JOURNALOFVIROLOGY,Nov. 1974, P. 1115-1125 Copyright0 1974 AmericanSociety for Microbiology
Vol. 14, No. 5 Printed inU.S.A.
Cold-Sensitive
Mutants
of
Bacteriophage
OX174.
II.
Comparison
of
Two
Cold-Sensitive Mutants
DAVID J. SEGAL' AND CLIFION E. DOWELL
Department ofGenetics, University of California, Davis, California, 95616andDepartmentofMicrobiology,
University
of Massachusetts, Amherst,
Massachusetts01002Receivedforpublication 17 June 1974
Cold-sensitive
bacteriophage 4X174 mutants,
another class of conditionallethals, were
examined with regard to growth parameters, DNA synthesis, and
particle
properties. Two mutants, cs7O and cs82, were examined. Mutant cs70
was
eclipse
defective,
showing
altered
eclipse kinetics
atpermissive
temperature
(40
C) and
failing entirely to eclipse at restrictive temperature (25 C). Mutant
cs7O
replicated well at 25 C if allowed prior eclipse at 40 C. Mutant cs82 had
wild-type
eclipse at both temperatures but was defective in single-strand
synthesis
at
25C,
which led
todelayed
progeny
phage appearance, decreased
progeny
phage synthesis rate, and greatly reduced burst size. The cs82 block
could
notbe
bypassed by
temperature
shift.
Since
complementation analysis
of
cs7O
and cs82 was not
feasible due to the unique properties of these
mutants,
those 4X174
properties
affected
by
the virus
coat wereexamined
as anindex of
amutation
in a coatprotein gene.
Mutantcs7O had aberrantattachment kinetics
at
both
25C and
40C,
evidence of
a coatprotein
alteration. Mutant cs70 also
exhibited
significantly
decreased thermal
stability,
further
evidence of an altered
virus
structure.Mutant
cs82had increased thermal
stability,
but the difference
was not
sufficient
to
allow
unequivocal assignment of this mutant to a coat
protein gene. Both mutants had
wild-type antiserum inactivation and host
range,
although
cs70
was
subject to less of (low-level) plating restriction by
endogenous
F+ factors.
Bacteriophage
OX174,
because of its limited
genome size of
about 5,500 nucleotides, has
been
examined
by
many
laboratories with the
objective
of
elucidating
all of the virus
func-tions.
To this
end,
nine
(and
perhaps
ten)
genes
have been defined (4, 5)
through
the
useof
conditional
lethal mutants of
the
temperature-sensitive
and nonsense types. In 1967, Dowell
(15) isolated a class of
4X174
mutants
which
failed
togrow
atlow temperature;
these
mu-tants were
called cold-sensitive
(cs) since they
were
phenotypically opposite to
temperature-sensitive
mutants. It
wasanticipated that such
mutants
might define
previously
uncharacter-ized
4X174
functions. In that preliminary
re-port,
twoclasses
ofcold-sensitive mutants were
described: those
defective
ineclipse, and those
whose cold
sensitivity
wasmanifest
insome
later
replication step(s).
Thepresent paper
compares the
growth properties
ofcs7O (an
eclipse mutant)
with cs82(a
replication
mu-tant)
atboth
permissive (40
C)
and restrictive
' Present address:DepartmentofPediatrics, Universityof Alberta,Edmonton,Alberta,Canada.
(25 C)
temperatures. The results of
single-step
growth
experiments and
intracellular
DNA
analyses show that the cs7O mutation
prevented
eclipse
completely
at 25C
and
partially
at 40C,
but
had little effect
onsubsequent replication
steps at either temperature.
The cs82 mutation,
on
the other
hand,
had
noeffect
oneclipse
at
either
temperature, but
severely inhibited
sin-gle-strand (SS) DNA synthesis at 25 C.
At
least four
4X174
genes
specify virus
struc-tural
proteins, and the
growth properties of the
cs7O
mutantsuggested
that it
would be
lo-cated
in a coatprotein gene.
Although
4X174
mutations have
been
mapped
through both
complementation
analysis (8, 39; C. A.
Hutchi-son, Ph. D.
thesis, California Institute of
Tech-nology,
Pasadena, 1969) and recombination (2,
4; C. A.
Hutchison,
Ph. D.
thesis),
several
properties
ofcs7O and
cs82precluded the use
ofthese
conventional
genetic
techniques (see
Dis-cussion).
Instead, evidence that cs7O
is a coatprotein
mutant wassought by examining those
properties determined
by
the viruscoat, such
asparticle
stability,
attachmentkinetics,
anti-1115
on November 10, 2019 by guest
http://jvi.asm.org/
serum
inactivation,
and host range. Otherin-vestigators have also used such
nongenetic
cri-teria to assign
OX174
mutations to coatproteingenes (2, 7, 14, 16, 21, 28; C. A. Hutchison, Ph. D. thesis).
Thispaper wastaken inpartfroma
disserta-tion submitted by D. J. Segal in partial
fulfill-ment.of the requirements for thePh. D. degree
at the
University
of California. A preliminary account of some of these results has beenpresented (D. J. Segal and C. E.Dowell, 1971,
Proc. Can. Fed. Biol. Sci. 14:161).
MATERIALS AND METHODS
Bacterial strains. The strains used were: (i) Esch-erichia coli C (BTCC 122), with genotype sup-, thy+,
oX8,
the usual host for kX174; (ii) E. coli HF4704, withgenotype sup-, thy-,OX8,
hcr- (27); (iii) E. coli HF4714, with genotype sup+, thy+,OX8,
a C-K-12 hybrid (C. A. Hutchison, Ph.D. thesis); and (iv) E. coli C-2, (v) HF4704F+,and (vi) CRF+, which are male derivatives of E.coli C, HF4704, and CR (15).Viral strains. Mutants cs7O andcs82 wereisolated and described previously (15). Amber mutants were
obtained from R. L. Sinsheimer;am33(A), aml4(B), amlO(D), am3(E), am9(G), and am23(H) were iso-lated by C. A. Hutchison (Ph. D. thesis),am88(F) by F. Funk (21), and amN-1(H) by M. Hayashi (34). Double mutants ofam3 and cs7O or cs82 were con-structedaccordingtoC.A.Hutchison (Ph. D. thesis).
Media. Top agar, bottom agar, and KC broth were
as previously described (16) except that KC broth contained0.001 MCaCl2. C-mediumisTPG medium (22) containing 0.1% Casamino Acids and 0.001 M CaCl2. Starvation buffer has been described (13).
Sodium tetraborate solution is a solution ofsodium tetraborate saturatedat 4C.
Chemicals. The chemicals [methyl-3H]thymine
(23 Ci/mmol), [methyl-3H]thymidine (50 to 60 Ci/
mmol),
[methyl-'4C]thymine
(1to5mCi/mmol),Tri-ton X-100, and Aquasol were purchased from New
England Nuclear (Boston, Mass.). Spectrafluorwas
from Amersham/Searle Corp. (Des Plains, Iowa).
Pronase (Calbiochem, Los Angeles, Calif.), was self-digested for2hat37C and2min at 80Casdescribed by Iwaya and Denhardt (26) Sarkosyl(Geigy Chemi-cal Corp. Ardsley, N.Y.) was 10% in 0.8% EDTA containing 0.1 M Tris buffer. Mitomycin C was obtainedfrom SigmaChemical Co. (St. Louis,Mo.),
andlyophilizedlysozymefromMann ResearchLabs., Inc. (New York, N.Y.). Phenol was redistilled and storedat -20C in asealedcontainer.
Biological assays. Assaysofinfectivecentersand intracellular phage were performed as described by Denhardt and Sinsheimer (13), using the soft-agar overlaymethod(1).Indicatorbacteriaweregrownto 3 x 108cells/mlandkeptinanicebath untilrequired.
Starvation synchrony. A stationary culture was
diluted at least 1:10,000 into growth medium and
allowedtoreach adensityofabout2x 108cells/mlat 37 C with aeration.Cellswerecollectedby centrifuga-tion, washed twice with starvationbuffer,andstarved
for90minat37C (13). A sampleofstarved cellswas
placed at 40C for 5 min and then infected at a multiplicity of infection (MOI) of about 5. After 15
min toalloweclipse, growthwasinitiatedbydiluting
1:100in starvation bufferat 4C and then 1:100 into growth medium at the desired temperature. For DNA-labeling experiments, after eclipse as previously described growthwasinitiatedby collecting infected cellsby centrifugation, washing once, and resuspend-ingat input density infresh growth medium at the propertemperature.
Burst sizedetermination. Cultures of E. coli C or HF4704weregrownto 2 x 108 cells/ml in KC broth or C-medium, and synchronously infected at a MOI of about 5asdescribed above. Growth was initiated by 1:10,000dilution intoappropriate medium at 40 C or 25C. Wild-type, cs7O, and cs82 burst sizes were calculated as the ratio of infectious centers at t = 60 (for 40C) or t = 240 (for 25 C) to input infectious centers at t = 0. Burst sizes foram3, cs70am3, and cs82am3were calculated as the ratio of intracellular phage att = 120(for 40C) ort = 240(for25C)tothe number ofinput-starved cells measured just prior to phage addition.
Preparation of '4C-labeled SS DNA. E. coli HF4704 was grown to 3 x 101 cells/ml in 500 ml of C-medium containing 2
,gg/ml
ofthymine, and syn-chronously infected with am3 (MOI = 5) in C-me-diumsupplemented only with 2.4 Mg of["4C]thymineper ml. After 180 min of infection, the cells were collected by centrifugation, suspended in Tris-lysozyme-EDTA (13), and lysed by six cycles of freeze-thaw. Saturated borate-eluted intracellular phage particles were purified twice by isopycnic centrifugation and dialyzed against 0.05 M sodium tetraborate. DNA was extracted by the hot-phenol method (24), with removal of residual phenol by dialysis against0.05Mborave.
Measurement of DNAsynthesis.At 40C,
OX174
DNA synthesis was measured by the method of
Lindqvist and Sinsheimer (27), in mitomycin C-treated cells growing in C-medium supplemented with 2gg of nonradioactive thymine per ml and 10
MCi
of[3H]thymine
per ml. At 25C, 4X174DNA syn-thesis was measured usinga method similar tothat ofIwaya and Denhardt(26); cellswerestarved, mito-mycin treated, andsynchronously infected in theab-sence of exogenous thymidine or thymine, since at
lowertemperaturesE.coli HF4704nolonger requires these supplements (12). Virus DNA synthesized at 25 C was labeled incells growing inC-medium sup-plemented with 10
MCi
of [3Hlthymidineperml plus20 ug of adenosine per ml to potentiate thymidine uptake (12).
Analysis of intracellular DNA. Infected 20-ml
portionsofE. coli HF4704 cultures were pulsed with
[3H]thymidinefrom 60 to 240min; the cultures were
chilledby addition of anequal volumeof 4C starva-tionbuffer; and the cellswere collected and washed threetimesby centrifugation. Thepelletwaslysedin
Tris-lysozyme-EDTA and treated withSarkosyl and
Pronase asdescribed (26).DNA wasdeproteinized by
roomtemperature phenol extraction, precipitated by isopropanol, and dissolved in 0.1 M NaCl as
on November 10, 2019 by guest
http://jvi.asm.org/
COLD-SENSITIVE4X174 MUTANTS
ouslydescribed (16);efficiencyofrecoveryof
phage-specific DNA was 67 to 77%. Samples containing
equal volumes of
['H]DNA
and['4C]SS
DNA werelayered over linear 5 to 20% sucrose gradients contain-ing 0.5 M NaCl and 0.05 M sodiumcitrate (pH 7) and centrifuged at37,000 rpm for 5.5 h in an SW50.1 rotor of a Beckman L2-65B ultracentrifuge. The gradient tube was punctured, and 3-dropfractions were col-lected in scintillation vials and counted using a
scintillationcocktailconsistingof1,875 ml oftoluene,
1,000ml of Triton X-100, 333 ml of water, and 125 ml
ofSpectrafluor.
Eclipse studies. E. coli C cultures were starved
and synchronously infected at 25 C or 40 C as
de-scribed above. Periodic samples were diluted 1:100
intoKCbroth at 4 C, and the number of uneclipsed particles was determined by intracellular assay at permissive temperature. Starvation conditions, which permit
4X174
attachmentandeclipse, but not DNA replication, were employed to prevent eclipse frombeingmaskedbysubsequentvirusreplication.
Thermalstabilitystudies. Virus stocks in 0.05 M
sodium tetraborate weredilutedto 1x1010 to 5 x
10"0
PFU/ml in saturated borate solution and left
over-nightat 4C.The experiment wasbegun by adding0.1
ml ofphage solution to 9.9 ml of saturated borate
maintainedat56.5Cin aconstant-temperature water
bath. Samples were diluted 1:100 intoKC broth at
4C, and survivors wereplatedonE. coli HF4714at
37C. Wild-type survival was 1 x 10-5to 3 x 10-'
after 180 min ofexponentialinactivationunder these conditions. Inactivation rates and survival ratios were
calculatedasdescribed(35, 39; C. A.Hutchison, Ph.
D. thesis).
Attachment studies. Attachment kinetics were measured essentially as described by Newbold and Sinsheimer (28). E. coli C, grown to 2.5 x 10' cells/ml
and starved in the usualfashion,wasequilibratedat
25 or 40C. Phage stocks were diluted in starvation buffer to 1.2 x 10'PFU/ml and
temperature-equili-brated;equalvolumes ofcs7O(or cs82)and am3were
combined, and the experiment was begun by
addi-tion of 0.1ml ofphagemixture(1.2 x 107phage)to5 ml ofcells(1.2 x 10' cells).The culture wasaerated,
and 0.1 ml samples werediluted into 9.9 ml ofKC
broth (at4C) containing2 x 109E. coliK-12 W-6 carrier cells, astrain to which 4X174cannot attach (15, 35). Cells and adsorbed virus were collected by
centrifugation, and the supernatant fluids were
ti-trated for unattachedcsphagebyplating onE. coli C at 37C and for am3 phage by plating on E. coli HF4714 at 25C.Wild-typeattachment kinetics were measured in the same way, but not in the same tube aswith am3.Thedesignfulfilled the following crite-ria. (i) am3 andcs7O(orcs82) were in the same tube to normalizesamplingerrors;(ii)MOIswere lowenough toensure nocooperative effect of attachment on
ad-sorptionofsubsequent particles(1). (iii) A dilution of
1:100 effectively stopped further attachment by
re-ducing cell concentration to below 2 x 106 cells/ml
(20). (iv) Calculations were based on only the
first-orderportion of attachment kinetics. Attachment rate constants weredetermined by the method of Adams
(1).
Antiserum inactivation. Experiments were per-formed by the method of Rolfe and Sinsheimer (31). Virusstocks were diluted at least 100-fold into 10 ml of KC broth at 37C,to afinalvirus concentration of 5
X 107PFU/ml.Anequalnumber of am3particles was
added to each tube, and the phage mixtures were equilibrated at 37 C for 15 min. The
OX174
antiserum(attachmentrate constant k = 2 x
104/min;
kindly
supplied by D. T. Denhardt) was diluted 1:500 into KC broth, and the experiment was begun by adding 0.1ml of antiserum mixture to10ml of virus mixture
(final k = 4 x 10-
'/min). Samples
taken at 1-minintervals were diluted 1:100 into KC broth. Wild-type and cssurvivors were assayed on E. coli C at 37 C, and am3survivors were assayed on E. coli HF4714 at 25 C. Phage mixtures provided an internal control against variation in sampling time and dilution. Underthese
conditions, itwaspossible to obtain 10 points on the
inactivation curveand yet remain within the quanti-tative limits of the Adams equation for calculating antiserum k values (1).
RESULTS
Eclipse
studies. Examining the efficiencies
of
plating
of
wild-type
and
cs7O and
cs82 at 25C
and
40C,
it was found that wild type had anEOP
of 0.55 at 25C relative
to 40C,
but that
cs7O
and
cs82had efficiencies of
plating
of 1.6 x10-7 and
1.7 x 10-5,respectively,
overthe
samerange.
The
initial
report onthese
mutants(16)
suggested that
cs7O
cold
sensitivity
wasdue
to ablock
ineclipse,
although
cs82 wasthought
tobe cold sensitive
insubsequent replication.
Eclipse
kineticsexperiments
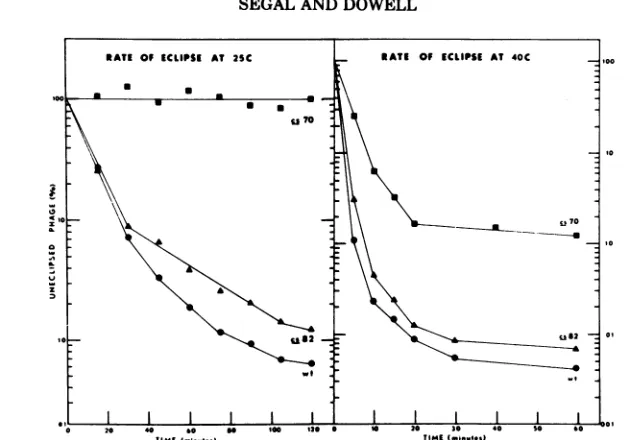
(Fig. 1) showed
that
at25C,
nocs7O PFUs
werelost
during
the
interval
inwhich
morethan
98%of
cs82and
wild-type
particles became
eclipsed.
In
addi-tion,
cs7O
eclipsed
at aslower
rate at40C and
had
ahigher
level of
noneclipsed particles
than
the
other
twoviruses. These
results showed
conclusively
that
the
cs7O mutation inhibited
eclipse completely
at25C and
partially
at40C.
The
cs82mutation,
onthe
other
hand,
had little
effect on
eclipse
ateither
temperature.Experi-ments
with am3,
cs70am3,
and
cs82am3 showed
a
similar
relationship.
Growth
experiments.
Single-stop growth
ex-periments
wereperformed
to see ifthe
cs7O
mutation
affected
anyposteclipse
function
and
also
todefine the
cs82replication
defect.
Atpermissive
temperature(40
C),
cs7O, cs82, and
wild
type hadsimilar
growth kinetics,
witheclipse
times of 12min, latentperiods
of14min,and burst
sizes of148,
83, and
89 forcs7O,
cs82,
and
wild-type,
respectively.
The
growth
ofcs7O
at 25
C
(Fig. 2),
aftereclipse
at40C,
wasverymuch
likewild-type,
whichsuggested
that
thecs7O
mutationhad
little effect on anyreplica-tion steps
subsequent
toeclipse.
Thegrowth
of1117
VOL.14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
TIME
FIG. 1. Eclipse ofwildtypecs7O, and cs82 at 25 C and 40 C. Starved E. coli C was infected at 25 or 40 C with
wild type, cs7O, or cs82 at MOIabout 4.Samples were diluted 1:100 into KCbroth at 4 C and assayed for intracellularphage. Uneclipsedphage is percentage(PFUper ml, after intracellular assay, of sample at t =X dividedbyPFU per ml, after intracellular assay, of sample at t = 0).
cs82 at 25 C, on the other hand, showed delayed
onset
of
intracellular phage
production,re-tarded
rateof
intracellular
synthesis, andgreatly reduced
burst size. This growthinhibi-tion of cs82
at 25C,
contrasted with the
wild-type
growth of cs70
atboth
temperatures(after
permissive
temperatureeclipse),
wasobserved
in
avariety of
growth
conditions
(Table 1).
The data in
Fig.
2suggested
that
cs82 wasdefective
notonly
in initiation of
intracellular
phage synthesis but also in
somesubsequent
process(es). An
infected
culture
wasshifted
from
40C
to25C after
20min ofinfection, with
the
rationale that
ifthe
cs82mutation
affected
a
function
continually
required
for
phage
syn-thesis, production would
ceaseafter
adown-shift.
The data in
Fig.
3showed that wild
type(am3)
continued
replication after such
ashift,
but that the cs82am3
culture
ceased
replication.
Atthe end of a 20-min
period,
replicative
form(RF) formation, replication,
transcription,and
translation
areessentially complete (32, 35),
sothat
anRF
defect
was initself
notsufficient toexplain
cs82cold sensitivity. It
wasalso
notlikely that
alysis defect
wasresponsible
for cs82cold sensitivity,
since inthe absence
oflysis
at25
C
the cs82 mutation stilldelayed
appearanceof
intracellular
phage, retarded intracellular
synthesis
rate,and reduced burst
size(experi-ment not
shown).
DNA
synthesis.
The growth experimentsshowed
that cs82growth inhibition
at25C
wasdue
toablock
in somelate
replication
processother than
RF
replication
orlysis. To determine
if the
cs82mutation
affected
progenySS DNA
synthesis
at 25C, cs82am3-infected cultures
were
labeled with [3H]thymidine from 60
to 240min
during the
period
of
SS
synthesis, and the
intracellular
phage DNA
wasanalysed
on asucrose
gradient. The results (Fig. 4) show that
the
cs82mutation
severely
inhibited
SS
synthe-sis
at 25C,
but
permitted normal synthesis of
RF I and RF II,
relative
toboth
am3and
cs70am3.
The
[3H]SS DNA peak sedimented
two to
three fractions
ahead
ofthe
"4C-labeled
SS
DNA
marker;
it
isdoubtful that this
re-flected altered
intracellular
SS
DNA in
the
infected
cultures, but instead probably reflects
the effects of
different methods of DNA
isola-tion
(19).
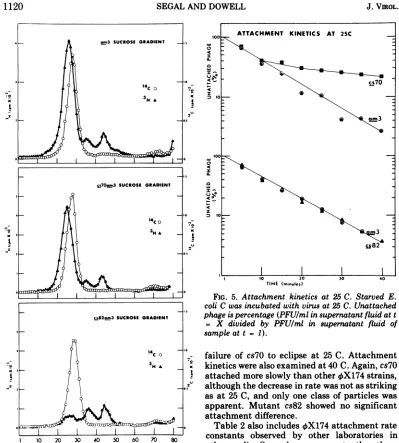
Attachment
kinetics.
The effect of the
cs7O
and cs82 mutations on virus attachmentrate at
25
C
wasexamined
by incubating
virusand
cells under conditions which
permitted phage
attachment
but
notreplication,
and then
by
assaying the
amountof virus
present inthe
supernatant
fluids
ofsamples taken
atvarioustimes
(Fig. 5). Wild
type(not
shown),
am3, andcs82 had
similar linear
kinetics, but
cs7O
exhib-ited
adistinctly biphasic
pattern at 25C. This
suggested that
twoclasses
ofcs7O particles
werepresent
which attached
atdifferent
rates(Table
2).
These two classes may havebeen
present inthe
lysate,
ormay have arisenthrough
activa-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.498.104.423.55.275.2]COLD-SENSITIVE
OX174
MUTANTStion of cs7O
during the experiment (6, 9).
Alternatively, cs7O may have spontaneously
detached
morefrequently
than wild type, am3,
or cs82
(see cs7O
eclipse
kinetics at 40 C, in Fig.
0 40 s0 120 160 200 240 TIME (minutes)
FIG. 2. Growthofcs7O and cs82at25C.Starved E.
coli Cwassynchronously infectedwithcs7O
(MOI
=4.2) or cs82 (MOI = 5.3) at 40C, and growth was
initiatedby 1:10,000dilution intoKCbrothat25C.
Mutantcs7Ohadan
eclipse
timeof
38min,a latentperiod of50min,andaburst size
of
264;mutantcs82hadaneclipse timeof65min,a latentperiod of70
min,andaburst size
of
4.4;wild type(notshown)
hadaneclipsetime
of
40min, alatentperiod
of
50min,and a burst size of230. Relative titer is
(infective
centers orintracellularPFUper
ml) dividedby (input
infectiouscenterspermlduringthelatent
period).
1).
Regardless,
the presence of
aclass of
cs7Oparticles with
asignificantly decreased
attach-ment
ratesupported
the hypothesis that the
cs7O mutation affects some
component of the
0X174 coat. However, this attachment defect
was not
sufficient to account for the complete
0
'0
5
'0
10
E
'0
'c
/g cs~~~~~~r82amt3
(40-25)
0 20 40 60 so 100 120
TIME (mI nutes)
FIG. 3. Growthof am3 and cs82am3 after tempera-turedownshift late in infection. E. coli Cwas
synchro-nouslyinfected with am3 (MOI=5)orcs82am3(MOI
= 5) at40C in thepresence of 0.009 M KCN(16).
Growthwasinitiatedby 1:100 dilutioninto KC broth
at 40C. After 20 min of infection, portions were
diluted 1:10into KC broth at25C. Lysis defective am3 derivatives were used to prevent lysis from prematurelyterminating the experiment.
TABML 1. Burst sizesofcold-sensitive mutantsa
Burst size0
Virus
40C KCbroth 40CC-medium 25C KC broth 25 CC-medium
Wildtype 78 17(6) 241± 102(6) 235 26(6) 534 ± 63 (4)
cs7O 160 34(6) 234 (1) 322±79(4) N.D.c
cs82 103 ± 16(6) 192 ± 63 (6) 2.9 ± 1.2 (5) 15.3 ± 2.2 (3)
am3 1045 ± 45(2) 415 44(3) 347±21(3) 176 ±+-63 (3)
cs70am3 1940(1) 2647 400(3) 401 114 (2) 400 (1) cs82am3 862±87(2) 611 ± 112(3) 3.5 ±0.5(3) 9.0 ± 1.2 (3)
aStarvedE. coli C cultureswere infectedatMOI about 5, and growth initiatedby 1:10,000 dilution into
appropriategrowthconditions.
bMean ± standarddeviation,
figure
inparenthesis isthe number of separate determinations.cND, Not done.
VOL. 14, 1974
1119
EFFECT OF SHIFT FROM 40C TO 25C _
ON THE GROWTH OF am3 ANDcs82pm3
am3
-(NO
SHIFT)-2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.498.44.442.48.500.2] [image:5.498.44.447.516.621.2]FRACTON NUMBER
FIG. 4. Analysis ofintracellular DNA synthesized
at 25C. Starved E. coli HF4704 was treated with
mitomycin C (50ugper10K cellsperml) for15minat 40C, and 25-mi portions were infected with am3
(MOI = 4.7), and cs70am3 (MOI = 5) or cs82am3
(MOI = 4.8). Growth wasinitiated bysuspensionin
C-medium (containing20 ugofadenosineperml)at 25C. After60min [9H]thymidine (specific activity 50
to60Ci/mmol)wasaddedtoalevelof10 MCi/ml,and
at 240 min infected cells were subjected to DNA extraction andgradient analysis. Counts have been corrected for isotope spillandfor background. RF I sedimented withapeak aboutfraction35, and RF II aboutfraction44.
[image:6.498.59.458.51.494.2]TIME (minutes)
FIG. 5. Attachment kinetics at 25 C. Starved E.
coli Cwasincubated with virus at 25 C. Unattached
phageis percentage(PFU/mlinsupernatant fluidat t
= X divided by PFU/ml in supernatant fluid of
sampleat t= 1).
failure of
cs7O
toeclipse
at 25C. Attachment
kinetics
werealso examined
at40C.
Again, cs70
attached
moreslowly
than other
4X174
strains,
although
the decrease in
rate was not asstriking
as at 25
C, and
only
oneclass of
particles
wasapparent.
Mutant
cs82showed
nosignificant
attachment difference.
Table
2also
includes
OX174
attachment
rateconstants
observed
by
otherlaboratories
inother
media. Our values
weregreaterthan those
literature
values,
but this
mayhave
been due
toincreased
adsorption
that
occurswhen
cells
areaerated (1), or when temperature is increased
(20,
37).
Alternatively, the increased
attach-ment rates may
reflect
increased
ability
ofstarved cells
toadsorb
4X174.
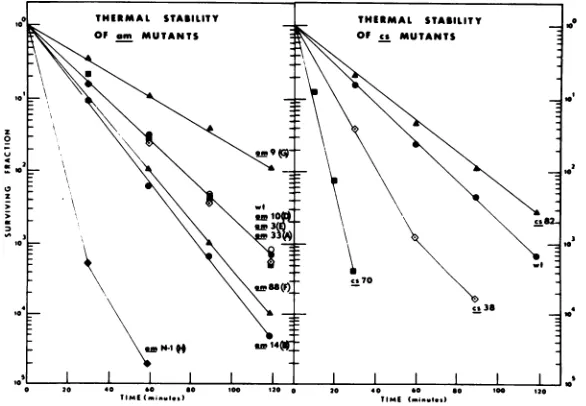
Thermal
stability.
Heat inactivationstudies
were
carried
out oncs7O and
cs82 todetermine
whether either
cold-sensitive
mutationaffected
virus
particle
stability.
Various4X174
mutants,whose
particle stabilities (except
for am88and
amN-1)
havebeen
previously examined
(35;C.
A.
Hutchison, Ph. D.
thesis;
D.Tseui,
Ph. D.thesis, University
ofCalifornia, Davis, 1969),
I2
x
LI
11X
1-1.9
I.,X
I
S)
LX
z
on November 10, 2019 by guest
http://jvi.asm.org/
COLD-SENSITIVE ,X174MUTANTS
were
incubated in saturated sodium tetraborate
at 56.5
C. The
data
in
Fig. 6
showed
that
only
those amber mutants with altered coat proteins
(suppressed, but still missense mutations)
ex-hibited altered
inactivation kinetics, thus
dem-TABLE 2. Attachment rate constants of cs mutants
VirusTemp (C) Attachmentrate
con-Virus Temp (C)
stants
(k)a
Wildtype 25 3.02 +0.20(2) am3 25 3.51 + 0.14 (4)
cs82 25 3.76+0.40(2)
cs7O 25 3.45 + 0.55 (2)
0.80+0.17(2)b
Wildtype 40 14.8
am3 40 15.7
cs82 40 14.3
cs7O 40 12.9
Wildtype 37 7.8 1.8(15)c;6A01;8.2e am3 37 8.6 + 0.59 (4)e
aExperimentswereperformed asdescribedin
Ma-terials and Methods, and k values (1) expressed as
milliliters per minute x 109. Both cs7O and cs82 determinations were from a tube containing am3 as well.Multiplevaluesrepresentthe results from more than one experiment.
bThe second set of values represent the biphasic
component ofthe curves; the first set derives from the first 10 min of theexperiments, the second set from the final 20 min.
cInKC broth (3).
dIn 0.1 M
CaCl2
at36C (20).eInstarvationbuffer(28).
THERMAL STABILITY OF am MUTANTS
0
* 2
Z
onstrating the ability of heat inactivation
stud-ies
todistinguish coat from noncoat
4X174
amber mutants. The greatly increased heat
sensitivity of
cs70 was therefore
considered
evidence of an altered cs7O coat protein;
cs82showed decreased heat sensitivity, but the
dif-ference
wasnot
sufficiently compelling to claim
that cs82 also is
acoat
protein
mutant.Although other
laboratories have used
parti-cle stability to define coat protein mutations,
the
useof different temperatures and
incuba-tion media
makes
the
comparison of heat
stabil-ities
difficult. To determine whether the
differ-ences observed for cs7O and cs82 were
signifi-cant
according to criteria adopted in other
studies, we chose experimental times when the
extent of wild-type inactivation was equivalent
to
that
observed by others. For example,
Sin-shiemer (35)
and Hutchison
(Ph.
D.
thesis)
observed 10-3 wild-type survival after 30 min in
0.025 M
Tris
buffer at 60
C, and the data in Fig.
6 showed about the same survival at 120
min.
Therefore, by applying the same criterion to the
t
=120
values in Fig. 6 as used
by
Hutchison
and
Sinsheimer
at t
=30 to
define a significant
change,
we canconclude from the
resulting
comparison (Table 3) that
cs7Oand
perhaps
cs82
had significantly altered particle
stabili-ties.
Similarly, by applying
the
criteria of Baker
and Tessman
(2)
tothe
t = 30value
inFig.
3(which gave the
wild-type inactivation
equiva-lent to
21min
at51.5
C in 0.1 M phosphate
buffer),
it was found that cs7O
again exhibited
what other investigators would have considered
jo
0 20 40 *0 00 00 t20 0 20 40 *0 0 100 220
-TIME
6Tmsbtoaa cm n a5 CnatroeTIME2.lutiona sd
FiG.
6. Thermalstabilitiesof
amandcsmutants at56.5C in saturated sodium tetraborate solution.VOL.
14,
19741121
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.498.45.234.142.300.2] [image:7.498.100.393.432.635.2]TABLE 3. Thermalstabilitiesof4X174mutants
Inactivation Survivalratio"
Mutant Cistron (mutant/wild
ratea
(mutant/wild type)type)
t=30 t=120am33 A 1.07 0.89 0.63
am14 B 1.47* 0.49* 0.043*
amlO D 1.03 1.03 0.89
am3 E 1.04 1.00 0.85
am88 F 1.30* 0.56 0.119*
am9 G 0.62* 2.00* 15.8*
am23 H 1.33* 0.59 0.128*
amN-1 H 4.68* 0.0009* < 0.0009*
cs7O ? 4.50* 0.0024* < 0.0024* 4.98* 0.0006* <0.0006*
cs70am3 ?, E 3.12* 0.016* <0.0005*
cs82 ? 0.78 1.50 4.88*
0.81 1.33 4.26* cs82am3 ?,E 0.78 1.42 4.38*
aCalculated asdescribed by Sinsheimer(35);
val-ues with asterisk (*) are considered significantly
different, where significance is a value <0.77 or
> 1.25 (35).
bCalculated as described by Tessman and Tessman (39) and Hutchison (Ph.D. thesis). Values with as-terisk(*) areconsideredsignificantlydifferent; signif-icanceat t= 30 isa valueless than 0.5 or greater than 1.5 (39), and at t = 120 is avalue less than 0.2 or greater than 4.0(Hutchison, Ph.D. thesis).
to
be an
altered
virus coat.
Serology
and
host
range. The effects of
the
cs7O and
cs82 mutations on other coat
protein-determined
4X174
functions
werealso
exam-ined.
4X174-neutralizing antibody
reacts
with
the virus coat
(35;
C.
A.
Hutchison, Ph.D.
thesis),
sothat
coatprotein
mutantsmay
well
exhibit altered
antiserum inactivation kinetics.
Furthermore,
4X174
host range mutants may
also show
altered
antiserum inactivation
(6,
7,
9).
However, examination of the responses
of
cs7O and
cs82 toinactivating antiserum
showed
nosignificant
differences from
wild-type
inactivation, with k values of
2.01x
103to
2.33
x
10'/min
for
cs7O,
cs82, am3,
and
wild-type.
With
regard
tohost
range, many -E. coli
strains
wereexamined
including
somesensitive
only
to/X174
host
range mutants, but the only
strains
showing any differences among
wild-type, cs7O, and
cs82 werehost cells
containing
an F+factor. The
data
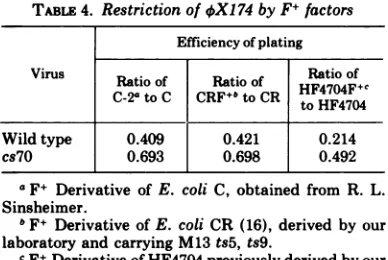
inTable 4showed that in
these
strains, cs7O plated relatively better than
wild
type
(which
wassimilar tocs82),
although
all were
restricted
by the F+ factor. The degree
of
restriction
wasslight, but similar to a
previ-ous
observation
of4X174
restriction
by
F+factors
(23).
DISCUSSION
Conditional
lethal
mutations,
apowerful
ge-netic
technique
developed
by
Campbell (11)
for
phage A and
by
Epstein et al. (17) for phage
T4,
can
define
moregenes than mutations
affecting
specific
functions.
Considerable progress
inun-derstanding
4X174
replication has
been made
with such
mutants (4, 5, 35,
C. A.
Hutchison,
Ph. D.
thesis), but most
investigators have
concentrated on mutants of the
temperature-sensitive
and nonsense types. Our
laboratory
has
sought additional
types of4X174
condi-tional lethal mutants, and we have isolated
cold-sensitive mutants
(15)
aswell as
strains
capable
of
growth
athigh
temperatures
(30)
and in
strains
normally
excluding
OX174
(7).
The present
paper
extends the initial
descrip-tion
(15) of the cold-sensitive
4X174
mutantscs7O
and cs82. We show that the
cs7Omutant
was blocked in
eclipse at 25 C (Fig. 1) but
wascapable of wild-type
replication
atthis
temper-ature if
allowed
prior
eclipse
at 40C
(Fig. 2).
The cs82
mutantexhibited
wild-type
eclipse
kinetics at both temperatures, but
at25
C
wasseverely
impaired
inSS DNA
synthesis (Fig. 4),
with
concomitant
delayed initiation of
intracel-lular
phage production, decreased rate of
prog-eny
synthesis, and reduced burst size. The
rapid
cessation of
intracellular cs82am3 production
following
temperature downshift late
ininfec-tion
(Fig.
3) shows that the
cs82gene function
was
continually
required during replication,
and that the
decreased SS
DNA
synthesis
observed
at 25C was not due
simply
to afailure
to
initiate SS DNA
synthesis.
At
permissive temperature (40
C)
both
cs7O
and
cs82grew
in away
similar to wildtype.
Wewere
intrigued
by the consistently higher burst
sizes of
cs7O relative
towild type
(Table
1),
[image:8.498.60.253.79.268.2]even
though
the
rateand
quantity
ofcs70am3
DNA
synthesis
was the same as am3(Fig.
7).
This
implied
that
the cs7O gene
product,
inTABLE 4. Restrictionof
4X174
byF+ factorsEfficiencyofplating Virus Ratioof Ratio of Ratioof
C-2ato C CRF+"toCR
HF4704F+c
tWildtype 0.409 0.421 0.214
cs7O 0.693 0.698 0.492
aF+ Derivative of E. coli C, obtained from R. L.
Sinsheimer.
bF+ DerivativeofE. coliCR (16), derived by our laboratory and carrying M13 ts5, ts9.
cF+Derivative of HF4704
previously
derivedby ourlaboratory (10).
1122
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.498.264.458.511.641.2]COLD-SENSITIVE kX174MUTANTS
rl-I0
x
E
0
0
t!
z
c4
4
0
u
z
0 20 40 60 80
TIME (minutes)
FIG. 7. DNA synthesis at 40C. Starved E. coli HF4704 was treated withmitomycin C asdescribed
(27) and infected with am3 (MOI = 4.4), cs70km3
(MOI=4.4)orcs82am3(MOI=4.4)at40C. Growth
wasinitiatedby suspension in C-medium containing2
;&gof nonradioactive thymineper ml and 10
,gCi
of[3H]thymidine
perml(specific activity 23Ci/mmol).Samples (0.5 ml each)wereassayed for[3HJuptakeas
described(27). Theburstsizeswere57for am3,88for cs82am3, and 245forcs70am3.
addition to controlling eclipse, may also have
had a regulatory role in 4X174 replication.
Further support for this hypothesis was that
both cs7O and cs82 could replicate at 44 C, a
temperature at which wild-type 4X174 would
notplate. As such,these mutantsmaybe useful
in studies of viral replication in host strains
temperature-sensitive for DNAsynthesis (7, 9,
30; C. E.Dowell, unpublished data).
The 4X174 mutations have beenassignedto
specific genes by both complementation and
recombination analysis, although
complemen-tation analysis is the more fruitful approach
because of low recombination frequencies (29,
40; C. A. Hutchison, Ph. D. thesis) and
site-specific effects (4) in 4X174. It was our
inten-tion to use complementation testing to
deter-mine whether cs7O orcs82 defined new 4tX174
functions.
Complementation
mapping involvesmixed-infection
under restrictive conditionswith the degree
of intracellularcomplementa-tion
determined
byplating
atrestrictivecondi-tions. It
canbe
seen thatthe very nature of the cs7O defect (inability to eclipse at 25 C)pre-cluded
complementation
testing
ofthis
mutant,since
atthe
restrictive temperature thecs7O
genome cannot even enter the cell to participate
in
complementation. Since
cs7O was thus notamenable
tocomplementation
analysis,non-genetic
methods
ofassigning cs7O to a geneticfunction
wereinvestigated.
Virus attachment is
obviously
a propertyof
the virus protein
coat,and
OX174
mutationsaffecting
attachment
arefound in
coatprotein
genes
(28, 36,
38). The aberrant attachment
kinetics of
cs7O atboth
25and
40C
weretherefore considered
asevidence of
analtered
cs70
coat protein.The
mostcommonly used
nongenetic criterion
toidentify
coatprotein
mutants is
particle thermal
stability. Intact
phage
arediluted into
achemically defined
solution maintained
at constant temperature(about
60C), and the
rateof inactivation
deter-mined
by
plating survivors
atspecific intervals.
This
treatmenthas
little effect
onDNA
primary
structure or
infectivity
(6, 18); thus, the
inac-tivating
eventis the
breaking
of
the
phage
coat.Mutants with heat
stability
different from
wild-type
arethought
tohave
analtered
particle
structure,
and hence
amutated
coatprotein
(33, 35, 39, C. A.
Hutchison,
Ph. D.
thesis).
The
significantly decreased heat
stability
of
cs7O
insaturated borate
solution
at 56.5C
wasagain
interpreted
asevidence of
acoatprotein
altera-tion. A
previous heat inactivation of cs7O
(15)
showed increased heat
stability,
but that
exper-iment was
performed
in adifferent
buffer and
ata
lower
temperature.Benbow
etal.
(4) have
placed
cs7O in
geneH
on
the
basis
ofthree- and four-factor
crosses,and
someof
the cs7O
properties
supportthis
hypothesis.
For
example,
the
only
other
mutantaffecting eclipse
isthe
geneH S13
mutanttl069
(14),
and the
only
other
mutantexhibiting
such
extreme
heat sensitivity
is the
geneH
mutantamN-1.
Also, cs7O
seems toaffect
the efficiency
of
maturing
progenyDNA into infectious
parti-cles
(contrast the increased
cs7O burst sizes inTable
1 withequal cs7O
DNAsynthesis
inFig.
7), and
the three gene H mutantsamN-1,
am80,
and
ts4also
affecttheability
ofSS
DNAtobe
matured
into infectious virusparticles
(16, 21,
33).
With
regard
tocs82and
conventionalgenetic
approaches,
preliminary
complementation
testsPHAGE DNA SYNTHESIS AT 40C
am3 e
cs7Oam3 U cs82am3 A
I
I
VOL.14, 19'74
1123
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.498.45.241.57.366.2]SEGAL DOWELL
TABLE 5. Effect ofcsmutationsonco-infecringam3
replication at25C
Virus MOT
Total
burst' am(%) cs(%)am3 4.9 193 100
cs70am3 5.2 386 100
cs82am3 5.6 1.4 100
cs70am3+am3 4.3 294 545 46b
4.0
cs82am3+am3 4.0 38 38C 14c
3.8
aE. coli C was infected at 40 C with the desired
mutants inthe presence of 0.009 M KCN(16). After 15 min at 40C with mildaeration, growthwasinitiated by 1:10,000 dilution into KC broth at 25 C. Samples were assayed for intracellular phage at t = 180, and total burst size determined by plating on E. coli HF4714 at 37 C.
bam3 contribution determined by plating at 25 C,
cs70am3 as the difference between 25 C and 37 C
values. The calculation considered the efficiency of plating of 0.7 for am3 at 25 C relative to 40 C.
ccs82am3 contribution determined at 44 C (since
am3 will not plate at this temperature), and am3 contribution as the difference between 37 C and 44 C values. Similar percentages were determined from 37C versus 25 C plating, as in the prior footnote.
suggested that the
cs82 mutationsomehow
interfered
withcoinfecting
virusreplication.
This
occurred
evenwhen
the
coinfecting virus
was am3
(Table 5). Although
am3gave
aburstof
193 at 25C, the
presenceof
co-infecting
cs82am3
reduced the burst
size to38,
mostof
which
(86%)
wasam3 progeny. This
trans-domi-nance
effect
suggests that the cs82 defect at25
C is not due
tothe
absenceof
anecessary
gene
product, but
isdue instead
tothe presenceof anaberrant gene
product capable
ofinhibit-ing expression
of
aco-infecting
wild-type
ge-nome. The
possibility
that the cs82 geneprod-uct at
25
C
is aprotein
present in aninactive
configuration
issupported by
theprevious
downshift
experiment (Fig. 3),
which showedimmediate cessation of
intracellulargrowth
fol-lowing
temperaturechange.
These areprecisely
the effects
predicted
fromchanges
inprotein
configuration at different temperatures.
ACKNOWLEDGMENTS
This investigation was supported by National Science Foundation grant 18639.
We wish to thankMichael Bowes, Donald Bone, and David Yeltonfor fruitful discussions, and also the Department of Microbiology, University of Massachusetts, where most of thisinvestigation wasperformed.
LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages. Interscience Pub-lishers,Inc., New York.
2. Baker, R., and L. Tessman. 1968. Heat stability of mutants in genes II,Illa and VI of phage S13. Virology 35:179-181.
3. Bayer, M. E., and T. W.Starkey.1972.Theadsorptionof bacteriophage OX174and its interaction with Esche-richia coli; a kinetic and morphological study. Virology 49:236-256.
4. Benbow, R. M., C. A. Hutchinson III, J. D.Fabricant, and R. L. Sinsheimer. 1971. Genetic map of bacterio-phage 4X174. J. Virol. 7:549-558.
5. Benbow, R. M., R. F. Mayol, J. C. Picchi, and R. L. Sinsheimer. 1972.Direction oftranslationand size of bacteriophage OX174cistrons. J. Virol.10:99-114. 6. Bleichrodt, J. F., J. Blok, and E. R. Berends-Van
Abkoude.1968.Thermal inactivation ofbacteriophage *X174andtwoof itsmutants.
Virology
36:343-355. 7. Bone, D. B., and C. E. Dowell. 1973. A mutant ofbacteriophage OX174which infects E. coli K12 strains. I. Isolation and partial characterization of *XtB. Virology 52:319-329.
8. Borrias,W.E.,J. H.Van DePol, C.VanDeVate,andG. A. Van Arkel. 1969. Complementation experiments between conditional lethal mutants ofbacteriophage *X174. Mol. Gen. Genet. 105:152-163.
9. Bowes, J. M., and C. E. Dowell. 1974. Purification and some properties of bacteriophage ST-1. J. Virol. 13:53-61.
10. Brown, L. R., and C. E. Dowell. 1968. Replicationof coliphageM-13. II. Intracellulardeoxyribonucleicacid forms associated with M-13 infection ofmitomycin C-treated cells. J. Virol. 2:1296-1307.
11. Campbell,A. 1961.Sensitive mutants ofbacteriophageA. Virology14:22-32.
12. Denhardt, D. T. 1969. Formation ofribosylthymine in Escherichiacoli.J. Biol.Chem.244:2710-2715. 13. Denhardt,D. T.,and R.L.Sinsheimer.1965.Theprocess
of infection with bacteriophage OX174. III. Phage maturation and lysis aftersynchronized infection. J. Mol.Biol. 12:641-646.
14. Doniger, J., and I. Tessman. 1969. AnS13capsid mutant that makes no replicative-form DNA. Virology 39:389-394.
15. Dowell, C.E. 1967. Cold-sensitivemutantsof bacterio-phage *X174. L. A mutant blocked in the eclipse function at low temperature. Proc. Nat. Acad. Sci. U.S.A.58:958-961.
16. Dowell,C.E.,andR. L. Sinsheimer. 1966. The process of infection with bacteriophage *X174. IX. Studies on the physiology ofthree OX174 temperature-sensitive mutants.J. Mol. Biol. 16:374-386.
17. Epstein, R. H., A. Bolle, C. M. Steinberg, E. Kellen-berger, E. BoyDe LaTour, R. Chevalley, R.S. Edgar, M. Susman, G.H.Denhardt, andA.Lielausis. 1963. Physiological studies ofconditionallethalmutantsof bacteriophage T4D. Cold Spring HarborSymp. Quant. Biol.28:375-394.
18. Fiers, W. F., and R. L. Sinsheimer. 1962. The structure of the DNA of bacteriophage OX174.IL.Thermal inacti-vation.J. Mol. Biol. 5:420-423.
19. Forsheit, A. B., and D.S.Ray.1970.Conformations of the single-strandedDNA ofbacteriophageM13. Proc. Nat. Acad. Sci. U.S.A. 67:1534-1541.
20. Fujimura,R.,and P.Kaesberg.1962.Theadsorptionof bacteriophageOX174toits host.Biophys.J. 2:433-449. 21. Funk, F., and R. L. Sinsheimer, 1970. The process of infection withbacteriophageOX174. XXXV. Cistron VIII. J. Virol. 6:12-19.
22. Greenlee,L.L., and R. L. Sinsheimer. 1968. The process ofinfection withbacteriophageOX174.VII.Effects of specific metabolic interruptions. J. Mol. Biol. 32:303-320.
23. Groman,N.B. 1969. Restriction ofbacteriophageOX174
124
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.498.63.258.71.197.2]COLD-SENSITIVE 0X174 MUTANTS
by F+ factor. Biochem. Biophys. Res. Commun. 37:691-696.
24. Guthrie, G. D., and R L Sinsheimer.1963.Observations
on the infection of bacterial protoplasts with the
deoxyribonucleic acid of bacteriophage OX174. Bio-chim. Biophys. Acta 72:290-297.
25. Hutchison, C. A., M. H. Edgell, and R. L. Sinsheimer. 1967. The process of infection with bacteriophage
0X174. XII. Phenotypic mixing between electropho-reticmutantsofOX174.J. Mol. Biol. 23:553-575. 26. Iwaya, M., and D. T. Denhardt. 1971. The mechanism of
replication of 0174 single-stranded DNA. H. The role of viralproteins. J. Mol. Biol. 57:159-175.
27. Lindqvist, B. H., and R. L. Sinsheimer. 1967. The proc-essof infection with bacteriophage #X174. XV.
Bac-teriaphage DNA synthesis in abortive infections witha
set of conditonal lethalmutants. J. Mol. Biol. 30:69-80.
28. Newbold, J. E., and R. L. Sinsheimer. 1970. Theprocess
of infection with bacteriophage sX174. XXXIV. Ki-netics of the attachment and eclipsestepsof the infec-tion. J.Virol. 5:427-431.
29. Pfeifer, D. 1961. Genetishe untersuchungenam bakterio-phagenOX174I.Aufbau eines selektivensystemsund nachweis genetischer rekombination. Z. Vererbung-slehre 92:317-329.
30. Primrose, S. B., W. M. Landers, and C. E. Dowell. 1971. Replication of bacteriophageOX174ina temperature-sensitive deoxyribonucleic acid host cell. J. Virol. 8:594-596.
31. Rolfe, U., and R. L. Sinsheimer. 1965. Antigens of
bacteriophage#X174.J. Immunol. 94:18-21. 32. Sedat, J. W., and R. L. Sinsheimer. 1970. The invivoOX
mRNA. Cold Spring Harbor Symp. Quant. Biol. 35:163-170.
33. Siegel, J.E. D., and M. Hayashi. 1969. 0X174
bacterio-phage structural mutants which affect deoxyribonu-cleic acid synthesis. J. Virol. 4:400-407.
34. Siegel, J. E. D., M. N. Hayashi, and M. Hayashi. 1968.
*X174coatproteinmutantsaffectingDNAsynthesis.
Biochem. Biophys. Res. Commun. 31:774-778. 35. Sinsheimer, R. L. 1968. BacteriophageOX174and related
viruses. Progr.Nucl. AcidRes. Mol. Biol. 8:115-169. 36. Sinsheimer, R. L., C. A. Hutchison m, and B. H.
Lindqvist. 1967. Bacteriophage 0X174: viral functions,
p. 175.InJ. S. Colter and W. Paranchych (ed.), The molecular biology of viruses. Academic Press Inc., New York.
37. Stent, G.S.1963. Molecularbiology of bacterial viruses. W. H. Freeman Co., San Francisco.
38. Tessman, E.S. 1965. Complementationgroupsinphage S13.Virology25:303-321.
39.Tessman,I., and E. S. Tessman. 1966. Functional units of phage S13: identification oftwogenesthat determine thestructureofthephagecoat.Proc. Nat.Acad. Sci. U.S.A. 55:1459-1462.
40. Weisbeek, P. L., J. H. Van De Pol, and G. A. Van Arkel. 1973.Mapping of hostrangemutantsofbacteriophage 0X174.Virology52:408-416.
41. Zuccarelli, A. J., R. M. Benbow, and R. L. Sinsheimer. 1972.DeletionmutantsofbacteriophageOX174.Proc. Nat. Acad. Sci. U.S.A. 69:1905-1910.
VOL. 14, 1974

![FIG. 7.Samplesdescribed;&gcs82am3,HF4704was(MOI[3H]thymidine(27) DNA synthesis at 40 C](https://thumb-us.123doks.com/thumbv2/123dok_us/1579142.110526/9.498.45.241.57.366/fig-samplesdescribed-gcs-hf-moi-thymidine-dna-synthesis.webp)