Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Interaction between Hepatitis B Virus Core Protein and
Reverse Transcriptase
LISA LOTT,1,2BURTON BEAMES,1LENA NOTVALL,1ANDROBERT E. LANFORD1,2*
Department of Virology and Immunology, Southwest Regional Primate Research Center, Southwest Foundation for
Biomedical Research, San Antonio, Texas 78227,1and Department of Microbiology, University of Texas Health Science
Center at San Antonio, San Antonio, Texas 782292
Received 27 April 2000/Accepted 18 September 2000
Previous mutagenesis studies with hepatitis B virus (HBV) suggest that continued interactions with core are required for several steps in genomic replication. To examine core-polymerase (Pol) interactions, insect cells were coinfected with baculovirus constructs that independently expressed core and Pol. The results demon-strated several features with implications that core plays an interactive role with HBV Pol: (i) core coprecipi-tated with constructs expressing full-length Pol as well as the terminal protein (TP), reverse transcriptase (RT) and RNase H domains of Pol, independently; (ii) coprecipitation of core was not dependent on the presence of an epsilon stem-loop sequence; and (iii) core-Pol complexes migrated as intact capsid particles, as detected by sucrose gradient analysis. To analyze the structural and sequence requirements of core in recognition of Pol, a series of core mutants with two- to four-amino-acid insertions or carboxy-terminal deletions were assessed for Pol interaction. The results indicated that capsid formation is required but not sufficient for interaction with Pol and that the TP and RT domains of Pol have different requirements for interaction with core. To map the core binding sites on Pol, a panel of amino- and carboxy-terminal deletion mutants of the TP and RT domains of Pol were analyzed for interaction with core. At least three separate core binding sites on Pol were detected. This analysis begins to define basic requirements for core-Pol interactions, but further study is necessary to delineate the effects of these interactions on encapsidation and genome replication.
Hepatitis B virus (HBV) is a member of theHepadnaviridae
family. Mature HBV virions are enveloped and contain par-tially double stranded, non-covalently closed, circular DNA with the viral polymerase (Pol) covalently bound to the first deoxyribonucleotide of the 5⬘end of the minus-strand DNA. HBV replicates by reverse transcription of a pregenomic RNA molecule which occurs inside of a capsid particle (for reviews see references 11, 32, and 39). For encapsidation and subse-quent genomic replication to occur, a ribonucleoprotein (RNP) complex forms between Pol and the epsilon stem-loop structure on a pregenomic RNA molecule (2). The epsilon stem-loop is present on both the 5⬘and 3⬘ends of pregenomic RNA, but only the 5⬘ copy of epsilon functions in vivo as a packaging signal (14, 16, 18, 34). A sequence in a bulge in the 5⬘ copy of epsilon functions as a template for the first four nucleotides in DNA synthesis by a protein-primed reverse transcription reaction catalyzed by Pol (41, 42). Minus-strand DNA synthesis continues after a translocation step in which the primed-Pol complex translocates to a complementary se-quence in the 3⬘copy of DR1 (8, 26, 29, 36–38, 44). Whether the priming reaction occurs after packaging or prior to capsid assembly is not known, nor is the mechanism by which the RNA-Pol RNP complex interacts with the capsid protein to produce replication-competent HBV particles.
The HBV capsid is composed of a single type of protein that is referred to as the core protein and consists of 183 amino acids. The core protein spontaneously forms capsids in the absence of the pregenomic RNA or Pol when expressed in insect cells (23), bacteria (7), yeast (28), orXenopusoocytes
(47). The core protein contains numerous phosphorylation sites, with a cluster of sites located at the carboxy terminus. The phosphorylation state of core is associated with confor-mational changes in the capsid particle (46) which may have significant function after the assembly of core particles and the encapsidation of viral RNA (17, 35). The carboxy terminus of the HBV core protein is very basic and contains four arginine-rich domains. Although this region is not necessary for particle formation, the carboxy terminus binds nucleic acids, which could facilitate encapsidation. Core exhibits DNA and RNA binding abilities in vitro, but no specificity between core and HBV RNA has been demonstrated (10, 12, 27, 33) in the absence of Pol. Both the Pol molecule and an epsilon stem-loop structure are essential for packaging, although neither is required to be replication competent for encapsidation to oc-cur (1, 13).
Previous mutagenesis studies with HBV and duck hepatitis B virus (DHBV) core examined the requirements of the capsid protein during encapsidation and viral DNA synthesis (3, 4, 31, 35, 45). Capsid formation and RNA encapsidation are not sufficient to ensure efficient viral DNA synthesis. Schlicht and coworkers reported that the deletion of 36 amino acids from the carboxy terminus of the DHBV core protein abolished genome replication, although mutant cores formed capsid par-ticles competent for genome packaging (35). Yu and Summers examined a series of deletions covering the 3⬘ end of the DHBV capsid open reading frame (ORF) and detected mu-tant phenotypes supporting various levels of DNA synthesis, ranging from wild-type (WT) DNA replication, to single-stranded DNA in the absence of relaxed circular DNA, to mutants in which replication was severely reduced (45). Studies with HBV core particles from transfected Huh7 cells revealed that when 12 amino acids were deleted from the carboxy ter-minus, core particles supported WT levels of RNA packaging but were defective in minus-strand DNA synthesis (3). Mutant
* Corresponding author. Mailing address: Department of Virology and Immunology, Southwest Regional Primate Research Center, Southwest Foundation for Biomedical Research, 7620 N.W. Loop 410, San Antonio, TX 78227. Phone: (210) 258-9445. Fax: (210) 670-3329. E-mail: [email protected].
11479
on November 9, 2019 by guest
http://jvi.asm.org/
proteins truncated to amino acid 163 or 164 package RNA but are drastically reduced in DNA synthesis (3, 31). Core proteins with further deletion to amino acid 144 were assembly compe-tent but did not package RNA (3, 31). The observation that deletions in the carboxy terminus of core cause a progressive loss of genome replication and ultimately loss of RNA pack-aging suggests that continued and specific interactions with core are required for several steps in genome replication.
In this study, we used a series of core insertion and carboxy-terminal deletion mutants to examine the structural and se-quence requirements of core for interacting with HBV Pol. Previous studies using this system have demonstrated that pu-rified human HBV Pol expressed in insect cells is active in protein-primed reverse transcription (24). Furthermore, inde-pendent expression of the TP and RT domains of Pol within the same cells results in the formation of a stable RNP complex with epsilon that is active in nucleotide priming and reverse transcription (25). In this study, insect cells were coinfected with baculovirus constructs independently expressing core and Pol. Immunoprecipitation reactions and treatment with RNase A confirmed that an interaction exists between HBV core and Pol that is not dependent on an epsilon stem-loop structure. In
addition, core coprecipitated with both the TP, RT, and RNase H domains, suggesting that core interacts with at least three sites on Pol. Analysis of Huh7 cells coinfected with vaccinia virus constructs expressing core and Pol revealed that core-Pol interaction extends to mammalian cells. Sucrose gradient anal-ysis demonstrated that Pol interacts with cores from capsid fractions; however, coprecipitation assays between mutant cores and the TP or RT domain revealed that capsid formation is required but not sufficient for these core-Pol interactions.
MATERIALS AND METHODS
Cells.The Sf9 cell line derived fromSpodoptera frugiperdawas maintained in spinner culture as described previously (23). Sf9 cells were cultivated in TNMFH supplemented with 5% fetal bovine serum and 0.1% pluronic F68. After infec-tions, the medium was changed to Grace’s medium supplemented with 2% fetal bovine serum. The human hepatoma cell line Huh7 (30) was maintained in Dulbecco modified Eagle medium–F-12 medium (1:1) plus 10% fetal bovine serum, 2 mM glutamine, and 50g of gentamicin per ml.
Plasmid constructs.HBV sequences of theaywsubtype are numbered as specified by Galibert and coworkers (9). The core deletion and insertion mutants (Fig. 1A) were generated as described previously (3, 4). Core proteins with carboxy-terminal deletions are designated Cd followed by a number representing the last carboxy-terminal amino acid. Insertions of two to four amino acids in core are designated Ci followed by a number representing the location of the
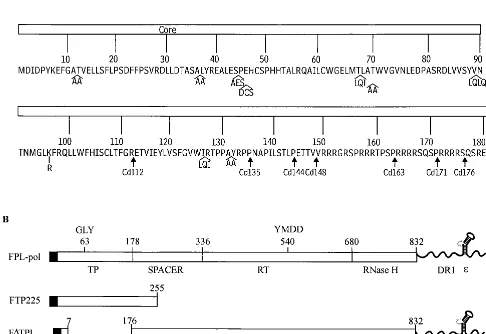
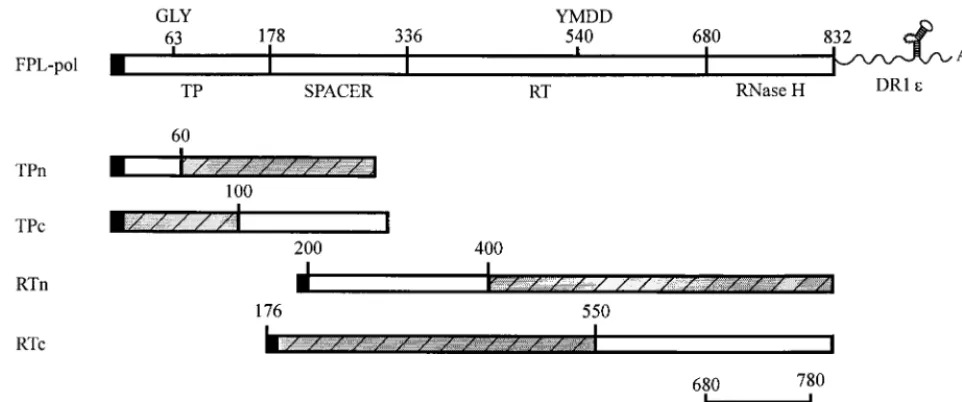
FIG. 1. (A) Amino acid sequence of the core gene. Insertions within the core gene are illustrated with the inserted amino acid sequence in single-letter code. A single point mutation of Lys to Arg is represented by a line at residue 96. Core deletions are illustrated below the amino acid sequence and designated by arrows. (B) Structures of FLAG-Pol constructs. FPL-pol depicts the structure of the WT Pol construct. ThepolORF is designated by open rectangles. The TP, spacer, RT, and RNase H domains are indicated below, and the GLY and YMDD motifs are labeled above, thepolORF. The F indicates the presence of a 10-amino-acid FLAG sequence at the amino terminus of Pol, which is illustrated by a solid bar; the L denotes an epsilon stem-loop structure, which is present at the 3⬘end of the Pol contructs. FTP225 denotes the amino-terminal 225 amino acids of Pol with a FLAG epitope; F⌬TPL denotes a deletion in the TP domain, but the presence of a FLAG epitope and an epsilon stem-loop structure.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.61.547.88.422.2]insertion. The point mutation used for this study is designated Cp96 to denote a Lys-to-Arg mutation at amino acid 96.
The construct expressing full-length Pol, designated FPL-pol (Fig. 1B), has the 10-amino-acid FLAG epitope (International Biotechnologies Inc., New Haven, Conn.) fused to the amino terminus as specified previously (24). The 3⬘terminus was extended beyond thepolORF to include DR1 and epsilon stem-loop se-quences. All Pol mutants were generated as previously described (22, 25). FTP225 (Fig. 1B) contains the amino-terminal 225 amino acids of Pol with a FLAG epitope. F⌬TPL (Fig. 1B) denotes a deletion in the TP domain of Pol, but the presence of a FLAG epitope and an epsilon stem-loop. F⌬TP⌬3⬘denotes a deletion in the TP domain and deletion of the 3⬘extension containing DR1 and epsilon. FTP199 expresses the amino-terminal 199 amino acids of Pol with a FLAG epitope. The HCV protein NS5B contains a FLAG epitope and was provided as a control (C. Bigger, unpublished data).
The vaccinia virus constructs FPL-pol and core contain an insert identical to the FPL-pol in baculovirus. FPL-pol and core were cloned into the pVOTE2 vaccinia virus vector (43), in which expression is regulated by T7 RNA polymer-ase and is underlacrepression.
The deletion mutants of the TP and RT domains were created by PCR mutagenesis as described previously (22). The amino-terminal deletions of TP and RT are named by the first amino acid at the amino terminus (e.g., FRTn300 for FLAG-RT from N-terminal amino acid 300 through C-terminal amino acid 832 with an epsilon stem-loop, and FTPn20/c300 for FLAG-TP from N-terminal amino acid 20 through C-terminal amino acid 300.) The carboxy-terminal dele-tions of TP and RT are named for the carboxy-terminal amino acid (e.g., FTPc300 spans amino acids 1 to 300, and FRTc800 spans amino acids 176 to 800). A minimal RNase H construct (FRNH) spans amino acids 680 to 780 and contains a FLAG epitope.
Immunoprecipitation.Pol proteins were immunoprecipitated with anti-FLAG affinity beads (M2 monoclonal antibody beads; Sigma). Insect cells were infected or coinfected with baculovirus constructs, harvested at 48 h postinfection by washing two times in phosphate-buffered saline (PBS), and extracted with PEB (PBS containing 10% glycerol, 0.5% Nonidet P-40, and protease inhibitors [100 M leupeptin, 1 mM Pefabloc, 10M aprotinin, 10g of pepstatin per ml, and 1 mM EDTA]). Clarified cell lysates were immunoprecipitated with anti-FLAG affinity beads for 2 h at 4°C. The beads were washed with PEB, one time with PEB–1 M NaCl, and a final time with PEB. Huh7 cells were coinfected with vaccinia virus constructs, harvested at 24 h postinfection and immunoprecipi-tated as described above.
SDS-PAGE and immunoblot analysis.Immunoprecipitated complexes were disrupted in electrophoresis sample buffer containing 2% sodium dodecyl sulfate (SDS) and 2% 2-mercaptoethanol and were heated to 100°C for 10 min. Proteins were separated by SDS-polyacrylamide gel electrophoresis (PAGE) as previously described (19, 20), and immunoblot analysis was performed by electrophoreti-cally transferring proteins to a Flurotrans polyvinylidene difluoride blotting membrane (Pall Biosupport, Glen Cove, N.Y.). Membranes were processed as previously described (21), using iodinated M2 monoclonal antibody to detect the FLAG epitope on Pol or with a rabbit anticore polyclonal antibody followed by
125I-protein A (NEN, Boston, Mass.).
Sucrose gradient analysis.Core and Pol proteins expressed in insect cells were analyzed by sucrose gradient centrifugation followed by immunoblotting. Clari-fied cell lysates from infected Sf9 cells were applied to 5-ml gradients of 10 to 50% sucrose in PBS, and sedimentation was performed in a Beckman SW55 rotor at 40,000 rPM (approximately 200,000⫻g) for 2 h. Ten 0.5-ml fractions were collected, and a portion of each fraction was analyzed by SDS-PAGE and immunoblotting as described above.
RESULTS
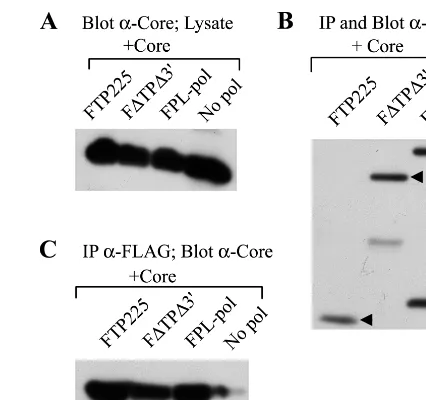
Core coprecipitates with the TP and RT domains of Pol in the absence of epsilon.To examine potential interactions be-tween core and Pol, insect cells were coinfected with baculo-viruses independently expressing core and Pol proteins, and core-Pol complex formation was evaluated by coprecipitation studies. All Pol constructs contained a FLAG epitope fused to the amino terminus of Pol; thus, Pol was immunoprecipitated with anti-FLAG antibodies, and coprecipitation of core was detected by Western blotting with anticore antibodies. Immu-noprecipitates were immunoblotted with anti-FLAG antibod-ies to demonstrate the level of immunoprecipitated Pol (Fig. 2B). A fraction of the cell lysates were immunoblotted with anticore antibodies to detect core protein expression (Fig. 2A). Coinfection of insect cells with core and FPL-pol indicated that core is able to coprecipitate with full-length Pol (Fig. 2C). No core was immunoprecipitated in the absence of Pol. Coinfec-tion of insect cells with baculoviruses expressing WT core and the TP (FTP225) or RT (F⌬TP⌬3⬘) domain of Pol revealed
that core coprecipitates with at least two sites on Pol (Fig. 2C). The fact that core was able to coprecipitate with TP (FTP225) and RT (F⌬TP⌬3⬘) lacking epsilon stem-loop structures re-vealed that epsilon is not required for these core-Pol interactions. To determine whether core was interacting with the FLAG epitope rather than Pol itself, we coinfected insect cells with baculoviruses expressing WT core and the HCV protein NS5B that contains a FLAG epitope. NS5B was immunoprecipitated with anti-FLAG antibodies, and Western blot analysis was performed with a mixture of anti-FLAG and anticore antibod-ies to detect precipitation of NS5B and coprecipitation of core. A fraction of the cell lysate was immunoblotted with anti-FLAG antibodies and anticore antibodies to confirm protein expression (Fig. 3A). The results revealed that NS5B but not core immunoprecipitated with anti-FLAG antibodies, indicat-ing that core does not interact with the FLAG epitope (Fig. 3B).
Detection of core-Pol complex formation in Huh7 cells.To determine if a core-Pol complex forms when the proteins are expressed in mammalian cells, Huh7 cells were coinfected with vaccinia virus constructs expressing WT core and full-length Pol. The Pol proteins were immunoprecipitated with anti-FLAG antibodies, and Western blot analysis was performed with anticore and anti-FLAG antibodies. A fraction of the cell lysate was immunoblotted to confirm core protein expression (Fig. 4A). Pol protein expression is lower than core expression and was detected only after immunoprecipitation with anti-FLAG antibodies (Fig. 4B). Coprecipitation of core confirmed that core-Pol interaction extends to mammalian cells.
[image:3.612.322.535.71.271.2]Treatment with RNase A does not disrupt core-Pol interac-tions.Although epsilon was not required for core-Pol complex formation, this did not rule out the possibility that the inter-action was dependent on an RNA intermediate. To test this possibility, we coinfected insect cells with baculoviruses ex-pressing core and TP, RT, or full-length Pol and then immu-noprecipitated the Pol proteins with anti-FLAG antibodies. FIG. 2. Interaction of core with the TP and RT domains of Pol. Insect cells were coinfected with baculoviruses expressing core and a Pol construct. (A) A fraction (1/20) of the cell lysate was immunoblotted with anticore antibodies to confirm that the core protein was expressed. (B) Pol proteins were immunopre-cipitated (IP) and immunoblotted with anti-FLAG antibodies. (C) Coprecipita-tion of core was detected by Western blot analysis with anticore antibodies. The negative control (No pol) is a single infection with WT core, illustrating that core does not immunoprecipitate with M2 affinity beads. Core coprecipitates with full-length Pol (FPL-pol) and with FTP225 and F⌬TP⌬3⬘, TP and RT constructs that lack epsilon stem-loop structures.
on November 9, 2019 by guest
http://jvi.asm.org/
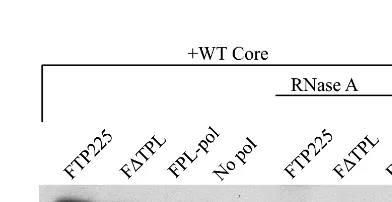
Identical sets of immunoprecipitates were incubated for 30 min at 37°C in the presence or absence of RNase A (50g/ml), while the proteins were still bound to the affinity beads. The beads were washed to remove any core released during the RNase treatment, and the resulting immunoprecipitates were examined by Western blotting with anticore antibodies (Fig. 5). No difference in the level of coprecipitated core was observed in the treated and untreated immunoprecipitates. Although the presence of a small, protected RNA fragment cannot be excluded, the results suggest that RNA is not required to maintain the interaction between core and Pol.
Interaction of TP and RT with carboxy-terminal deletion mutants of core.To analyze the structural and sequence re-quirements of core for recognition of Pol, we used a series of carboxy-terminal truncations of the 183-amino-acid core pro-tein. Insect cells were coinfected with baculoviruses expressing a core deletion mutant and TP or RT. Core mutant proteins are designated Cd followed by the carboxy-terminal amino acid number. A fraction of the cell lysate was immunoblotted with anticore antibodies to detect core protein expression (Fig. 6A). lmmunoprecipitation and Western blot analysis was performed with anti-FLAG antibodies to detect the Pol polypeptide (Fig. 6B), and immunoblotting with anticore antibodies was used to detect coprecipitation of core (Fig. 6C). The patterns of RT binding to the core deletion proteins were identical to that of TP. The core deletion proteins Cd163, Cdl71, and Cd176 co-precipitated with TP and RT (Fig. 6C). Deletion to amino acid 148 abolished coprecipitation of core, although further dele-tion to amino acid 144 restored core-Pol interacdele-tion. No bind-ing to core deletions beyond amino acid 144 was detected. These results suggest that progressive deletion of amino acids from core may induce a change in conformation, consequently preventing interaction between the core deletion mutant Cd148 and the TP and RT domains of Pol. Further deletion to amino acid 144 may restore a conformation that permits a binding interaction to occur. In addition, these results revealed that at least one TP and RT binding site resides within the first 144 amino acids of core. The data do not exclude the existence of additional binding sites from amino acids 163 to 183.
[image:4.612.319.547.70.368.2]Interaction of core insertion mutants with TP and RT.To investigate the influence of two- to four-amino-acid insertions within the core protein on core-Pol interactions, insect cells were coinfected with baculoviruses expressing core insertion proteins and TP or RT. Immunoprecipitation was performed with anti-FLAG antibodies for Pol, followed by Western blot FIG. 3. Core does not interact with the FLAG epitope on Pol. (A) Insect
[image:4.612.71.275.72.236.2]cells were coinfected with baculoviruses expressing WT core and the HCV protein NS5B that contains a FLAG epitope. A fraction (1/20) of the cell lysate was immunoblotted with anticore antibodies to confirm that the core protein was expressed. (B) Immunoprecipitation (IP) was performed with FLAG anti-bodies, and Western blot analysis was performed with a mixture of anticore and anti-FLAG antibodies. NS5B immunoprecipitates with anti-FLAG antibodies. Core does not coprecipitate with anti-FLAG antibodies or with NS5B.
FIG. 4. Core-Pol interaction in mammalian cells. Huh7 cells were coinfected with vaccinia virus constructs expressing core and Pol. The negative control (No pol) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblotted with anti-FLAG and anticore antibodies. (B) Lysates were im-munoprecipitated (IP) with anti-FLAG antibodies to Pol and immunoblotted with anti-FLAG and anticore antibodies. WT core coprecipitates with HBV Pol expressed in liver cells.
FIG. 5. Treatment with RNase A does not disrupt core-Pol interactions. Insect cells were coinfected with baculoviruses expressing core and TP, RT, or full-length Pol, and the Pol proteins were immunoprecipitated with anti-FLAG antibodies. The control (No pol) is a single infection with WT core followed by immunoprecipitation with anti-FLAG antibodies. Core-Pol complexes were in-cubated for 30 min at 37°C in the presence or absence of RNase A (50g/ml), while the proteins were still bound to the affinity beads. The affinity beads were washed to remove any core released by RNase treatment, and coimmunopre-cipitated core was analyzed by SDS-PAGE and Western blot analysis with anti-core antibodies.
on November 9, 2019 by guest
http://jvi.asm.org/
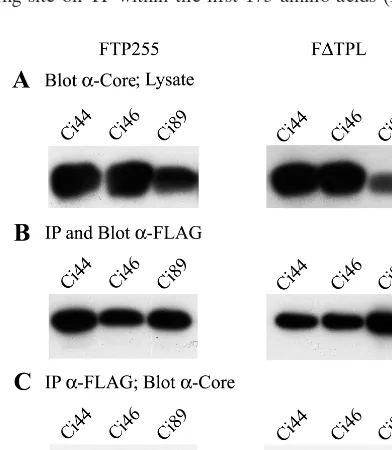
[image:4.612.331.527.542.643.2]analysis with anticore antibodies. Core insertion mutants are designated Ci to denote a two- to four-amino-acid, in-frame insertion followed by the amino acid number that corresponds to the site of insertion. Cp96, a point mutation at amino acid 96, was used in this study. Core mutants with insertions at the amino terminus, Ci11 and Ci36, and carboxy terminus, Ci126 and Ci131, were unable to coprecipitate with TP or RT (Table 1). Mutants with insertions in the central domain of core (Ci46, Ci67, Ci69, Ci89, and Cp96) coprecipitated with both TP and RT. The core insertion mutant Ci44 coprecipitated with TP but not RT (Fig. 7C), revealing that in this instance, TP and RT have different requirements for interacting with core.
Pol interacts with HBV capsid particles. Sucrose gradient analysis was performed to determine the molecular form of core that coprecipitated with Pol. Insect cells were coinfected with baculoviruses expressing WT core and full-length Pol. The clarified lysates were separated by velocity sedimentation on 10 to 50% sucrose gradients and immunoblotted with anticore antibodies. The WT core protein formed capsid particles which sedimented near the middle of the gradient, and a small
amount of dimeric core sedimented at the top of the gradient (Fig. 8A). Immunoblot analysis of gradient fractions of FPL-pol expressed from a single infection (Fig. 8B), or from a coinfection with baculoviruses expressing core (Fig. 8C), re-vealed similar distribution patterns in which Pol proteins were distributed throughout the top and middle of the gradients (fractions 2 to 5). Immunoblot analysis of the core protein coprecipitating with Pol revealed that Pol interacts with cores from capsid fractions (Fig. 8D). Pol was detected primarily in noncapsid fractions; however, the amount Pol in association with core is below our level of detection. The failure to detect Pol in the fractions in which core coprecipitated suggests that a small percentage of the Pol enters into the core-Pol interac-tion, indicative of a high core-to-Pol ratio, perhaps approach-ing the stoichiometry observed in virions.
Analysis of amino- and carboxy-terminal deletion mutants of TP for interaction with core.To map core binding sites on the TP polypeptide, we analyzed several amino- and carboxy-terminal deletion mutants of TP for the ability to coprecipitate core. Within the carboxy-terminal deletion panel, the largest FIG. 6. Interactions of TP and RT with carboxy-terminal deletion mutants of core. Insect cells were coinfected with baculoviruses expressing a core deletion mutant and TP or RT (F⌬TPL). The control (No TP or No RT) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblotted with anticore antibodies to confirm that the core protein was expressed. Immunoprecipitation (IP) was performed with anti-FLAG antibodies followed by SDS-PAGE and Western blot analysis with anti-FLAG antibodies (B) to show the presence of TP and RT or with anticore antibodies (C) to detect coprecipitation of core. Core mutants Cd144, Cd163, Cd171, and Cd176 coprecipitated with TP and RT, while core mutants Cd112, Cd135, and Cd148 failed to coprecipitate with TP or RT.
on November 9, 2019 by guest
http://jvi.asm.org/
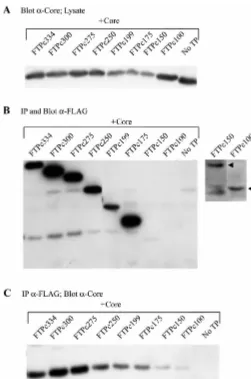
and smallest constructs spanned amino acids 1 to 334 and 1 to 100, respectively, and contained an amino-terminal FLAG epitope. The amino-terminal deletion panel contained dele-tions of 20, 30, 50, and 60 amino acids from the 1-300 TP construct. Insect cells were coinfected with baculovirus con-structs expressing core and a deletion mutant of the TP do-main. Immunoprecipitation and Western blot analysis was per-formed with anti-FLAG antibodies to detect the TP deletion mutants (Fig. 9B and 10B), followed by immunoblotting with anticore antibodies to detect coprecipitation of core (Fig. 9C and 10C). Carboxy-terminal deletions spanning to amino acid 175 were positive for coprecipitation of core, indicating a core binding site on TP within the first 175 amino acids (Fig. 9C).
Constructs spanning from amino acids 1 to 150 and smaller were not well expressed, and coprecipitation of core could not easily be interpreted; however, the level of core that coprecipi-tated with the 150- and 100-amino-acid TP polypeptides was greater than the negative control. The amino-terminal deletion mutants of TP coprecipitated core, with the largest deletion extending to amino acid 60 (Fig. 10C). This indicates that a core binding site exists on TP between amino acids 60 and 300. Taken together, these data suggest a common binding site for core on TP within amino acids 60 to 100. Alternatively, the data are also consistent with the presence of at least two separate binding sites, one in the amino terminus and one in the carboxy terminus of the TP domain.
Analysis of amino- and carboxy-terminal deletion mutants of RT for interaction with core.To determine the core binding pattern on the RT polypeptide, several amino- and carboxy-terminal deletion mutants of the RT domain were analyzed for interaction with core. The largest RT construct with a carboxy-terminal deletion spanned amino acids 176 to 800 (FRTc800), and deletions extended to amino acid 550 (FRTc550). Within the amino-terminal deletion panel, the largest construct spanned amino acids 300 to 832 (FRTn300), and the smallest construct spanned amino acids 400 to 832 (FRTn400). Insect cells were coinfected with baculovirus constructs expressing core and a deletion mutant of the RT domain. Immunopre-cipitation and Western blot analysis were performed with anti-FIG. 7. TP and RT differ in requirements for interacting with core. Insect
[image:6.612.331.532.69.341.2]cells were coinfected with baculoviruses expressing a core insertion mutant (Ci44, Ci46, or Ci89) and TP or RT (F⌬TPL). (A) A fraction (1/20) of the cell lysates were immunoblotted with anticore antibodies to confirm that the core insertion mutants were expressed. (B) Immunoprecipitation (IP) was performed with FLAG antibodies, followed by immunoblotting with FLAG anti-bodies to confirm that the TP and RT polypeptides were expressed and immu-noprecipitated. (C) Immunoprecipitation was performed with FLAG anti-bodies, followed by immunoblotting with anticore antibodies. Ci46 and Ci89 coprecipitated with both TP and RT, while Ci44 failed to coprecipitate with RT.
[image:6.612.53.294.92.218.2]FIG. 8. Pol interacts with capsid particles. Insect cells were infected with baculoviruses expressing FPL-pol or coinfected with WT core and FPL-pol. The clarified lysates were sedimented in 5-ml 10 to 50% sucrose gradients at 200,000⫻gfor 2 h. Ten 0.5-ml fractions were collected, and a portion was analyzed by SDS-PAGE and Western blot analysis. (A) The WT core protein from a coinfection with FPL-pol formed capsid particles that sedimented pri-marily in fractions 5 to 7. Immunoblot analysis was performed with anticore antibodies. Immunoprecipitation and immunoblot analysis were performed with anti-FLAG antibodies to FPL-pol expressed from a single infection (B) or from a coinfection with WT core (C). (D) Gradient fractions from a coinfection with FPL-pol and WT core were immunoprecipitated (IP) with anti-FLAG antibodies and immunoblotted with anticore antibodies to determine the molecular form of core that interacts with Pol.
TABLE 1. Coprecipitation of core insertion mutants with the RT and TP domains of Pola
Mutant Coprecipitation of core with:
RT TP
Ci11 ⫺ ⫺
Ci36 ⫺ ⫺
Ci44 ⫺ ⫹
Ci46 ⫹ ⫹
Ci67 ⫹ ⫹
Ci69 ⫹ ⫹
Ci89 ⫹ ⫹
Cp96 ⫹ ⫹
Ci126 ⫺ ⫺
Ci131 ⫺ ⫺
aInsect cells were coinfected with baculoviruses expressing a core insertion mutant and the RT or TP domain of Pol. Pol polypeptides were immunopre-cipitated with anti-FLAG antibodies, and the coprecipitation of core (⫹) or absence of core (⫺) was evaluated by Western blot analysis with anticore anti-bodies.
on November 9, 2019 by guest
http://jvi.asm.org/
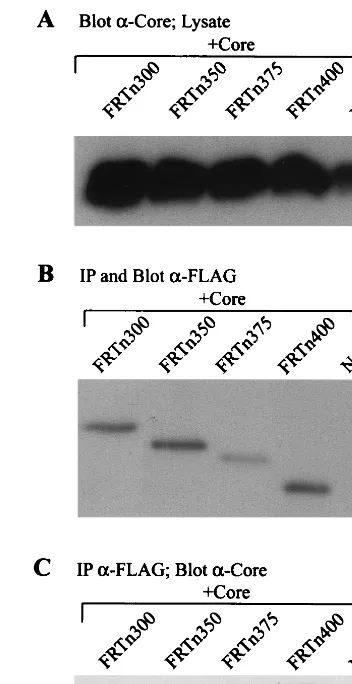
[image:6.612.75.271.418.643.2]FLAG antibodies to detect the RT deletion mutants (Fig. 11B and 12B) followed by immunoblotting with anticore antibodies to detect coprecipitation of core (Fig. 11C and 12C). All car-boxy-terminal deletion mutants were positive for coprecipita-tion of core, indicating a core binding site on RT between amino acids 176 and 550 (Fig. 11C). Core also coprecipitated with each of the amino-terminal deletion mutants of the RT domain, identifying a core binding site on Pol between amino acids 400 and 832 (Fig. 12C). One possibility to explain the nature of these interactions is the presence of a common core binding site on Pol between amino acids 400 and 550. How-ever, the data are also consistent with the presence of two or more core binding sites, one in the amino terminus and one in the carboxy terminus of the RT polypeptides.
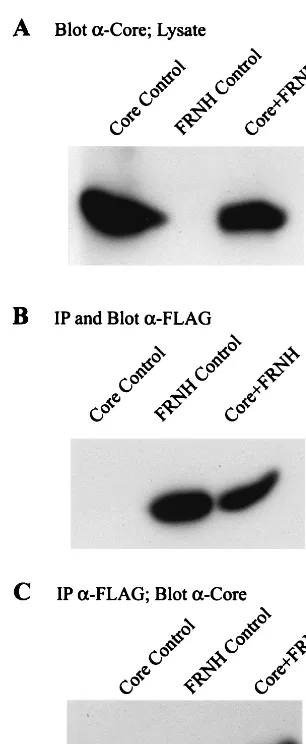
To test whether a core binding site was present in the car-boxy terminus of the RT constructs, we examined FRNH, a minimal RNase H construct which spanned amino acids 680 to 780. Insect cells were coinfected with baculovirus constructs expressing FRNH and WT core. Immunoprecipitation was performed with anti-FLAG antibodies to FRNH, followed by Western blot analysis with anti-FLAG and anticore antibodies. The results demonstrate that core coprecipitates with a mini-mal RNase H polypeptide (Fig. 13C).
DISCUSSION
[image:7.612.313.549.69.478.2]In this study, we described a model system for the analysis of core-Pol interactions to begin to define the requirements of FIG. 9. Coprecipitation of core with carboxy-terminal deletion mutants of
the TP domain. Insect cells were coinfected with baculoviruses expressing car-boxy-terminal deletion mutants of the TP domain and WT core. The control (No TP) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblotted with anticore antibodies to confirm that the core protein was expressed. Immunoprecipitation (IP) was performed with anti-FLAG antibodies followed by SDS-PAGE and Western blot analysis with anti-FLAG antibodies (B) to show the presence of the TP polypeptides. A longer exposure demon-strating polypeptides FTPc150 and FTPc100 is shown to the right. (C) Western blot analysis with anticore antibodies demonstrates that a definitive core binding site exists on TP between amino acids 1 and 175 and may exist in the polypeptide between amino acids 1 and 100.
FIG. 10. Coprecipitation of core with amino-terminal deletion mutants of the TP domain. Insect cells were coinfected with baculoviruses expressing amino-terminal deletion mutants of the TP domain and WT core. The control (No TP) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblotted with anticore antibodies to confirm that the core protein was expressed. Immunoprecipitation (IP) was performed with anti-FLAG antibodies followed by SDS-PAGE and Western blot analysis with anti-FLAG antibodies (B) to show the presence of the TP polypeptides or with anticore antibodies (C) to detect coprecipitation of core. A core binding site was detected on TP between amino acids 60 and 300.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.52.303.71.450.2]these interactions in HBV encapsidation and replication. Core-Pol interaction was observed in insect cells coinfected with baculoviruses expressing core and Pol, as well as in the Huh7 liver cell line coinfected with vaccinia virus core and Pol. The results indicate the occurrence of a protein-protein inter-action involving Pol and core that was not dependent on an epsilon stem-loop sequence and was not disrupted after treat-ment with RNase. In addition, core interacts with at least three sites on Pol, within the TP, RT, and RNase H domains. One binding site was detected within the amino terminus of the TP domain. Binding was also observed with individual fragments spanning amino acids 176 to 550 of the RT polypeptides and amino acids 680 to 780 of the RNase H domain (Fig. 14). Interaction of core with the RNase H domain could conceiv-ably be involved in encapsidation, since point mutations in the RNase H domain disrupt packaging (1, 5, 6, 13).
Sucrose gradient analysis revealed that Pol exists in a com-plex with capsid particles, suggesting either that the core-Pol interaction occurs between Pol and capsid particles or that Pol interacts with a core monomer/dimer which is subsequently incorporated into a newly formed capsid particle. Two lines of evidence support Pol interaction with capsid particles. Core protein in the monomer/dimer fraction of sucrose gradients did not coprecipitate with Pol, while interaction was observed between Pol and capsid particles. Moreover, the analysis of a large panel of core mutants suggested that capsid formation was an essential attribute of core for interaction with Pol. The results indicate that at least the FLAG epitope was exposed on the exterior of the particle. Although during replication Pol would be expected to reside on the interior of the particle, this model system could be detecting a trapped intermediate in the encapsidation process. The possibility that the Pol-epsilon RNP complex interacts with a precapid-like particle during encapsidation has not been formally excluded.
[image:8.612.342.518.71.413.2]At this time, we are unable to form core-Pol complexes using purified proteins from baculovirus constructs or by the mixing FIG. 11. Coprecipitation of core with carboxy-terminal deletion mutants of
the RT domain. Insect cells were coinfected with baculoviruses expressing ami-no-terminal RT deletion mutants and WT core. The control (No RT) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblot-ted with anticore antibodies to detect core protein expression. Immunoprecipi-tation (IP) was performed with anti-FLAG antibodies followed by SDS-PAGE and Western blot analysis with anti-FLAG antibodies (B) to show the presence of the RT polypeptides or with anticore antibodies (C) to detect coprecipitation of core. A core binding site extending from amino acids 176 to 550 was detected on the RT polypeptide.
FIG. 12. Coprecipitation of core with amino-terminal deletion mutants of the RT domain. Insect cells were coinfected with baculoviruses expressing carboxy-terminal RT deletion mutants and WT core. The control (No RT) is a single infection with WT core. (A) A fraction (1/20) of the cell lysate was immunoblot-ted with anticore antibodies to detect core protein expression. Immunoprecipi-tation (IP) was performed with anti-FLAG antibodies followed by SDS-PAGE and Western blot analysis with anti-FLAG antibodies (B) to show the presence of the RT polypeptides or with anticore antibodies (C) to detect coprecipitation of core. A core binding site was detected on Pol between amino acids 400 and 832.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.66.279.73.456.2]of cell lysates containing core and Pol. A number of factors could contribute to the requirement for coexpression. Pol may interact in a nascent form before complete folding has oc-curred. This is not likely to be the case for core, since it would imply interaction at the monomer/dimer level. Alternatively, host factors may be involved in the conformational require-ments for interaction or more directly in a mechanism sup-porting initial contact. The role of Hsp90 in Pol-epsilon inter-action has been demonstrated in the DHBV system. In DHBV, RNP formation depends on cellular factors that include Hsp90 and the chaperone partner p23, which are incorporated into the viral nucleocapsid (15). The requirement of cellular factors for core-Pol interactions was not explored in this study.
Delineation of the steps in HBV replication represented by the core-Pol interaction examined in this study is difficult in the absence of an in vitro encapsidation and genome replication system. This study may represent one step in the development
of such a system. Core-Pol interactions could play a role in multiple steps in HBV replication. Encapsidation of the pre-genomic RNA may require direct core-Pol interaction even though the encapsidation substrate is an epsilon-Pol RNP complex. The role of epsilon in this process could be to induce a proper Pol conformation for interaction with core. Studies with DHBV Pol support the contention that epsilon induces a conformational change in Pol that is required for protein prim-ing (40). The fact that core-Pol interaction in insect cells was not dependent on epsilon does not imply that this interaction is not essential for packaging. Overproduction of Pol in insect cells may result in Pol polypeptides with a variety of confor-mations, some of which may mimic the conformation induced by epsilon, perhaps by interaction with cellular RNAs with epsilon-like properties. Evidence for this also comes from our studies on in vitro nucleotide priming with HBV Pol from insect cells. With full-length Pol this event appears to be inde-pendent of epsilon, although an RNA template is required (24). Epsilon is required for the transcomplementation of nu-cleotide priming between independently expressed TP and RT in insect cells (25). In this instance, epsilon may serve to cor-rectly align the interacting TP and RT polypeptides and induce a conformation permissive for priming. Together, these studies and the observation that Pol cannot be encapsidated in the absence of epsilon suggest that one of the primary roles of epsilon may be to induce a permissive conformation on Pol for priming activity and interaction with core.
Core-Pol interactions are probably essential in genome rep-lication beyond the encapsidation step. Evidence for a contin-ued interaction of core and Pol during genome replication can be extrapolated from mutagenesis studies on core. The dele-tion of amino acids from the carboxy terminus of core results in a progressive deficiency in completion of HBV genome replication. Previous work with these core deletion mutants revealed that the deletion of carboxy-terminal amino acids from core resulted in the progressive loss of DNA synthesis and RNA packaging. Deletion to amino acid 163 resulted in the formation of mutant capsids that were defective in DNA synthesis. Core proteins with further deletion to amino acid 144, although defective for RNA packaging, formed capsid particles, as shown by sucrose gradient analysis (3). In our present study, all of the capsid-forming deletion mutants ex-cept Cd148 coimmunoprecipitated with the TP and RT do-mains of Pol. Further deletion of core to amino acids 135 and 112 resulted in a failure to form capsid particles and an inabil-ity to coprecipitate with TP or RT. The results suggest that although required, capsid formation is not sufficient for core-Pol interactions.
[image:9.612.97.250.69.441.2]The formation of capsid-like particles was again revealed to be necessary for interaction with Pol in the analysis of core insertion mutants. Ci11, Ci36, Ci126, and Ci131 are the four core insertion mutants that do not readily form capsids, and each of them failed to coprecipitate with TP and RT. The capsid-forming mutant Ci44 coprecipitated with TP but failed to coprecipitate with RT. These results indicate that the TP and RT domains of Pol have different requirements for inter-acting with core. The differences in binding activities between Ci44 and Ci46 suggest that subtle differences in core influence RT recognition. Previous studies that analyzed capsid forma-tion in these mutants also examined the influence of the inser-tions on RNA encapsidation and genome replication (4). The core mutants Ci44 and Cp96 encapsidate WT levels of pre-genomic RNA and support reduced levels of both plus-strand and minus-strand DNA synthesis. The core insertion mutants Ci46 and to a small degree Ci69 encapsidate low levels of pregenomic RNA, and the core mutants Ci67 and Ci89 are FIG. 13. Core interacts with the RNase H domain. Insect cells were
coin-fected with baculovirus constructs expressing FRNH and WT core. The negative controls are a single infection with baculoviruses expressing FRNH (FRNH Control) and a single infection with baculoviruses expressing WT core (Core Control). (A) A fraction (1/20) of the cell lysate was immunoblotted with anti-core antibodies to detect anti-core protein expression. Immunoprecipitation (IP) was performed with anti-FLAG antibodies followed by Western blot analysis with anti-FLAG antibodies (B) to indicate immunoprecipitation of FRNH and with anticore antibodies (C) to demonstrate coprecipitation of core. Core interacts with a minimal RNase H domain extending from amino acids 680 to 780.
on November 9, 2019 by guest
http://jvi.asm.org/
deficient in encapsidation (4). All of these capsid-forming core mutants interact with TP, and all but Ci44 interact with RT. These results indicate that meeting the requirement for at least one core-Pol interactive site is not sufficient for encapsidation and certainly does not guarantee the formation of a replica-tion-competent particle. The results suggest that core-Pol as-sociations are essential for encapsidation and genomic repli-cation, yet the assembly of a replication-competent particle is considerably complex, and the requirement for multiple inter-actions is evident.
ACKNOWLEDGMENT
This work was supported by grant CA53246 from the National In-stitutes of Health.
REFERENCES
1.Bartenschlager, R., M. Junker-Niepmann, and H. Schaller.1990. The P gene product of hepatitis B virus is required as a structural component for genomic RNA encapsidation. J. Virol.64:5324–5332.
2.Bartenschlager, R., and H. Schaller.1992. Hepadnaviral assembly is initi-ated by polymerase binding to the encapsidation signal in the viral RNA genome. EMBO J.11:3413–3420.
3.Beames, B., and R. E. Lanford.1993. Carboxy-terminal truncations of the HBV core protein affect capsid formation and size of the encapsidated HBV RNA. Virology194:597–607.
4.Beames, B., and R. E. Lanford.1995. Insertions within the hepatitis B virus capsid protein influence capsid formation and RNA encapsidation. J. Virol.
69:6833–6838.
5.Chen, Y., W. S. Robinson, and P. L. Marion.1992. Naturally occurring point mutation in the C terminus of the polymerase gene prevents duck hepatitis B virus RNA packaging. J. Virol.66:1282–1287.
6.Chen, Y., W. S. Robinson, and P. L. Marion.1994. Selected mutations of the duck hepatitis B virus P gene RNase H domain affect both RNA packaging and priming of minus-strand DNA synthesis. J. Virol.68:5232–5238. 7.Cohen, B. J., and J. E. Richmond.1982. Electron microscopy of hepatitis B
core antigen synthesized inE. coli. Nature296:677–678.
8.Condreay, L. D., T.-T. Wu, C. E. Aldrich, M. A. Delaney, J. Summers, C. Seeger, and W. S. Mason.1992. Replication of DHBV genomes with muta-tions at the sites of initiation of minus- and plus-strand DNA synthesis. Virology188:208–216.
9.Galibert, F., E. Mandart, F. Fitoussi, P. Tiollais, and P. Charnay.1979. Nucleotide sequence of the hepatitis B virus genome (subtype ayw) cloned in
E. coli. Nature281:646–650.
10. Gallina, A., F. Bonelli, L. Zentilin, G. Rindi, M. Muttini, and G. Milanesi.
1989. A recombinant hepatitis B core antigen polypeptide with the prota-mine-like domain deleted self-assembles into capsid particles but fails to bind nucleic acids. J. Virol.63:4645–4652.
11. Ganem, D.1996. Hepadnaviridae and their replication, p. 2703–2737.In
B. N. Fields (ed.), Virology. Lippincott-Raven, Philadelphia, Pa. 12. Hatton, T., S. Zhou, and D. N. Standring.1992. RNA- and DNA-binding
activities in hepatitis B virus capsid protein: a model for their roles in viral replication. J. Virol.66:5232–5241.
13. Hirsch, R. C., J. E. Lavine, L.-J. Chang, H. E. Varmus, and D. Ganem.1990. Polymerase gene products of hepatitis B viruses are required for genomic RNA packaging as well as for reverse transcription. Nature344:552–555. 14. Hirsch, R. C., D. D. Loeb, J. R. Pollack, and D. Ganem.1991.cis-acting
sequences required for encapsidation of duck hepatitis B virus pregenomic RNA. J. Virol.65:3309–3316.
15. Hu, J. M., D. O. Toft, and C. Seeger.1997. Hepadnavirus assembly and reverse transcription require a multi-component chaperone complex which is incorporated into nucleocapsids. EMBO J.16:59–68.
16. Junker-Niepmann, M., R. Bartenschlager, and H. Schaller.1990. A short
cis-acting sequence is required for hepatitis B virus pregenome encapsida-tion and sufficient for packaging of foreign RNA. EMBO J.9:3389–3396. 17. Kann, M., and W. H. Gerlich.1994. Effect of core protein phosphorylation
by protein kinase C on encapsidation of RNA within core particles of hep-atitis B virus. J. Virol.68:7993–8000.
18. Knaus, T., and M. Nassal.1993. The encapsidation signal on the hepatitis B virus RNA pregenome forms a stem-loop structure that is critical for its function. Nucleic Acids Res.21:3967–3975.
19. Laemmli, U. K.1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature227:680–685.
20. Lanford, R. E., and J. S. Butel.1979. Antigenic relationship of SV40 early proteins to purified large T polypeptide. Virology97:295–306.
21. Lanford, R. E., K. D. Carey, L. E. Estlack, G. C. Smith, and R. V. Hay.1989. Analysis of plasma protein and lipoprotein synthesis in long-term primary cultures of baboon hepatocytes maintained in serum-free medium. In Vitro Cell. Dev. Biol.25:174–182.
22. Lanford, R. E, Y. H. Kim, H. Lee, L. Notvall, and B. Beames.1999. Mapping of the hepatitis B virus reverse transcriptase TP and RT domains by transcomplementation for nucleotide priming and by protein-protein inter-action. J. Virol.73:1885–1893.
23. Lanford, R. E., and L. Notvall.1990. Expression of hepatitis B virus core and precore antigens in insect cells and characterization of a core-associated kinase activity. Virology176:222–233.
24. Lanford, R. E., L. Notvall, and B. Beames.1995. Nucleotide priming and reverse transcriptase activity of hepatitis B virus polymerase expressed in insect cells. J. Virol.69:4431–4439.
25. Lanford, R. E., L. Notvall, H. Lee, and B. Beames.1997.
Transcomplemen-FIG. 14. Map summarizing core binding sites on Pol. FPL-pol depicts the structure of the WT Pol construct; core binding sites on amino- and carboxy-terminal deletion mutants of TP and RT are illustrated below the diagram. The shaded areas represent the minimal binding domains for each series. TPn, amino-terminal deletions of TP; TPc, carboxy-terminal deletions of TP. The TP deletions share a common area from amino acids 60 to 100, indicating the mapping of a common binding site or possibly two binding sites, one in the amino-terminal domain and one in the carboxy-terminal domain. RTn, amino-terminal deletions of RT; RTc, carboxy-terminal deletions of RT. The RT deletions share a common area from amino acids 400 to 550, which again indicates the mapping of either a common core binding site or at least two separate binding sites. RNH, carboxy-truncated RNase H domain. Core binds to a minimal RNase H domain spanning from amino acids 680 to 780.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:10.612.60.541.74.275.2]tation of nucleotide priming and reverse transcription between indepen-dently expressed TP and RT domains of the hepatitis B virus reverse tran-scriptase. J. Virol.71:2996–3004.
26. Lien, J., D. J. Petcu, C. E. Aldrich, and W. S. Mason.1987. Initiation and termination of duck hepatitis B virus DNA synthesis during virus maturation. J. Virol.61:3832–3840.
27. Matsuda, K., S. Satoh, and H. Ohori.1988. DNA-binding activity of hepatitis B e antigen polypeptide lacking the protaminelike sequence of nucleocapsid protein of human hepatitis B virus. J. Virol.62:3517–3521.
28. Miyanohara, A., T. Imamura, M. Araki, K. Sugawara, N. Ohtomo, and K. Matsubara.1986. Expression of hepatitis B virus core antigen gene in Sac-charomyces cerevisiae: synthesis of two polypeptides translated from different initiation codons. J. Virol.59:176–180.
29. Molnar-Kimber, K. L., J. W. Summers, and W. S. Mason.1984. Mapping of the cohesive overlap of duck hepatitis B virus DNA and of the site of initiation of reverse transcription. J. Virol.51:181–191.
30. Nakabayashi, H., K. Taketa, K. Miyano, T. Yamane, and J. Sato.1982. Growth of human hepatoma cells lines with differentiated functions in chem-ically defined medium. Cancer Res.42:3858–3863.
31. Nassal, M.1992. The arginine-rich domain of the hepatitis B virus core protein is required for pregenome encapsidation and productive viral posi-tive-strand DNA synthesis but not for virus assembly. J. Virol.66:4107–4116. 32. Nassal, M.1999. Hepatitis B virus replication: novel roles for virus-host
interactions. Intervirology42:100–116.
33. Petit, M.-A., and J. Pillot.1985. HBc and HBe antigenicity and DNA-binding activity of major core protein P22 in hepatitis B virus core particles isolated from the cytoplasm of human liver cells. J. Virol.53:543–551.
34. Pollack, J. R., and D. Ganem.1993. An RNA stem-loop structure directs hepatitis B virus genomic RNA encapsidation. J. Virol.67:3254–3263. 35. Schlicht, H.-J., R. Bartenschlager, and H. Schaller.1989. The duck hepatitis
B virus core protein contains a highly phosphorylated C terminus that is
essential for replication but not for RNA packaging. J. Virol.63:2995–3000. 36. Seeger, C., D. Ganem, and H. E. Varmus.1986. Biochemical and genetic evidence for the hepatitis B virus replication strategy. Science232:477–484. 37. Seeger, C., and J. Maragos.1990. Identification and characterization of the
woodchuck hepatitis virus origin of DNA replication. J. Virol.64:16–23. 38. Seeger, C., and J. Maragos.1991. Identification of a signal necessary for
initiation of reverse transcription of the hepadnavirus genome. J. Virol.
65:5190–5195.
39. Seeger, C., and W. S. Mason.2000. Hepatitis B virus biology. Microbiol. Mol. Biol. Rev.64:51–68.
40. Tavis, J. E., and D. Ganem.1996. Evidence for activation of the hepatitis B virus polymerase by binding of its RNA template. J. Virol.70:5741–5750. 41. Tavis, J. E., S. Perri, and D. Ganem.1994. Hepadnavirus reverse
transcrip-tion initiates within the stem-loop of the RNA packaging signal and employs a novel strand transfer. J. Virol.68:3536–3543.
42. Wang, G.-H., and C. Seeger.1993. Novel mechanism for reverse transcrip-tion in hepatitis B viruses. J. Virol.67:6507–6512.
43. Ward, G. A., C. K. Stover, B. Moss, and T. R. Fuerst.1995. Stringent chemical and thermal regulation of recombinant gene expression by vaccinia vectors in mammalian cells. Proc. Natl. Acad. Sci. USA92:6773–6777. 44. Will, H., W. Reiser, T. Weimer, E. Pfaff, M. Bu¨scher, R. Sprengel, R.
Cat-taneo, and H. Schaller.1987. Replication strategy of human hepatitis B virus. J. Virol.61:904–911.
45. Yu, M., and J. Summers.1991. A domain of the hepadnavirus capsid protein is specifically required for DNA maturation and virus assembly. J. Virol.
65:2511–2517.
46. Yu, M., and J. Summers.1994. Phosphorylation of the duck hepatitis B virus capsid protein associated with conformational changes in the C terminus. J. Virol.68:2965–2969.
47. Zhou, S., and D. N. Standring.1991. Production of hepatitis B virus nucleo-capsidlike core particles inXenopusoocytes: assembly occurs mainly in the cytoplasm and does not require the nucleus. J. Virol.65:5457–5464.