Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Neutralization Profiles of Primary Human Immunodeficiency Virus
Type 1 Isolates in the Context of Coreceptor Usage
D. CECILIA,
1VINEET N. KEWALRAMANI,
1JEANNE O’LEARY,
2BARBARA VOLSKY,
1PHILLIPE NYAMBI,
1SHERRI BURDA,
1SERENA XU,
1† DAN R. LITTMAN,
1,3AND
SUSAN ZOLLA-PAZNER
1,2*
New York University Medical Center,
1New York Veterans Affairs Medical Center,
2and
Howard Hughes Medical Institute,
3New York, New York
Received 10 March 1998/Accepted 22 May 1998
Most strains of human immunodeficiency virus type 1 (HIV-1) which have only been carried in vitro in
peripheral blood mononuclear cells (primary isolates) can be neutralized by antibodies, but their sensitivity to
neutralization varies considerably. To study the parameters that contribute to the differential neutralization
sensitivity of primary HIV-1 isolates, we developed a neutralization assay with a panel of genetically engineered
cell lines (GHOST cells) that express CD4, one of eight chemokine receptors which function as HIV-1
coreceptors, and a Tat-dependent green fluorescent protein reporter cassette which permits the evaluation and
quantitation of HIV-1 infection by flow cytometry. All 21 primary isolates from several clades could grow in the
various GHOST cell lines, and their use of one or more coreceptors could easily be defined by flow cytometric
analysis. Ten of these primary isolates, three that were CXCR4 (X4)-tropic, three that were CCR5 (R5)-tropic,
and four that were dual- or polytropic were chosen for study of their sensitivity to neutralization by human
monoclonal and polyclonal antibodies. Viruses from the X4-tropic category of viruses were first tested since
they have generally been considered to be particularly neutralization sensitive. It was found that the X4-tropic
virus group contained both neutralization-sensitive and neutralization-resistant viruses. Similar results were
obtained with R5-tropic viruses and with dual- or polytropic viruses. Within each category of viruses,
neu-tralization sensitivity and resistance could be observed. Therefore, sensitivity to neuneu-tralization appears to be
the consequence of factors that influence the antibody-virus interaction and its sequelae rather than coreceptor
usage. Neutralization of various viruses by the V3-specific monoclonal antibody, 447-52D, was shown to be
dependent not only on the presence of the relevant epitope but also on its presentation. An epitope within the
envelope of a particular virus is not sufficient to render a virus sensitive to neutralization by an antibody that
recognizes that epitope. Moreover, conformation-dependent factors may overcome the need for absolute fidelity
in the match between an antibody and its core epitope, permitting sufficient affinity between the viral envelope
protein and the antibody to neutralize the virus. The studies indicate that the neutralization sensitivity of
HIV-1 primary isolates is a consequence of the complex interaction between virus, antibody, and target cell.
The sensitivity of human immunodeficiency virus type 1
(HIV-1) strains to neutralization depends on several factors.
For example, the level of intercellular cell adhesion molecule
type 1 (ICAM-1) on a virus particle affects the sensitivity with
which it can be neutralized by antibody (15, 40). Sawyer et al.
(43), using laboratory-adapted and primary isolates, showed
that the host cells used for growing the virus stock influenced
the sensitivity of the virus to neutralization and that the type of
target cells used in the neutralization assay, i.e., T-cell lines or
unstimulated or phytohemagglutinin (PHA)-activated
periph-eral blood mononuclear cells (PBMCs), also contributes to the
sensitivities with which neutralization of HIV and other viruses
is detected (34, 53, 54).
The isolates that have been adapted to T-cell lines (TCLA
strains) have frequently been described as neutralization
sen-sitive. However, data show that there are TCLA strains which
are highly sensitive to neutralization, e.g., MN, and TCLA
strains that are relatively less so, e.g., RF (28).
A consensus concerning primary isolates suggests that they
are difficult to neutralize. However, many reports document
that there is a spectrum of neutralization sensitivity among
primary isolates just as there is among TCLA strains (19, 22,
38, 49, 52). There has also been a consensus that the
neutral-ization sensitivity of HIV isolates is linked to the phenotype of
isolates, that is, that syncytium-inducing (SI) or CXCR4-tropic
(X4) viruses (including all laboratory-adapted strains) are
more easily neutralized than non-syncytium-inducing (NSI) or
CCR5-tropic (R5) viruses (the phenotype of the majority of
primary isolates) (50). This is not supported by published data.
For instance, Hogervorst et al. (23) made chimeric LAI viruses
with the envelopes of an NSI or an SI isolate from the same
individual; both chimeric viruses, regardless of NSI or SI
phe-notype, were neutralized by a heterologous serum pool. With
the identification of the HIV coreceptors, CXCR4 and CCR5,
coreceptor usage was thought to play a role in the greater
sensitivity of TCLA strains to neutralization. However, it was
shown recently that whether a strain uses CXCR4 or CCR5, its
susceptibility to neutralization remains unchanged (27, 34, 45):
Trkola et al. (45) used CD4-blocking reagents and monoclonal
antibodies (MAbs) against dualtropic TCLA or primary
iso-lates and showed that neutralization was unaffected by the
coreceptor used. La Casse et al. (27) used V3-binding MAbs
against a primary isolate and the TCLA clone of the same
isolate and came to the same conclusion, as did Montefiori et
al., using polyclonal HIV-positive human sera (34).
* Corresponding author. Mailing address: New York VA Medical
Center, 423 East 23rd St., Room 18125N, New York, NY 10010.
Phone: (212) 263-6769. Fax: (212) 951-6321. E-mail: Zollas01@mcrcr6
.med.nyu.edu.
† Present address: Schering-Plough Research Institute, Kenilworth,
N.J.
6988
on November 9, 2019 by guest
http://jvi.asm.org/
To quantify the differential neutralization sensitivities of
pri-mary isolates, we developed a new assay which is subject to less
variability than previously described assays and used it to test
a broad panel of primary isolates for sensitivity to Ab-mediated
neutralization. “GHOST cells” which were described
previ-ously (24, 45), were used as the target cells in this assay. They
are human osteosarcoma cells (HOS) that express CD4 and
one of several HIV coreceptors. These cells also contain a gene
for green fluorescent protein (GFP) under the control of the
HIV-2 promoter, which, in the presence of Tat, acts as an
indicator of infection, generating a fluorescent cytoplasmic
sig-nal which can be detected and enumerated by flow cytometry.
The GHOST cells were found to be infectable by all of the
primary isolates tested, with the coreceptor preference
reflect-ing the tropism of each isolate. The ease with which infection
is detected by flow cytometry was used to advantage in
devel-oping a sensitive, reproducible, and convenient neutralization
assay which demonstrated that, within each category of viruses
defined by phenotype and coreceptor usage, there were
neu-tralization-sensitive and neutralization-resistant strains. Thus,
the sensitivity or resistance of a primary isolate to Ab-mediated
neutralization is a function of the virus particle and the effects
of its interaction with Ab, not a characteristic of any category
of viruses defined to date.
MATERIALS AND METHODS
Virus isolates.A total of 21 primary isolates which had been passaged exclu-sively in PBMCs were used. These included isolates SF33 and SF2, obtained from J. Levy, University of California at San Francisco, San Francisco, Calif.; isolates CA1, CA5, CA13, CA20, VI191, VI525, VI313, and MAI, obtained from G. van der Groen, Institute of Tropical Medicine, Antwerp, Belgium; isolates 92HT593, 92HT594, 91US056, 92RW021, BK131, SM993, 92TH080, JR-FL, and 89.6, supplied by the National Institutes of Health (NIH) AIDS Research and Ref-erence Reagent Program; and BZ167, supplied originally by J. Mascola, Walter Reed Army Institute of Research, Rockville, Md. MNp, a primary isolate of the MN strain, which had never been passaged in cell lines, was obtained from J. Sullivan, University of Massachusetts Medical School, Worcester, Mass. The clade designation and MT-2-defined phenotype of each primary isolate are shown in Table 1. All virus stocks were prepared by infecting PHA-activated
human PBMCs (54). Briefly, frozen PBMCs from HIV-1-negative blood donors were thawed, stimulated with PHA (3mg/ml; Difco Detroit, Mich.) for 3 days, centrifuged, and infected with 1 ml of virus-infected culture supernatant. After a 1-h exposure of the cells to the virus, the volume of the cell suspension was adjusted to a concentration of 23106cells/ml and the culture was maintained
in RPMI 1640 medium with 10% fetal bovine serum and interleukin-2 (IL-2) (20 U/ml; Boehringer Mannheim Biochemicals, Indianapolis, Ind.) at the same cell concentration. The concentration of p24 in the infected culture supernatant was checked every 3 to 4 days by a noncommercial enzyme-linked immunosorbent assay (26). The infected culture supernatant was collected at 1 to 2 weeks postinfection when the concentration of p24 was at least 100 ng/ml.
Cells.The GHOST cell lines used herein were derived from HOS cells (24). Briefly, HOS cells were transduced with the human CD4 gene encoded by the murine leukemia virus retroviral vector, pMV7neo, to generate a CD4-positive HOS.T4 clone. HOS.T4 cells were subsequently stably cotransfected with a reporter construct consisting of the HIV-2 long terminal repeat directing the expression of humanized green fluorescent protein (GFP) and a selection con-struct composed of the human cytomegalovirus immediate-early (IE) promoter driving the expression of hygromycin phosphotransferase. Cells stably trans-fected with the GFP reporter construct were checked for sensitivity to HIV-1 Tat-mediated gene activation. One clone; clone 34, which expressed GFP strongly after Tat transactivation, was designated the parental cell line and chosen for further development. GHOST cl.34 parental cells were transduced with one of the chemokine receptors (CCR1, CCR2, CCR3, CCR5, CXCR4, Bonzo/STRL33, or BOB/gpr15) encoded on the murine leukemia virus vector, pBABEpuro. Cells expressing the CCR5 coreceptor are referred to below as GHOST-R5, those expressing the CXCR4 coreceptor are referred to as GHOST-X4, etc. About 40% of the cells from each of the GHOST cell lines were positive for CD4; 84% of the GHOST-X4 cells were positive for CXCR4; 66% of the GHOST-R5 cells were positive for CCR5. The parent GHOST cells were 3.8% positive for CXCR4 and 0.8% positive for CCR5.
GHOST cells bearing chemokine receptors were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum, 1% glu-tamine, 2% penicillin plus streptomycin, Geneticin (200mg/ml), hygromycin (25
mg/ml), and puromycin (1mg/ml). The cultures were maintained at 37°C in a 5% CO2humidified incubator. Cell monolayers, when confluent, were resuspended
by using 0.25% trypsin. The cells were maintained for up to 15 passages and then replaced with fresh cells from the cryopreserved stock which had been frozen at the second or third passage.
MAbs and polyclonal Abs.Five polyclonal serum samples and two MAbs were used in the neutralization tests. Sera F and N are from asymptomatic HIV-positive volunteers from the Veterans Affairs Medical Center (New York, N.Y.) and the University of California Los Angeles Medical Center (Los Angeles, Calif.), respectively. Serum N was provided by S. Miles (University of California Los Angeles Medical Center, Los Angeles, Calif.). Serum FDA-2 is a serum pool derived from four bleeds from an HIV-positive patient obtained from the NIH AIDS Research and Reference Reagent Program. HIVIG-Ug is the immuno-globulin G (IgG) fraction from pooled HIV-positive serum obtained in Uganda (supplied by B. Jackson, Johns Hopkins University, Baltimore, Md.); it was used at a starting concentration of 0.5 mg/ml. Pool 2 is a serum pool from 33 randomly selected HIV-positive subjects at the Veterans Affairs Medical Center, New York. An HIV-negative human serum specimen was used in the experiments as a negative control. All sera were heat inactivated at 56°C for 30 min prior to use. Two human MAbs were used in this study, 447-52D and IgG1b12. 447-52D, produced in this laboratory, was previously described as a broadly cross-reactive V3-specific MAb (18, 19). IgG1b12, a recombinant antibody which recognizes an epitope overlapping the CD4 binding domain of the HIV-1 envelope (8), was obtained from the NIH AIDS Research and Reference Reagent Program. Stocks of both MAbs were stored in frozen aliquots as purified IgG (1 mg/ml) and used at concentrations ranging from 0.1 to 25mg/ml.
Infectivity assay.GHOST cells were seeded in 24-well plates (Falcon; Fisher Scientific, Springfield, N.J.) at 63104cells/well/0.5 ml. On the following day, the
medium was removed and the monolayers, about 70% confluent, were infected with undiluted virus stocks (100ml/well). To each well was added DEAE-dextran to a final concentration of 8mg/ml. The virus was allowed to adsorb overnight, after which the virus-containing medium was removed and the cell monolayers were washed once with phosphate-buffered saline. Subsequently, 1 ml of com-plete medium, as described above, was added per well. The day on which the virus was added was considered day 0. Cells were harvested on day 4 or 5 postinfection (p.i.). On the day of harvest, the cell monolayers were once again washed with phosphate-buffered saline, resuspended in 300ml of 1 mM EDTA in PBS, and fixed in formaldehyde at a final concentration of 2%. The cells were then analyzed with a FACScan flow cytometer (Becton Dickenson, San Jose, Calif.). The live cells were gated on the basis of forward and side scatter. Because of the autofluorescence of uninfected GHOST cells due to basal expression of the indicator cassette, the gain on the FL 1 channel was set to bring the mean channel fluorescence of uninfected cells to,102. The number of infected cells
was determined by using a scattergram of fluorescence versus forward scatter after setting the gates with uninfected cells. A total of 15,000 to 20,000 events was scored. The total number of cells was about 106/well on the day of harvest. Hence
[image:2.612.50.291.89.309.2]the number of cells scored was approximately 1/50 of the total cells in culture. As noted above, 3.8% of the GHOST parental cells are CXCR4 positive, and hence
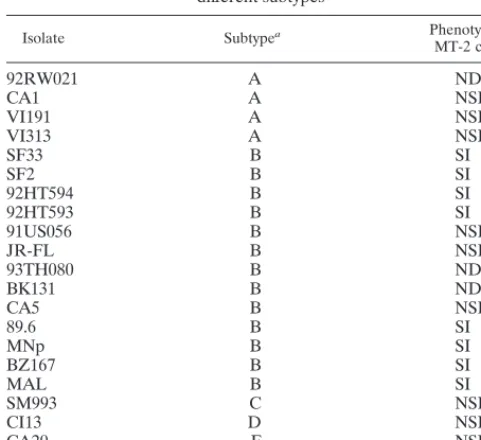
TABLE 1. Syncytium-inducing phenotype of primary isolates from
different subtypes
Isolate Subtypea Phenotype in
MT-2 cellsa
92RW021
A
ND
bCA1
A
NSI
VI191
A
NSI
VI313
A
NSI
SF33
B
SI
SF2
B
SI
92HT594
B
SI
92HT593
B
SI
91US056
B
NSI
JR-FL
B
NSI
93TH080
B
ND
BK131
B
ND
CA5
B
NSI
89.6
B
SI
MNp
B
SI
BZ167
B
SI
MAL
B
SI
SM993
C
NSI
CI13
D
NSI
CA20
F
NSI
VI525
G
SI
aThe subtypes and phenotypes of each virus are listed as reported in the AIDS Research and Reference Reagent Program Catalog or in reference 37.
bND, not done.
on November 9, 2019 by guest
http://jvi.asm.org/
all the coreceptor derivatives express a low but detectable level of CXCR4. To account for this, GHOST-CCR1 cells were used to establish background infect-ability. The mean number of fluorescent GHOST-CCR1 cells12 standard deviations after infection with the viruses used in this study was considered the cutoff. On this basis,.99 fluorescent cells/15,000 cells had to be present for a virus to be considered positive for infectivity in each of the GHOST cell lines tested.
GHOST cell neutralization assay.The method for the GHOST cell neutral-ization assay was essentially as described above for infectivity studies except that a fixed dilution of each virus stock was used, based on predetermined infectivity titers; the virus dilution used was chosen to give about 200 to 800 fluorescent cells per 15,000 events in the absence of anti-HIV antibodies. Polyclonal Ab or MAb preparations were diluted serially in fivefold dilutions. Equal volumes of diluted Ab preparations and virus were mixed and incubated for 1 h at 37°C; 100ml of the virus-Ab mixture was added to duplicate wells and incubated overnight in the presence of DEAE-dextran (8mg/ml). Subsequent washes, incubations, harvest, and readout procedures were performed as described above. Neutralization assays were typically terminated 3 to 4 days p.i. Care was taken to terminate the assay before cell lysis occurred. Formation of moderate-sized syncytia did not seem to affect the flow analysis, since the forward and side scatter gates included almost all of the viable cells.
RESULTS
Determination of coreceptor preferences by primary isolates
from different clades.
A panel of 21 primary isolates belonging
to different clades and demonstrating different SI/NSI
pheno-types when grown in MT-2 cells was chosen for infectivity
studies (Table 1). GHOST cells expressing CCR1, CCR2,
CCR3, CCR5, CXCR4, BOB, or Bonzo were infected with
virus stocks in the presence of DEAE-dextran since this
com-pound was required for optimal infectivity by some viruses. For
example, while isolate SF33 was not significantly affected by
the presence of DEAE-dextran, isolate CA5 showed infectivity
only in its presence (data not shown). Thus, for all infectivity
and neutralization assays, DEAE-dextran was included during
virus adsorption.
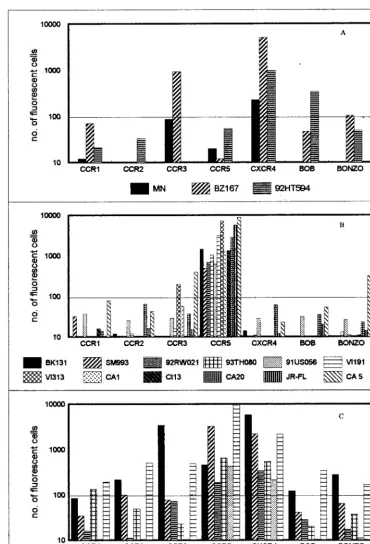
Infection by primary isolates of GHOST cells expressing the
various chemokine receptors was readily detected by flow
cy-tometry, as depicted in Fig. 1. Of nine SI viruses tested, only
three, BZ167, MNp and 92HT594, were X4, using CXCR4 but
not CCR5. However, BZ167 could also use CCR3, while
92HT594 also used BOB (Fig. 1A). The other six SI viruses
were dualtropic, using both CXCR4 and CCR5, or polytropic,
using both major coreceptors and CCR2, CCR3, and/or
Bonzo; these three coreceptors generally mediated infection
with lower efficiencies than did CXCR4 and CCR5 (Fig. 1C).
The 12 NSI viruses tested were R5, defined as using CCR5 but
not CXCR4. Viruses CA5 and VI313 also used CCR3 and
Bonzo, respectively (Fig. 1B). Again, these two coreceptors
mediated infection with lower efficiency.
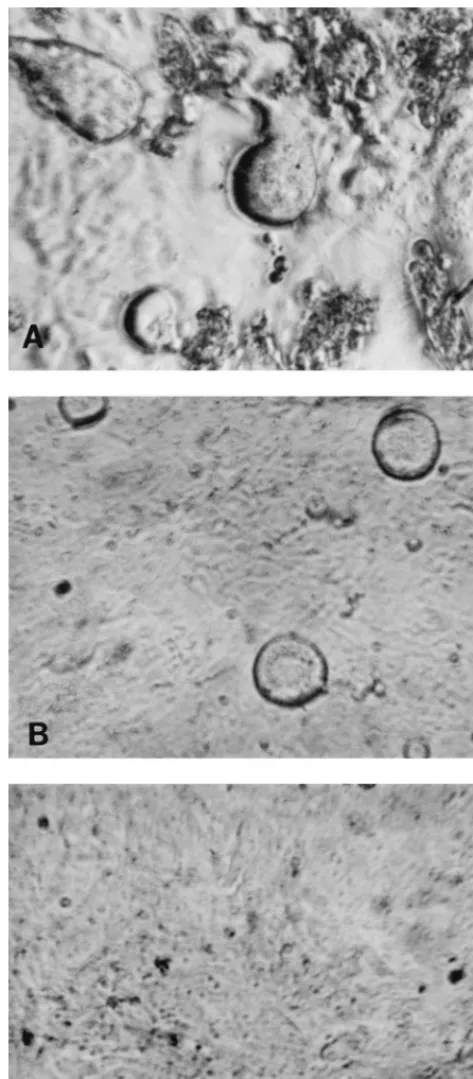
All the isolates, irrespective of coreceptor usage or SI/NSI
phenotype, caused syncytium formation in the GHOST cells.
For the majority of R5 viruses, the cytopathic effect was
ap-parent early compared to that seen with viruses which used
other coreceptors. Figures 2A and B show syncytium formation
in GHOST cells by the R5 virus VI313 and by the X4/R5 virus
89.6, respectively, on day 3 p.i. Uninfected cells are shown in
Fig. 2C.
Infection kinetics.
The kinetics of infection for a polytropic
virus (SF33) and a dualtropic virus (92HT593), as assessed
fluorocytometrically, are shown in Fig. 3A for GHOST-X4 and
GHOST-R5 cells. SF33 showed a lower efficiency of infection
in GHOST-R5 cells compared to that in GHOST-X4 cells,
although the rates of infection in the two cell lines were
par-allel. Isolate 92HT593 showed similar kinetics in the two cell
lines.
Ab-mediated neutralization of primary isolate infection of
GHOST cells.
The neutralization assay was first performed
with the polytropic clade B SF33 isolate in GHOST-X4 and
GHOST-R5 cells. The positive serum pool 2 and an
HIV-negative serum specimen were used at a final dilution of 1:40.
The cells were harvested from days 2 to 6 p.i. The number of
fluorescing GHOST-X4 or GHOST-R5 cells in wells which
were infected with virus treated with normal serum was about
700 on day 6 (Fig. 3B and C). This was reduced to background
levels throughout the observation period when virus was
pretreated with the anti-HIV pool 2 (Fig. 3B and C). The
day of harvest did not affect the degree of neutralization
observed.
A checkerboard neutralization assay was also carried out
with SF33 and MAb 447-52D or the HIV-positive serum F.
Varying the virus input by 1 order of magnitude, resulting in
170 to 1,165 infected cells/15,000 cells in the absence of
anti-body, did not influence the neutralization titers observed in the
assay (data not shown).
Subsequently, 10 primary isolates from clade B with different
coreceptor preferences were tested in the GHOST cell
neu-tralization assay against individual positive sera, the
HIV-positive serum pool, HIVIG-Ug, and two human MAbs. Each
Ab-virus combination was tested in duplicate, and each assay
was repeated at least twice. There was very little variability in
the duplicates, with the variation ranging from 0.3 to 10%.
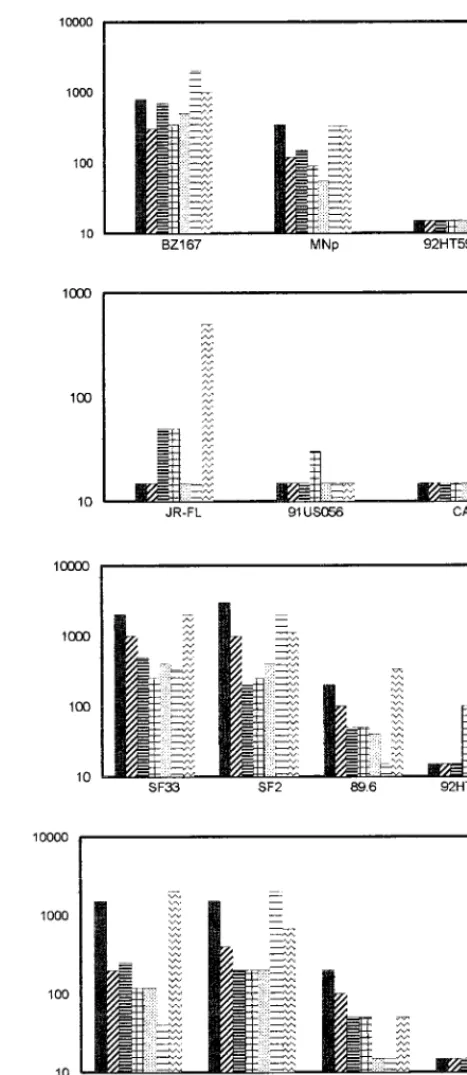
Neutralization curves are shown in Fig. 4 for the polytropic
primary isolate SF2 with three individual immune sera in both
GHOST-X4 and GHOST-R5 cells. Similar neutralization
curves were used to determine the 50% neutralization titers,
which are shown in Fig. 5.
Of the X4 viruses, BZ167 was neutralized best, with 50%
serum neutralizing titers of 1:500 to 1:1,000 and 50%
neutral-izing MAb concentrations of 1 to 5
m
g/ml (Fig. 5A). The
primary isolate MN was also neutralized by all the Ab
prepa-rations but with lower titers. 92HT594 could not be neutralized
by any of the Ab preparations tested; the lowest dilution of
antiserum tested was 1:40 and the highest concentration of
MAb tested was 25
m
g/ml for 447-52D and 13.5
m
g/ml for
IgG1b12 (Fig. 5A).
Of the R5 viruses, JR-FL could be neutralized by sera N and
FDA-2 at titers of 1:50 and by MAb IgG1b12 at 2
m
g/ml. Only
44% neutralization was achieved with isolate 91USO56 and
FDA-2 at a 1:40 dilution. Isolate CA5 could not be neutralized
by any of the Ab preparations tested (Fig. 5B).
With dual- and polytropic viruses, the patterns of
neutral-ization were essentially the same whether the viruses were
assayed on GHOST-X4 or GHOST-R5 cells (Fig. 5C and D,
respectively). Only SF33 showed slightly lower titers when
tested on GHOST-R5 (compared to GHOST-X4) with pool
2, the three individual HIV-positive sera, and the anti-V3
MAb, 447-52D. SF33 (in GHOST-X4) and SF2 could be
neutralized best, at serum titers and MAb concentrations of
;
1:1,000 and 1 to 2
m
g/ml, respectively. Isolate 89.6 showed
moderate neutralization, and 92HT593 showed poor
neu-tralization.
Thus, in each of the different virus categories defined by
coreceptor usage, there was a spectrum of differential
suscep-tibilities to Ab-mediated neutralization. R5 viruses appeared
to be somewhat more resistant than X4, dual-, or polytropic
viruses, but this conclusion may be affected by the particular
viruses used from the panel available for testing.
As shown in Fig. 5, neither 447-52D nor IgG1b12
neutral-ized all the isolates. MAb 447-52D neutralneutral-ized 4 of 10 strains,
and IgG1b12 neutralized 7 of 10 strains. When 50%
neutral-izing concentrations were achieved for MAb 447-52D, they
ranged from
,
1 to 20
m
g/ml; the range for IgG1b12 ranged
from
,
0.5 to 10
m
g/ml. The core epitope of MAb IgG1b12 has
not been identified, and therefore it was not possible to
on November 9, 2019 by guest
http://jvi.asm.org/
relate its activity with any particular virus sequence. The core
epitope of MAb 447-52D was previously identified as GPXR
by Pepscan analysis with overlapping peptides from the V3
sequence of the MN strain (18). Binding to the V3
MNpeptide,
however, accounts for only about 10% of the binding energy of
[image:4.612.116.486.69.613.2]the MAb (18); while neutralizing activity was associated with
this V3 sequence (12), it was also correlated with the
dissoci-ation rate constant (48), which is affected by both sequence and
conformation. This information suggested that the sequence
at the crown of the V3 loop might not be sufficient to confer
FIG. 1. Twenty-one primary isolates were tested for their coreceptor preference in GHOST cells expressing CCR1, CCR2, CCR3, CCR5, CXCR4, BOB, or Bonzo. The virus isolates were either SI (A), NSI (B), or dual- or polytropic (C). One representative experiment of two or three is shown, with values being the mean of duplicate observations. The mean number of fluorescent cells observed in GHOST-CCR1 cells with all 21 viruses12 standard deviations was the cutoff value (horizontal line).on November 9, 2019 by guest
http://jvi.asm.org/
Ab binding that would lead to neutralization and that under
certain conditions, conformation might overcome the need
for absolute fidelity in the core epitope. To examine this,
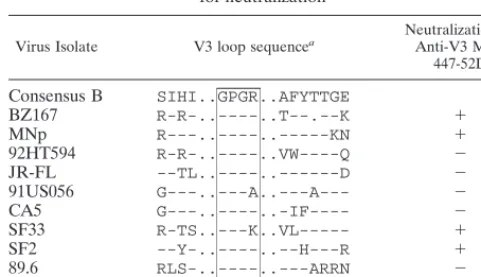
partial V3 sequences of the 10 isolates used in the
neutral-ization experiments were analyzed; they are shown in Table
2. Of the 10 viruses, 8 contain the GPXR core epitope, but
only 3 of these (BZ167, MNp, and SF2) were neutralized by
MAb 447-52D. This suggests that the presence of the core
epitope is not sufficient to confer neutralization sensitivity.
Conversely, one of the two isolates that does not contain the
core epitope (SF33) was neutralized by MAb 447-52D,
sug-gesting that conformation-dependent structures can confer
sufficient binding energy to effect neutralization.
DISCUSSION
[image:5.612.310.543.60.523.2]We have shown that within each class of HIV-1 primary
isolates categorized by coreceptor usage, there is a spectrum of
neutralization sensitivity. The neutralization sensitivity of the
isolates was determined by a new assay developed with
genet-ically engineered GHOST cell lines which express CD4 and
one of several chemokine receptors known to function as HIV
FIG. 2. Syncytia seen in GHOST-C5 cells infected with NSI virus VI313 (A)or with dualtropic virus 89.6 (B). (C) Uninfected cells.
FIG. 3. (A) Kinetics of infection of SF33 in GHOST-X4 (■) and GHOST-R5 (h) cells and of 92HT593 in GHOST-X4 (F) and GHOST-R5 (E) cells. (B and C) Effect of pretreatment with HIV immune serum pool 2 (Œ) and normal serum (■) on the infection kinetics of SF33 in GHOST-X4 (B) and GHOST-R5 (C) cells. Sera were tested at a final dilution of 1:40.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.52.289.71.610.2]coreceptors. The use of the GHOST cell lines as target cells in
this neutralization assay substantially reduces the variability
inherent in PBMC-based neutralization assays, which is a
re-sult of donor variation. The GHOST cell assay is also
advan-tageous because it measures the number of infected cells
di-rectly, in contrast to the measurement of p24 or reverse
transcriptase in the PBMC neutralization assay, which provides
only an indirect assessment of the level of infection.
The applicability of the GHOST cells to the development of
a neutralization assay useful with a broad range of primary
HIV-1 isolates was established by determining the
susceptibil-ity of the cells to infection with 21 different primary isolates.
Simultaneously, the range of coreceptor usage was also
deter-mined for these isolates. Coreceptor usage was easily discerned
for each virus by simply measuring the induced fluorescence of
the GFP reporter gene upon infection of the GHOST cells
carrying the coreceptor used by the virus being studied. Until
now, identification of coreceptor usage by primary isolates has
depended on quantitation of p24 or syncytium formation with
cell lines expressing one of the coreceptors (3, 45). In addition
to CXCR4 and CCR5, CCR3 was used by several viruses, but
it was used at a slightly lower efficiency. CCR3 is present on a
wide range of cells (21, 42, 47), and the ability to use CCR3
may be relevant to the progression of disease. Other
corecep-tors, e.g., BOB and Bonzo, were similarly used by a minority of
viruses and at relatively low efficiency.
Conflicting conclusions about the sensitivity of primary
iso-lates to neutralization have been drawn. Many groups have
shown that patients’ sera display comparable neutralization
titers with laboratory-adapted strains and primary isolates (1,
28, 43–45). Several other studies, however reported a
differ-ence in neutralization sensitivities between laboratory-adapted
and primary isolates, based on results with sera from vaccinees
(29), HIV-positive sera (29, 34), and MAbs (14). Since TCLA
isolates are SI and a majority of primary isolates are NSI (50),
the data suggesting a greater neutralization sensitivity for
lab-oratory-adapted strains has inaccurately been transformed into
a consensus that SI isolates are more sensitive to neutralization
than are NSI isolates. Our studies show that there is a
spec-trum of sensitivities within each virus phenotype to
Ab-medi-ated neutralization and that some SI (X4) primary isolates are
difficult to neutralize while some NSI (R5) isolates are
neu-tralization sensitive, indicating that phenotype has little to do
with virus sensitivity to neutralization.
[image:6.612.61.281.70.185.2] [image:6.612.312.546.81.619.2]Compared to other viruses, serum neutralization titers to
HIV appear to be low (32). One of the reasons for this
obser-vation could be a lower sensitivity of the neutralization assays
FIG. 4. Neutralization of SF2 by three immune sera (N, F, and FDA-2) in GHOST-X4 (bold lines) and GHOST-R5 (dashed lines) cells. Each serum dilu-tion was tested in duplicate, and the percent neutralizadilu-tion was calculated by using the mean. The dose-response curves obtained are from one representative experiment of two or three carried out with each serum sample.FIG. 5. The 50% neutralizing titers shown on the y axis were determined for 10 primary isolates with seven antibody preparations. The X4-tropic viruses were tested on GHOST-X4 cells (A), the R5-tropic viruses were tested on GHOST-R5 cells (B), and the polytropic viruses were tested on GHOST-X4 (C) or GHOST-R5 (D) cells. Fivefold dilutions of the Ab preparations were tested for neutralization against a fixed dilution of virus. The stocks of the MAb preparations were adjusted to 1 mg/ml; thus, a titer of 1:1,000 is equivalent to 1
mg/ml. HIVIG-Ug was used at a starting concentration of 0.5 mg/ml; thus, an HIVIG-Ug titer of 1:1,000 corresponds to 0.5mg/ml. Antibody preparations that did not neutralize an isolate at the lowest dilution tested (1:40) are shown graphically with an arbitrary titer of 1:15, which should be considered negative.
on November 9, 2019 by guest
http://jvi.asm.org/
currently used for HIV. Multiple approaches to developing a
sensitive and convenient neutralization assay have been tried.
Back et al. (4) developed a transfection-neutralization assay in
which CD4-negative cells transfected with proviral DNA of
molecular clones were cocultured with PBMCs. Candotti et al.
(9) described an assay in which, instead of quantitation of p24,
HIV provirus synthesis was measured by PCR. The sensitivity
of detecting neutralization was not improved by these
tech-niques. Several assays that directly measure the reduction in
infectivity commonly use T-cell lines, which restricts the assays
to SI viruses. Most commonly, PBMCs are used as target cells
in measuring the neutralization of primary isolates, and
am-plification products such as p24 or RT are quantified for the
readout. The conditions for these PBMC-based assays vary
widely (reviewed in reference 51) and result in broad variations
in the neutralizing activity detected. In assays where virus is
exposed to unstimulated PBMCs (54), target cells include
CXCR4-expressing T cells and CCR5-expressing lymphocytes
and monocytes (7), permitting the assessment of neutralization
of both SI and NSI viruses. In neutralization assays with
PBMCs activated with PHA and maintained in IL-2, PHA
down-regulates CCR5 while IL-2 up-regulates CCR5 (7); thus,
the status of the target cells varies with respect to coreceptor
expression and depends on the particular conditions used for
this “conventional” assay system. An alternative system with
less variable target cells would therefore be highly desirable.
Potential target cells for this purpose include genetically
engi-neered cells expressing CD4, coreceptors, and an indicator
gene. Such cell lines, with one of several indicator genes
con-trolled by an HIV promoter, have been used to detect HIV
infection. The indicator genes that have been used include
chloramphenicol acetyltransferase (11),
b
-galactosidase (25),
and luciferase (13). Recently, secreted alkaline phosphatase
was used as the indicator gene in a neutralization assay in
which the output was measured as chemiluminescence (33).
In the studies described above, GHOST cells served as
tar-get cells and both MAbs and polyclonal Abs were used to
mediate neutralization. The MAbs provide more quantitative
analyses and provide more refined information for analyzing
specific epitopes involved in neutralization. The two MAbs
used in this study were against functionally different sites: MAb
447-52D is a V3-specific MAb (18, 20), and IgG1b12 is
di-rected to an epitope that overlaps the CD4 binding domain (8,
41). The 50% neutralizing concentrations ranged from
,
1 to
20
m
g/ml for 447-52D and from
,
0.5 to 10
m
g/ml for IgG1b12.
Analysis of the neutralization sensitivities of the 10 isolates
tested with MAb 447-52D revealed that effective neutralization
did not always correlate with the presence of the core epitope
defined for this MAb. Thus, the core epitope could be present
in the envelope of a virus, e.g., 92HT593, that the MAb failed
to neutralize, or could have a substitution in the core epitope,
e.g., SF33, and still be neutralized. Since it is known that MAb
447-52D recognizes both linear and conformational aspects of
the virus envelope (18), the data suggest that presentation of
the same epitope on the envelopes of different isolates varies,
affecting the binding of Ab or the conformational changes that
the MAb induces in the virus envelope. This could occur, for
example, as the result of a change in the dissociation rate of the
Ab from the virion; this is known to profoundly affect the
neutralizing capacity of MAbs (48). It could also occur if
changes at sites other than the core epitope affect epitope
exposure or conformation. For example, changes in the
glyco-sylation of gp120 affect the exposure of epitopes (5) and
changes in amino acids affect epitopes at distant sites (36, 39).
Other factors, such as the presence of adhesion molecules on
the surface of virions, may also play a role in changing the
neutralization characteristics of the virus (6, 16, 40).
Loss of neutralization sensitivity is not necessarily
accompa-nied by loss of antibody binding, indicating that changes in the
epitope may affect the way in which the MAb interferes with
the process of virus infectivity, e.g., fusion and uncoating (31,
36). The finding that a single passage of plasma virus through
PHA-stimulated PMBCs changes the neutralization profiles
and surface characteristics of primary isolates (6, 17) highlights
the mutability of HIV. However, what actually contributes to
the neutralization sensitivity of an isolate is not known.
Prob-ably several factors contribute, since different Abs function at
different steps in virus infection. Thus, an anti-V3
conforma-tion-dependent Ab blocks infection at a postinternalization
step (2), the Fab fragment of IgG1b12 neutralizes at a
postfu-sion step, the whole IgG1b12 molecule inhibits virus fupostfu-sion
(30), and receptor blocking has been reported as a mechanism
of action for several human anti-HIV MAbs (10, 46).
There-fore, the neutralization of an isolate is defined not just by the
presence of an epitope or by virus interaction with CXCR4 or
CCR5, but also by the presentation of the epitope, the way it
interacts with the Ab, and the effect of this interaction on the
virus.
ACKNOWLEDGMENTS
This work was supported in part by NIH grants HL 59725, AI 36085,
and AI 32424; by NIAID grant AI 27742 supporting the NYU Center
for AIDS Research; and by funds from the Research Center for AIDS
and HIV Infection of the Department of Veterans Affairs, New York,
N.Y. V.N.K. is a postdoctoral fellow of the Damon Runyan-Walter
Winchell Foundation, and D.R.L. is an investigator of the Howard
Hughes Medical Institute.
REFERENCES
1. Arendrup, M., A. Sonnerborg, B. Svennerholm, L. Akerblom, C. Nielsen, H.
Clausen, S. Olofsson, J. O. Nielsen, and J. S. Hansen.1993. Neutralizing antibody response during human immunodeficiency virus type 1 infection: type and group specificity and viral escape. J. Gen. Virol. 74:855–863. 2. Armstrong, S. J., T. L. McInerney, L. McLain, B. Wahren, J. Hinkula, M.
Levi, and N. J. Dimmock.1996. Two neutralizing V3 monoclonal anti-bodies act by affecting different functions of human immunodeficiency virus type 1. J. Gen. Virol. 77:2931–2941.
3. Auewarakul, P., S. Louisirirotchanakul, R. Sutthent, T. Taechowisan, C.
Kanoksinsombat, and C. Wasi.1996. Analysis of neutralizing and enhancing antibodies to human immunodeficiency virus type 1 primary isolates in plasma of individuals infected with env genetic subtype B and E viruses in Thailand. Viral Immunol. 9:175–185.
4. Back, N. K., L. Smit, E. Hogervorst, A. C. van Wijk, J. Goudsmit, and M.
Tersmette.1994. Development and evaluation of an HIV-1
transfection-TABLE 2. Sequence comparison of the V3 loop of viruses studied
for neutralization
Virus Isolate V3 loop sequencea Neutralization byAnti-V3 MAb 447-52D
Consensus B
SIHI..GPGR..AFYTTGE
BZ167
R-R-..----..T--.--K
1
MNp
R---..----..---KN
1
92HT594
R-R-..----..VW----Q
2
JR-FL
--TL..----..---D
2
91US056
G---..---A..---A---
2
CA5
G---..----..-IF----
2
SF33
R-TS..---K..VL---
1
SF2
--Y-..----..--H---R
1
89.6
RLS-..----..---ARRN
2
92HT593
R-S-..----..--RA-.K
2
aThe sequences were obtained from the database of Myers et al. (35), and the box denotes the core epitope recognized by MAb 447-52d. Dots are for align-ment; dashes denote homology.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.49.290.89.227.2]neutralization assay. J. Acquired Immune Defic. Syndr. 7:531–538. 5. Bandres, J. C., Q. F. Wang, J. O’Leary, F. Baleaux, A. Amara, J. Hoxie, S.
Zolla-Pazner, and M. K. Gorny. 1998. Human immunodeficiency virus (HIV) envelope binds to CXCR4 independently of CD4, and binding can be enhanced by interaction with soluble CD4 or by HIV envelope deglycosyla-tion. J. Virol. 72:2500–2504.
6. Bastiani, L., S. Laal, S. Zolla-Pazner, and M. Kim. 1997. Host cell-depen-dent alterations in envelope components of HIV-1 virions. J. Virol. 71:3444– 3450.
7. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. MacKay. 1997. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regu-lated on human T lymphocytes. Proc. Natl. Acad. Sci. USA 94:1925–1930. 8. Burton, D. R., J. Pyati, R. Koduri, S. J. Sharp, G. B. Thornton, P. W. H. I.
Parren, L. S. W. Sawyer, R. M. Hendry, N. Dunlop, P. L. Nara, M. Lamac-chia, E. Garratty, E. R. Stiehm, Y. J. Bryson, Y. Cao, J. P. Moore, D. D. Ho, and C. F. Barbas III.1994. Efficient neutralization of primary isolates of HIV-1 by a recombinant human monoclonal antibody. Science 266:1024– 1027.
9. Candotti, D., M. Rosenheim, J. M. Huraux, and H. Agut. 1997. Two PBMC-based neutralization assays depict low reactivity of both anti-V3 monoclonal antibodies and immune sera against HIV-1 primary isolates. J. Virol. Meth-ods 64:81–93.
10. Chen, Y. H., and M. P. Dierich. 1996. Identification of a second site in HIV-1 gp41 mediating binding to cells. Immunol. Lett. 52:153–156.
11. Ciminale, V., B. K. Felber, M. Campbell, and G. N. Pavlakis. 1990. A bioassay for HIV-1 based on env-CD4 interaction. AIDS Res. Hum. Retro-viruses 6:1281.
12. Conley, A. J., M. K. Gorny, J. A. Kessler II, L. J. Boots, D. Lineberger, E. A.
Emini, M. Ossorio, S. Koenig, C. Williams, and S. Zolla-Pazner.1994. Neutralization of primary HIV-1 virus isolates by the broadly-reactive an-ti-V3 monoclonal antibody, 447-52D. J. Virol. 68:6994–7000.
13. Deng, H., D. Unutmaz, V. N. Kewalramani, and D. R. Littman. 1997. Ex-pression cloning of new receptors used by simian and human immunodefi-ciency viruses. Nature 388:296–300.
14. D’Souza, M. P., D. Livnat, J. A. Bradac, S. H. Bridges, and The AIDS
Clinical Trials Group Antibody Selection Working Group.1997. Evaluation of monoclonal antibodies to human immunodeficiency virus type 1 primary isolates by neutralization assays: performance criteria for selecting candidate antibodies for clinical trials. J. Infect. Dis. 175:1056–1062.
15. Fortin, J., R. Cantin, G. Lamontagne, and M. Tremblay. 1997. Host-derived ICAM-1 glycoproteins incorporated on human immunodeficiency virus type 1 are biologically active and enhance viral infectivity. J. Virol.
71:3588–3596.
16. Frank, I., H. Stoiber, S. Godar, H. Stockinger, F. Steindl, H. W. D. Katinger,
and M. P. Dierich.1996. Acquisition of host cell-surface-derived molecules by HIV-1. AIDS 10:1611–1620.
17. Gauduin, M., G. P. Allaway, P. J. Maddon, C. F. Barbas III, D. R. Burton,
and R. A. Koup.1996. Effective ex vivo neutralization of human immuno-deficiency virus type 1 in plasma by recombinant immunoglobulin molecules. J. Virol. 70:2586–2592.
18. Gorny, M. K., A. J. Conley, S. Karwowska, A. Buchbinder, J. Xu, E. A. Emini,
S. Koenig, and S. Zolla-Pazner.1992. Neutralization of diverse HIV-1 vari-ants by an anti-V3 human monoclonal antibody. J. Virol. 66:7538–7542. 19. Gorny, M. K., T. C. VanCott, C. Hioe, Z. R. Israel, N. L. Michael, A. J.
Conley, C. Williams, J. A. Kessler II, P. Chigurupati, S. Burda, and S. Zolla-Pazner.1997. Human monoclonal antibodies to the V3 loop of HIV-1 with intra- and inter-clade cross-reactivity. J. Immunol. 159:5114–5122. 20. Gorny, M. K., J. Xu, S. Karwowska, A. Buchbinder, and S. Zolla-Pazner.
1993. Repertoire of neutralizing human monoclonal antibodies specific for the V3 domain of HIV-1 gp120. J. Immunol. 150:635–643.
21. He, J., Y. Chen, M. Farzan, H. Choe, A. Ohagen, S. Gartner, J. Busciglio, X.
Yang, W. Hofmann, W. Newman, C. R. Mackay, J. Sodroski, and D. Gabuzda.1997. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. Nature 385:645–649.
22. Hioe, C. E., S. Xu, P. Chigurupati, S. Burda, C. Williams, M. K. Gorny, and
S. Zolla-Pazner.1997. Neutralization of HIV-1 primary isolates by poly-clonal and monopoly-clonal human antibodies. Int. Immunol. 9:1281–1290. 23. Hogervorst, E., J. De Jong, A. Van Wijk, M. Bakker, M. Valk, P. Nara, and
J. Goudsmit.1995. Insertion of primary syncytium-inducing (SI) and non-SI envelope V3 loops in human immunodeficiency virus type 1 (HIV-1) LAI reduces neutralization sensitivity to autologous, but not heterologous, HIV-1 antibodies. J. Virol. 69:6342–6351.
24. KewalRamani, V. N., B. Volsky, D. Kwon, W.-K. Xiang, J. Gao, D. Unutmaz,
C. M. Hill, R. E. Sutton, and D. R. Littman.Unpublished data.
25. Kimpton, J., and M. Emerman. 1992. Detection of replication-competent and pseudotyped human immunodeficiency virus with a sensitive cell line on the basis of activation of an integratedb-galactosidase gene. J. Virol. 66: 2232–2239.
26. Laal, S., S. Burda, S. Sharpe, and S. Zolla-Pazner. 1993. A rapid, automated microplate assay for measuring neutralization of HIV-1. AIDS Res. Hum. Retroviruses 9:781–785.
27. LaCasse, R. A., K. E. Follis, T. Moudgil, M. Trahey, J. M. Binley, V.
Planellas, S. Zolla-Pazner, and J. H. Nunberg.1998. Coreceptor utilization by human immunodeficiency virus type 1 is not a primary determinant of neutralization sensitivity. J. Virol. 72:2491–2495.
28. Mascola, J. R., J. Louwagie, F. E. McCutchan, C. L. Fischer, P. A. Hegerich,
K. F. Wagner, A. K. Fowler, J. G. McNeil, and D. S. Burke.1994. Two antigenically distinct subtypes of HIV-1: viral genotype predicts neutraliza-tion immunotype. J. Infect. Dis. 169:48–54.
29. Mascola, J. R., S. W. Snyder, O. S. Weislow, S. M. Belay, R. B. Belshe, D. H.
Schwartz, M. L. Clements, R. Dolin, B. S. Graham, G. J. Gorse, M. C. Keefer, M. J. McElrath, M. C. Walker, K. F. Wagner, J. G. McNeil, F. E. McCutchan, and D. S. Burke, for the NIAID AIDS Vaccine Evaluation Group.1996. Immunization with envelope subunit vaccine products elicits neutralizing antibodies against laboratory-adapted but not primary isolates of human immunodeficiency virus type 1. J. Infect. Dis. 173:340–348. 30. McInerney, T. L., L. McLain, S. J. Armstrong, and N. J. Dimmock. 1997. A
human IgG1 (b12) specific for the CD4 binding site of HIV-1 neutralizes by inhibiting the virus fusion entry process, but b12 Fab neutralizes by inhibiting a postfusion event. Virology 233:313–326.
31. McKeating, J. A., J. Bennett, S. Zolla-Pazner, M. Schutten, S. Ashelford,
A. L. Brown, and P. Balfe.1993. Resistance of a human serum-selected human immunodeficiency virus type 1 escape mutant to neutralization by CD4 binding site monoclonal antibodies is conferred by a single amino acid change in gp120. J. Virol. 67:5216–5225.
32. McLain, L., and N. J. Dimmock. 1994. Single- and multi-hit kinetics of immunoglobulin G neutralization of human immunodeficiency virus type 1 by monoclonal antibodies. J. Gen. Virol. 75:1457–1460.
33. Means, R. E., T. Greenough, and R. C. Desrosiers. 1997. Neutralization sensitivity of cell culture-passaged simian immunodeficiency virus. J. Virol.
71:7895–7902.
34. Montefiori, D. C., R. G. Collman, T. R. Fouts, J. Y. Zhou, M. Bilska, J. A.
Hoxie, J. P. Moore, and D. P. Bolognesi. 1998. Evidence that antibody-mediated neutralization of human immunodeficiency virus type 1 by sera from infected individuals is independent of coreceptor usage. J. Virol. 72: 1886–1893.
35. Myers, G., B. Korber, B. Foley, R. F. Smith, K. Jeang, J. W. Mellors, and A.
Wain-Hobson.1996. Human retroviruses and AIDS. Theoretical Biology and Biophysics, Los Alamos National Laboratories, Los Alamos, N.M. 36. Nara, P. L., L. Smit, N. Dunlop, W. Hatch, M. D. Waters, J. Kelliher, R. C.
Gallo, P. J. Fischinger, and J. Goudsmit.1990. Emergence of viruses resis-tant to neutralization by V3-specific antibodies in experimental human im-munodeficiency virus type 1 (IIIB) infection of chimpanzees. J. Virol. 64: 3779–3791.
37. Nyambi, P. N., L. Heyndrickx, W. Janssens, F. Daeyaert, P. Lewi, K.
Fransen, B. Willems, S. Coppens, K. Vereecken, P. Piot, and G. van der Groen.Identification of specific amino acid motifs and biophenotypic prop-erties among primary HIV-1 group M (subtype A-H) and O isolates belong-ing to different neutralization clusters. Submitted for publication. 38. Nyambi, P. N., J. Nkengasong, P. Lewi, K. Andries, W. Janssens, K. Fransen,
L. Heyndrickx, P. Piot, and G. van der Groen.1996. Multivariate analysis of human immunodeficiency virus type 1 neutralization data. J. Virol. 70:6235– 6243.
39. Reitz, M. S., C. Wilson, C. Naugle, R. C. Gallo, and M. Robert-Guroff. 1988. Generation of a neutralization-resistant variant of HIV-1 is due to selection for a point mutation in the envelope gene. Cell 54:57–63.
40. Rizzuti, C. D., and J. G. Sodroski. 1997. Contribution of virion ICAM-1 to human immunodeficiency virus infectivity and sensitivity to neutralization. J. Virol. 71:4847–4851.
41. Roben, P., J. P. Moore, M. Thali, J. Sodroski, and C. F. Barbas III. 1994. Recognition properties of a panel of human recombinant Fab fragments to the CD4 binding site of gp120 that show differing abilities to neutralize human immunodeficiency virus type 1. J. Virol. 68:4821–4828.
42. Sallusto, F., C. R. Mackay, and A. Lanzavecchia. 1997. Selective expression of the eotaxin receptor CCR3 by human T helper 2 cells. Science 227:2005– 2007.
43. Sawyer, L. W., M. T. Wrin, L. Crawford-Miksza, B. Potts, Y. We, P. A.
Weber, R. D. Alfonso, and C. V. Hanson.1994. Neutralization sensitivity of human immunodeficiency virus type 1 is determined in part by the cell in which the virus is propagated. J. Virol. 68:1342–1349.
44. Scarlatti, G., J. Albert, P. Rossi, V. Hodara, P. Biraghi, L. Muggiasca, and
E. M. Fenyo.1993. Mother-to-child transmission of human immunodefi-ciency virus type 1: correlation with neutralizing antibodies against primary isolates. J. Infect. Dis. 168:207–210.
45. Trkola, A., T. Ketas, V. N. KewalRamani, F. Endorf, J. M. Binley, H.
Katinger, J. Robinson, D. R. Littman, and J. P. Moore.1998. Neutralization sensitivity of human immunodeficiency virus type 1 primary isolates to an-tibodies and CD4-based reagents is independent of coreceptor usage. J. Vi-rol. 72:1876–1885.
46. Ugolini, S., I. Mondor, P. W. H. I. Parren, D. R. Burton, S. Tilley, P. J.
Klasse, and Q. J. Sattentau.1997. Inhibition of virus attachment to CD41
target cells is a major mechanism of T cell line-adapted HIV-1 neutraliza-tion. J. Exp. Med. 186:1287–1298.
47. Uguccioni, M., C. R. Mackay, B. Ochensberger, P. Loetscher, S. Rhis, G. J.
on November 9, 2019 by guest
http://jvi.asm.org/
LaRosa, P. Rao, P. D. Ponath, M. Baggiolini, and C. A. Dahinden.1997. High expression of the chemokine receptor CCR3 in human blood basophils. Role in activation by eotaxin, MCP-4, and other chemokines. J. Clin. Invest.
100:1137–1143.
48. VanCott, T. C., F. R. Bethkes, V. R. Polonis, M. G. Gorny, S. Zolla-Pazner,
R. R. Redfield, and D. L. Birx.1994. Dissociation rate of antibody-gp120 binding interactions is predictive of V3-mediated neutralization of HIV-1. J. Immunol. 153:449–459.
49. Weber, J., E.-M. Fenyo, S. Beddows, P. Kaleebu, A. Bjorndal, and the WHO
Network for HIV Isolation and Characterization.1996. Neutralization sero-types of HIV-1 field isolates are not predicted by genetic subtype. J. Virol.
70:7827–7832.
50. Zhang, L., T. He, K. Kuntsman, S. Wu, Y. Guo, A. U. Neumann, D. D. Ho,
and S. M. Wolinsky.1998. HIV-1 phenotype, co-receptor usage and disease progression, abstr. 281, p. 132. In Abstracts of the 5th Conference on Ret-roviruses and Opportunistic Infections.
51. Zolla-Pazner, S. 1996. Mechanisms contributing to the neutralization of HIV-1. Immunol. Lett. 51:89–93.
52. Zolla-Pazner, S., C. Alving, R. Belshe, P. Berman, S. Burda, P. Chigurupati,
M. L. Clements, A.-M. Duliege, J.-L. Excler, J. Kahn, M. J. McElrath, S. Sharpe, F. Sinangil, K. Steimer, M. C. Walker, N. Wassef, and S. Xu.1997. Neutralization of a clade B primary isolate by sera from HIV-uninfected recipients of candidate AIDS vaccines. J. Infect. Dis. 175:764–774. 53. Zolla-Pazner, S., M. Lubeck, S. Xu, S. Burda, R. J. Natuk, F. Sinangil, K.
Steimer, R. C. Gallo, J. W. Eichberg, T. Matthews, and M. Robert-Guroff.
1998. Induction of neutralizing antibodies in T-cell line-adapted and primary human immunodeficiency virus type 1 isolates with a prime-boost vaccine regimen in chimpanzees. J. Virol. 72:1052–1059.
54. Zolla-Pazner, S., and S. Sharpe. 1995. A resting cell assay for improved detection of antibody-mediated neutralization of HIV-1 primary isolates. AIDS Res. Hum. Retroviruses 11:1449–1457.
on November 9, 2019 by guest
http://jvi.asm.org/