Sarcoma-Associated Herpesvirus (KSHV) ORF57 Protein Is Not
Required for Efficient KSHV Lytic Replication
Da-Jiang Li,aDinesh Verma,aand Sankar Swaminathana,b
Division of Infectious Diseases, Department of Medicine,a
and Department of Pathology,b
University of Utah School of Medicine, Salt Lake City, Utah, USA
Kaposi’s sarcoma-associated herpesvirus (KSHV) ORF57 protein is expressed early during lytic KSHV replication, enhances ex-pression of many KSHV genes, and is essential for virus production. ORF57 is a member of a family of proteins conserved among all human and many animal herpesviruses that are multifunctional regulators of gene expression and act posttranscriptionally to increase accumulation of their target mRNAs. The mechanism of ORF57 action is complex and may involve effects on mRNA transcription, stability, and export. ORF57 directly binds to REF/Aly, a cellular RNA-binding protein component of the TREX complex that mediates RNA transcription and export. We analyzed the effects of an ORF57 mutation known to abrogate REF/Aly binding and demonstrate that the REF-binding mutant is impaired in activation of viral mRNAs and noncoding RNAs confined to the nucleus. Although the inability to bind REF leads to decreased ORF57 activity in enhancing gene expression, there is no demonstrable effect on nuclear export of viral mRNA or the ability of ORF57 to support KSHV replication and virus production. These data indicate that REF/Aly-ORF57 interaction is not essential for KSHV lytic replication but may contribute to target RNA stability independent of effects on RNA export, suggesting a novel role for REF/Aly in viral RNA metabolism.
K
aposi’s sarcoma-associated herpesvirus (KSHV), a human gammaherpesvirus, is a causative agent of Kaposi’s sarcoma (KS), primary effusion lymphoma (PEL), and a subset of Multi-centric Castleman’s disease (MCD), (for a review, see reference 10). The ORF57 protein is expressed early in the lytic phase of KSHV replication during KSHV reactivation from latency (54) and is essential for lytic virus replication (15,28). KSHV recom-binants carrying targeted deletions of ORF57 fail to replicateinvitroand are markedly deficient in expression of several early and
late lytic cycle KSHV genes (15,28). Replication of recombinants with ORF57 deleted can be specifically rescued by transfection of ORF57 (15,28). Although ORF57 is functionally similar to ho-mologs Epstein-Barr virus (EBV) SM and herpes simplex virus (HSV) ICP27, each protein cannot cross-complement the others in rescuing virus replication (3,13,15).
Although deficient lytic gene expression may explain the re-quirement for ORF57 during replication, the mechanisms by which ORF57 enhances gene expression remain incompletely characterized and are likely to be complex. Two major mecha-nisms have been proposed: that ORF57 enhances nuclear export of lytic KSHV mRNAs or, alternatively, that ORF57 increases total lytic mRNA levels by enhancing stability and/or transcription, without affecting their nuclear export. Despite possible transcrip-tional effects (32,39), it has been clearly established that ORF57 induces accumulation of reporter gene mRNA by posttranscrip-tional mechanisms. ORF57 increases reporter gene expression in a promoter-independent fashion and does not increase transcrip-tion from promoters where transcriptranscrip-tional initiatranscrip-tion rate has been directly examined (2,14,24).
ORF57 increases the levels of its target mRNAs in both the nucleus and the cytoplasm, suggesting that at least part of its ac-tivity is nuclear and due to effects on RNA stability (46). Consis-tent with this view, ORF57 increases the accumulation of KSHV PAN RNA, a noncoding RNA that is confined to the nucleus (14, 24,51). A recent report also demonstrated that ORF57 stabilizes
PAN RNAs, thereby increasing their half-lives and intranuclear accumulation (46). An alternative, but not mutually exclusive, hypothesis is that ORF57 facilitates nucleocytoplasmic RNA ex-port by recruiting various cellular proteins to KSHV mRNAs (4, 17,29). According to this model, KSHV lytic mRNAs, the majority of which are intronless, are inefficiently exported due to an inabil-ity to assemble ribonucleoprotein transcription/export (TREX) complexes whose formation is associated with the process of pre-mRNA splicing (4). ORF57 binds the cellular RNA-binding pro-tein, REF/Aly, which is a component of TREX, via an RNA-inde-pendent protein interaction (33,36). A site-specific mutant of ORF57 carrying two amino acid changes in the REF/Aly binding domain (PP208,211AA) was shown to be incapable of binding REF/Aly (4). This mutant ORF57, referred to as ORF57Pmut, un-like wild-type ORF57 (wtORF57), did not bind TREX complexes
in vitro. It was also reported that this mutation specifically resulted
in inefficient export and decreased cytoplasmic accumulation of KSHV ORF47 mRNA in cotransfection assays. In contrast to these findings, REF/Aly knockdown was reported to have a negligible effect on ORF57 enhancement of gene expression in KSHV-in-fected cells (31). Furthermore, in a separate report, ORF57 mu-tants unable to bind REF/Aly nevertheless retained partial ability to enhance KSHV target mRNA accumulation (36).
These findings left unanswered the question of whether REF/ Aly binding is required for ORF57’s essential role in KSHV repli-cation and virion production. We therefore directly examined the
Received11 May 2012Accepted25 June 2012
Published ahead of print3 July 2012
Address correspondence to Sankar Swaminathan, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01190-12
on November 7, 2019 by guest
http://jvi.asm.org/
ability of the ORF57 PP208,211AA mutant, which is defective for REF/Aly binding, to rescue replication of a KSHV strain with ORF57 deleted. Importantly, ORF57Pmut, which is unable to bind REF/Aly, was nevertheless capable of rescuing replication of an ORF57-knockout KSHV. Although the ORF57Pmut was less active in increasing target mRNA levels, in contrast to the previous report that impairment of REF/Aly binding inhibits nuclear ex-port of KSHV mRNAs, mRNA exex-port itself was not affected. Fur-ther, the reduced function of the ORF57Pmut was particularly apparent in its effect on PAN RNAs, which are exclusively nuclear. These data indicate that the primary effect of the REF/Aly-ORF57 interaction may be to enhance nuclear accumulation and stability of RNAs, which is consistent with a recent report that REF/Aly may decrease the turnover of nuclear RNA (50).
MATERIALS AND METHODS
Cells, plasmids, and DNA transfection.HeLa and 293 cells were grown at 37°C in Dulbecco modified Eagle medium (Invitrogen) supplemented with 10% fetal calf serum and glutamine. 293T cells are human embryonic kidney cells transformed with adenovirus type 5 DNA (12). The Vero cell line was originally initiated from the kidney of a normal adult African green monkey (48). Generation of Vero cells infected with an ORF57-knockout KSHV was previously described (15). The Bac36 ORF57-null mutant cell line (Bac36-⌬57) was kindly provided by Zhi-Ming Zheng, which contains a full-length KSHV genome with ORF57 deleted and a hygromycin B resistance gene and expresses green fluorescent protein as a marker (28). Cells were maintained in Dulbecco modified Eagle medium and 10% fetal calf serum. All transfections were performed with Lipo-fectamine Plus (Invitrogen) according to the manufacturer’s instructions. Transfections were performed with 1g of DNA per 10 cm2of adherent
cells in either six-well dishes or 25-cm2flasks. When multiple plasmids
were used in transfections, empty pcDNA3 vector (Invitrogen) was used to equalize the amount of DNA in each transfection mix to a total of 1g of DNA per 10 cm2of cells.
The ORF47, ORF56, and ORF59 coding regions were PCR-amplified from BCBL1 cells (41) and cloned in cytomegalovirus (CMV) promoter driven expression vector pcDNA3. The HA-tagged ORF57 cDNA has been previously described (14). An ORF57 mutant in which prolines 208 and 211 were replaced by alanine (MUT ORF57) gene was generated by PCR and cloned in pCDNA3 with an N-terminal influenza virus hemag-glutinin (HA) epitope tag. The ORF50 expression plasmid DD267, in which ORF50 is cloned in pCDNA3, has been previously described (18). The FLAG-tagged REF/Aly expression vector was kindly provided by Gideon Dreyfuss (21). The sequence of each plasmid was verified by DNA sequencing. Primer sequences are available on request.
Immunofluorescence microscopy.HeLa cells for immunofluores-cence confocal microscopy were grown on glass coverslips. Cells were plated at 2⫻105cells per well in six-well dishes and transfected with 1g
of DNA per 10 cm2of surface area 48 h prior to fixation. Cells were washed
with phosphate-buffered saline (PBS), fixed in 4% paraformaldehyde at 20°C for 15 min, washed again, and stored at 4°C. Fixed cells were blocked with 20% goat serum and incubated with primary antibodies. The follow-ing primary antibodies were used in combination as appropriate: mono-clonal anti-HA or polymono-clonal rabbit anti-HA for wtORF57 and HA-ORF57Pmut and polyclonal anti-REF for endogenous cellular REF/Aly. The cells were then washed and incubated with both fluorescein isothio-cyanate-conjugated anti-mouse IgG and Alexa Fluor 594-conjugated goat anti-rabbit IgG. Nuclei were counterstained with DAPI (4=,6= -diamidino-2-phenylindole) and slides were mounted for microscopy with ProLong Gold antifade reagent (Molecular Probes). Slides were visualized and im-aged using an Olympus Fluoview FV1000 confocal microscope.
RNA isolation and analysis.Total cellular RNA was isolated from washed cell pellets using Qiazol and Qiagen miRNeasy columns according to the manufacturer’s protocols. Cytoplasmic and nuclear RNA fractions
were prepared by lysing cells in hypotonic lysis buffer. Briefly, after cells were detached using 0.48 mM EDTA in PBS at room temperature, washed cell pellets were incubated for 10 min at 4°C in hypotonic buffer (1.5 mM MgCl2, 10 mM KCl, 0.5 M dithiothreitol [DTT], 10 mM HEPES [pH 7.9])
with 80 U of RNasin (Promega)/ml and homogenized with 16 to 18 strokes of a microfuge tube as described previously (23). Dounced lysate was centrifuged for 5 min at 400⫻gto pellet nuclei. Qiazol was added to the supernatant at a ratio of 7.5:1 and placed at 4°C. Nuclear pellets were washed twice in low-salt buffer (1.5 mM MgCl2, 0.02 mM KCl, 0.2 mM
EDTA, 25% glycerol, 0.5 M DTT 20 mM HEPES [pH 7.9]) with 80 U of RNasin/ml to remove cytoplasmic contamination. Nuclei were lysed by addition of 250l of Qiazol and sonication on ice with 10 pulses at a 30% duty cycle of a Branson sonicator, followed by the addition of 500l of Qiazol. Both supernatant and nuclear fractions were then further pro-cessed according to the manufacturer’s protocol and eluted in RNase-free Tris-EDTA. RNA was quantitated by spectrophotometry. Northern blot-ting was performed as previously described after electrophoresis in 1% agarose (45). Blots were hybridized with32P-labeled, gene-specific DNA
probes generated by random oligonucleotide primed Klenow DNA poly-merase (Ready-To-Go DNA labeling beads kit; Amersham). Blots were visualized by using autoradiography and a Typhoon phosphorimager (GE Life Sciences), and quantitation was performed with ImageQuant soft-ware and normalized to U6 RNA and GAPDH (glyceraldehyde-3-phos-phate dehydrogenase) levels.
Immunoprecipitation and immunoblotting.Immunoprecipitations were performed with lysates of COS7 or HeLa cells harvested 48 h after transfection. The cells were washed and lysed in 250 mM NaCl, 50 mM Tris-HCl (pH 7.2), 0.5% NP-40, and protease inhibitor cocktail (Sigma) by incubation at 4°C for 15 min with vortexing. Clarified lysates were precleared with normal rabbit IgG (Bethyl) and protein A-agarose beads (Sigma), followed by immunoprecipitation with either anti-HA mono-clonal antibody (Covance), anti-REF polymono-clonal antibody (Bethyl), or normal rabbit IgG. RNase treatment was performed with 100g of RNase A/ml for 30 min at 37°C. Immunoprecipitations were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and im-munoblotting with anti-FLAG or anti-HA antibody. Imim-munoblotting was performed with polyclonal anti-ORF57 antibody (36), monoclonal anti-HA antibody, or anti-FLAG antibody and horseradish peroxidase-conjugated secondary antibody, followed by chemiluminescence detec-tion (Amersham).
qPCR. Quantitative reverse transcription-PCR (qPCR) was per-formed with SYBR green One-Step RT-PCR Master Supermix (Applied Biosystems) according to the manufacturer’s protocol using a StepOne-Plus real-time PCR system (Applied Biosystems). No template controls and no reverse transcription controls were included in each analysis. Each sample was analyzed in triplicate with gene specific primers and GAPDH was used as the endogenous control. Unless specified otherwise, RNAs from at least two independent transfections were analyzed simultane-ously. The gene-specific primers were as follows: ORF47 Q1F, 5=-AGCC
TCTACCCTGCCGTTGTTCT-3=; ORF47 Q1R, 5=-ACGACCGCGACTA
AAAATGACCT-3=; ORF59Q1F, 5=-CTCCCTCGGCAGACACAGAT-3=;
ORF59Q1R, 5=-GCGTGGTGCACACCGACGCCC-3=; PAN-Q1F, 5=-CC
GCCGATTGTGGGTTGATT-3=; PAN-Q1R, 5=-TTTTGTTCTGCGGGC
TTATGGAG-3=; GAPDH Q1F, 5=-AGGGTCATCATCTCTGCCCCCT
C-3=; and GAPDH Q1R, 5=-TGTGGTCATGAGTCCTTCCACGAT-3=. Induction of lytic gene expression and virus replication.To induce KSHV lytic gene expression or virus production in 293 or Vero cells in-fected with Bac36 ORF57-null mutant KSHV, cells were either transin-fected with ORF50 expression vector or infected with adenovirus expressing ORF50 (AdRTA; kindly provided by Don Ganem) (1). As appropriate, ORF57 plasmid or empty vector and ORF56 plasmid was included in the transfection. Where indicated, 12-O-tetradecanoylphorbol-13-acetate (TPA) was added to the growth medium at a concentration of 20 ng/ml the day after transfection to enhance lytic replication. For virus passage, supernatant was harvested from the cultures 5 days after transfection, KSHV ORF57-REF/Aly Binding Dispensable for Replication
on November 7, 2019 by guest
http://jvi.asm.org/
cleared by centrifugation twice, and filtered through a 0.45-m-pore-size cellulose acetate filter. Uninfected 293T cells were infected by centrifuga-tion and incubacentrifuga-tion in virus-containing supernatant for a minimum of 1.5 h. Cells were examined daily and photographed at 48 h postinfection under fluorescence microscopy to detect green fluorescent protein (GFP) expression in newly infected cells. All passage experiments were per-formed from a minimum of three independent transfections.
RESULTS
Characterization of mutant ORF57 expression, intracellular
lo-calization, and interaction with REF/Aly.In order to fully
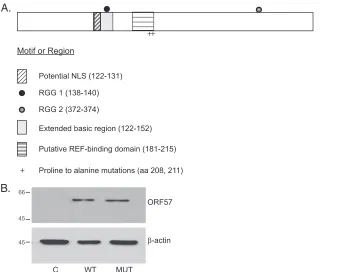
char-acterize the effects of mutating ORF57 amino acids previously reported to affect REF/Aly binding, we introduced two proline-to-alanine substitution mutations at amino acids 208 and 211 in the ORF57 cDNA by site-directed PCR-mutagenesis (Fig. 1). The mutant ORF57 gene (henceforth referred to as ORF57Pmut) was sequenced to verify the mutations and was cloned in the expression vector pCDNA3 with an HA epitope tag at the amino terminus. In order to verify that the mutant protein would be expressed at levels comparable to wild-type ORF57 protein, ORF57Pmut and wtORF57 were transfected into HeLa cells, and expression was compared by immunoblotting. As shown inFig. 1B, ORF57Pmut was expressed at levels compa-rable to those for wtORF57. Similar results were obtained with either polyclonal anti-ORF57 rabbit antiserum or anti-HA monoclonal antibody and in 293 cells (results not shown).
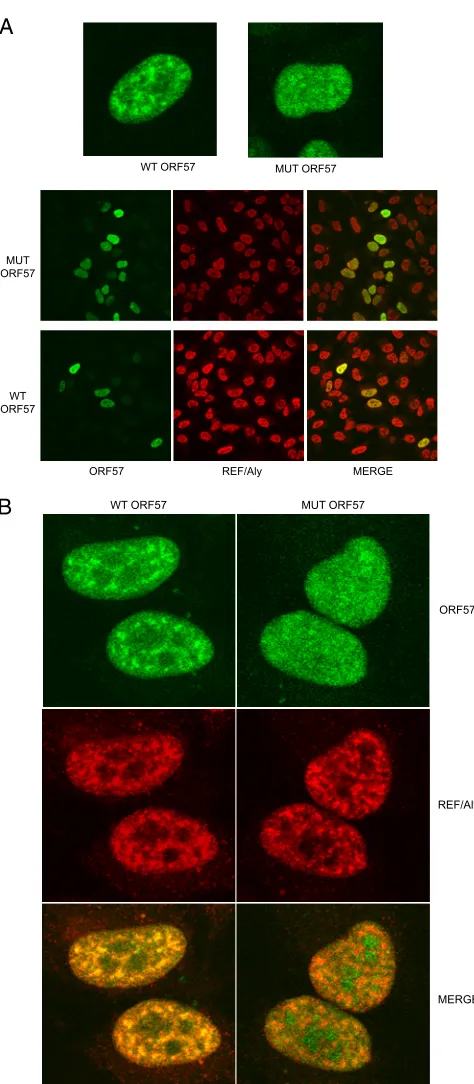
The intracellular localization of mutant and wtORF57 was next compared by immunofluorescence confocal microscopy. HeLa cells were transfected with HA-ORF57Pmut or HA-wtORF57
plasmids, fixed, and stained with anti-HA antibody. As shown in Fig. 2A, wtORF57 displayed a diffuse, speckled distribution in the nucleus, with greater localization in heterogeneous nuclear foci. This pattern of localization is typical of ORF57 and is exclusively nuclear, as we have previously described (36). The distribution of ORF57Pmut, was also nuclear, but in a more homogeneous, gran-ular pattern (Fig. 2A, upper panel). Cells transfected with HA-wtORF57 or HA-ORF57Pmut were also stained with anti-HA monoclonal antibody and polyclonal anti-REF/Aly to determine whether REF/Aly and ORF57 proteins colocalize. As shown inFig. 2A, lower panel andFig. 2B(higher magnification), while both wtORF57 and REF/Aly are distributed throughout the nucleus, the pattern of localization is different. In cells expressing wtORF57, REF/Aly is concentrated in foci with ORF57. However, in cells expressing ORF57Pmut, REF/Aly did not demonstrate such colocalization and ORF57Pmut maintains a more diffuse localization.
In order to confirm that the mutant ORF57 gene we had gen-erated was deficient in REF/Aly interaction, coimmunoprecipita-tion was performed. Cells were cotransfected either HA-wtORF57 or HA-ORF57Pmut. Immunoprecipitations were performed with anti-REF or anti-ORF57 antibodies and immunoblotting was used to detect both proteins. As shown inFig. 3, wtORF57 was efficiently coimmunoprecipitated with REF/Aly, but ORF57Pmut was not.
Enhancement of RNA accumulation by mutant and wtORF57.
ORF57 strongly enhances the steady-state accumulation of several KSHV mRNAs both in transfection assays and in KSHV-infected
FIG 1Construction and expression of mutant ORF57 defective for REF/Aly binding. (A) Diagram of ORF57 protein motifs and sites of REF/Aly-binding mutation. The nuclear localization signal (NLS), two RGG motifs, and other potential functional domains are shown. Amino acids 208 and 211 in the REF/Aly-binding domain that were mutated from proline to alanine are shown. (B) Expression of wild-type ORF57 and mutant ORF57 proteins. Lysates of cells transfected with wtORF57 or ORF57Pmut were analyzed by SDS-PAGE and immunoblotting with anti-ORF57 antibody. The blot was stripped and reprobed with anti-actin antibody (lower panel). The molecular mass in kilodaltons is indicated at the left.
on November 7, 2019 by guest
http://jvi.asm.org/
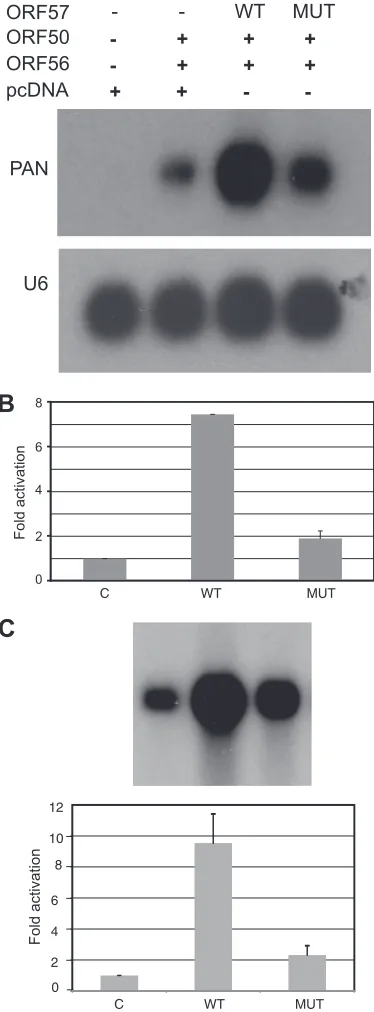
[image:3.585.135.472.64.336.2]cells undergoing lytic replication (2,14,15,24,28). If REF/Aly interaction is important for ORF57 effects on its target mRNAs by facilitating export, the PP208,211AA mutant ORF57 should be significantly impaired in function since it is deficient in the ability to bind REF/Aly as shown above (4). We therefore compared ORF57Pmut and wtORF57 in transfection assays using KSHV ORF59 as a reporter mRNA. As shown inFig. 4A, ORF59 is ex-tremely dependent on ORF57 for efficient expression. ORF59 mRNA was virtually undetectable by Northern blotting in the ab-sence of ORF57 but was readily detected when wtORF57 was cotransfected. ORF57Pmut also increased total ORF59 mRNA, but to a lesser degree than wtORF57. Quantitation of the ORF59 levels using a phosphorimager indicated that ORF59 accumula-tion was enhanced 18-fold by wtORF57 and 12-fold by ORF57Pmut over the levels in the absence of ORF57 (Fig. 4B). In order to confirm the Northern blotting data, qPCR was also per-formed on the RNAs that were used for Northern blotting. As shown inFig. 4C, the results were similar, with wtORF57 enhanc-ing ORF59 accumulation more than ORF57Pmut (⬃20-fold ver-sus 13-fold over control). Thus, loss of REF/Aly binding leads to
⬃35% reduction in ORF57’s ability to enhance accumulation of ORF59 mRNA (P⬍0.01).
[image:4.585.44.281.59.601.2]The enhanced accumulation of mRNA in the presence of ORF57 has been attributed to effects of ORF57 on export via REF/ Aly. Alternatively, we and others have shown that ORF57 in-creases nuclear mRNA accumulation without significant changes in cytoplasmic/nuclear (C:N) ratio of the mRNA, suggesting that ORF57 exerts its effects primarily by increasing RNA stability, consequently increasing both nuclear and cytoplasmic accumula-tion (36,46). We therefore wanted to determine whether REF/Aly binding was required for ORF57 to enhance RNA stability, inde-pendent of any effects on RNA export. Since the polyadenylated nontranslated KSHV transcript PAN is exclusively nuclear, and ORF57 increases PAN RNA stability (46,51,52), we examined the effect of the REF/Aly binding mutation on the ability of ORF57 to enhance PAN RNA accumulation. Vero cells infected with a re-combinant KSHV deleted for ORF57 were transfected with either wtORF57 or ORF57Pmut and with ORF50 to induce KSHV lytic gene expression. RNA was harvested from transfected cells, and PAN expression was measured by Northern blotting. As shown in Fig. 5A, ORF57Pmut was less active than wtORF57 in enhancing PAN RNA levels. By phosphorimager analysis, and normalizing levels to U6 RNA, a primarily nuclear cellular RNA, ORF57Pmut had 38% of the activity of wtORF57 in enhancing PAN accumu-lation (Fig. 5B). PAN RNA levels in these samples was also mea-sured by qPCR, and the results confirmed the pattern observed on the Northern blots, with ORF57Pmut having⬃25% of the activity of wtORF57. To further confirm the decreased activity of
FIG 2Nuclear localization patterns of wtORF57, ORF57Pmut, and REF/ Aly. (A) HeLa cells transfected with either wtORF57 or HA-ORF57Pmut were fixed and stained with monoclonal anti-HA and poly-clonal anti-REF antibodies and examined by confocal immunofluo-rescence microscopy. Individual cells stained with anti-HA to detect ORF57 (upper panel) and lower power view of cells stained with each antibody and merged (lower panel). (B) Higher magnification of cells transfected and stained in panel A above. Cells were transfected with either HA-wtORF57 or HA-ORF57Pmut and stained with anti-HA and anti-REF/ Aly antibodies and visualized as described above.
FIG 3Coimmunoprecipitation of ORF57 and REF/Aly. 293T cells were trans-fected with either HA-wtORF57 (WT) or HA-ORF57Pmut (MUT) or empty vector (C). Lysates of transfected cells were immunoprecipitated with anti-REF/Aly antibody (lanes Ref), anti-ORF57 (lanes 57), or normal rabbit serum (lanes C) and immunoblotted with anti-HA antibody to detect ORF57. A total of 1% of each input lysate (lanes I) was loaded onto each gel.
KSHV ORF57-REF/Aly Binding Dispensable for Replication
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.311.530.67.114.2]ORF57Pmut on PAN accumulation compared to wtORF57, a PAN expression plasmid was transfected into HeLa cells with ei-ther wtORF57, ORF57Pmut, or empty vector. RNA from these cells was analyzed by Northern blotting and qPCR. The results were consistent with the observed effects of wild-type and mutant ORF57 on PAN when it was expressed in KSHV-infected cells, with ORF57Pmut being 25% as active as wtORF57. Thus, a signif-icant reduction in activity of ORF57Pmut on a nonexported RNA (PAN) is observed, and this reduction is more pronounced than that which occurs when the target is an mRNA that undergoes nuclear export. These data therefore indicate that REF/Aly re-cruitment may have effects on RNA stability that are independent of its role in nuclear RNA export. These data also demonstrate that the effect is promoter-independent, as we have previously
dem-onstrated for wtORF57 (14) and therefore unlikely to be due to any effects on transcription.
The effect of ORF57 on export of KSHV target mRNAs remains to be fully characterized. Although an effect on export has been
FIG 4Effect of mutant or wtORF57 on ORF59 expression. (A) RNA was isolated from HeLa cells transfected in duplicate with ORF59 and either wtORF57 (WT), ORF57Pmut (MUT), or empty vector (C), and Northern blotting was performed with ORF59 probe. The blot was stripped and re-probed with U6 probe as an internal loading control. (B) Quantitation of samples in panel A was performed by phosphorimager. (C) RNA from cells transfected with ORF59 and either empty vector (C), wtORF57 (WT), or ORF57Pmut (MUT) as in panel A was analyzed by qPCR to quantitate ORF59 mRNA.
FIG 5Effect of wtORF57 or ORF57Pmut on nuclear PAN RNA expression. (A) Northern blotting was performed to measure the levels of PAN RNA in Vero cells infected with ORF57-knockout KSHV. HeLa cells were transfected with ORF50 and either wtORF57 (WT), ORF57Pmut (MUT), or empty vector (C). (B) qPCR was performed on the RNAs from panel A. (C) RNA from HeLa cells transfected with PAN and either ORF57 or ORF57Pmut plasmids was analyzed by Northern blotting (upper panel) and qPCR (lower panel). qPCR was performed on RNA from three transfections.
on November 7, 2019 by guest
http://jvi.asm.org/
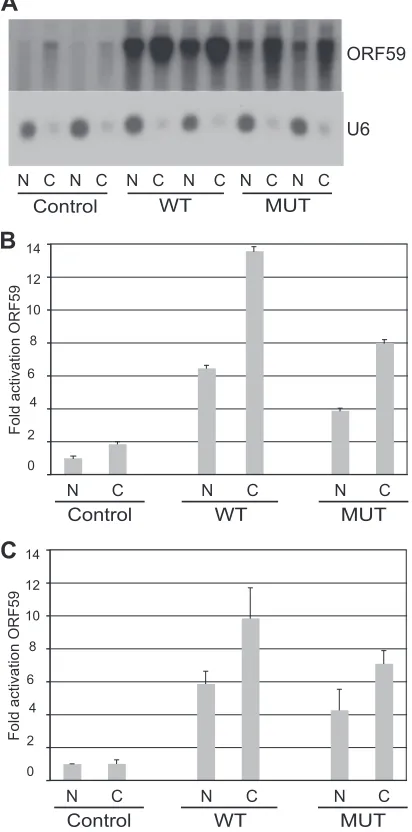
[image:5.585.68.260.73.463.2] [image:5.585.328.515.78.584.2]suggested based on reports of increased cytoplasmic accumula-tion of some KSHV mRNAs in the presence of ORF57, such find-ings have not been universally replicated (4,6,36). In order to compare the cytoplasmic accumulation of representative KSHV mRNAs in the presence of ORF57Pmut and wtORF57, we sepa-rated and measured cytoplasmic and nuclear RNA from cells ex-pressing specific KSHV target mRNAs and either wtORF57 or ORF57Pmut. Northern blotting of the samples demonstrated that wtORF57 significantly increased accumulation of both nuclear and cytoplasmic ORF59 mRNA (6- and 13-fold, respectively). Al-though ORF57Pmut was less active in enhancing ORF59 accumu-lation, it nevertheless enhanced ORF59 accumulation in both the nucleus and the cytoplasm at levels 50% those of wtORF57 (Fig. 6AandB). Importantly, the C:N ratio of ORF59 target mRNA was similar, regardless of whether ORF57Pmut or wtORF57 was ex-pressed (approximately 2: 1). In order to obtain further quantita-tive data and confirm the Northern blotting results, we performed qPCR analysis of the RNAs with primers specific for ORF59. As shown inFig. 6C, mutant and wild-type ORF57 both enhanced nu-clear and cytoplasmic ORF59 RNA expression, with ORF57Pmut be-ing less active. The C:N ratio of ORF59 mRNA was 1.67 with both ORF57Pmut and wtORF57 (P⫽0.37, indicating that the ratios are unlikely to be significantly different), confirming that wtORF57 is not superior in facilitating relative cytoplasmic accumulation of ORF59 mRNA.
The effect of ORF57 may be target RNA specific, and it was therefore possible that effects on export might be detectable only with some mRNAs. ORF57 has previously been reported to increase cytoplasmic to nuclear KSHV ORF47 mRNA ratios, suggesting an export effect (4). We therefore examined the effect of both wtORF57 and ORF57Pmut on ORF47 expres-sion, in experiments similar to those performed with ORF59 as a target mRNA. Northern blotting to detect ORF47 mRNA demonstrated that both wtORF57 and ORF57Pmut increased accumulation of ORF47 mRNA, with ORF57Pmut exerting 60 to 70% of the activity of wtORF57. (Fig. 7A andB). Precise quantitation by qPCR of the nuclear and cytoplasmic levels was performed with normalization to cellular RNA, confirming the Northern analysis (Fig. 7C). Importantly, the N:C ratios of ORF47 RNA were essentially identical to those seen with ORF57Pmut and wtORF57 (1.6: 1,P⫽0.94), indicating that although the overall activity of ORF57Pmut was decreased, this effect was unrelated to changes in export.
Rescue of ORF57-knockout KSHV by ORF57Pmut.An
unre-solved question regarding the ORF57-REF/Aly interaction is whether this interaction is required for ORF57’s role in KSHV lytic replication and virion production. Although ORF57 ho-mologs in EBV and HSV also interact with cellular export pro-teins, whether these interactions are essential for replication re-mains unclear. We and others have shown that ORF57 enhances expression of several KSHV lytic genes and enables efficient virion production in vitro by complementation of ORF57-deficient recombinant KSHV (15, 28). The partially retained ability of ORF57Pmut to increase KSHV target gene RNA expression sug-gested that it might be sufficient to enable KSHV replication and virion production. We therefore tested the ability of ORF57Pmut to rescue replication of ORF57-defective recombinant KSHV. KSHV virion production and replication of ORF57-knockout KSHV can be rescued by transfection with ORF57, followed by induction of the lytic cycle by chemical treatment or ORF50
trans-gene expression (15,28). Further, Majerciak et al. have demon-strated that disruption of ORF57 leads to impaired ORF56 expres-sion incis, and cotransfection of ORF56 more efficiently rescues virion production (28). In order to compare the ability of wtORF57 and ORF57Pmut to rescue the⌬57 mutant, 293 cells infected with Bac36⌬57 were transfected with ORF56 and either empty vector, wtORF57 or ORF57Pmut. Twelve hours after trans-fection, KSHV replication was induced by transduction of ORF50 using replication-incompetent adenovirus (AdRTA). Cell super-natant was harvested after 96 h, filtered, and used to infect unin-fected 293 cells. Cells that are inunin-fected by KSHV passaged in this manner express GFP from the Bac36⌬57 genome and are readily visualized (15,28). Live infected cells were examined by
fluores-FIG 6Effect of wild-type or mutant ORF57 on nuclear and cytoplasmic ac-cumulation of ORF59 mRNA. (A) Northern blotting was performed with ORF59 probe on RNA isolated from cytoplasmic and nuclear fractions of lysates from HeLa cells transfected with ORF59 and either empty vector (Con-trol), wtORF57 (WT), or ORF57Pmut (MUT). Blot was stripped and reprobed with U6 as a control (lower panel). (B) Quantitation of samples in panel A was performed by phosphorimaging. (C) qPCR of nuclear and cytoplasmic RNAs isolated as in panel A above.
KSHV ORF57-REF/Aly Binding Dispensable for Replication
on November 7, 2019 by guest
http://jvi.asm.org/
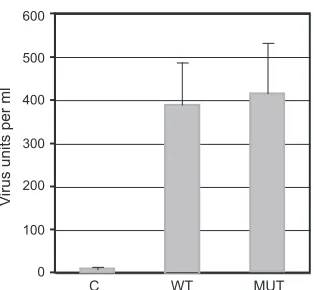
[image:6.585.319.525.71.487.2]cence microscopy 48 h after infection and infected cells that ex-pressed GFP were counted. As shown inFig. 8, wtORF57 and ORF57Pmut were both able to significantly rescue KSHV virion production, whereas ORF56 and ORF50 alone were not. The mean infectious virus units/ml of supernatant were 477 per ml and 445 per ml from wtORF57- and ORF57Pmut-transfected cells, respectively, compared to⬍10 U/ml from empty vector-trans-fected cells. Thus, the ability of ORF57 to bind REF/Aly did not appear to required for the essential function of ORF57 in enhanc-ing virion productionin vitro.
DISCUSSION
KSHV ORF57 has been shown to play roles in RNA transcription, processing, export, stability, and translation (2,4,5,15,24,28–32, 36,39,46). Whether one or more of these activities is predomi-nant in specific situations has not been established. The inter-relatedness of these processes also makes it difficult to determine whether ORF57 has multiple independent functions.
The majority of lytic cycle KSHV genes are encoded as single
open reading frames, a property shared with other herpesviruses (35,42,44). The fact that splicing of cellular intron-containing RNAs is required for efficient formation of export complexes has given rise to the hypothesis that intronless herpesvirus mRNAs may be intrinsically incapable of being efficiently exported. Be-cause association of REF/Aly and other export adapters with mRNA is thought to be dependent on splicing for intron-contain-ing genes (9,27,53), the finding that ORF57 interacts with the cellular REF/Aly protein has led to the model that ORF57 is in-volved in export of viral mRNAs to the cytoplasm (4). Although REF/Aly is not essential for cellular mRNA export, it plays an important role in TAP recruitment and RNA export (11,22,40, 49). ORF57 binds REF/Aly directly, and possibly CBP80, leading to the model that ORF57 recruits REF/Aly to its target mRNAs and facilitates TAP recruitment and formation of functional export complexes. In support of this model, a REF/Aly nonbinding ORF57 mutant was reported to be deficient in enhancing cyto-plasmic accumulation of ORF47 mRNA (4). However, depletion of REF with small interfering RNAs had minimal effects on ORF57’s ability to increase KSHV gene expression (31). Recently, ORF57 was shown to interact with UIF, another cellular export factor that can recruit TAP to mRNA in the absence of REF/Aly (16,17). Knockdown of both UIF and REF/Aly was shown to have an additive effect, decreasing ORF57’s ability to activate KSHV gene expression, similar to the effect on cellular gene expression (16). The interaction of ORF57 with REF/Aly may therefore play a redundant role in mRNA export which can be replaced by UIF. Alternatively, these interactions may be important for ORF57’s other functions such as RNA stabilization.
The experiments described here allow several conclusions. First, the REF/Aly interaction contributes significantly to ORF57’s ability to increase RNA levels. In order to address the role of the REF/Aly interaction in ORF57 function, we utilized several tran-scripts that are responsive to ORF57. Mutation of ORF57 so that it no longer binds REF/Aly led to impaired nuclear accumulation of ORF59, ORF47 and PAN. Although the activating effect of
FIG 7Effect of ORF57 on nuclear and cytoplasmic accumulation of ORF47 mRNA. (A) Northern blotting was performed with ORF47 probe on RNA isolated from cytoplasmic and nuclear fractions of lysates from HeLa cells transfected with ORF47 and either empty vector (Control), wtORF57 (WT), or ORF57Pmut (MUT). (B) Quantitation of samples in panel A was performed by phosphorimaging. (C) qPCR of nuclear and cytoplasmic RNAs isolated as in panel A above.
FIG 8Rescue of⌬57-KSHV replication by transfection of ORF57. 293 cells stably carrying⌬57 KSHV bacmid were transfected with either empty vector (C), wtORF57 (WT), or ORF57Pmut (MUT). At 24 h posttransfection, cells were infected with Ad50 and treated with TPA to induce KSHV replication. After an additional 96 h, supernatants were collected, filtered and used to infect 293T cells. Cells were examined 24 to 48 h after infection by fluorescence microscopy, and GFP-positive cells were counted. In all experiments, when cells transfected with empty vector (C), fewer than 10 GFP units (infected cells)/ml of supernatant were obtained. The results are representative of four independent transfection and passage experiments.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.344.501.63.208.2] [image:7.585.69.256.67.430.2]ORF57Pmut was compromised by the mutation, the C:N ratios of KSHV mRNAs were not significantly altered. Similar findings have been reported in herpesvirus saimiri, where the ORF57 ho-molog markedly increases both nuclear and cytoplasmic accumu-lation of ORF47 mRNA but does not alter C:N ratios (47). These findings differ significantly from a previous study in which ORF47 export was stimulated by wtORF57 but not by an identical REF/ Aly interaction mutant ORF57 (4). The ORF57 effects on nuclear mRNA accumulation described in our study are most likely due to effects on stability rather than increased transcription, especially when ORF59 and ORF47 were transcribed from a CMV promoter, which we and others have shown is not affected by ORF57 (14,24, 46). Therefore, these data suggest that the REF-ORF57 interaction may play a role in stabilizing RNAs.
Consistent with the model that REF/Aly recruitment may play a novel role in RNA stabilization in addition to export, ORF57Pmut was also significantly deficient in its ability to in-crease PAN RNA levels, which is exclusively nuclear, and not ex-ported. These data are consistent with the recent report by Stubbs et al. that tethering of REF/Aly to KSHV RNAs increases their nuclear stability (50). We and others have also demonstrated that a larger deletion mutant of ORF57, which also cannot bind REF, is similarly impaired in its effects on PAN (36,50). Interestingly, polyadenylation of PAN may be enhanced by REF (50). Whether REF plays a similar role in polyadenylation of viral or cellular mRNAs remains to be determined.
Although KSHV ORF57, EBV SM, and HSV ICP27 are re-quired for viral replication and interact with various cellular ex-port factors, it remains unclear whether these interactions are es-sential for viral replication. We have directly examined the effect of the REF-binding mutation on the ability of ORF57 to support KSHV replication and demonstrate that, surprisingly, the inability to bind REF/Aly does not significantly affect rescue of an ORF57-knockout virus. The most likely interpretation is that while an inability to bind REF/Aly impairs KSHV gene expression, the de-crease is not limiting for virus productionin vitro. Our findings are similar to those made in the study of ORF57 homolog CMV UL69, which interacts with UAP56 and the related URH49, but UAP56/URH49 binding was not required for replication (25). A caveat regarding the limitations of such rescue experiments is in order. Although rescue of a lethal mutation by transfection of the deleted gene is adduced as evidence of its essential nature, rescue by a mutant gene does not prove that the mutation does not im-pair function. For example, the inability to bind REF/Aly by ORF57 may slow the kinetics of virus production without affect-ing the ultimate yield of virus measured at the endpoint of the assay.
Several lines of evidence now suggest that in addition to roles in export, TREX proteins may play a role in preventing the rapid nuclear degradation that is observed with cDNA-derived tran-scripts and artificial noncoding RNAs (20, 34, 37, 38, 43,53). Intronless cellular mRNAs contain sequences within their coding regions that increase nuclear stability and bind TREX complex proteins independent of splicing (26). Although REF/Aly binding by HSV ICP27 was initially thought to be important for viral mRNA export, it has been shown to be dispensable (7, 8,19). Thus, while interaction of ORF57 homologs with the cellular ex-port machinery may also be imex-portant for viral mRNA exex-port, our findings support the model that recruitment of these factors may
serve to perform other functions such as protection of viral mRNA.
ACKNOWLEDGMENTS
This study was supported by grants RO1 CA119905 and CA81133 to S.S. from the National Cancer Institute, National Institutes of Health.
We thank Zhi-Ming Zheng, Don Ganem, Gideon Dreyfuss, and Rolf Renne for their generosity with reagents, cell lines, and advice.
REFERENCES
1.Bechtel JT, Liang Y, Hvidding J, Ganem D.2003. Host range of Kaposi’s sarcoma-associated herpesvirus in cultured cells. J. Virol.77:6474 – 6481. 2.Bello LJ, et al.1999. The human herpesvirus-8 ORF 57 gene and its
properties. J. Gen. Virol.80:3207–3215.
3.Boyer JL, Swaminathan S, Silverstein SJ.2002. The Epstein-Barr virus SM protein is functionally similar to ICP27 from herpes simplex virus in viral infections. J. Virol.76:9420 –9433.
4.Boyne JR, Colgan KJ, Whitehouse A.2008. Recruitment of the complete hTREX complex is required for Kaposi’s sarcoma-associated herpesvirus intronless mRNA nuclear export and virus replication. PLoS Pathog.
4:e1000194. doi:10.1371/journal.ppat.1000194.
5.Boyne JR, Jackson BR, Taylor A, Macnab SA, Whitehouse A. 2010. Kaposi’s sarcoma-associated herpesvirus ORF57 protein interacts with PYM to enhance translation of viral intronless mRNAs. EMBO J.29:1851– 1864.
6.Boyne JR, Whitehouse A.2009. Nucleolar disruption impairs Kaposi’s sarcoma-associated herpesvirus ORF57-mediated nuclear export of in-tronless viral mRNAs. FEBS Lett.583:3549 –3556.
7.Chen IH, Li L, Silva L, Sandri-Goldin RM.2005. ICP27 recruits Aly/REF but not TAP/NXF1 to herpes simplex virus type 1 transcription sites al-though TAP/NXF1 is required for ICP27 export. J. Virol.79:3949 –3961. 8.Chen IH, Sciabica KS, Sandri-Goldin RM.2002. ICP27 interacts with the
RNA export factor Aly/REF to direct herpes simplex virus type 1 intronless mRNAs to the TAP export pathway. J. Virol.76:12877–12889.
9.Cheng H, et al.2006. Human mRNA export machinery recruited to the 5=
end of mRNA. Cell127:1389 –1400.
10. Ganem D.2007. Kaposi’s sarcoma-associated herpesvirus, p 2847–2888. InKnipe DM, Howley PM (ed), Fields virology, 5th ed, vol 2. Wolters Kluwer/Lippincott/Williams & Wilkins Co, Philadelphia, PA.
11. Gatfield D, Izaurralde E.2002. REF1/Aly and the additional exon junc-tion complex proteins are dispensable for nuclear mRNA export. J. Cell Biol.159:579 –588.
12. Graham FL, Smiley J, Russell WC, Nairn R.1977. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol.36:59 –74.
13. Gruffat H, et al.2002. Epstein-Barr virus mRNA export factor EB2 is essential for production of infectious virus. J. Virol.76:9635–9644. 14. Gupta AK, Ruvolo V, Patterson C, Swaminathan S.2000. The human
herpesvirus 8 homolog of Epstein-Barr virus SM protein (KS-SM) is a posttranscriptional activator of gene expression. J. Virol.74:1038 –1044. 15. Han Z, Swaminathan S.2006. Kaposi’s sarcoma-associated herpesvirus
lytic gene ORF57 is essential for infectious virion production. J. Virol.
80:5251–5260.
16. Hautbergue GM, et al.2009. UIF, a New mRNA export adaptor that works together with REF/ALY, requires FACT for recruitment to mRNA. Curr. Biol.19:1918 –1924.
17. Jackson BR, et al.2011. An interaction between KSHV ORF57 and UIF provides mRNA-adaptor redundancy in herpesvirus intronless mRNA ex-port. PLoS Pathog.7:e1002138. doi:10.1371/journal.ppat.1002138. 18. Jeong J, Papin J, Dittmer D.2001. Differential regulation of the
overlap-ping Kaposi’s sarcoma-associated herpesvirus vGCR (orf74) and LANA (orf73) promoters. J. Virol.75:1798 –1807.
19. Johnson LA, Li L, Sandri-Goldin RM.2009. The cellular RNA export receptor TAP/NXF1 is required for ICP27-mediated export of herpes sim-plex virus 1 RNA, but the TREX comsim-plex adaptor protein Aly/REF appears to be dispensable. J. Virol.83:6335– 6346.
20. Katahira J, Inoue H, Hurt E, Yoneda Y. 2009. Adaptor Aly and co-adaptor Thoc5 function in the Tap-p15-mediated nuclear export of HSP70 mRNA. EMBO J.28:556 –567.
21. Kataoka N, Dreyfuss G.2004. A simple whole cell lysate system for in vitro splicing reveals a stepwise assembly of the exon-exon junction com-plex. J. Biol. Chem.279:7009 –7013.
KSHV ORF57-REF/Aly Binding Dispensable for Replication
on November 7, 2019 by guest
http://jvi.asm.org/
22. Kim VN, Dreyfuss G.2001. Nuclear mRNA binding proteins couple pre-mRNA splicing and post-splicing events. Mol. Cells12:1–10. 23. Kingston RE.2012. Preparation and analysis of RNA. Curr. Protoc. Mol.
Biol.98:4.0.1– 4.0.2.
24. Kirshner JR, Lukac DM, Chang J, Ganem D.2000. Kaposi’s sarcoma-associated herpesvirus open reading frame 57 encodes a posttranscrip-tional regulator with multiple distinct activities. J. Virol.74:3586 –3597. 25. Kronemann D, Hagemeier SR, Cygnar D, Phillips S, Bresnahan WA.
2010. Binding of the human cytomegalovirus (HCMV) tegument protein UL69 to UAP56/URH49 is not required for efficient replication of HCMV. J. Virol.84:9649 –9654.
26. Lei H, Dias AP, Reed R.2011. Export and stability of naturally intronless mRNAs require specific coding region sequences and the TREX mRNA export complex. Proc. Natl. Acad. Sci. U. S. A.108:17985–17990. 27. Luo MJ, Reed R.1999. Splicing is required for rapid and efficient mRNA
export in metazoans. Proc. Natl. Acad. Sci. U. S. A.96:14937–14942. 28. Majerciak V, Pripuzova N, McCoy JP, Gao SJ, Zheng ZM. 2007.
Targeted disruption of Kaposi’s sarcoma-associated herpesvirus ORF57 in the viral genome is detrimental for the expression of ORF59, K8␣, and K8.1 and the production of infectious virus. J. Virol.81:1062–1071. 29. Majerciak V, et al.2011. Kaposi’s sarcoma-associated herpesvirus ORF57
interacts with cellular RNA export cofactors RBM15 and OTT3 to pro-mote expression of viral ORF59. J. Virol.85:1528 –1540.
30. Majerciak V, et al.2008. Kaposi’s sarcoma-associated herpesvirus ORF57 functions as a viral splicing factor and promotes expression of intron-containing viral lytic genes in spliceosome-mediated RNA splicing. J. Vi-rol.82:2792–2801.
31. Majerciak V, Yamanegi K, Nie SH, Zheng ZM. 2006. Structural and functional analyses of Kaposi sarcoma-associated herpesvirus ORF57 nu-clear localization signals in living cells. J. Biol. Chem.281:28365–28378. 32. Malik P, Blackbourn DJ, Cheng MF, Hayward GS, Clements JB.2004.
Functional co-operation between the Kaposi’s sarcoma-associated her-pesvirus ORF57 and ORF50 regulatory proteins. J. Gen. Virol.85:2155– 2166.
33. Malik P, Blackbourn DJ, Clements JB.2004. The evolutionarily con-served Kaposi’s sarcoma-associated herpesvirus ORF57 protein interacts with REF protein and acts as an RNA export factor. J. Biol. Chem.279: 33001–33011.
34. Masuyama K, Taniguchi I, Kataoka N, Ohno M. 2004. RNA length defines RNA export pathway. Genes Dev.18:2074 –2085.
35. Neipel F, et al.1997. Human herpesvirus 8 encodes a homolog of inter-leukin-6. J. Virol.71:839 – 842.
36. Nekorchuk M, Han Z, Hsieh TT, Swaminathan S.2007. Kaposi’s sar-coma-associated herpesvirus ORF57 protein enhances mRNA accumula-tion independently of effects on nuclear RNA export. J. Virol.81:9990 – 9998.
37. Nojima T, Hirose T, Kimura H, Hagiwara M.2007. The interaction between cap-binding complex and RNA export factor is required for in-tronless mRNA export. J. Biol. Chem.282:15645–15651.
38. Ohno M, Segref A, Kuersten S, Mattaj IW.2002. Identity elements used in export of mRNAs. Mol. Cell9:659 – 671.
39. Palmeri D, Spadavecchia S, Carroll KD, Lukac DM.2007. Promoter-and cell-specific transcriptional transactivation by the Kaposi’s sarcoma-associated herpesvirus ORF57/Mta protein. J. Virol.81:13299 –13314. 40. Reed R, Cheng H.2005. TREX, SR proteins, and export of mRNA. Curr.
Opin. Cell Biol.17:269 –273.
41. Renne R, et al.1996. Lytic growth of Kaposi’s sarcoma-associated her-pesvirus (human herher-pesvirus 8) in culture. Nat. Med.2:342–346. 42. Rezaee SA, Cunningham C, Davison AJ, Blackbourn DJ.2006. Kaposi’s
sarcoma-associated herpesvirus immune modulation: an overview. J. Gen. Virol.87:1781–1804.
43. Rodrigues JP, et al.2001. REF proteins mediate the export of spliced and unspliced mRNAs from the nucleus. Proc. Natl. Acad. Sci. U. S. A.98: 1030 –1035.
44. Russo JJ, et al. 1996. Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8). Proc. Natl. Acad. Sci. U. S. A.93:14862– 14867.
45. Ruvolo V, Wang E, Boyle S, Swaminathan S.1998. The Epstein-Barr virus nuclear protein SM is both a post-transcriptional inhibitor and ac-tivator of gene expression. Proc. Natl. Acad. Sci. U. S. A.95:8852– 8857. 46. Sahin BB, Patel D, Conrad NK.2010. Kaposi’s sarcoma-associated
her-pesvirus ORF57 protein binds and protects a nuclear noncoding RNA from cellular RNA decay pathways. PLoS Pathog. 6:e1000799. doi: 10.1371/journal.ppat.1000799.
47. Shin YC, Desrosiers RC. 2011. Rhesus monkey rhadinovirus ORF57 induces gH and gL glycoprotein expression through the posttranscrip-tional accumulation of target mRNAs. J. Virol.85:7810 –7817.
48. Simizu B, Rhim JS, Wiebenga NH.1967. Characterization of the Tac-aribe group of arboviruses. I. Propagation and plaque assay of TacTac-aribe virus in a line of African green monkey kidney cells (Vero). Proc. Soc. Exp. Biol. Med.125:119 –123.
49. Strasser K, et al.2002. TREX is a conserved complex coupling transcrip-tion with messenger RNA export. Nature417:304 –308.
50. Stubbs SH, Hunter OV, Hoover A, Conrad NK. 2012. Viral factors reveal a role for REF/Aly in nuclear RNA stability. Mol. Cell. Biol.32: 1260 –1270.
51. Sun R, Lin SF, Gradoville L, Miller G.1996. Polyadenylylated nuclear RNA encoded by Kaposi sarcoma-associated herpesvirus. Proc. Natl. Acad. Sci. U. S. A.93:11883–11888.
52. Zhong W, Ganem D.1997. Characterization of ribonucleoprotein com-plexes containing an abundant polyadenylated nuclear RNA encoded by Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8). J. Virol.
71:1207–1212.
53. Zhou Z, et al.2000. The protein Aly links pre-messenger-RNA splicing to nuclear export in metazoans. Nature407:401– 405.
54. Zhu FX, Cusano T, Yuan Y.1999. Identification of the immediate-early transcripts of Kaposi’s sarcoma-associated herpesvirus. J. Virol.73:5556 – 5567.