0022-538X/78/0025-0553$02.00/0
Copyright ©1978 AmericanSocietyforMicrobiology Printed inU.S.A.
Impaired
Processing of
Precursor
Polypeptides
of
Temperature-Sensitive
Mutants of Rauscher Murine
Leukemia
Virus
W. J. M. VAN DEVEN,* D.VAN ZAANE,t C. ONNEKINK,AND H. P. J. BLOEMERS Departmentof Biochemistry, University of
Nijmegen,
Nijmegen,
The NetherlandsReceived forpublication 24 August 1977
The synthesis and processing of virus-specific precursor polypeptides in
NIH/3T3cellsinfectedatthepermissivetemperature(31°C) with
temperature-sensitive (ts) mutantsof Rauscher murine leukemia viruswas studied in
pulse-chase experiments at the permissive and nonpermissive (39°C) temperatures.
The newly synthesized virus-specific polypeptides were analyzed by sodium
dodecyl sulfate
polyacrylamide
gelelectrophoresis
after immunoprecipitationwith polyvalent and monospecific antisera against Rauscher murine leukemia
virus proteins. In cells infected with ts mutants defective in early replication
steps(the
early
mutantstsl7andts29), andtsmutantsdefective inpostintegrationsteps
(the
latemutants ts25 and ts26), theprocessing
of theprimary
gag geneproduct was
impaired
at thenonpermissive
temperature. gag-pr75 ofall fourmutants was converted into gag-pr65; however, gag-pr65 accumulated at the
nonpermissive
temperature, and the main internal virionpolypeptide
p30wasnot formed.
Therefore,
theproteolytic cleavage
is blockedbeyond
gag-pr65.Concomitantly,
theformation of the envgene-related polypeptide p12(E)
of allfour mutantswasblockedattherestrictivetemperature.In contrast, cells infected with the latemutantts28,which
produced
noninfectiousvirionsat39°C,
showedanormalturnoverof the gagandenvprecursor
polypeptides.
Instudiesonthemolecularbiology of type C
RNA tumor viruses, the isolated conditional
lethalmutants of avian (6, 9, 22, 26) and
mam-malian (13, 15, 17,25) RNA tumor viruses
pro-videuseful markers forthe analysisof theviral
genome.Temperature-sensitive (ts) mutants of
murine
leukemia
virus (MuLV) have beende-rived from the Kirsten (17), Rauscher (15), and
Moloney (25) strains. The mutants of the
Rauscher strain (R-MuLV) have been divided
intothreedifferent classes baseduponthestage
at which virus
replication
is impaired at therestrictivetemperature (15). Through the
cour-tesy of J. R.
Stephenson,
we have obtainedrepresentatives
of the tsmutants of class I (at39°C, defective in
replication
functions beforethe
synthesis
of viral group-specific antigens),class II(at
39°C,
defective in functionsbetweenthesynthesisofviralgroup-specificantigens and
production ofvirions), and class III
(noninfec-tious virions
produced
at39°C).
The virus-specific structural polypeptides of
the RNA tumor viruses are formed through
proteolytic cleavage of precursor polypeptides
tPresent address:DepartmentofVirology,Central Veter-inaryInstitute,Lelystad, The Netherlands.
(for a review see 14). Thus, in the case of
R-MuLV, the
primary
gag geneproduct,
gag-pr75,is converted into gag-pr65, which is
split
intocertain
intermediary
products (with
molecularweights
between40,000 and50,000)
and,
subse-quently,
intothepolypeptides
of the viralcore,p15, p12,
p30,
and plO (2).Likewise,
gp69/71and
p15(E),
thevirus-specific polypeptides
lo-cated in the viral
envelope
as well as in theplasma membrane of infected
cells,
arederivedfrom the
primary
env geneproduct env-pr82.
Finally,
p12(E),
found in the viralenvelope,
ispresumably
derived fromp15(E) (12,
24; D.vanZaane, thesis, University of Nijmegen, 1977).
Using
gel
filtration columnsthatdidnotresolvethe two gag precursor
polypeptides
gag-pr75and
gag-pr65,
Stephenson et al. (18) showedthattwo ts mutants of
R-MuLV,
ts25 andts26,
possessed temperature-sensitive defects in the
proteolytic cleavageof thegaggene-related
pre-cursorpolypeptides. These studieswere under-takentolocalize the defectsmorepreciselyand
toinvestigatewhether the other thermosensitive
replication mutants of R-MuLV also possess
thermosensitive defects in gene
expression.
Therefore, the gene expressions of two
early
(ts17 and
ts29)
and one late (ts28) ts mutants553
on November 10, 2019 by guest
http://jvi.asm.org/
(WT) R-MuLV.
MATERIALS AND METHODS Materials.i,-[n'S]methionine (specific activity,380 Ci/mmol) anda'4C-labeled amino acid mixture
(spe-cificactivity,54mCi/matom)wereobtained fromthe Radiochemical Centre, Amersham, England. Anti-gp69/71 and anti-p15sera were the generous gift of M. Strand and J. T. August. Characteristics of these antisera weredescribed by Strand and August (20).
Anti-R-MuLV serum, anti-p30 serum, anti-p15(E),p12(E) serum, andanantiserum raisedagainst bovine serum albumin (anti-BSA serum) were pre-pared and describedbyvanZaaneetal.(24).
Cells and viruses. NIH/3T3 cells (21) were cul-tured as monolayers in Dulbecco's modification of Eagle mediumsupplementedwith 10%fetal calfserum (Grand IslandBiologicalCo.Bio-CultLtd.).
The viruses used included a clonal isolate of WT R-MuLV (WT-248) and five R-MuLV ts mutants, tsl7, ts25, ts26, ts28, and ts29 (15). NIH/3T3 cells were infected with virusasdescribed byStephenson
and Aaronson (15). Infection with ts R-MuLV was performed at31°C,and that with WT R-MuLV was
performedat37°C.
JLS-V9 cells, derived from bone marrow cells of BALB/c mice (4) infected with and producing R-MuLV (V9-R-MuLV), were cultured as monolayers
inEagle minimal essential mediumsupplementedwith 10%calfserum.V9-R-MuLV labeled with
L-[nS]me-thionine was isolated as described byDuesberg and Robinson (5) andwasusedas areference.
Labeling of cells andpreparation of lysates. Subconfluent monolayers of cells growing in plastic
tissue culture flasks (Falcon Plastics, 25 cm2) were starved for 10min inlabelingmedium (Hanksbasic salt solutionsupplementedwith 10%dialyzedcalf se-rumand amino acids except for the radioactive ones). Then, the mediumwasreplaced by labeling medium with aradioactive amino acid(s), and the cellswere
incubated for30min. Afterbeing pulse-labeled, the cells werewashed once, and4 ml of normal culture medium was added; thereafter, the cellseitherwere
lysedimmediatelyorradioactivitywaschased for
var-ious times. Insomeexperimentsthetemperaturewas
shiftedduringthe chaseperiod.Whenindicated,the temperatureshift from 39 to31°Cwasperformed2h afterthebeginningofthe chaseperiod;the tempera-tureshiftfrom31 to39°Cwasperformed immediately
afterpulse-labelingof the cells.
Afterpulsesaswellasafterchases,thecellswere lysed by the addition to the cultures (i.e., cellsand medium)of1ml ofabuffercontaining50mMsodium phosphate (pH 7.2),4.5%sodiumchloride,5.0%Triton X-100, 2.5% deoxycholate, and0.5% sodium dodecyl sulfate (fivefold-concentrated immunoprecipitation
buffer [24]).Afterclearing thelysates by
centrifuga-tion at 220,000 x g for 10min in aTi5Orotor ofa
Spinco L-50 centrifuge, the lysates were stored at -800C.
Immunoprecipitation and polyacrylamide gel
electrophoresis. Detection of R-MuLV-specific
duredescribed byvanZaaneetal. (24). Immunopre-cipitates were analyzed by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis according to
Laemmli (10) on 7 to 18% polyacrylamide gradient
slabgels. Radioactivitywasvisualizedbyscintillation
autoradiography(3).
RESULTS
Synthesis and identification of
virus-spe-cific
polypeptides
inNIH/3T3 cells infectedwith
WT R-MuLV(WT-248).
The synthesisofvirus-specific polypeptides in NIH/3T3 cells
infected with WT R-MuLV was examined in
pulse-chase experiments at 37°C (Fig. 1A-G).
Cells
werepulse-labeled
for30minwith a"C-labeled amino acid mixture, and,thereafter,
ra-dioactivitywaschased for0and20h.Theviral
structural polypeptideswereidentified through
immunoprecipitation
analysis with well-defined(see Materials and Methods)
monospecific
anti-sera and a polyvalent antiserum against
R-MuLV polypeptides.
Radioimmunoprecipita-tionanalysis of pulse-labeled
cells
withanti-R-MuLV serum revealed that in addition to the
virus-specific precursor polypeptides gag-pr75,
gag-pr65, and env-pr82
(cf.
references 1, 24),small amounts of two high-molecular-weight
polypeptides
(indicated with0)
wereprecipi-tated
(Fig.
1A).Uponanalysis with monospecificantisera it appeared that both
high-molecular-weight
polypeptides
carried antigenicdetermi-nantsofthegag-relatedstructuralpolypeptides
p30
andp15,
butnotoftheenv-related structuralpolypeptide gp69/71 (not shown). These
high-molecular-weight polypeptides are therefore
probably virus-specific polypeptides, which in
addition to the antigenic determinants of the
gag-precursorcontainother virus-specific
anti-genic determinants (possibly of reverse
tran-scriptase [8]). They might represent
virus-spe-cific precursor polypeptides whose synthesis is
due to an occasional readthrough at the gag geneterminationsite on aviralmRNA (14).
Moreover, afterpulses as well as after chases, someadditionalhigh-molecular-weight polypep-tides (>90,000) were precipitated with the
anti-seraused,
especially
withtheanti-R-MuLVse-rum(Fig.
1A-O)
and theanti-p15(E),p12(E)se-rum(Fig. 1D, indicated with *). Probably, many of these polypeptidesarenot virusspecific and represent cross-reacting host
cell
polypep-tides. However, some of them might be degra-dation products of the high-molecular-weightprecursor polypeptidessynthesizedaccording to a readthrough mechanism. We will not discuss
thesignificanceof these products here, because
on November 10, 2019 by guest
http://jvi.asm.org/
PRECURSOR POLYPEPTIDES OF ts R-MuLV
pzr-82 _
pr-75,_
c3jl69/71
p-65-
-F_ _
W--..
-pr-82
&a- ,am --Pr75
-r
_ 4--o gp69/71_oi__p-.-pr65
__
p)30_
p151EI -.-p122rE pl15[E]
pl5-p12[EI
A B C D E F
Chlase- ' 11hr ) 20 20 20 20 20 c 20
H J K M N 0
0 2 7 20 0 2 7 20
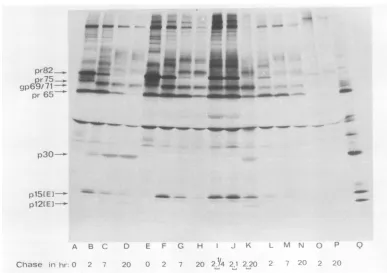
FIG. 1. Normalprocessing of gag and env products of WT R-MuLV (WT-248). Subconfluent monolayers
ofNIH/3T3 cellsinfectedwith WTR-MuL V were pulse-labeled for 30minwith 100
,LCi
ofa'4C-aminoacid mixture (A-G) or with 75liCiofL-[PS]methionine
(H-O),andradioactivity was chased for 0, 2, 7, and 20 h as indicated. The pulse-chase experiments wereperformedat 31°C(H-K), 37°C (A-G), or 39°C(L-O).Afterlysis ofthe cellsasdescribed in the text, virus-specific polypeptides were analyzed by sodium dodecyl
sulfate-polyacrylamide gelelectrophoresisandscintillation autoradiography after immunoprecipitation with: anti-R-MuLVserum(A, B,H-O);anti-gp69/71serum(C);anti-pl5(E),pl2(E) serum (D); anti-p30 serum(E); anti-p15serum(F);oranti-BSAserum(G). Bands marked with * and*are discussed in the text.
theirsynthesis does not seem to be affectedin
anyofthemutants wehaveinvestigated. After
a chase period of20 h,
radioimmunoprecipita-tion analysis of labeled cultures (i.e., cells and
medium)withanti-R-MuLVserumshowedthat
the gag and env precursor polypeptides had
disappeared
and that the viral structuralpoly-peptides gp69/71, p30,
p15(E),
andp12(E)wereformed(Fig.
1B).
These viral structuralpolypep-tideswereidentifiedwithmonospecificantisera
(Fig.
1C-F).
Inacontrolexperiment
noneof thepolypeptides
could beprecipitated
withanti-BSA serum
(Fig.
1G).
It should be noted thatthegag
products
p12andplOarenotrecognized
bytheanti-R-MuLVserumused.
Asacontrolfor the
analysis
of thetsmutants,Fig.
l(H-O)
showsthe results ofapulse-chase
experimentat31°C
(Fig. 1H-K)
and39°C
(Fig.
1L-O) with
NIH/3T3
cells infected with WTR-MuLV
(WT-248).
Cells werepulse-labeled
with L-[35S]methionine for 30 min, and,
there-after,
radioactivity
was chased for 0, 2, 7, and20 h.
Radioimmunoprecipitation analysis
wasperformed
with anti-R-MuLV serum. Itap-pearedthatatbothtemperaturesthe precursor
polypeptide
env-pr82, as well asgag-pr75 andgag-pr65, was cleaved into virus-specific
struc-tural polypeptides (Fig. 1H-O). It should be
noted thatp15does notcontainmethionine (7)
andconsequentlyisnotdetected whencellsare
labeledwithL-[35S]methionine.
Localization
of theblockageinthecleav-ageof the gag
gene-related
precursorpoly-peptides
of ts25 and ts26. Thecleavage
ofthegaggene-related precursor
polypeptides
ofts25 and ts26 was shown to be blocked atthe
nonpermissive temperature (2, 19). To localize
the
blockage
in thecleavage scheme,
thesyn-thesisandprocessingofvirus-specific precursor
polypeptides was examined in NIH/3T3 cells
infectedat31°C withts25andts26.Pulse-chase
experiments were performed at 31 and 39°C,
andthe
virus-specific polypeptides
wereimmu-noprecipitated with anti-R-MuLV serum. The results obtained with ts25 arepresented inFig.
2. At31°C no differences in the synthesisand
processing of the precursorpolypeptidesofts25
and WT-248 could be observed (cf. Fig. 2A-D
and Fig. 1H-K). In accordance with the work
of Stephenson et al. (19), Fig. 2(E-H) shows
that the processing of the gag gene precursor
polypeptideswasimpairedatthe
nonpermissive
VOL. 25,1978 555
I-anit-- -
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.49.444.77.308.2]pt-82_, pr-75
gl.)69/71
-pr 65*
a
a*~~~~b
O. - ama
4o
~~~~~~~.-. w4__~ _
p30
--p15[E I---
-121E2I1 E
A B (. D E F G H
Chasse' iTA("0 20 ( 2 20 2 4
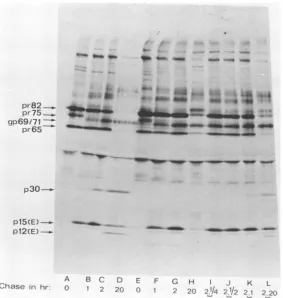
FIG. 2. Thermosensitiveprocessing ofgagproductsofts25. Subconfluent monolayers of NIH/3T3 cells
infectedat31°Cwithts25werepulse-labeled for30min with 75,uCiof L-[35S]methionine, andradioactivity was chasedforvarious times asindicated (A-P). Pulse-chase experiments wereperformed at 31°C(A-D,
O-P)or39°C(E-H).Insome casesthe temperaturewasshifted forachaseat31°C (I-K)or39°C(L-N), as
described in the text. Thus, "chase in h2l/4" indicates a2-h chase at39°C followed by a 0.25-h chaseat
31°C,etc.Virus-specific polypeptideswereanalyzedasdescribed in thelegendtoFig.1.Immunoprecipitation
with: anti-R-MuLVserum(A-N);anti-BSAserum(O-P); reference V9-R-MuLV labeled with
L-[35S]methio-nine(Q).
temperature; gag-pr65 of ts25 is accumulated
and no p30isformed. On the other
hand,
likeWTgag-pr75, thegag-pr75 ofts25isconverted
into gag-pr65. When the temperature was
shiftedto31°C2hafter
pulse-labeling
the cellsat 39°C (Fig.
21-K),
theproteolytic
cleavagewas resumed and p30was produced. When, in
another experiment, the temperature was
shifted from 31°C (during the pulse) to 39°C
(duringthechase), theproteolytic cleavage was blocked (Fig. 2L-N).
The results of parallel experiments with NIH/3T3 cells infected with ts26 (not shown) weresimilartothose observed with ts25; again,
at39°Cgag-pr65wasaccumulated, but the
mu-tationwasslightly leaky:cellsinfected with ts26
producedsmallamountsofp30attherestrictive
temperature(Fig. 3G-H).
A more detailed analysis of the blockage in
the proteolytic cleavage of the gag precursor
polypeptidesofts25andts26 wasobtained after
radioimmunoprecipitation analysis with
mono-specific antisera
(Fig.
3). Cells infectedwith ts25orts26werelabeledfor30minwitha
"'C-amino
acid mixture at 31 or
39°C,
and radioactivitywaschasedfor20h. Figure3showsthatatthe
nonpermissive
temperaturegag-pr65 wasaccu-mulatedincells infectedwith ts25
(Fig. 3M-O)
orts26(Fig. 3G-I);furthermore, the
production
of the
gag-related
structuralpolypeptides
p30
and
p15
was inhibited. In addition to theim-pairedprocessingof the gagprecursor
polypep-tides of ts25and ts26,ablockagein the
produc-tion of an env
gene-related
structuralpoly-peptidewasalsoobservedattherestrictive
tem-perature.
Radioimmunoprecipitation
analysiswith anti-R-MuLVserum (Fig.2H and N; Fig.
3G and M), anti-gp69/71 serum (Fig. 3K and
Q), and
anti-p15(E),p12(E)
serum (Fig. 3J andP) revealed that production of
p12(E)
was in-hibited. Production of the structural envelopepolypeptides gp69/71 and
p15(E),
however,seemedtobe normal (Fig.2F-H andFig. 3G, J,
K, M, P,andQ).Itshould be noted that
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.67.454.76.349.2]PRECURSOR POLYPEPTIDES OF ts R-MuLV
I
gp69/71-
d
pr65 _I1.
a
_.-_-*
.l"i
p15(E) _ _
p15 a
p12(E)
I:
t-O
lb
7-
-_
^o
_im- --- ---P15(FE)
_-
_--A B C D E F G H J K L M N
O p Q
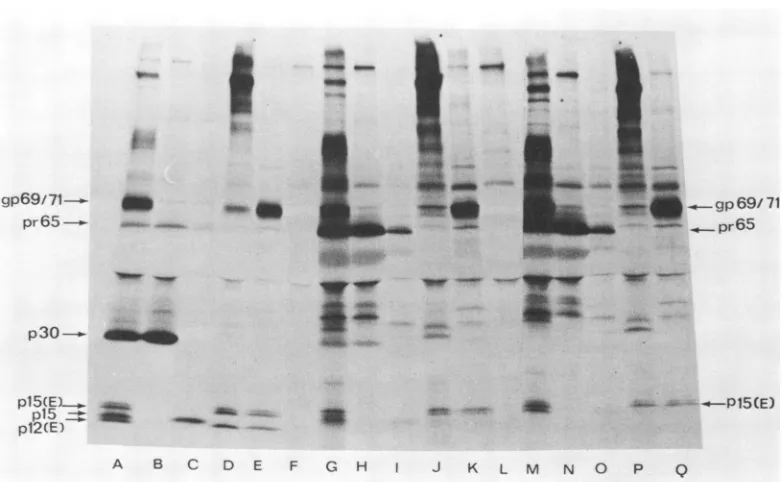
FIG. 3. Leakythernosensitive processing ofgagproducts ofts26; thermosensitive productionof the env
productp12(E)in additiontothethermosensitiveprocessing ofgagproducts ofts25andts26.NIH/3T3 cells
infected at31°Cwith ts26 (A-L) orts25(M-Q) werepulse-labeled for30min with 100,uCi ofa 14C-amino
acidmixture,andradioactivitywaschasedfor20 h.Pulse-chaseexperimentswereperfornedat31°C (A-F)
or39°C(G-Q). Virus-specific polypeptideswereanalyzedasdescribed in thelegendtoFig.1. Immunoprecip-itation with: anti-R-MuLV serum (A, G, M); anti-p30 serum (B, H, N); anti-p15 serum (C, I, 0);
anti-p15(E),pI2(E)serum(D, J, P); anti-gp69/71serum(E, K, Q); anti-BSAserum(F, L).
times pl5(E) seemed to be composed of two components(Fig. 1, 2, and 3). Bothpolypeptides
were immunoprecipitated with anti-R-MuLV serum(Fig. 1, 2, and 3A, G, and M), anti-gp69/71 serum(Fig. 3E, K, and Q), andanti-p15(E),p12(E) serum (Fig. 3D, J, and P). [Incidentally, the
precipitation of p15(E) and p12(E) with anti-gp69/71 serum is presumably due to the pres-enceofacomplex betweengp69/71, p15(E), and
p12(E)asdiscussedbyvanZaaneetal. (24) and
Leamnsonetal. (11).]
Synthesis ofvirus-specific polypeptides
in NIH/3T3 cells infected with tsl7, ts28,
andts29. Twoearlytemperature-sensitive
mu-tants ofR-MuLV, tsl7andts29,wereshownto
bedefective inone or morefunctionsbefore the
synthesisofgroup-specific antigens (15). To
ex-amine the possibility that these mutants also
hadadefect in theproteolytic cleavageoftheir precursor polypeptides, the synthesis of virus-specific polypeptideswasstudiedat31and39°C, after infection at the permissive temperature.
Cells were labeledwith L-[35S]methionine, and
radioimmunoprecipitation was performed with
anti-R-MuLVserum.Theresultsobtained with
ts17 and ts29 are presented in Fig. 4 and 5,
respectively. In bothcases,gag-pr75 was
proc-essedtogag-pr65atthenonpermissive
temper-ature.However,proteolytic cleavagewasblocked
beyond gag-pr65, since no p30 was formed.
When the temperature was shifted from 39 to
31°C, cleavage was resumed, as could be
con-cluded from theproduction of p30. Furthermore, the inhibitionoftheproductionofp12(E)atthe
restrictive temperature, asfound incultures
in-fectedwithts25andts26,wasalsoobserved.On
the otherhand, the latemutant ts28, similarly analyzedinparallel experiments,wasnot defec-tive fortheprocessing of itsprecursor
polypep-tides. The relevant pulse-chase experimentsat 31and39°CareshowninFig. 6.
DISCUSSION
Some temperature-sensitive mutants of
R-MuLVarepartiallycharacterized (15, 16, 18, 19, 23,27). Amongthemarethe mutantsts17, ts25,
ts26, ts28, and ts29, which were used in this
study.
At39°C,someof thesemutantsaredefective in either early replication steps (class I
repre-sentedby tsl7andts29)orpostintegrationsteps (classIIrepresented by ts25andts26, andclass
'71
pr65
VOL. 25,1978 557
E-0 t
ago
ob.ft .
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.500.45.436.69.310.2]."! 8-), _a
p6t75
-!!)69
J71:.- (I5z 0
" .
_NW *N -0
maegm-
a*MOW
ow-.
_soAmmww ---__ _w _4m- op
r)30
;5(L,([- _
;,12(ir ;:
rF L aj :A'\^ I'
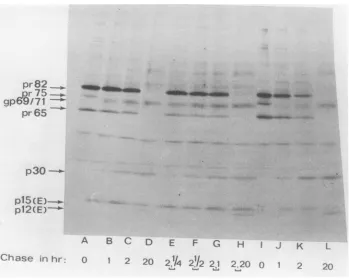
FIG. 4. Thermosensitiveprocessing of gag products of tsl7and concomitantfailure to produce the env productp12(E).NIH/3T3cellsinfectedat31°C with tsl7werepulse-labeledwith75,iCi ofL-[35S]methionine
for 30 min, and radioactivity waschasedfor various times asindicated (A-M). Pulse-chase experiments wereperformed at 39°C (A-D) or31°C (H-M). The temperature was shifted from 39 to 31°C (E-G), as
described in thetext(seealsolegend toFig. 2). Virus-specificpolypeptideswere analyzedasdescribed in thelegendtoFig.1.Immunoprecipitationwith: anti-R-MuLVserum(A-K);anti-BSAserum(L-M);reference
V9-R-MuLV labeled withL-[35S]methionine (N).
IIIrepresented by ts28) (15). ts25andts26 are
postintegration defectivemutantswhose defect
is known to involve the cleavage of the gag
gene-related
precursorpolypeptides
(2, 19).Mu-tantts29 possessesathermolabilereverse
tran-scriptase (23) and ts28 has an unidentified ts
defectat alate step in itsreplication (15, 27).
In this study cells were infected with these
mutants atthepermissive temperature (31°C),
and subsequently thesynthesis and processing
ofvirus-specificprecursorpolypeptideswere
ex-amined at the permissive and nonpermissive
temperatures. At 31°C no differences in the processing of the gag and env gene-related
pre-cursor polypeptides were observed in cells in-fected with either WT R-MuLV or oneof the
ts mutants (Fig. 1, 2, 4, 5, and 6). However, at
39°C,ablockagein theprocessingofagag
gene-related precursor polypeptide of the mutants
tsl7, ts25, ts26, and ts29 was observed (Fig. 2,
3, 4,and 5). When the temperature wasshifted
down to
31°C
afterpulse-labeling
the cells at39°C, processing ofthegag precursor
polypep-tidesto the internal structural virion
polypep-tides p30 (Fig. 2, 3, 4, and 5) and p15 (Fig. 3)
was observed. The impaired processing of the gag precursor polypeptides at the restrictive temperature was localized beyond gag-pr65, since gag-pr75 disappearedbut gag-pr65
accu-mulated andp30wasnotproduced (Fig. 2, 3, 4, and 5). In all analyses, impaired production of
p12and plOescaped detection, since these
virus-specific polypeptideswere notrecognized by the
anti-R-MuLV serum. However, presumably
noneof the gaggene-relatedstructural polypep-tides isproduced at39°Csince all four subgene
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.72.459.69.382.2]PRECURSOR POLYPEPTIDES OF ts R-MuLV
pr82 _ pr75_
gFp69/71 pr65
II 01- i
.*.. lo M a 4*
k._.
~~~~~~~~~~~mub-1
~ ~ ~ ~ ~ ~ ~~omkl 4
wql qwp w w qW
Pl5CE)
--p12(E}
Ch-ase in hr: A0 B1 C2 20D E0 1F G2 20H 21/4 21/2J 21K L
2.20
FIG. 5. Thermosensitiveprocessing of gag products of ts29 and concomitant failure to produce the env gene-related productp12(E). NIH/3T3 cellsinfectedat31°C withts29 werepulse-labeled for30minwith75
yCiof
L-[3S]methionine,
andradioactivity waschasedfor various timesas indicated(A-L). Pulse-chaseexperiments wereperformed at31°C (A-D) or39°C (E-H). The temperature wasshifted from 39to 31°C
(I-L), as described in the text (see also legend to Fig. 2). Virus-specific polypeptides were analyzed as
describedin thelegendtoFig. 1.Immunoprecipitationwith: anti-R-MuLVserum(A-L).
products, p15, p12,p30, and plO,arepresent in
the smallergag precursor
polypeptide
gag-pr65(van Zaane,
thesis).
This conclusion does notexclude the
possibility
thateventually
gag-pr65isdegraded andmay give rise to anincomplete
setofgagsubgene products (cf. Barbacid etal.
[2]). The impaired processingof the gag
precur-sor polypeptide of ts25 and ts26 is consistent
with the observationsofStephensonetal. (19).
Thepresenceof thesamedefectinthemutants
ts17 and ts29 is
quite
unexpected, since bothwerecharacterizedasearlymutants(class I).In
the case of ts29 the early replication defect is associated withathermolabilereverse
transcrip-tase (23).Furtherevidence forasecond
replica-tion defect in ts29 is obtained from an electron
microscopic study by Yegeretal. (27).
Upon ashift to the nonpermissive tempera-ture after infection atthe permissive tempera-ture, these authors found an accumulation of
immature virions at the cell surface. The
im-maturevirionswereinan
early
stage ofbudding,suggestinga
replication
defectat anearly
stageof virion
assembly.
Whereas cultures infectedwith ts17, ts25, ts26, and ts29revealed an
im-paired
processing
of the gag precursorpolypep-tides at the
nonpermissive
temperature,produc-tion of theenv
gene-related polypeptide p12(E)
wasalso inhibited (Fig. 2, 3, 4, and5). It isnot
yet clear whether or not there is a relation
between these defects.For
instance,
the defectcould be
primarily
invirionassemblage, and,
asaconsequence, thecleavageof different
precur-sorpolypeptidescould be blocked. As discussed
above, two of the mutants of this
phenotype
(ts17 andts29)possessanadditional
early
defect onthe basis of whichtheywerelistedasclass I mutants. Again, it is not known whether theyare double (or triple) mutants or single
pleio-tropicmutants.
25,1978 559
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.500.101.384.74.372.2].%*-i:W%m qa.*r- " *
-~~~~~~~~~~~~m._.
VISA,
pI5cE)---p12(
E)-A B C D E F G H
Chase in hr: 1 2 20 22/4 21/221 2,200
21 2, f ,2
FIG. 6. Normalprocessing ofgagandenvproductsofts28.NIH/3T3cellsinfectedat31°Cwith ts28were
pulse-labeled for30minwith 75 jCiofL-[35S]methionine,andradioactivitywaschasedforvarious timesas
indicated(A-L).Pulse-chaseexperimentswereperformedat39°C (A-D)or31°C(I-L).Thetemperaturewas
shifted from39to31°C (E-H),asdescribed in the text.Virus-specific polypeptideswereanalyzedasdescribed
in thelegendtoFig.1 (seealsolegendtoFig.2). Immunoprecipitationwith: anti-R-MuLVserum(A-L).
Finally,the structuralpolypeptidesof ts28are
produced normally at the restrictive
tempera-ture(Fig. 6). However,thedistortedmorphology of the virions formed at 390C (27) suggests a
defectinalatestageof theassembly.
ACKNOWLEDGMENTS
We thank J. R. Stephensonand S. A. Aaronson for the clonal stockofwild-typeR-MuLV and the ts R-MuLV
mu-tants, andwethankM. Strand and J. T.Augustfor the anti-gp69/71 andanti-p15sera.
These investigationswere supported by the Foundation
for Medical Research FUNGO,which is subsidized by the NetherlandsOrganizationforAdvancement of PureResearch (Z.W.O.).
LITERATURE CITED
1. Arcement, L. J., W. L. Karshin, R. B. Naso, G. Jamjoom,and R. B.Arlinghaus. 1976. Biosynthesis ofRauscher leukemiaviralproteins: presence ofp30 andenvelope p15sequencesinprecursorpolypeptides.
Virology 69:763-774.
2. Barbacid, M.,J. R.Stephenson,and S. A.Aaronson. 1976. Gag-gene of mammalian type-C RNA tumour
viruses. Nature(London) 262:554-559.
3. Bonner, W. M., and R. A. Laskey. 1974. A film detection method for tritium-labeled proteins and nucleic acids inpolyacrylamidegels. Eur. J. Biochem. 46:83-88.
4. Chopra, H. C., and G. P. Shibley. 1967. Infection ofan
establishedmousecell line with cell-free Rauscher
leu-kemia virus.I.Cytological studywith the electron
mi-croscope.J. Natl.Cancer Inst. 39:241-256.
5.Duesberg, P. H.,and W. S. Robinson. 1966. Nucleic
acids andproteinsisolated from the Rauschermouse
leukemia virus (MLV). Proc. Natl. Acad. Sci. U.S.A. 55:219-227.
6.Friis, R. R., K. Toyoshima, and P. K. Vogt. 1971.
Conditionallethal mutants of aviansarcomaviruses. I.
Physiologyof ts75 and ts149.Virology43:375-389.
7. Ikeda, H.,W.Hardy,Jr.,E.Tress,and E. Fleissner.
1975.Chromatographic separationandantigenic
anal-ysis of proteins of theoncornaviruses. V. Identification
ofanewmurineviralprotein, p15(E).J. Virol. 16:53-61.
8. Jamjoom,G.A.,R. B.Naso,and R. B.Arlinghaus. 1977. Further characterizationof intracellularprecursor polyproteins of Rauscher leukemia virus. Virology 78:11-34.
9. Kawai, S., and H. Hanafusa. 1972. Plaqueassay for
somestrains of avian leukosis virus.Virology48: 126-135.
10. Laemmli, U. K. 1970. Cleavage of structural proteins during the assemblyof the head ofbacteriophageT4.
Nature(London)227:680-685.
11. Leamnson,R.H.,M. H. M.Shandler,and M. S.
Hal-pern.1977. A structuralprotein complexinMoloney leukemiavirus.Virology76:437-439.
12. Naso, R. B., L.J. Arcement,W. L.Karshin, G. A.
Jamjoom, and R. B. Arlinghaus. 1976. A fucose-deficientprecursortoRauscherleukemia virusgp69/71. Proc. Natl. Acad. Sci. U.S.A. 73:2326-2330.
13. Scolnick,E.M.,J. R.Stephenson,and S. A.
Aaron-Pr
82I
pr
75gP9/7
1 0Pr65
0
J K
1 2
L
20
MWAO.Immmm.-.... ,WI; .110mmo.. .:...
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.90.439.71.350.2]son. 1972.Isolationoftemperature-sensitive mutants
of murinesarcomavirus. J. Virol.10:653-657.
14. Shapiro, S. Z., and J. T. August. 1976. Proteolytic cleavage events in oncornavirus protein synthesis. Biochim.Biophys.Acta 458:375-396.
15. Stephenson, J. R., and S. A. Aaronson. 1973. Charac-terizationoftemperature-sensitive mutantsofmurine leukemia virus.Virology 54:53-59.
16. Stephenson, J. R., and S. A. Aaronson.1974. Temper-ature-sensitive mutantsof murineleukemia virus: III. Mutants defective inhelper functions forsarcomavirus fixation.Virology 58:294-297.
17. Stephenson,J.R.,R. K.Reynolds, and S. A.
Aaron-son. 1972. Isolation oftemperature-sensitive mutants
ofmurineleukemia virus.Virology48:749-756. 18. Stephenson, J. R., S. R. Tronick, and S. A. Aaronson.
1974.Temperature-sensitivemutantsofmurine leuke-mia virus. IV. Furtherphysiological characterization and evidence for genetic recombination. J. Virol. 14:918-923.
19. Stephenson,J.R., S. R. Tronick, and S. A. Aaronson.
1975. Murineleukemia virusmutantswith temperature-sensitive defects inprecursorpolypeptide cleavage. Cell 6:543-548.
20. Strand, M.,and J.T.August.1976.Structural proteins of ribonucleic acid tumor viruses. J. Biol. Chem. 251:559-564.
21.Todaro, G., and H. Green. 1963. Quantitative studies
onthegrowth ofmouse embryo cells inculture and theirdevelopment into established lines. J. Cell. Biol. 17:299-313.
22.Toyoshima, K., and P. K. Vogt. 1969. Temperature-sensitivemutantsofanaviansarcomavirus.Virology
39:930-931.
23.Tronick, S. R., J.R. Stephenson, L.M.Verma, and S. A.Aaronson. 1975. Thermolabilereverse
transcrip-taseofamammalianleukemia virusmutant tempera-ture-sensitivein itsreplication andsarcomavirushelper functions. J. Virol. 16:1476-1482.
24.vanZaane, D., M. J. A. Dekker-Michielsen,andH. P. J. Bloemers.1976.Virus-specificprecursor polypep-tides in cells infected with Rauschermurineleukemia virus:synthesis, identificationandprocessing. Virology 75:113-129.
25.Wong, P. K. Y., and J. A. McCarter. 1973. Genetic studies of temperature-sensitive mutants ofMoloney murineleukemia virus. Virology 53:319-326. 26. Wyke, J.A.1973.The selectiveisolation of
temperature-sensitive mutants of Rous sarcoma virus. Virology 52:587-590.
27. Yeger, H., V.I.Kalnins, and J. R. Stephenson. 1976.
Electron microscopy ofmammalian type-C RNA
vi-ruses: useof conditional lethalmutants instudies of virion maturation andassembly. Virology 74:459-469.
561