DEAE buffercontaining1 MNaCl. The
twoflow-through poolsand thetwo1MNaCl eluate
poolsweredialyzedat4°Cfor2days against several
changesof50mMNH4HCO3-0.01%TritonX-100. An
analysisof thefractionsbypolyacrylamide gel
electro-phoresisinSDS(SDS-PAGE) indicated thatmostof
theE3-gp25Kwasin theflow-through fractions,but
on November 10, 2019 by guest
http://jvi.asm.org/
442 KORNFELD AND WOLD
that it was contaminated by a number of cellular
glycoproteins.Thepulseandpulse-chaseflow-through
fractions contained 2.2 x 107 and 1.2 x 107 cpm,
respectively.
E3-gp25Kwasfurtherpurifiedby gelifitration,as
follows. The dialyzed DEAE flow-through fractions
were lyophilized, suspended in about 5ml of0.4 M
glycine, mixed with 5volumes of cold acetone, and
precipitated for 2 h at 0 to 2°C. The proteins
co-precipitated with thelargeflocculentglycine
precipi-tate that formed in the presence of acetone. The
acetone extractsthe Triton X-100(atahigh
concen-trationafter thelyophilizationstep), which mustbe
removed because it forms micelles which preclude
partitioning of proteins in the gel filtration column.
Theglycine-proteinpelletsweresuspended in50mM
NH4HCO3-0.01%TritonX-100, lyophilized, and
resus-pended in0.7ml of 8 Mguanidinehydrochloride-2%
2-mercaptoethanol-10 mM EDTA-50 mM Tris-hy-drochloride, pH 8.5. The recovery of counts was >95%.
Thesamples wereheatedat 560Cfor45minto
dis-sociatethepolypeptidesandwereneutralized with0.1
ml of1 M sodium phosphate, pH 6.5. Thesamples
were mixed withasmall volume ofphenolred
con-taining100 mgofsucroseandwereappliedto acolumn
(1.5by100cm) of Bio-Gel A-1.5mequilibratedin6M
guanidinehydrochloride-10mMdithiothreitol-1 mM phenylmethylsulfonyl fluoride-20 mM sodium
phos-phatebuffer, pH6.5.One-hour(3.5-ml)fractionswere
collected. All fractionswereadjustedto0.02%Triton
X-100andwere dialyzed against 50mM
NH4HCO3-0.02% Triton X-100.Each fractionwascounted,and
selected fractionswereassayed bySDS-PAGE, using
the deoxycholate-trichloroacetic acid precipitation
method (seebelow).
SDS-PAGE. SDS-PAGEwascarriedoutwith
dis-continuous 13%polyacrylamidegelsasdescribed
pre-viously (27).ThtonX-100causes"smearing"ongels;
therefore, itwasextracted fromthe proteinsamples
beforeSDS-PAGE,asdescribedbelow.Thesamples,
adjusted to0.5 ml in 1.5-ml Eppendorf tubes, were
mixed(Vortex) with10
pl
of 2% sodiumdeoxycholateandincubated at roomtemperaturefor 15min.
Tri-chloroacetic acidwasaddedto afinal concentration of
10%, and thesampleswereincubatedat 0 to2°C for
30min.Theheavytrichloroacetic
acid-deoxycholate-proteinprecipitatethatformedwasremovedby
cen-trifugationfor4mininanEppendorfmicrofuge.The
pelletswerewashed twice with1 ml ofcoldacetone
andfinally were suspended in 20 to 50 ul of
SDS-PAGEsample buffer (27).
Preparation oflabeledglycopeptides. The re-tained fractions of the Bio-Gel A-1.5m column that
containedpurified E3-gp25K(seeFig. 1)werepooled
andlyophilized.Eachpoolwasdissolved in1.5mlof water and extracted twice with 2 ml of toluene to removeTritonX-100.The somewhat turbid solutions
wereincubatedat600Cwith7mgof pronase in2mM
CaCl2-0.1MTris-hydrochloride,pH8.0.After4h,an
additional7mgof pronasewasadded.After20h, each
reaction was boiled for 2 min and subjected to gel
filtrationon acolumn(1.5by98cm) of Bio-Gel P-6to
isolate theglycopeptides(seeFig.2below).
Preparation ofoligosaccharides.The
glycopep-tide fractions from each sample, obtained from the
Bio-Gel P-6 column, were lyophilized and redissolved
in 2.5ml of water. Onemilliliter of each glycopeptide
samplewastaken to dryness and incubated for 19 h at
370Cin 100
pl
of 50 mM citratephosphate buffer, pH6.5, containing2mUof
endo-,8-N-acetylglucosamini-dase H(endo H) (Miles Laboratories, Inc.). The
sam-ples, diluted to1.0 ml with water, were boiled for 3
minand againsubjected to gelfiltration on Bio-Gel
P-6as shown in Fig. 2.The pooledoligosaccharide
frac-tions from the Bio-Gel P-6 column were lyophilized,
redissolved in water, spotted onWhatman no. 1 paper,
andsubjected todescending paper chromatography in
pyridine-ethyl acetate-acetic acid-water (5:5:1:3),
sol-ventI, for7days to separateoligosaccharides
accord-ing to size. The sample lanes and a lane containaccord-ing oligosaccharide standards of known size were cut in
1-cmsegmentswhich wereeluted in 1.0 ml of water. A
sample of each was counted, and eluates were pooled that corresponded to Man9GlcNAc, Man8GlcNAc, Man7GlcNAc, and Man6GlcNAc (see Fig. 3).
Structural analysis ofoligosaccharides. The
oligosaccharides were methylated by the method of Hakomori (7), and the permethylated products were
hydrolyzed in 2 NH2S04 andanalyzed by thin-layer
chromatographyasreported by Li et al. (16). Labeled
oligosaccharides werereduced with NaBH4 and
sub-jectedto acetolysis aspreviously described (2). The
Man- Mandisaccharides obtained by acetolysis were
reduced withNaBH4andsubjected tomolybdate
elec-trophoresis as previously described (2) to separate
Man 2al,2 mannitol from Man 21,3 mannitol.
Oligosaccharidesweredigested withjack bean
a-man-nosidase preparedby the method of Li and Li (17) by
incubationat370Cwith 1U of enzyme in 20
pl
of 50mMacetatebuffer, pH 4.6, for 1.5h. The digestions
were then subjected todescending chromatography
on Whatman no. 1 paper in solvent I for 16 h to
separatefreemannosefrom Man-*GlcNAc
disaccha-ride.
RESULTS
Purity
of[2-3Hlmannose-labeled
E3-gp25K. Figure 1A shows the Bio-Gel A-1.5m column elution profile of the labeled
glycopro-teins from thepulse-labeledcells. Fractions20,33, 34, 35, 36,and37-41poolwere
analyzed by
SDS-PAGE. Themajority
ofE3-gp25K
wasfoundinthemajorretained
peak,
fractions33to-37 (Fig. 1B). Some E3-gp25K wasalso found
in the void volume, despite the use of 6 M
guanidine
hydrochloride
and10mMdithiothre-itol in the column
buffer,
which attests to thetendency of this
polypeptide
to aggregate. A smallquantity
ofE3-gp22K wasfound in frac-tions 35 to 41. The 44Kglycopolypeptide
infractions34 to 36ishighly relatedtoE3-gp25K,
because it has a very similar
[35S]methionine-containingpeptide
map (27;Q. Kapoor
and W.Wold,
unpublished
data).
Theapparent molec-ularweightestimate of44K forthis glycopoly-peptidedoes notruleoutthepossibility
that itis a dimer of gp25K, because the
mobility
ofJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
Vo
It
A
Cpu'
1a
X1D0
10
8
6
4
2
0 10 20 30 40 5
FRACTION NO I3.5ml/fraction)
B
fl %I 5 -. ! -W-
93K-69K- _
->l1.K _
...:5 _K-_40
13K-s ii_
41
FIG. 1. Gel filtration of[2-3Himannose
pulse-la-beledE3-gp25Kon acolumnofBio-Gel A-1.5m. The
column wasprepared and run as described in the
text. (A) Elutionprofile ofthe labeled
glycopolypep-tides. (B) SDS-protein gelwhich illustrates the
gly-copolypeptides infractions20, 33, 34, 35, 36, and
37-41pool. "M"refersto'4C-labeledproteinstandards
(NewEngland NuclearCorp.), whichwere
phospho-rylase b(93K), bovineserum albumin(69K),
ovalbu-min(46K), carbonicanhydrase (31K),andcytochrome
c(13K).
glycopolypeptidesinSDS-gelsis not an accurate
indication oftheirmolecular weights. The60K glycopolypeptide in fraction 35 may have been yet another form(e.g.,trimer)ofE3-gp25Korit may have been a contaminant. If itwas a
con-taminant,itsconcentrationwasmuchtoolowto
compromiseouranalysisof theoligosaccharides
ofE3-gp25K. Similarresults were obtained with
the glycoproteins from the
pulse-chase-labeled
sample (data notshown). Fractions 33to41 inFig. 1 were pooled,as were analogousfractions
from the pulse-chase-labeled sample, and the
glycopeptides were prepared bypronase
diges-tion.
Purification of the oligosaccharides of E3-gp25K. Figure 2 shows theBio-Gel P-6 col-umnprofileof theglycopeptides obtainedonthe pronasedigestofthepulse-labeled sample (solid line) andonthe oligosaccharides released from theseglycopeptides by endoHdigestion(dashed
line). Essentially thesameprofilewas obtained
for the pulse-chase sample. EndoHcleavesthe twoN-acetylglucosamine residues of the chito-biose core of high-mannose, asparagine-linked oligosaccharides, liberating an oligosaccharide withoneN-acetylglucosamine residueat its
re-ducing end and leavingoneN-acetylglucosamine
U residue attached to asparagine in the peptide. Kobata(12) hasrecently reviewedthespecificity studies thatshowthat endoH can act on
high-mannoseoligosaccharides, butnot onthe
com-plex type. In both the pulse and pulse-chase samples, all ofthe
[2-3H]mannose-labeled
oli-gosaccharidesweresusceptibletoendoH
cleav-ageand thusmustbe of thehigh-mannose-type
structure.
The oligosaccharide fractions from the
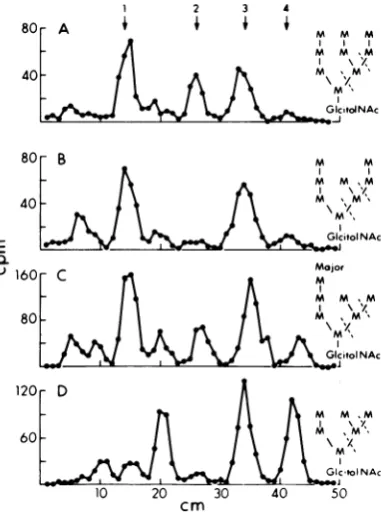
Bio-GelP-6columnsweresubjectedtopaper chro-matographytoresolve the different sized oligo-saccharides. Thepulse sample contained almost equal amounts ofoligosaccharides comigrating with authentic
MangGlcNAc
(ninemannoseres-idues linked to N-acetylglucosamine) and
Man8GlcNAc,aswell asmuchsmaller amounts
3,000
Vo Man
P'.
9
0
11 1.
I
0
II
(r
2,000k
E
1,000_
U 20 40 60 80
FRACTION
FIG. 2. Bio-Gel P-6gel filtration ofthe glycopep-tidesin the pronasedigest(0) andthe
oligosaccha-ridesreleasedfrom them after endo H digestion (0).
The Bio-Gel P-6 column (1.5 by98cm) was
equili-bratedandeluted with0.1Mammoniumbicarbonate,
and2-mlfractionswerecollected. Samples of every
fraction were counted in asolutioncontaining 4 ml
of 3a70 scintillationfluidand0.4mlofwater.
Frac-tions 32 to 50 werepooled fromtheglycopeptide peak,
andfractions49 to 60werepooled from the
oligosac-charidepeak.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.497.50.243.77.424.2] [image:4.497.256.446.421.564.2]444 KORNFELD AND WOLD
ofmaterialcomigrating with Man7GlcNAc and Man8GlcNAc (Fig. 3A). In contrast, the
pulse-chasesample(Fig.3B) containedpredominantly
MansGlcNAc,
with much less Man9GlcNAc.Upon rechromatography, each of the pooled
eluates from the first separation migrated as
single peaks coincident with Man96GlcNAc standards. The quantitative difference in
Man9GlcNAc
between thepulse and the pulse-chase samples indicated that processing of the oligosaccharidetoitsmatureform hadoccurred during thechaseperiod.Structural studies on each oligosaccharide species from both the pulse and pulse-chase sampleswereperformedasdescribedbelow, but dataareshownonly for thepulse samples.
Re-sults were essentially identical for the pulse-chasesample oligosaccharides.
Structure of the
Man9GlcNAc
oligosac-charide. When the oligosaccharide with the same mobility as Man9GlcNAc was digested with a-mannosidase, 89% of the radioactivitymigrated upon paper chromatography as free
mannoseand 11%migratedasthedisaccharide
Man
-.1,4
GlcNAc(data
notshown). Thus, itcontainedeighta-mannoseresidues andcanbe
represented as Man8 -a-, Man- - GlcNAc.
Todetermine whichhydroxylsonthemannose residues in
MangGlcNAc
were involved ingly-cosidic bonds, MangGlcNAc was subjected to methylation, and the methylated mannose
spe-cies released by acid hydrolysiswereseparated
by thin-layer chromatography. The resultsare
showninFig.4andaresummarized inTable 1.
MangGlcNAc
gaveriseto2,4-dimethyl, 3,4,6-tri-methyl, and 2,3,4,6-tetramethylmannosein theratio of 2:4:3.
To localize the Man
.1,6
Man linkages,MangGlcNAc
wasreduced with NaBH4andsub-jectedtoacetolysis (which preferentially cleaves
Man
--16,
Manlinkages).Fragmentsobtained12 3 4 5
MgM8 M7 M6 M5
400
A
200
E
D-)
B
10 20 30 40 50
cm
FIG. 3. Preparativepaperchromatography of oli-gosaccharides released by digestion with endo H.
Thechromatogramsweredevelopedandanalyzedas
described in the text. (A) Pulse sample; (B)
pulse-chasesample. Thearrowsshow themigration posi-tionsof authenticMan9GlcNAc-Man5GlcNAc
oligo-saccharides. Fractions corresponding toeachpeak
werepooledandanalyzed further.
r Man8
5 10 15
cm
FIG. 4. Thin-layer chromatography ofthe
methyl-atedmannosespeciesderivedfromthepooled
oligo-saccharidefractionsshown inFig.3. Thethin-layer platesweredevelopedandanalyzedasdescribedin
thetext.Thearrowsatthetopdenote themigration positions ofthefollowingauthenticmethylated man-nose species: (1) 2,3-dimethyl, (2) 2,4-dfmethyl, (3)
3,4,6-trimethyl; (4) 2,4,6-trimethyl, and(5) 2,3,4,6-tet-ramethylmannose.
E
a-u
200[
1600 1200
E
a- 800
-U
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.497.290.430.262.579.2] [image:5.497.66.243.307.590.2]VOL.40,1981OLIGOSACCHARIDES OF THE E3 GLYCOPROTEIN OF Ad2 445
TABLE 1. Methylation ofpulse-labeled oligosaccharidesandacetolysisfragments
No. ofmannoseresidues0a
Sample 2,3,4,6-Tetramethyl 2,4,6-Trimethyl 3,4,6-Trimethyl- 2,4-Dimethyl mannose mannose niannose mannose
Intactoligosaccharides
Man9GIcNAc 3.2 0 3.8 2.0
MansGlcNAc 3.2 0 2.9 1.9
Man7GlcNAc 2.8 0.4 2.1 1.7
Man6GlcNAc 2.8 0.2 1.2 1.8
Acetolysis fragmentsofMan9GIcNAc
Man--.Man --.Man 1.1 0.9 1.0 0
Man4GlcitolNAc 1.1 0.9 2.0 0
0Thesumof theradioactivityin
all
speciesofmethylatedmannosein eachsampleinFig.4wassetequaltothenumberofmannoseresidues in thatsampleand the counts per minute permannoseresiduewerecalculated
foreachsample.This valuewasthen usedtodeterminethe number ofmannoseresidues of everymethylated species.
had the same mobilities as authentic
Man4GlcitolNAc, Man3,
and Man2(Fig.
5A).
[image:6.497.254.445.262.519.2]These results
suggested
thestructure shown inFig. 5A,
where thedashed lines indicatecleavage
sites at
al,6
linkages.
Thetwolarger fragments
werecharacterized further
by methylation
(Ta-ble
1). Man2
was shown to be linkedal,2
by
subjecting
the materialtoNaBH4 reductionfol-lowed
by
high-voltage
electrophoresis
inmolyb-date
buffer,
whichseparates
Man 2,man-nitol from Man al3 mannitol
(data
notshown).
The results establish that
Man9GIcNAc
con-tains two
branching
mannose residues substi-tutedat0-3and0-6,
giving
riseto2,4-dimnethyl
mannose aftermethylation,
which must be linked to one another via anal,6
linkage
asshown in
Fig.
5Atoproduce
thefragments
ob-served uponacetolysis.
TheMan3fragment
con-tainsoneterminalmannoseresidue
(giving
riseto
2,3,4,6-tetramethyl mannose),
one residue substituted at0-2(giving
riseto3,4,6-trimethyl
mannose),
and one residue substituted at 0-3(giving
rise to2,4,6-trimethyl
mannose) whichmust have
originally
been one of the branchpoint
mannose residues. The otheroriginal
branch
point
mannoseis in theMan4GlcitolNAc
fragment,
where it issubstituted at 0-3(giving
riseto
2,4,6-trimethyl mannose).
Alsopresent
in thisfragment
are oneterminalmannoseresidue and two mannose residues substituted at 0-2.These results
provide
evidence that theMan9GIcNAc
has thestructureshown inFig.
6.Structure of the Man5GleNAc
oligosac-charide. When theMan8GIcNAc
oligosaccha-ride was
digested
witha-mannosidase,
88% of theradioactivity
was releasedasfree mannose,and 12% had the
chromatographic mobility
ofMan
-~---P,
aGIcNAc,
indicating
the presence of1201
60~
1 2 3 4
E
0. 1u
M M
GlcitodNAc
160-C MMajor
80-M ,m
GlcitoINAc
D
,M\ XY
~ ~ ~ m
Gc.tolNAc
FIG. 5. Paper chromatography of the fr-agments producedby acetolysis oftheoligosaccharides. The structures attherightindicate thestructuresdeduced
fr-om
the data, with the dashed lines showing thelocation of the al,6 linkages that are
preferen-tiaily
cleaved by acetolysis. (A) Man9G1cNAc, (B) Man8GlcNAc, (C) Man7GlcNAc, and (D)Manr4-GleNAc. Thearrowsatthetop denote themigration
positionsofthefollowingstandardoligosaccharides:
(1)Man4GlcitolNAc,(2) Man3, (3) Man2,and(4) man-n-ose.
sevena-mannose residues. As indicated in
Fig.
4 and Table 1,
methylation
of theoligosaccha-ride gave rise to
2,4-dimethyl,
3,4,6-trimethyl
and
2,3,4,6-tetramethyl
mannosein the ratio of40,
1981on November 10, 2019 by guest
http://jvi.asm.org/
446 KORNFELD AND WOLD
Man Man Man
al a2al al,2
Man Man Man
al,21 xl
,3\\
/l,6Man Man
al,3 X\..4
//al
,6Man
Ia
G1lcNAc
MangGlcNAc
Man Man
al,2 Ial,2
Man Man Man
al,21
al,3\\
Sal,6Man Man
al,3
\*.
//al
,6 ManGIcNAc
Man8GlcNAc
Man al,21
Man Man Man
al,2| al,3\ /al,6
Man Man
al,3 /al ,6
Man
GlcNAc
A (major)
Man Man
al 21 |al,2
Man Man
al 2 1xl 3
Man Man
al,3\ /l,6
Man G1cAc
B
Man
|al,2
Man Man Man
al,2 al,3\ / l,6
Man Man
al ,3 'Nl\ /cal,6 Man
Ia
G1lcNAc
(mi nor) C
Man7GlcNAc
Man Man Man
al,21 al,3
\
/cl ,6Man Man
al,3 \ /l611 Man
GlcNAc
Man
al
.21
Man Man
al,21
cl,3\
Man Man
al,3 \ /al,6
Man
GlcNAc
(minor) (major)
Man6GlcNAc
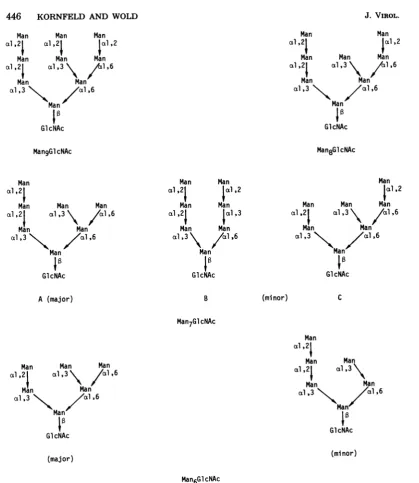
FIG. 6. Structuresproposedfor theoligosaccharides of E3-gp25K.
2:3:3. Acetolysis of Man8GlcNAc produced two
major fragments corresponding to Man4-GlcitolNAc and Man2(Fig.5B).Molybdate
elec-trophoresis (data not shown) resolved the
re-duced Man2 into two peaks, one comigrating
with Man
012
mannitol and the othercomi-gratingwith Man
al,3,
mannitol in the ratio of1.5:1.0.Thestructureexpectedtoyield these fragments isshown in Fig. 5B and Fig. 6. TheexcessMan
al2
mannitolobtainedprobably
reflectssomeunderdegradationintheacetolysis reaction, in which only a single cleavageoc-curred, giving rise to Man
a1,2,
Man andMan6GlcitolNAc.
The latter wouldaccountfortheslowest-moving peakonthechromatogram inFig.5B. Themethylationandacetolysisdata
indicate that
MansGlcNAc
has the structureproposedinFig.6.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.497.52.472.49.533.2]Structure of the
Man7GlcNAc
oligosac-charide. WhenMan7GlcNAc wastreated witha-mannosidase,
85% of theradioactivity
wasre-leasedasfreemannoseand 15%comigrated with Man
1,4,
GlcNAcdisaccharide,
indicating thepresence of six a-mannose residues. The
methylation products
shown inFig.
4and Table1 and the
acetolysis
cleavage
pattern shown in Fig.5C both indicate thepresenceofmorethanoneisomeric
Man7GlcNAc
structure inthisoli-gosaccharide.
Acetolysis
produced major
frag-ments
corresponding
toMan4GlcitolNAc, Man2,
and free mannose; this is consistent with thestructureand
cleavage
sites shown inFig.
5C for the majorMan7GlcNAc species. However,
sig-nificantamountsoffragments corresponding
toMan3GlcitolNAc
andMan3werealsoproduced, andmolybdateelectrophoresis
revealed that the Man2 contained both Man al,3- Man(ex-pected
from themajor
species)
and Manal,2 Man.Toaccountfor theMan3
fragment
andfor theproduction
of2,4,6-trimethyl
man-nose and less thantwo full residues of
2,4-di-methylmannosefromMan7GlcNAc,
weproposethataminor
species
with the Bstructure occursalong with the major A structure
(Fig. 6).
Toaccountfor the Man3GlcitolNAc
fragment
andthe Mn al,2 Man
fragment,
we alsoproposethat another minor
species
with theCstructureshownin
Fig.
6alsooccursinMan7GlcNAc.
Structure of theMan^GlcNAc
oligosac-charide.
When theMan6GlcNAc
was treated witha-mannosidase,
84% of theradioactivity
wasreleasedasfreemannoseand16%migrated with Man
1,4-
GlcNAc. This indicates fivea-mannose residues.
Methylation
of Man6-GlcNAc gave the results shown inFig.
4 and Table 1,andacetolysis
produced the fragmen-tationpattemshown inFig.
5D. Thefragments
corresponded
toMan3GlcitolNAc, Man2,
and free mannose and fit the structure with thecleavage
sites shown inFig.
5D.Allof theMan2wasshown tobe Man
a1,3,
Manby molybdateelectrophoresis;
thus, the structure of the vastmajority of the Man6GlcNAc
oligosaccharide
mustbe as shown inFig.6forthemajor species.
This oligosaccharide should give rise to 2,4-di-methyl, 3,4,6-tri2,4-di-methyl, and2,3,4,6-tetramethyl
mannoseintheratio 2:1:3 andproduceno
2,4,6-trimethyl
mannose.Infact,
themethylation
re-sults
(Table
1) indicate thepresenceof0.2 resi-dueof2,4,6-trimethylmannose, 0.2 residue more3,4,6-trimethyl
mannosethanexpected,and 0.2 residueless2,4-dimethyl
andtetramethyl man-nose than expected; this suggests that 20%of
Man6GlcNAc
has thestructure shown in Fig. 6 asthe minor species. This structurewouldpro-duce a Man4GlcitolNAc fragment upon
aceto-lysis, and inspection of Fig. 5D shows a small
radioactive peak at that position.
DISCUSSION
Glycosylation of proteins in mammalian cells
occursbyenbloctransfer ofa
Glc3Man9GlcNAc2
moiety fromalipid-linked
oligosaccharide donor(16). Typically, the varioustypes of
high-man-noseand complexoligosaccharides foundon gly-coproteins are generated by processing of
Glc3Man9GlcNAc2
asthenewly synthesized gly-coproteins travel from the rough endoplasmic reticulum through the Golgi region, withre-moval of the threeglucoses
occurnng
veryrap-idly(14). In ouranalyses of theoligosaccharides released by endo H, we found that the pulse-labeled sample, which should contain newly
synthesized
andmatureE3-gp25K,gaverise toequal
amounts ofMan9GlcNAc
and Man8-GlcNAc and much smaller amounts ofMan7-GlcNAc
and Man6GlcNAc. In contrast, thepulse-chase-labeled sample,
which shouldcon-tain only mature
E3-gp25K,
gave rise tomainly
MansGlcNAc,
withconsiderably
lessMansGlcNAc,
Man7GlcNAc,
andMan6GlcNAc. This implies thatMan9GlcNAc
was processed mainly into Man8GlcNAc during the 3-h chaseperiod.
The branching pattern and linkage of the mannose residues that we found in theMan9GlcNAc oligosaccharide
from E3-gp25Kare identical to those in the lipid-linked
Glc3Man9GlcNAc2 previously
described (16), suggesting that E3-gp25K is initially glycosyl-atedby thisprecursoroligosaccharide and then threeglucose
residuesaretrimmedoff.In the case of
glycoproteins
havinghigh-mannose
oligosaccharides
smaller thanMani-GlcNAc2,
there isgenerally
somedegree
ofhet-erogeneity
in the number ofmannose residues introducedduring
theprocessing reactionscat-alyzed by
theGolgi
al,2-mannosidase (26). Thistypeof sizeheterogeneity has been described for theheavy-chain
oligosaccharides
ofhumanim-munoglobulin
Mmyeloma
proteins (2, 3, 5), andinthe presentstudy, this mechanism probably
accountsfor the small amounts ofMan7GlcNAc andMan6GlcNAc
species
observed. In additiontosize
heterogeneity,
high-mannose oligosaccha-ridescandisplay structural heterogeneityamongoligosaccharides
of the same size. Although threepossible
Man8GlcNAc oligosaccharide
iso-merscanbeformedby
thecleavage
of asingle terminal mannose residue fromMan9GlcNAc,
we found only one isomer, indicating thatthe
removalof the first mannose residue from the
on November 10, 2019 by guest
http://jvi.asm.org/
448 KORNFELD AND WOLD
E3-gp25K oligosaccharide is specific and not
random. A similar finding has been made for several otherglycoproteins (3, 5).However, since
several Man7GlcNAc isomers and two
Man6GlcNAc isomers were
found,
thesubse-quent mannose
processing
was either lessor-dered, ordifferent patterns of ordered removal occurredatseparate
glycosylation
sitesto gen-eratethesesmaller,quantitatively
minorspecies. Theoligosaccharides ofE3-gp25Karealltypical high-mannose oligosaccharides having struc-tures identical to those found in a variety of other membrane and solubleglycoproteins
(13). Other viralglycoproteins,
for example, the G protein of vesicular stomatitis virus (14), areinitially
glycosylated by
the host cell with ahigh-mannose
oligosaccharide,
which is then processed and convertedto acomplex-type
oli-gosaccharideby
the addition ofouter chainN-acetylglucosamine,
galactose,
and sialic acidres-idues and by the addition offucosetothe core
N-acetylglucosamine.
Inthe case ofE3-gp25K,
wefoundnoevidence that suchcomplex chains
were formed, as shown by the fact that the oligosaccharideswere
quantitatively
cleavedby endo H and could be degradedcompletely
toMan g GlcNAc
by
treatmentwitha-manno-sidase. The factorsthat determine whether the oligosaccharideat agiven
glycosylation
site on aparticular protein will remainahigh-mannosetype orbeconvertedtoacomplextypeby
gly-cosyl
transferases of theGolgi
apparatus are notknown, but probably the
conformation
ofthepolypeptide isonefactor.
The DNA sequence of the putativegene for E3-gp25K reveals two potential sites for
Asn-linkedglycosylation (8, 19). Although thereare nodirectdata, circumstantial evidencesuggests
that both sitesmaybe
glycosylated.
Forexam-ple,Persson etal. (20)found4mol of N-acetyl-glucosaminepermol ofE3-gp25K, which is
con-sistent with2mol of
ManxGlcNAc2.
Further, theapparent molecular weight of nonglycosylated gp25K, obtained
by
infecting
cells in the pres-ence oftunicamycin (19, 20) or by cleaving with endo H (Q. S. Kapoor, W. S. M.Wold, and G.Chinnadurai, Virology, inpress), isabout 4.5K to 6K smaller than glycosylated gp25K as
estimated by SDS-PAGE. Only 1 mol of
Man8GlcNAc2,
with amolecularweight of 1.7K, isunlikely
to cause such a large difference. If there are, in fact, 2 mol of the sameMansGlcNAc2
moiety per molofgp25K and ifthey are linked to the two Asn-Val-Thr sites
predictedby theDNAsequence, it is ofinterest
that there are no obvious similarities in the amino acidsproximaltothesesites(8, 19).Thus,
the signals that dictate the oligosaccharide
structure at a given glycosylation site are not
obvious from a casualinspection of the amino
acidsequenceof theglycosylation sites; perhaps
these signals are degenerate, or perhaps they
involve protein folding.
ACKNOWLEDGMENTS
Wethank Maurice Green for support and for reading the manuscript and Catherine Devine and Karen Fujimoto for excellenttechnical assistance.
Thiswork was supported by Public Health Service research grantsCA-24710 and CA-08759 and Cancer Center Core grant CA-25843from the National Institutes of Health. W.S.M.W. wassupported by Public Health Service Research Career Development award CA-00650 from the National Institutes of Health.
LITERATURE CITED
1. Brackmann,K.H.,M.Green,W.S. M.Wold,and M.
Cartas. 1980. Identification andpeptide mapping of adenovirus 2-induced earlypolypeptides isolated by two dimensionalgel electrophoresis. J. Biol. Chem. 255: 6772-6779.
2. Chapman, A., and R. Kornfeld. 1979.Structure of the high mannoseoligosaccharides of a human IgM mye-lomaprotein. I. The major oligosaccharides of the two high mannoseglycopeptides. J. Biol. Chem. 254:816-823.
3. Chapman,A., and R.Kornfeld.1979.Structure of the high mannose oligosaccharides of a human IgM mye-loma protein. II. The minor oligosaccharides of high mannoseglycopeptideI. J.Biol. Chem.254:824-828. 4. Chin,W.W.,and J. V.Maizel,Jr.1976.Polypeptides
ofadenovirus.VII. Further studies of early polypeptides in vivo andlocalization of E2 and E2Atothe cellplasma
membrane. Virology 71:518-530.
5. Cohen,R. E., and C. E. Ballou. 1980. Linkage and sequence analysis of mannose-rich glycoprotein core oligosaccharidesbyprotonnuclear magnetic resonance spectroscopy.Biochemistry 19:4345-4358.
6. Green, M., and W.S. M.Wold. 1979.Preparation of humanadenoviruses.MethodsEnzymol. 58:425-435. 7. Hakomori,S.1974.Arapidpermethylationofglycolipid
andpolysaccharide catalyzed by methylsulfinyl carb-anion indimethyl sulfoxide. J. Biochem. (Toyko) 55: 205-208.
8. Herisse,J., G.Courtois,and F. Galibert. 1980. Nu-cleotide sequence of the EcoRI Dfragment of adeno-virus 2 genome. Nucleic Acids Res. 8:2173-2192. 9. Ishibashi, M.,and J. V.Maizel,Jr.1974.The
polypep-tides of adenoviruses. VI.Early and late
glycopolypep-tides.Virology 58:345-361.
10.Jeng, Y-H., W. S. M. Wold, and M. Green. 1978. Evidence for an adenovirustype 2-coded early
glyco-protein. J.Virol. 28:314-323.
11.Johansson, K., H. Persson, A.M.Lewis,U. Petters-son, C.Tibbetts,andL.Philipson.1978.Viral DNA sequences and geneproducts in hamstercells trans-formedby adenovirus type2.J.Virol.27:628-639. 12.Kobata, A.1979. Use of endo-andexoglycosidasesfor
structural studies ofglycoconjugates. Anal. Biochem. 100:1-14.
13. Kornfeld,R., and S. Kornfeld.1980.Structureof
gly-coproteins and theiroligosaccharide units,p.1-34.In W.J. Lennarz(ed.), Thebiochemistryofglycoproteins
and proteoglycans. Plenum Publishing Corp., New York.
14. Kornfeld, S.,E.Li, and I. Tabas.1978.Thesynthesis of complex-typeoligosaccharides. II. Characterization of theprocessing intermediates in the synthesis of the complex oligosaccharideunits of the vesicular
stomati-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
tis virus G protein. J. Biol.Chem.253:7771-7778. 15. Kvist,S., LOstberg,H.Persson, L.Philipson,and
P. A. Peterson. 1978.Molecular association between transplantation antigens and cell surface antigen in adenovirus-transformed cell line. Proc. Natl. Acad. Sci. U.S.A. 75:5674-5678.
16.Li, E.,L. Tabas, and S. Kornfeld. 1978. The synthesis of complex-type oligosaccharides. I. Structure of the lipid-linked oligosaccharide precursor of the complex-type oligosaccharides of thevesicular stomatitis virus G pro-tein. J. Biol.Chem.253:7762-7770.
17. Li, Y. T., and S. C. Li. 1972. a-Mannosidase,fl-N-acetyl
hexosaminidase, and B-galactosidase from Jack Bean Meal.Methods Enzymol. 28:702-713.
18.Persson,H., M. Jansson, and L.Philipson.1980. Syn-thesis and genomic site for an adenovirus type 2 early
glycoprotein.J.Mol. Biol. 136:375-394.
19. Persson,H., H. Jornvall, and J.Zabielski 1980. Mul-tiple mRNA species for the precursor to an adenovirus-encoded glycoprotein: identification and structure of thesignal sequence. Proc. Natl. Acad. Sci. U.S.A. 77: 6349-6353.
20. Persson,H.,C.Signas,andL.Philipson. 1979. Purifi-cation and characterization of an early glycoprotein fromadenovirus type 2-infected cells. J. Virol.
29:938-948.
21. Rogers, J., P.Early, C. Carter, K. Calamine, M. Bond, LHood,and R. Wall. 1980. Two mRNAs with differ-ent 3'ends encodemembrane-bound and secreted forms
ofimmunoglubulin g chain. Cell 20:303-312. 22. Rose,J. K., W. J. Welch, B. M. Sefton, F. S. Esch,
andN.C. Ling. 1980. Vesicular stomatitis virus gly-coprotein isanchored in the viral membrane by a hy-drophobic domain near the COOH terminus. Proc. Natl. Acad. Sci. U.S.A. 77:3884-3888.
23.Ross, S. R., S. J. Flint, and A. J. Levine. 1980. Identi-fication of the adenovirus early proteins and their ge-nomic mappositions. Virology 100:419-432.
24. Ross, S., and A. J. Levine. 1979. The genomic map positionof the adenovirus type 2glycoprotein.Virology 99:427-430.
25. Storch, T. G., and J. V. Maizel, Jr. 1980. Theearly
proteins of the nondefectiveAd2-SV40 hybrid viruses: the 19Kglycoprotein is coded by Ad2 earlyregionE3. Virology 103:54-67.
26. Tabas, I., andS. Kornfeld. 1979. Purification and char-acterization of a rat liverGolgi a-mannosidasecapable
of processing asparagine-linked oligosaccharides. J. Biol.Chem. 254:11655-11663.
27. Wold, W. S. M., and M. Green. 1979. Adenovirus type 2early polypeptidesimmunoprecipitated byantisera to fivelines ofadenovirus-transformedratcells. J.Virol. 30:297-310.
28. Wold,W. S.M.,M.Green,and J. K. Mackey.1978. Methods and rationale foranalysis of human tumors for nucleic acid sequences ofoncogenichuman DNA vi-ruses, p. 69-161. In E. Busch(ed.),Methods in cancer research, vol. 15. AcademicPress,Inc., New York.