Copyright ©D 1977 American Society for Microbiology Printed in U.S.A.
Further
Characterization of the Complementation Group B
Temperature-Sensitive
Mutant of Respiratory Syncytial
Virus
ROBERT B. BELSHE,* LINDA S. RICHARDSON, THOMAS J. SCHNITZER, DAVID A. PREVAR, ENA CAMARGO, AND ROBERT M. CHANOCK
Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, Bethesda, Maryland 20014
Received for publication 18 May 1977
ts-2, atemperature-sensitive and plaque morphology mutant of respiratory syncytial virus and sole representative of complementation group B, was
com-pared with members of the other complementation groups of respiratory syncy-tialvirus (group A [ts-1]andgroup C [ts-7]). ts-2 was found to be 10- to 1,000-fold morerestrictedingrowthand abilitytospreadat restrictivetemperatures (37,
38,and 390C) than at the permissive temperature(320C). In temperature shift-up experiments, thetsdefect ofts-1 and othermembers of complementation group A wasfoundtoaffect a late function that was required for at least 13 hinthe replicativecycle. Thetslesion of ts-7 affected afunction earlyinthe replication cycle. In contrast, ts-2 was not temperature sensitive when studied by the shift-uptechnique. The discrepancy between the ts plaque property and failure to detect temperature sensitivity during the shift-up experiment was resolved when it was shown that ts-2 had a defect in adsorption or penetration or both at the restrictive temperature. Clonal analysis of revertant ts-2 showed a
coordi-nate restoration of ts+ phenotype and syncytium-forming capacity. It appears thatts-2has a defectina proteinthat isinvolvedinadsorptionand/or penetra-tionof virus and isalso responsible for cell fusionactivity.
Seven temperature-sensitive (ts) mutantsof
theA2 strain of respiratorysyncytial (RS) virus
weredescribed and partially characterized
pre-viously (2, 4, 9, 10). Each was selected on the basis of a 100-foldorgreaterreductioninplaque
formationatthe restrictive temperatureof 39°C
compared with the permissive temperature of 32°C. Further characterization revealed that
the efficiency ofplaque formation at 3900 was
reduced at least 105-fold with each of the
mu-tants, exceptts-5,whichwas a"leaky"mutant.
Geneticanalysis of thesemutantsdefinedthree complementation groups: group A contained ts-1, ts-3, ts-4, ts-5, and ts-6; groups B and C
contained single mutants,ts-2and ts-7, respec-tively (10). Inaddition, ts-2 wasnoted to be a
plaque morphology mutant that did not fuse cellstoform syncytiaduring plaque formation
atthepermissive temperature (2).Inthis
com-munication we report on further studiesof the
complementation group B mutant, ts-2, and
compare itspropertiestothose ofgroupAandC mutants.
MATERIALS AND METHODS
Proceduresforgrowthof virusand assay of infec-tivity, aswell as the indirect immunofluorescence
technique, have been describedpreviously (9). Viruses and cell cultures. Thedevelopmentof the seven RS ts mutants was described previously in detail (2, 10). Six mutants, ts-1 to ts-6, were pro-duced after exposure of RS virustoeither 10-4 M 5-fluorouridine (ts-1tots-4)or10-2.5 M5-fluorouracil (ts-5,ts-6) in thegrowthmedium. ts-7wasisolated by Louis Potash after treatmentofRS virus with ethylmethane sulfonate. HEp-2 cellcultureswere obtained from Flow Laboratories (Rockville, Md.) andmaintainedasdescribed(2).
Growthof ts-2. The ts-2 mutant wasinoculated into a set ofHEp-2 roller tube tissuecultures at a multiplicityofinfection(MOI) of0.1 or0.001. Incu-bationwascarriedout at32, 37, 38,or39°C. Daily for5days,apairof tissueculturesfrom each experi-mental groupwasharvested andassayed for infec-tious virusat320Caspreviouslydescribed(9).
Temperature shift analysis. Each ts virus was inoculated at an MOI of0.1 into a setofHEp-2roller tube cultures thatwereincubatedatroom tempera-ture for 90 min. The cultures were then washed threetimeswithmedium, refedwithmedium, and allowedtoincubate at32°C. At varioustimes, two cultures wereshifted from 32 to 39°C. After24 h, each of the cultures was harvested, and the com-bined cellsand fluid mediumwereassayedat32°C for infectiousvirusbytheplaque technique.
Adsorption-penetration experiments. Theability of a repPesentative of each ts complementation
on November 10, 2019 by guest
http://jvi.asm.org/
GROUP B ts MUTANT OF RS VIRUS group to adsorb and penetrate HEp-2 cells at the
permissive or restrictive temperature was examined using twoindexes ofinfection, namely, production of infectious virus and production of viral antigens.
Production of infectious virus after adsorption at 32 or3900 was tested in the following manner. HEp-2 roller tube tissue cultures were maintained at either 32or39TCthroughout the adsorption-penetra-tion period. Virus was warmed to 32 or 39TC and then inoculated onto the HEp-2 cells, which were incubated at the same temperature. After 90min of adsorption, unattached virus was removed by wash-ingthe culture three times with medium warmed to 32 or39TC;the culture was thenoverlaid with pre-warmed medium containing 1% human convales-cent serumcontaining a high level of RS virus anti-body (plaque reduction titer of 1:1,024) and then againwashed three times with prewarmed medium. At no time were the 39TC cultures allowed to cool during the adsorption-penetration period. After being overlaid with prewarmed maintenance me-dium, the cultures were incubated at either 32 or 39TC. After24hof incubation, cells and fluid media wereassayed for infectious virus by the plaque tech-nique.
Production of viral antigens after adsorption-pen-etration at 32 or 3900 was assayed with Leighton tubeHEp-2 tissue cultures. Leighton tube cultures wereinoculated with virus, adsorbed, washed, and incubated ateither 32 or3900in thesame manner as described above for roller tube tissue cultures. After 24 hof incubation, cells were fixed andassayed by indirect immunofluorescence for production of viral antigen. Underthese conditions, the viral antigen-containingcells were predominantly those infected withthe input virus (8).
Clonal analysis of revertant ts-2. Revertant ts-2 virus was generated by incubating
replicate-in-10o A
32/
370
390
U.
fected HEp-2 cultures for 6 days at 35, 36, 37, 38, or 390C. Each culture was subpassaged twice, from culture to culture, at the original temperature of incubation. The second and third passages were ini-tiated by inoculation ofundiluted, pooled cells and medium to maximize the input multiplicity and hence the emergence of altered virus with thets+ phenotype (9).Syncytial cytopathiceffects were ob-served in some ofthe second-passage cultures in-fected with ts-2, and ts+ virus was also detected in thesecultures. To characterize the relationship be-tween these two events, i.e., restorationofsyncytial cytopathic effect and restoration of abilityto pro-duce plaques at 38°C(ts+ property), a clonal analysis of virusderived from 10separate cultures was per-formed. Clones were selected by picking plaques produced at 320C and subsequently passaging the plaque-purified virus one time in HEp-2 roller tube cultures to increase the quantityof virus for analy-sis (3). Atotal of57clones from the 10 cultureswere examined for their ability to form syncytiaand for efficiency ofplaque formation at 380C.
RESULTS
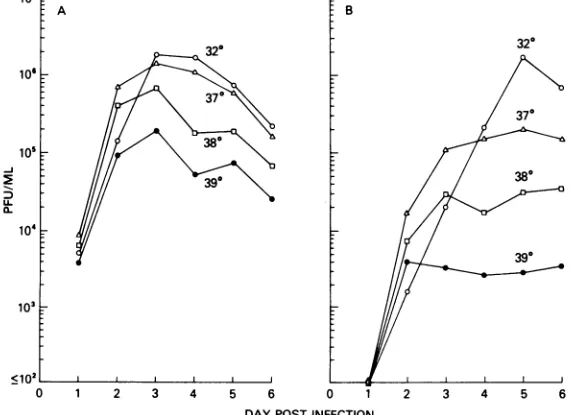
Effect of temperature on growth ofts
mu-tants. Growth occurred at each of the restric-tive temperatures (37, 38, or 390C); however, the level of replicationwasinverselyrelatedto
temperature (Fig. 1). Ineach instance, growth
at the restrictive temperature was reduced compared with growth at the permissive
tem-perature of 32°C. At the higher MOI (0.1), growth of ts-2 was much less restricted at390C
(10-fold reduction) than was the growth of ts-1 (100,000-fold reduction [9]) or ts-7 (1,000-fold reduction, datanotshown). The effect ofinput
5 6 0 1
DAYPOSTINFECTION
2 3 4 5 6
FIG. 1. Growthofts-2atpermissive (32°C)and restrictive(37,38, 39°C)temperatures.(A)MOI=0.1; (B)
MOI = 0.001.
0 1 2 3 4
9 VOL. 24, 1977
.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.504.109.398.443.651.2]multiplicityonthe peak titerattainedby ts-2at
restrictive temperature is shown in Fig. 1A
(MOI=0.1PFU/cell) andB(MOI=0.001PFU/ cell). At each of the restrictive temperatures,
the peak titerwashigherwith thehigher MOI.
However,aneffect ofMOIonlevel ofvirus
rep-licationwasnotseenatthe permissive
temper-ature. Inculturesincubated at390C, the viral titer peaked after 2 days of incubation, and
further growthwas not observed. Incontrast,
growth was seen after 2 days when cultures were incubated atthepermissivetemperature
(Fig. 1B, 320C growthcurve).
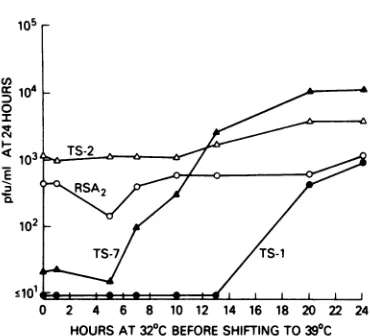
Temporal expression of thets lesion.
Tem-perature shift-up revealed that growth ofthe wild-typeviruswasnot affected when cultures were shifted from32 to 390C (Fig. 2). ts-1 was
previously found tohave alate lesion,
requir-ingabout 13 h of incubation atthe permissive temperaturebefore completion of thets-1 func-tion (L. S. Richardson, T. J. Schnitzer, R. B.
Belshe, E. Camargo, D. A. Prevar, and R. M. Chanock, Arch. Virol., in press). In thepresent study this was confirmed when it was shown
thatatleast13hofincubationwasrequiredat the permissivetemperatureforinfectiousvirus to be produced. When other complementation
group A viruses were tested by temperature
shift-up, eachexhibitedapatternsimilarto ts-1
(data notshown). ts-7had anearly lesion and
required 5 h of incubation at the permissive
temperature forproduction of infectious virus
(Fig.2). Anunanticipated findingwasthatthe
growth ofts-2wasnotaffected byashift-up in
temperature at any time in the 24 h of viral
growth (Fig. 2).
ts-2wastemperaturesensitivewith regardto
plaque formation, but the mutant was not ts
when studied by temperature shift-up. Possi-bly, thiscontradiction reflectedanearlydefect
which restricted the initial stage ofinfection,
but which didnot affect virusreplicationonce
infectionwasinitiated. To investigate this
pos-sibility, the ability of ts-2 to adsorb and
pene-tratecellswasstudiedat32and390C. Adefect
inthispropertycould explain theapparent
dis-crepancyinourfindingswith ts-2;the tempera-ture shiftexperimentmeasures onlyone cycle
of growth, but formation ofplaques involves multiplegrowth cycles (andthereforemultiple cycles of adsorption and penetration). Wild-type RS virus, ts-1, ts-2, and ts-7 were tested
for theirabilitytoabsorb andpenetrate at32or
390C (Tables1and2).Wild-typevirus adsorbed andgrewequallywellat32and 390C. ts-1 and ts-7 adsorbedequally well at32 and390C, but neithermutantgrewwellat390C (Table 1)nor
producedasmuch antigenat390Cascompared
Crn104 o
0
I-~~~~~~~~~~>
102
TS-7/ /TS-1
[image:3.504.269.454.69.237.2]0 2 4 6 8 10 12 14 16 18 20 22 24 HOURS AT320C BEFORE SHIFTING TO 390C
FIG. 2. Effect of time of shift from 32 to 39°C temperatureof incubationonthe titerof wild-type RS
(0), ts-1 (O), ts-2 (A), and ts-7 (A)at24 h; MOI=
0.1.
TABLE 1. Effect ofadsorptiontemperatureonthe
growth ofwild-type RSvirus, ts-1, ts-2,and ts-7 incubatedfor 24 hatpermissiveorrestrictive
temperature
Quantity ofvirus pro-Adsorption duced
after
incubation Virus temp attheindicatedtempVirus
tempICY
for24h320C 39'C
Wildtype 32 4.6b 4.3
39 4.6 4.5
ts-1 32 4.3 <1.0
39 3.9 <1.0
ts-2 32 3.8 3.6
39 2.5 1.6
ts-7 32 5.3 2.9
39 5.3 2.5
aHEp-2cells were inoculated with the indicated
virus atan MOI of 0.1(wild type,ts-1,and ts-2) to0.3 (ts-7),incubated for 90 min at 32 or39TC,and then washed with medium containing RS virus antibody, refed,andincubated at 32 or390C.
bPFU permilliliter(log1O).
with 320C (Table 2). ts-2, however, did not
ad-sorb and/or penetrate cellsefficiently at 390C.
Adsorption ofts-2 to HEp-2 cells was
approxi-mately20-to100-foldlessefficient at 390C than at320C (Tables 1and2). The decrease inviral
growth or production of viral antigens when
adsorption-penetration wasallowed to occurat 390C cannotbe ascribed toheat labilityof the ts-2mutant,sincethisviruswasshowntobe as
stable as wild-type virus at this temperature
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.266.459.305.488.2]GROUP B ts MUTANT OF RS VIRUS TABLE 2. Effectoftemperatureofadsorptionand incubation onproduction ofviralantigensinHEp-2cellsby
RSvirusandits ts mutants
Cells containing Fold reduction inpercentageofcellscontaining viralantigens viral antigens fo
32'C
adsorption39'C
adsorption39'C
adsorptionRS virus
(329C
adsorptionfor for 90min;then for 90min;then for 90min;then 90min;then 32C 39'Cincubation 32*Cincubation 39'Cincubation incubationfor 24h) for 24 h for24 h for 24hWild type 11.0 0 0 0
ts-1 15.0 75.0 1.9 37.5
ts-2 10.4 1.9 20.8 52.0
ts-7 15.0 6.3 1.7 21.4
a HEp-2 cells were inoculated with an MOI of approximately 0.1, incubated for 90 minat32 or
390C,
washed with medium containing RS virus antibody, refed, andincubated at 32 or 390C.bTested byindirect immunofluorescence withhumanconvalescentserum.
(4). Once ts-2 waspermittedtoadsorbtoHEp-2
cells at 320C, transfer to 390C for subsequent incubation did notreducevirusyield or antigen
productionasmarkedly aswith the ts-1 or ts-7
mutant.
ts phenotype and plaquemorphologyof re-vertant ts-2. Serial passage of ts-2 at 35 to390C
yielded some virus that produced plaques at
380C (i.e., ts+) andinduced formation of
syncy-tia. This phenomenonwasobservedduringthe second and thirdpassages.Inthethird-passage
cultures, mostof the virusappearedtobe wild
type since these virus suspensions produced plaquesat380Cwithanefficiency equivalentto
wild-typevirus, and mostplaqueswere
syncy-tial. Incontrast, viruses inthe second-passage
cultures were intermediate between the ts-2
mutantand wild-typevirus withrespect to
pla-quing efficiency at
380C
and plaquemorphol-ogy. Todeterminewhether all of the virus
par-ticles in the second-passage suspensions were
intermediate in ts+ and plaque morphology
phenotypeorwhetheramixed population ofts
andts+virusesand nonsyncytial and syncytial viruses were produced, a clonal analysis was
performed. Fifty-seven clones derived from 10
separate cultures, 5 incubated at 35°C and 5
incubated at 37°C, were selected by picking individual plaques at
320C.
Afterone passage in HEp-2 roller tube cultures at320C,
theseclonal populations were evaluated forts+ and
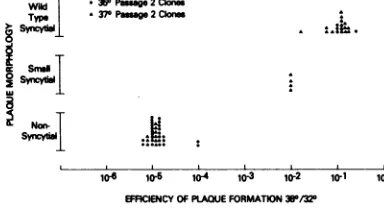
syncytialproperties(Fig. 3).Thirty-four clones
that did not produce syncytia plaqued with greatly reduced efficiency at
380C
or notat all(plaque titer at 38°C/titer at 32°C was 10-4 to
10-5). Nineteen clones, derived from sixofthe
second-passage cultures, formed wild-type syn-cytiaand formed plaques at38°Casefficiently
asthe wild-type virus. Inaddition, fourclones
that formed syncytia smaller than those of
wild-typevirus wereintermediate intheir
effi-ciency of plaque formationat
380C.
Thus,there wascoordinaterestoration of thets+andsyncy-Wild
Type
"I SynCYsu
E0
sT
Sn Tu25
2
Nom. Sagcy1iel*355 Pase 2 Clnes 37 Pasus 2 Clons
104 10-6 104
[image:4.504.254.446.237.345.2]EFRCIENCY OF PLAQUE FORMATION35'IW
FIG. 3. Plaque morphology of clones of ts-2 com-paredwithefficiency of plaqueformation atthe re-strictivetemperature
(38QC).
The cloneswereselected fromts-2 viruspassagedtwiceat35°C (0) or37°C (A).tial properties.Furthermore, virusesthatwere
intermediateinplaguing efficiencyat380Calso exhibitedanintermediate typeofsyncytial
cy-topathology.
DISCUSSION
Littleisknownofthestructureand function ofRS viral proteins, because the virus doesnot grow to high titer in tissue culture and it is
difficulttopurifythe virustopermit
biochemi-calanalysis of viralpolypeptides. Despite these
difficulties, two groups have presented
evi-dence for the existence of two to three viral
glycoproteins (5, 11). These glycoproteins
pre-sumably are the spikes that project from the
viralenvelope. The function of the spikes of RS virushasnotbeendefined, butitisreasonable
toassumethattheyareinvolvedinattachment to the host cell and in cell fusion, which is a
characteristic effect of thisvirus in a number of
cell cultures. Theparamyxoviruses, which
re-sembleRS virus inpossessing alimiting
enve-lope withspikelike projections and in causing cellfusion, havetwodistinct surface
glycopro-tein spikes. One of these glycoproteins
func-tions as ahemagglutinin and also has neura-VOL. 24, 1977
10-3 10-2 lo-' 10°
on November 10, 2019 by guest
http://jvi.asm.org/
12 ET AL.
minidase activity (1). The other glycoprotein causes cell fusion and appears to be involved in penetration of virus into the host cell (7). RS virus has not been shown to possess hemagglu-tination orneuraminidaseactivity (6) and thus cannotbeconsidered strictly comparableto the
paramyxoviruses. However, RSvirus must at-tach to the host cell, andthis function could be
considered comparable to hemagglutination.
The prominent property of syncytium forma-tionexhibitedby RS virussuggests that it pos-sesses aproteinthatcausescell fusion.
Against this background of relative
igno-rance of RS virus structure and function, the
group B ts mutant, ts-2, was studied in an attempt todescribe thetsdefectmoreprecisely withrespect tothetimeandnature of the
func-tion affected. Although ts-2 was temperature sensitive with respect to plaque formation, it was nottemperature sensitivewhenstudied by shift-up. In this procedure, the adsorption of
virus wasallowed to occur at room temperature
before incubation at 32 or 390C. The
discrep-ancy between the ts phenotype ofts-2 when studiedby the plaque technique anditslack of
temperaturesensitivitywhen characterized by shift-upsuggested that thismutantmight have
adefect that affected the initialstage of
infec-tion, namely, adsorption and penetration. Sub-sequently, this was shown to be the case. At
thistime adistinction between adsorption and penetrationcannotbemade,since240C, a tem-perature close to that which prevailed in the laboratory, ispermissive for growth of RS vi-rus, andthus both adsorption andpenetration
could have occurredatthis temperature. ts-2 is of special interest since it is a plaque
morphologymutant.Itdoesnotproduce
syncy-tialplaques under permissiveconditions (2). It istempting tospeculatethatthetsdefect ofts-2
affects the protein responsible for cell fusion and, presumably, the protein for penetrationas
well. Ifthisisthecase, asingledefectcould be invoked to explain the nonsyncytial plaque property atthe permissivetemperatureand the
failure ofadsorption-penetration at restrictive temperature. Such a unitary hypothesis
neces-sarilyimplies that the ts-2 defect isexpressedat the permissive temperature (i.e., nonsyncytial plaques) and becomes progressively more re-strictivewithincreasing temperature. It is also
possiblethat the ts-2 virion fusion protein,
al-thoughdefective atpermissive temperature, is stillsufficientlyactivetopermitpenetrationto
occur, whereasthe activity of the affected pro-tein is not sufficient at the membrane of the
infected cell to allowcell recruitment to occur
by fusionof membranes.
An alternate explanation for the
defective-ness ofts-2 is that the mutant contains two separatelesions, one that is ts and affects ad-sorption-penetration and another thatisnot ts
and affects cell fusion. This is not likely the case, however, since cell fusion is probably a
manifestation of the same activity responsible
for cell penetration as has been shown for the paramyxoviruses (7). In addition, coordinate restoration of syncytium formation and the ability to form plaques atrestrictive tempera-ture was observed in clones selected from six
independently derived virus populations
con-tainingrevertant ts-2.Thissuggeststhata sin-gle lesionwasresponsible for both the aberrant plaque morphology and thets property.
LITERATURE CITED
1. Choppin,P.W.,and R. W. Compans. 1975. Reproduc-tion ofparamyxoviruses, p. 95-178.In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 4. Plenum Press, New York. 2. Gharpure, M. A., P. F. Wright, and R. M.Chanock.
1969. Temperature-sensitive mutants of respiratory syncytialvirus. J.Virol. 3:414-421.
3. Hodes, D. S., H. W. Kim, R. H. Parrott, E. Camargo, and R. M. Chanock. 1974. Genetic alteration in a temperature sensitive mutant of respiratory syncy-tial virus afterreplicationin vivo. Proc. Soc. Exp. Biol. Med.145:1158-1164.
4. Kalica, A. R., P. F. Wright, F. M. Hetrick, and R. M. Chanock. 1973.Electron microscopicstudies of respi-ratory syncytial temperature-sensitive mutants. Arch.Gesamte Virusforsch. 41:248-258.
5. Levine, S. 1977. Polypeptides of respiratory syncytial virus.J.Virol. 21:427-431.
6. Richman, A.V., F. A. Pedreira, and N. M. Tauraso. 1971.Attemptstodemonstrate hemagglutination and hemadsorption by respiratory syncytialvirus. Appl. Microbiol. 21:1099-1100.
7. Scheid, A.and P. W. Choppin. 1974. Identification of biological activities of paramyxovirus glycoproteins. Activation of cellfusion, hemolysis, andinfectivity byproteolytic cleavageofan inactiveprecursor pro-teinofSendaivirus.Virology57:475-490.
8. Schieble, J. H., A. Kase, and E. H. Lennette. 1967.
Fluorescent cell counting as an assay method for respiratorysyncytialvirus. J. Virol. 1:494-499.
9. Schnitzer, T.J.,L. S.Richardson,andR.M.Chanock.
1976.Growthand genetic stabilityof thets-1mutant ofrespiratorysyncytialvirus atrestrictive
tempera-tures.J. Virol. 17:431-438.
10. Wright,P.F.,M.A.Gharpure,D.S.Hodes, andR. M.
Chanock.1973.Geneticstudies of respiratory syncy-tial virustemperature-sensitive mutants. Arch. Ge-samteVirusforsch. 41:238-247.
11. Wunner, W.H., andC. R. Pringle.1976. Respiratory syncytialvirusproteins.Virology73:228-243.
on November 10, 2019 by guest
http://jvi.asm.org/