JOURNALOFVIROLOGY, Aug.1993, p.4875-4885 0022-538X/93/084875-11$02.00/0
Copyright©1993, American Society for Microbiology
Vol. 67, No. 8
Separation
of the Complex
DNA
Binding Domain of EBNA-1
into
DNA
Recognition
and Dimerization Subdomains of
Novel Structure
MEI-RU
CHEN,'
JAAP M. MIDDELDORP,2AND S. DLANEHAYWARDl.3*
Molecular Virology Laboratories, Department ofOncology1 and Department of Pharmacology and Molecular Sciences,3JohnsHopkinsSchoolofMedicine, 725 North WolfeStreet, Baltimore, Maryland21205-2185,
andOrganon-Teknika, 5281 RM Boxtel, The Netherlands2 Received 28 January1993/Accepted 15 May 1993
EBNA-1 isessential for replication of the latent episomal form of the Epstein-Barr virus genome and is involvedinregulation of viral latency promoters. EBNA-1
activity
is mediated through direct DNA binding. The DNAbinding and dimerization functions of EBNA-1 havepreviously
been located to a carboxy-terminal domain, amino acids (aa) 459to607.Toidentifyanddefinethesubdomainsfor thesetwofunctions, we created anextensive series of deletions andpointmutationsinanEBNA-1 (aa 408to641)background.
Theability
of the EBNA-1 mutants to heterodimerize with awild-type EBNA-1 (aa 459to 641) polypeptidewas tested in immunoprecipitation assays withamonoclonal antibody,EBNA.OTlx, that recognizes EBNA-1 (aa 408 to 641) butnotEBNA-1 (aa459to641).Theseexperiments revealed that mutations affecting dimerization occurred over twoseparateregions, aa501to532 andaa554to598. DNAbindingwastested inmobility
shift assays againstapanelofoligonucleotide-binding sites. Dimerizationwas aprerequisitefor DNAbinding. The DNA recognition domainwaslocalizedto a separateregion, aa459to487, upstream of the dimerization domain. EBNA-1 variantscarryingsubstitutions ataa467 and 468 andataa477 gaveapatternofbindingtomutantoligonucleotide probes thatimplicatestheseparticular amino acidsin DNArecognition. EBNA-1 appears to utilize novel mechanisms for both DNA recognition and dimerization since neither domain conforms to
previously
described structural motifs.The EBNA-1proteinhasacentral role in the maintenance ofalatentEpstein-Barrvirus infection. EBNA-1 is theonly virus-encoded protein required for
replication
of the episo-mal form of the Epstein-Barr virus genome(39, 41).
The latency origin of replication, oriP, contains twoEBNA-1-binding loci: region I, the family of repeats, consists of 20 tandemcopiesofthe
EBNA-1-binding
site,
andregion
II,thedyad symmetry
region,
contains twopairs
ofoverlapping,
lower-affinity,binding sites
(15, 24, 26).
Thedyad
symmetryregionis the site ofinitiationof DNA
replication
(9)
while thefamily of repeats
provides
an enhancer function(25,
38).
Electron microscopic studies have indicated that EBNA-1
moleculesboundtothesetwoelementsoforiPcanmediate
loopingout of the intervening DNA sequences, a process that may facilitate formation of the replication initiation
complex(8, 35). An additional role for EBNA-1 in nuclear matrix attachment during replication has been
postulated
(14).EBNA-1 alsoparticipatesin
regulation
oflatency
genetranscription. EBNA-1-mediated transactivation of the la-tencyC promoter has been described
previously (36),
andimmediately downstreamof the
latency
F promoterare twoEBNA-1-binding
siteswhich maynegatively
modulate tran-scription from.thispromoter(2, 28, 29, 31,
33).
The
replication
andgeneregulation
functionsof EBNA-1 aremediatedthrough
DNAbinding
(15, 24).
EBNA-1 binds DNA as a dimer (1, 7), and both the DNA binding anddimerization functions are located in the
carboxy-terminal
third of the 641-amino-acid
(aa) protein (1, 13, 24).
Mapping
studies usingamino- and
carboxy-terminal
deletions of this segmentof the EBNA-1 openreading
frameplaced
theDNA*Correspondingauthor.
binding and dimerization domains between aa 459 and 607 (1). The relative positioning of these two functional domains andtheirnaturehave beenamatterof conjecture. An amino acid sequence comparison led Inoue et al. (13) to suggest that EBNA-1 contained a basic helix-loop-helix (bHLH) domain. The nature of the dimerization domain is of some relevance since certain motifs, such as the helix-loop-helix motif, allow heterodimerization with other proteins of the same class. Such interactions can significantly modify the functionof the individualproteins involved.
In a previous attempt to separate the DNA-binding and
dimerization domains, we created a set of relatively large internal deletions across the aa 459 to 607 region (32). However, these deletions all abolished DNA binding and greatly reduced dimerization, indicating that the entire re-gion spanningaa459to607wasvery sensitive to structural
perturbation. In the present study, we sought to minimize
nonspecific effects on EBNA-1 structure by introducing a series of amino acid substitutions and small five- to seven-aa deletions into the segment of EBNA-1 from aa 459 to 607. These mutants plus the availability of a new monoclonal
antibody against EBNA-1, EBNA.OTlx, allowed us to separate the individual DNAbinding and dimerization do-mains andtogainsomeinsightintoamino acids that may be involved in DNArecognition. Interestingly, EBNA-1 does not appeartobelong to anypreviouslyrecognized class of DNA-binding protein.
MATERIALSANDMETHODS
Plasmidconstructions.Wild-typeEBNA-1 (aa408 to641)
[referred
to hereafter as EBNA-1(408-641)] was expressed byinvitrotranscriptionandtranslation frompRA362(1)
and 4875on November 9, 2019 by guest
http://jvi.asm.org/
4876 CHEN ET AL.
EBNA-1 (aa 459to 641) [referred to as EBNA-1(459-641)] was generated similarly from pWS61 (32). The deletion series pRA374(479SRA481-520), pRA376(499SRA501-20), pRA378(A525-32), pWS52A(A537-53), pWS43(555SRA557-64), pWS46(576SRA578-616), and pWS47(A599-614,615SR) has been described previously (32), as have EBNA-1 con-structions containing uniqueXbaI sites at codons 479 and 480,499 and500,521 and522,555 and556,and 615 and616. Three of the XbaI mutations were transferred into the in vitro transcription vector pGH253 to create pMRC32 (479SR), pMRC30(521S), andpMRC31(576SR).
Theoligonucleotides LRA27(5'-CCGGATCCTGTAGGG GAAGCCGAT), which annealstothe EBNA-1 templateat codon 408, and LGH312 (5'-GGCCCTGCAGTCACTCCTG CCCTTlCCTC), which anneals to the EBNA-1 template at codon 641, served as the outside primers for polymerase
chain reaction (PCR)mutagenesis. Recombinant PCR with these outside primers and a pair of overlapping inside
primerswasusedtogeneratepMRC35(467VD) containinga SalI site at codons 467 and 468 and pMRC36 (SllVY) containing anAccIsite atcodons 511 and 512. Briefly,the
oligonucleotides LRA27 and5'-CACGGTCGACTCCAAAC CACCCTCC were used to amplify codons 408 to 473 and introduce mutated sequences at codons 467 and 468, and oligonucleotides 5'-TGGAGTCGACCGTGGTCAAGGAGGT
and LGH312wereused to amplifycodons 462 to641with
mutations at 467 and 468. The two PCRproducts were then
purified, denatured, annealed, andagain amplified with the outside primers LRA27 and LGH312 and cloned into
pGH253to createpMRC35.Similarly, plasmid pMRC36was generated with the primer pairs LRA27 and 5'-TACTGTA TACATATACGAACACACC to amplify codons 408to517 with mutationsatcodons 467 and 468, andoligonucleotides
5'-ATATGTATACAGTAAGACCTCCCTl
and LGH312wereused toamplify codons 506 to 641 withmutationsat511 and 512. A similar strategy was also used to generate
pMRC42(513A). The outside primers for pMRC42 were LRA27 andLRA28, 5'-CGGATCCTGCAGGAATTCCAAT TGCCCCATGTTGGTA (whichannealsoutside the EBNA-1 open reading frame), and the internal primer pair was
5'-GGAGGTClTTAGCACCTCCATAand5'-TATGGAGGT
GCTAAGACCTCC.
The second set of small deletionswere created by using
forward PCRprimerscontaining
XbaI,
SacI,orSalIrestric-tionenzymesitesto loopoutthe desiredcodons.The PCR
products obtained with these primers and LGH312 were then used to replace the equivalent wild-type DNA frag-ment. The PCR primers, with restriction sites underlined, were
5'-GCrATCJAGA
TITrACAAACTCATATA,
pMRC 45(555SRA557-64);
5'-ATTIT[CA.AAGAGCTCTCCTG-GCTAGG, pMRC46 (479SRA481-5);5'-TACCTCTAGATT
CGTATATGGAGGTAGT, pMRC47 (499SRA501-7); 5'-TT
AAGzAGCTfCTCGTAGAAAGGACTACC,
pMRC49(A489-93); and
5'-TGGAGTCiGACTCCAACCCGAAATIT,
pMRC57A1 (467VDA469-73). Plasmid pMRC57A2
(467VDA469-73,478V) contains the same deletion as pMRC57A1 plus a spontaneous mutation (aa 478, F to V). EBNA-1(468-641)
(pMRC53)wasmadebymoving the
SaiI-PstI
DNA fragment from pMRC35 into an in vitro transcription vector.UtilizingtheintroducedXbaI sites or the nativeSacl site in the EBNA-1 gene and mismatched PCR primers, the
followingadditionalpoint mutations were created: pMRC24
(485D;
reverse primer:5'-CAGGAGAGTCTCTATCACCTT
CTGCAATG; Sacl); pMRC26 (477A-SR; reverse primer5'-CAAT[CTAGAAAATGCCGGGTTGGAACC;
XbaI at479 and 480); pMRC28 (574DD; reverse primer 5'-AAT
CGCTCTACIAATCATCCTCAGCAAATAT; XbaI at 576 and577); pMRC29(517AAASS;reverseprimer 5'-AGTTCC
TCTAGATGAGGGGGCAGCGGAGGTCTTACT; XbaI at 521 and 522); pMRC33 (488R;
5'-TTAAGjAGCTCGCCTG
GCTAGGAGT; SacI site); and pMRC56 (490P; 5'-TTAA
GAGCTCTCCTGCCCAGGAGTCACGTA,
SacI site). A pair of oligonucleotides, 5'-CTAGAGGAACTTGGGTCC ACT and 5'-CTAGAGTGGACCCAAGTTCCT, were an-nealed and cloned into theXbaI site ofpMRC47to create pMRC50(499SR505HSR). Cloningin thereverseorientation created pMRC51(499SRSVAPSSS). All the mutations were confirmedby double-strandeddideoxysequencing. With theexception of the one case noted, no spontaneous PCR-generated alterations were observed.
In vitro transcription-translation. Plasmid DNAwas lin-earized downstream of thecodingsequenceof EBNA-1 and incubated with T7 RNA polymerase to prepare capped
mRNAby usinganinvitrotranscriptionkit(Stratagene, La
Jolla, Calif.). In vitro translation was carried out byusing rabbit reticulocyte lysates (Promega, Madison, Wis.). A standard in vitrotranslation reaction contained 1to2 ,ug of mRNA in a 50-,ul reaction mixture and 50 ,Ci of
[35S]me-thionine (800
Ci/mmol)
purchased from New England Nu-clear (Wilmington, Del.). Cotranslations contained equalamountsof the twodifferent RNAs. Labeledproteinswere storedat -70°C.
Immunoprecipitation assays usingEBNA.OTlx. The mono-clonal antibody EBNA.OTlx specifically recognizes an epitope between aa 408 and 459 of EBNA-1 (19a). Two microliters of each invitro-translatedproteinwasdilutedto 100 ,u with TSET buffer(150mM NaCl, 50 mMTris-HCl,
0.1 mM EDTA, 2% Triton X-100 [pH 8.0]), and 2 ,u of EBNA.OTlx hybridoma culture supernatant was added. The mixturewasincubatedat4°Cfor 1 h. Fivemilligramsof proteinA-Sepharose(in TSET)wasadded, andafter 30 min ofincubationat4°C,theprecipitatewaspelletedand washed onceinTSET and threetimesin NET(150mMNaCl,50 mM Tris-HCl [pH7.4],0.1 mMEDTA).The samplewas resus-pended in buffer(62.5mM Tris[pH 6.8], 10% glycerol, 4%
sodiumdodecyl sulfate[SDS],100 mMdithiothreitol, 0.01% bromophenolblue) andboiled for 3 min before
electrophore-sison anSDS-13% polyacrylamide gel.
Preparation ofoligonucleotide-binding substrates.
Symmet-ricconsensusand mutantbindingsiteprobeswereprepared
from 19-mer synthetic single-stranded oligonucleotides
which could self anneal and thus serve as primers for a Klenow fill-inreaction, generating double-stranded 30-mers
(2). The oligonucleotide-bindingsites usedwere20, 33, 36,
71, 73,74, 96, 97,100, 102, 104,196, and197,asdescribedin
Ambinderetal. (2). Oligonucleotides (10 pmol)wereheated to 75°C and slowly cooled to room temperature for self
annealing.
[32P]dATP
(10 ,Ci), and 10 nmol each ofdCTP, dGTP, anddTTJPwasaddedto a20-,u reaction, along with S U of Klenow polymerase. After 1 h of incubation, the reactionwasstopped byadding45 ,ul ofSTE(150mMNaCl,10 mMTris-HCl,0.1mMdisodium EDTA[pH 7.4]), and the DNAwaspurifiedinaG-25spin column(5Prime- 3Prime,
Inc., Boulder, Colo.).
Electrophoretic mobility shift assays. In vitro-translated EBNA-1 wasincubated withprobeDNA(20 fmol)for 30 min at room temperature in a 25-,ul total volume of 25 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid [pH 7.5])-1 mM dithiothreitol-5 mM MgCl2-1 mg of bovine serum albumin per ml-100 mM
KCl-0.1%
Nonidet P-40-1 ,ug of poly(dI-dC) (Pharmacia, Inc., Piscataway,N.J.).Forsupershiftassays, 2
RI
of 1:10 dilutedmonoclonal J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
EBNA-1 DNA RECOGNITION AND DIMERIZATION 4877
(A)
(B)-cotransiationwitn(459-641) cotranslation with(459-641V
0 CCC Ul Lr)
-O
U)
(m -C
Cd CD CD U)
N 00 m ,
U° n o r U1 "r 'r q v
o cr
O L -t0I
N 1-U In _3
I
o U)
ICC4 Ln LI) LO q Cf, < LO < ;-(I
.n tnU1 vn c
N 0) n LO to v UL c) Ln Ln N- un
< T <C ) L) <
kDa
46
-30-
-.1.5
21.5-_ -_M
1 2 3 4 5 6 7 S 9 10 2 I3 4 ) 7 8 9 10
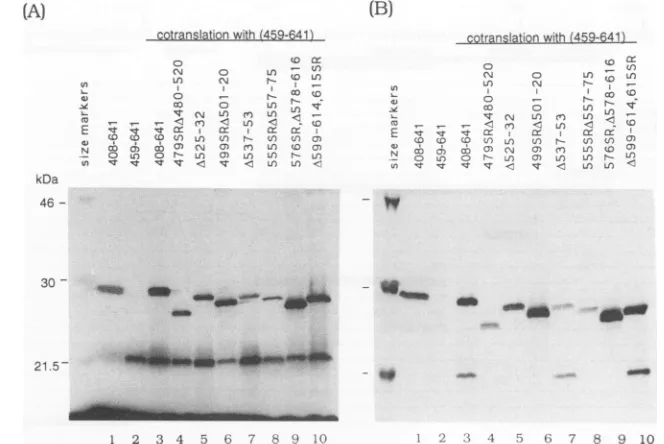
FIG. 1. Useof the monoclonalantibodyEBNA.OT1xtoassessEBNA-1 dimerization.(A)Invitro-translatedEBNA-1polypeptideswere
analyzed by SDS-PAGEon a13%gel.Each lane contains 3plof thein vitrotranslation mixture. Lanes1, wild-typeEBNA-1(408-641); 2, wild-typeEBNA-1(459-641); 3,cotranslation ofEBNA-1(408-641)andEBNA-1(459-641); 4to10, cotranslation ofEBNA-1(459-641)witha
previously describedsetof EBNA-1 deletions(31). (B)Immunoprecipitationoftheinvitro-translatedpolypeptidesshown inpanelA with
monoclonal antibody EBNA.OTlx. EBNA-1(408-641)wasimmunoprecipitated bytheantibody (lane 1)butEBNA-1(459-641)wasnot(lane 2). Cotranslation of EBNA-1(459-641) with EBNA(1408-641) resulted in the formation of a heterodimer, and EBNA-1(459-641) was
coimmunoprecipitatedwithEBNA.OT1x(lane 3).Lanes4to10,immunoprecipitationaftercotranslationof theindicated EBNA-1deletions
withEBNA-1(459-641).
antibody (EBNA.OTlx) culture supernatant was added to the mixture and incubatedfor15 minat roomtemperature.
The reaction mixtures were loaded onto a 6%
polyacryl-amide gel in HEE buffer (10 mMHEPES [pH 7.5], 1 mM EDTA, 5 mM EGTA) and electrophoresed at 10 V/cm at
roomtemperaturefor 1.5 h. The gelswere dried and auto-radiographed.
RESULTS
Useof the monoclonal antibodyEBNA.OTlx toassay for EBNA-1dimerization. Both the DNAbindingand dimeriza-tion domains of EBNA-1maptoaregion, aa459 to607, at thecarboxylend of theprotein (1).Inaprevious attemptto separatethedomains for thesetwofunctions, asetoflarge internal deletionsacrossthisregionwastested for dimeriza-tionability byusingglutaraldehyde cross-linking (32).
How-ever,each of the deletionshadseverelycompromised
dimer-ization abilityinthis assay.Thus, adifferentapproachwas
required to identify sequencesinvolved in EBNA-1 dimer-ization. Here we report the use of an EBNA-1-specific
monoclonal antibody and a new series of mutations that made itpossibletodissectregionsof theprotein requiredfor
dimerization.
EBNA.OTlxis a monoclonal antibodyraised against an
EBNA-1 protein thatlacked the Gly-Gly-Ala repeats. The antibodyrecognizesan epitopelocated betweenaa408 and
aa459 of EBNA-1 (19a). When this antibodywas used to immunoprecipitate different in vitro-translated EBNA-1 polypeptides, we found that EBNA.OTLx immunoprecipi-tated in vitro-translated EBNA-1(408-641) (Fig. 1B, lane 1) but notEBNA-1(459-641) (Fig. 1B, lane2). When cotrans-lated, EBNA-1(408-641) formed a heterodimer with the polypeptide, spanning aa 459 to 641, which could now be
coimmunoprecipitated with EBNA.OTLx (Fig. 1B, lane 3). Thus, coimmunoprecipitation of EBNA-1(459-641) could be
used to assay for the dimerization ability of deleted or
mutatedaa408to641polypeptides. The originalsetof large internal deletions was cotranslated with EBNA-1(459-641)
(Fig. 1A,lanes 4to10).Thecotranslated polypeptideswere
thenimmunoprecipitatedwithEBNA.OTlx (Fig. 1B, lanes4 to10).Inthismoresensitiveassay,twoof thedeletions (Fig. 1B, lanes 7 and 10) were able to heterodimerize with EBNA-1(459-641), indicating that aa537 to 553 and 599 to 614aredispensablefordimerization.
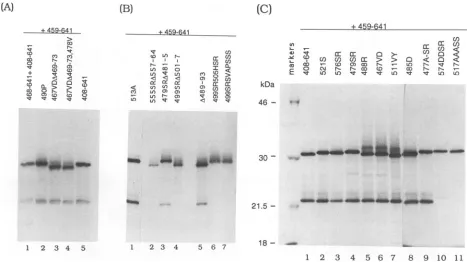
Identification of domains essential for EBNA-1 dimeriza-tion. To further localize the EBNA-1dimerization domain,
we constructed a series ofsmall deletions and point muta-tions inanEBNA-1(408-641) background (Fig. 2). Deletions are named with aA symbolfollowed by the numberof the first and last amino acid deleted. For point mutations, the numberof the firstamino acid altered is given followed by thesinglelettersymbolfortheintroduced aminoacid. Many of the mutations were created within EBNA-1-coding
se-quencescarryingauniqueXbaI restrictionsite,whichitself
introduces a serine and arginine (SR) mutation. (Note that
the XbaI siteatcodons 521and 522introducesonlyaserine
becausethe wild-type aa522 is arginine). None of the SR
mutations affected dimerization ability (Fig. 3Band C and datanotshown). Thenew series of EBNA-1mutantswere
cotranslated in vitro withEBNA-1(459-641)and immunopre-cipitated with EBNA.OTlx. The single exception to this protocolwasEBNA-1(468-641),which lacks theepitopefor the EBNA.OTlx antibody and was cotranslated with
EBNA-1(408-641). The results obtained with deletions and groupedmutationsarepresentedinFig. 3A and Band with point mutations predominantly in Fig. 3C. Each of the
Ln;
L-cU
Ln
IT
__<
mn U) U) U C)CD CD C;3 ) L.)
C0 U CC - N O U) O) Ln
St -' v-t S4
uc O Lt) to LO
U)U) U)-
-C~Ur < C to
) )CCl:C
uL) N Ul) UC c c) rm Ln CO c) C) L. U)N- LO
v C L)nv C
-wt
VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.135.467.72.294.2]4878 CHEN ET AL.
408 459 475 490 50!
1 1l
PVG-//- RKKGGWFGKHRGQGGSNPKFENIAEGLRALLARSHVERTTDEQTWVAG
--.
5 520
rVFVYQQSKTsLYNLRRGTALAIPQCRLT
_
-535 553
1 1
rPLSRLPFGMAPGPGPQPGP
---VD
---VD
_---VD-~----V--- ---SR ---A-SR
---s
---D---_______________________________________________ ---
R---SR
---SRM
l---SR--- HSR
--- SRSVAPSSS
---_---vY
--- A---
AAASS--
---
_-
_---_-SR--46B-641
467VD
467VDA469-73 467VDA469-738V 479SR
477A-SR
479SRA481-5 479SRA481-520
485D
488R
A489-93 490P 499SR 499SRA501-20 499SRA501-7 499SR505HSR
499SRSVAPSSS
511VY
513A 517AAASS 521S
A525-32 A537-53
554 570 585 600 615 630 641
l ~~~I l l lI I
LRBSIVCYFMVFLQTHIFAEVLkDAIKDLVMTKPAPTCNIRVTVCSFDDGVDLPPWFPPMVBQAAAEGDDGDDGDEGGDGDEGEEGQE
-SR--- 5555R -SR --- 555SRA557-64 -SR --- 5555RA557-75
---SR--- 576SR
---DDSR--- 574DDSR
---SR --- 576SRA578-616
---SR--- A599-614,615SR FIG. 2. Structure ofthedeletionsandpoint mutationsused in this study. The amino acid sequence of EBNA-1(408-641) is shown, with theamino acids subjectedto
mutagenesis
indicatedin bold type and the positions of deletions indicated by the stippled bars. Each mutant polypeptideisnamedintheright-hand
column,and the position and nature of the mutations carried by each is shown.constructions carryingdeletions located
between
aa459 and 493 retained the abilitytoheterodimerize (Fig. 3A, lanes 1,3, and 4, and 3B, lanes 3 and 5).
Point
mutations in thissegmentlikewise didnotaffect dimerization(Fig.3A, lane 2, and
3C,
lanes 5, 6, and 8). We had already shown that a downstream segment encompassingaa537 to553 was alsodispensable fordimerization (Fig. 1B, lane 7). However, a deletion located between these tworegions, 499SRA501-7,
abolished dimerization ability (Fig. 3B, lane 4). Further, mutation of these seven amino acids (499SRSVAPSSS) or even of aa
505
to 507(499SR505HSR)
also destroyed theability to dimerize (Fig. 3B, lanes 7 and 6, respectively).
Mutation ofnonhydrophobic residues immediately down-streamat aa511 and 512 andaa513 didnotaffect
dimeriza-tionability(Fig. 3C, lane7, and3B,lane1),butmutation of aa517to
520,
three of which arehydrophobicresidues,did abolish dimerization(Fig. 3C, lane11).Anadjacent deletion, A525-32, which was part ofthe previously described deletion set,also lacked dimerization ability (Fig. 1B, lane 5).There-fore, one of the dimerization domains involves residues locatedbetween aa 501 and 532.
Heterodimerization assayswith the set of large deletions hadimplicated a second region located between aa 554 and 598 in dimerization function (Fig. 1, lanes 8 to 10). The requirement for residues in this region was confirmed. A small
deletion,
555SRA557-64, abolished dimerization func-tion (Fig. 3B,lane
2), asdid a point mutation, 574 DDSR, whichchanged two hydrophobic residues (Fig. 3C, lane 10). The dimerization results are summarized in Table 1. Weconclude that EBNA-1has abipartite dimerization domain, withresidues located between aa 501 and 532 and between aa554 and 598beingessential.
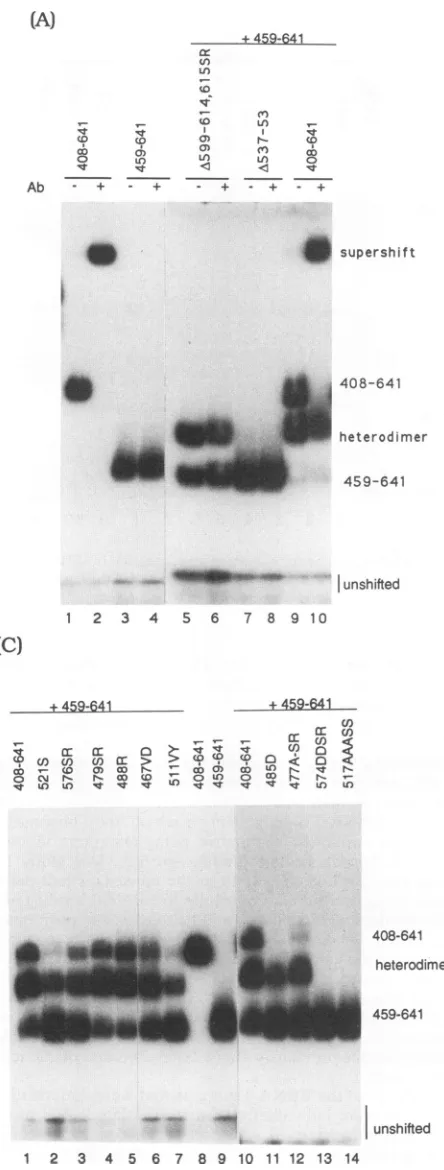
DNA binding by EBNA-1 heterodimers and specificity of
EBNA.OTix. The major goal of this studywas to separate the DNAbindingdomain of EBNA-1 from its dimerization domains. We firsttested DNAbindingof the new seriesof mutantsinthecontextofheterodimers. These experiments not only provided information on DNA-binding capability butalsoprovidedconfirmatory evidencefor
heterodimeriza-tion. Wedemonstrated that in vitro-translated EBNA-1(408-641) and EBNA-1(459-641) were capable of binding to a 32P-labeled 30-meroligonucleotide probe in a mobility shift assay(Fig.4A, lanes 1 and3).Theshiftedbandgenerated by
bindingof
EBNA-1(408-641)
was supershifted by theaddi-tion of the EBNA.OTlx antibody while, as expected, that generated bybinding of EBNA-1(459-641)was not(Fig. 4A, lanes 2 and 4). Thecotranslated polypeptides spanning aa 408 to 641 and 459 to 641 produced an additional shifted species (Fig. 4A, lane 9) of intermediate mobility, which resulted from the binding of heterodimers containing one subunit each of EBNA-1(408-641) and EBNA-1(459-641).
Interestingly, EBNA.OTlx supershifted the aa-408-to-641 homodimercomplexcompletely butwasunabletosupershift the heterodimer species (Fig. 4A, lanes 6 and 10). The
antibody apparently requires two epitopes to formastable
complex with DNA-bound EBNA-1 even though it can interact with, and immunoprecipitate, free heterodimers carryingonlyasinglecopyof theepitope.Thisrequirement J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.64.555.61.382.2]EBNA-1 DNA RECOGNITION AND DIMERIZATION 4879
(A)
(B)
(C)
+459-641
a)
co -c
O)co coN N.(
0 l
+ 0) 0)
,- < a
CD 0) CD CD 0
CO , CO a
+459-641 +±459-641
uz 03 F
It) X On
< V) V) Ur
co In a7) c
_ n r- CO CON v
(11n
a) (131
Cr) ° >
U})
CO0 cc
a:
0) 0) IT 0)0) < It
U,
4,
(a
E kDa
46- r X
C. CD 0
191
a: o >-U) (1) U)cc > >
- CD ) co
-c0rJrN CDC e) el) I' t T LO)
Ur
a) N N
L
t- 1
U-30 -
*mw
5mmmhmhI__ a21.5
-___-__ _
1 2 3 4 5 1 2 3 4 5 6 7 18- .
1 2 3 4 5 6 7 8 9 10 11
FIG. 3. Theeffect of small deletions(AandB)andpointmutations(C)onEBNA-1dimerization. ImmunoprecipitationbyEBNA.OTLx ofcotranslatedEBNA-1(459-641)and EBNA-1polypeptidescarryingtheindicated mutationswasperformedasdescribed in thelegendtoFig.
1.
maybebroughtaboutbythe
change
in conformation under-gone by EBNA-1 onbinding
(32).
The twooriginal large
deletions that could form heterodimers were also cotrans-lated with EBNA-1(459-641) and tested for DNAbinding.
Heterodimers between A599 to 614 and
EBNA-1(459-641)
werecapableof DNA
binding (Fig.
4A,lanes5 and6).
TheA537-553
polypeptide
remained unabletobind DNAeven as aheterodimer(Fig.
4A, lanes 7 and8).
This mayreflect a greaterperturbationofsecondary
structurebytheaa537to 553 deletion.Separation of the EBNA-1 DNAbinding domain from the
dimerization domains. Wenext examined the
ability
oftheseries of small deletions and
point
mutations inEBNA-1(408-641)tobindDNAasheterodimers with
EBNA-1(459-641). Dimerization is essential for DNA
binding.
The mu-tantsthat failedtoheterodimerize(Fig. 3;
Table1)
alsofailed tobind DNA(Fig.
4B,lanes2, 4, 7,and8;Fig. 4C,
lanes 13 and 14). The XbaI mutations(SR)
did not affect DNAbinding(Fig. 4C,lanes 2to4,and datanot
shown).
Twoof the mutations thatlay
within the boundaries of the firstdimerization domain but didnotaffect dimerization
ability,
511VY and
513A,
werecapable
ofDNAbinding
(Fig. 4C,
lane 7, and 4B, lane
1).
Ofgreatest interest were the fourconstructions carrying
deletions located betweenaa459and 485. Thesemutantpolypeptides
all failedtobind DNA(Fig.
4B, lanes 3, 9, 11, and
12), implicating
aminoacids in thisregion in DNA
recognition. Further,
anadjacent deletion,
A489-93, bound only as the heterodimer form and not as a
homodimer (Fig. 4B, lane
6),
suggesting
thatthese residuesmightcontributetothe
stability
of the DNA boundcomplex.
Asimilarresultwas obtained with the
point
mutation 485D(Fig. 4C, lane11). Other
point
mutations within thisregion
boundasboth homodimers and heterodimers
(Fig. 4B,
lane10, and 4C, lanes 5, 6, and 12).Theresults of these binding assays are summarized in Table 1. They indicate that an essential DNAbindingdomain of EBNA-1lies upstream of
thedimerization domains and islocatedbetween aa459 and 487.
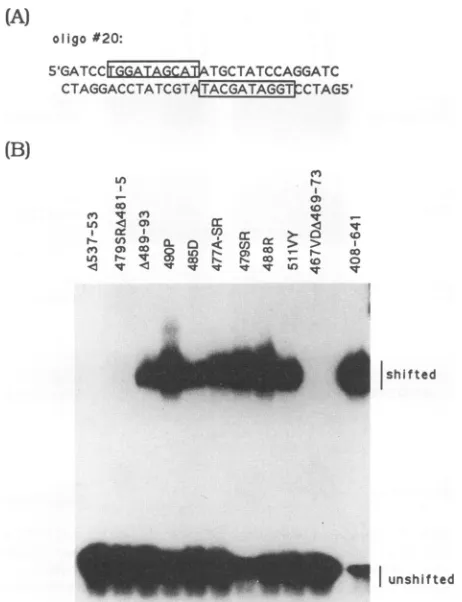
Differentialbindingto mutantoligonucleotidessuggests that aa 467, 468, and 477 may participate in DNA recognition. Previously,we had analyzed the precise sequence require-mentsfor EBNA-1 bindingusing a setof30-bp oligonucle-otides that contained symmetric point mutations in both
halves of the palindromic binding site (2). This study re-vealed that positions 9 and 10 in the consensus half-palin-dromic sequence didnotcontributetoEBNA-1binding and that considerableflexibilitycould betoleratedatpositions 1 and 2.Mutationsin positions3through8ofthe recognition
site had themostdeleterious effectonbinding.The EBNA-1 deletion constructions identified a DNA binding domain located betweenaa459 and 487. Togain further insight into theportionof this domain involved in DNArecognition,we examined theabilityof the different EBNA-1 constructions torecognizethepreviously characterizedmutant
oligonucle-otideprobes.
Firstly,10of the EBNA-1mutantsthatweredimerization competentwere individually invitro translated. The trans-lation products were analyzed on a denaturing polyacryl-amidegel(datanotshown),and the volumeswere adjusted
such thatequalamountsof
35S-labelled
proteinwereused in the subsequentmobility shift assays. Binding of these mu-tants totheconsensuswild-type oligomer20wasexamined inamobilityshift assay(Fig. 5). (The sequence ofoligomer20 is shown in Fig.
SA,
with both copies of the 10-bp consensushalf-siteboxed.)As was seenin the heterodimer-izationbindingassay(Fig. 4),there isagroupofmutantsthat VOL.67,1993no_ i
ask
WMI'amagaft
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.81.548.73.335.2]4880 CHEN ET AL.
+459-641
cc
_ n
IV
cn I?]
+ en c
LI) L/ 0
+459-641 +459-641
0
Li)N
cn CD N
Irc CO co
-)U) co I -eO> + (D
mc ro co 0
'-n q o) o a o N 10 Str < IT IT COt It CO I
supershift
408-641
heterod mer
459-641
....~
_ .unshifted1 2 3 4 5 6 7 8 9 10
(C)
+459-641 __+_459-641
(0 (D1a N o) 0 s 0t N
O cm N N 10O0CD -O 0 O N-T Lo X) -a ' W)e st "istt It Ln un
. 3W _ hA unshifted
[image:6.612.60.282.69.654.2]1 2 3 4 5 6 7 8 9 10 11 12 13
FIG. 4. DNAbindingofthemutantseriesof EBNA-1 polypep-tides cotranslated with wild-type EBNA-1(459-641). (A) Mobility shiftassayshowing binding of heterodimers and supershifting by EBNA.OTlx.Invitro-translated polypeptideswereincubatedwith
a32P-labeled symmetric 30-meroligonucleotide representinga wild-type EBNA-1-bindingsite (oligomer 20 [Fig. 5]) and subjectedto electrophoresisthrougha6%polyacrylamidegel.EBNA-1(408-641) andEBNA(1459-641)bothbindtotheprobe(lanes1 and3), but only EBNA-1(408-641) is supershifted by the addition ofEBNA.OTlx (lanes 2 and 4).Lanes 5 to10,binding of EBNA-1(408-641)and two mutantpolypeptidescotranslatedwithEBNA-1(459-641).The third shiftedspecies(heterodimer) results from bindingof aheterodimer between EBNA-1(459-641) and the wild-type or mutant EBNA-1(408-641)polypeptides. The effectofthe addition of EBNA.OTLx tothe boundcomplexes is showninlanes 6, 8 and10.Ab, antibody. (B and C) Mobility shiftassaysofEBNA-1polypeptides carrying small deletions and point mutations cotranslated with wild-type EBNA-1(459-641).
408-641 heterodimer
459-641
unshifted
3 4 5 6 7 8 9 10 11 12 13 14
cannot bind to the EBNA-1 target sequence. This group,
group I, includes A537-53, 479SRA481-5, and
467VDA469-73. All other EBNA-1 mutants bound oligomer 20with an affinitysimilartothat of thewild-typeEBNA-1(408-641).
Wethenperformed mobilityshift assaysto testbindingto a series ofoligomers that had mutations in the consensus binding site (boxed in Fig. 5A). To facilitate comparisons,
only the shifted complexes are displayed in Fig. 6 and 7. Also presented inthese figuresis the half-site sequence of the oligomer probes used, with the mutated nucleotides
underlinedand in bold type. When thehigh-affinityoligomer 74 was used, the binding pattern was verysimilar to that seen with the wild-type oligomer 20 (Fig. 6). The binding
behaviorof the group II mutants, 490P, 479SR and 488R, wasverysimilartothat ofwild-type EBNA-1(408-641) with all the oligomers presented in Fig. 6. Wild-type EBNA-1 binds oligomers 33, 104, 96, and 71 with an intermediate
affinity (2). However, several of the mutant polypeptides were completely unable to bind to these oligomer probes
(Fig. 6). This group, group III, includesA489-93, 485D, and 511VY. In contrast, 477A-SR was able to discriminate
(A)
(B)
(0 (t
CD
co
0
Ab - +
S
+459-641
,4-(0 LI
P-LO) 1 r
Ln v LI
= = cr
< LO L cn
C) L/) v) ) L/) '45-0
S
dM
1408-641
heterodimer
459-641
2
1 2
J.VIROL.
-_"
on November 9, 2019 by guest
http://jvi.asm.org/
EBNA-1 DNA RECOGNITION AND DIMERIZATION 4881
(A)
oligo#20:
51GATCCtGAIj
RabATGC
TATCCAGGATCCTAGGACCTATCGTA GT TCTAGS'
(B)
') n
c
en
q)(nI
Ln S aw a: S
en
C, S 8uD
r. ) co ' 'CD
a:: Go 0
4
shfte [image:7.612.69.300.68.369.2]unshifted
FIG. 5. ComparisonoftheDNA-bindingabilityofindividually translated wild-type and mutant EBNA-1
polypeptides.
(A) Se-quence ofthe oligomer 20 probe. The symmetric core EBNA-1-binding site is boxed.(B) Mobilityshift assaysusingoligomer20as theprobe.TABLE 1. Summary ofdimerization and DNA-binding characteristics of EBNA-1polypeptidesa
DNAbinding
EBNA-1polypeptide
Dimeri-zation Consensus Recognition
site"
characteristic408-641 + + Wildtype
468-641 +
-467VD + + Sitediscrimination
467VDA469-73 +
-467VDA469-73,478V +
-479SR + + Wild-typepattern
477A-SR + + Sitediscrimination
479SRA481-5 +
-479SRA481-520 -
-485D + + High-affinitysites
488R + + Wild-typepattern
A489-93 + + High-affinity sites
490P + + Wild-typepattern
499SR + +
499SRA501-520 -
-499SRA501-7 -
-499SR505HSR -
-499SRSVAPSSS -
-511VY + + High-affinitysites
513A + + Wild-typepattern
517AAASS
521S + +
A525-32 -
-A537-53 +
-555SR + +
555SRA557-64 -
-555SRA557-75 _ _
576SR + +
574DDSR -
-576SRA578-616 -
-A599-614,615SR + + Heterodimerbinds
aDatafromFig. 1 and 3to6.
bBindingtooligomer 20.
among the intermediate-affinity oligomer probes. It bound
oligomers33 and 104equallyaswellasthegroupII mutants butbound 96 moreweakly and failed tobind oligomer 71.
Surprisingly, 477A-SR appeared to show weak
binding
to oligomer 36, whichwas notbound under these conditions evenbythe wild-type protein. We classified 477A-SR as a group IVmutant. Themutation in 477A-SR that iscontrib-utingtoitsbinding phenotypeisclearlythe
lysine-to-alanine
changeatposition477. Bindingof theprotein carryingonly
theSR
mutation,
479SR,is shownalongside477A-SRinFig.
6, anditsbinding propertiesareindistinguishablefromthose ofwild-type
EBNA-1(408-641).
We next
compared
thebinding
pattern of another DNAbinding domain
point
mutant,467VD,
with that oftwoofthegroupIII mutantsand the group IVmutant477A-SR(Fig. 7).
The group III mutants,485D and511VY, againboundtothe
high-affinity oligomers20 and74, showedreducedbindingto thehigh-intermediate-affinity oligomer 102,anddidnotbind to any of the other
oligomers. Interestingly,
the mutant 467VD, like 477A-SR, discriminated among the differentintermediate-affinity oligomers. It bound strongly to oligo-mers73and 96 andweaklyto
oligomer
33, 71,and197, and it did not bind tooligomer
104.Although
477A-SR and 467VDshared this group IVbinding behavior, theirbindingaffinities for theindividualmutant
oligomers
werestrikingly
different. Mutant 467VD boundtooligomer
96 much more stronglythan did477A-SRandwasabletobindtooligomer
71, which wasnot boundby
477A-SR. On the otherhand,
477A-SR bound strongly to oligomers 33 and 104, which werebound only weakly or not at all by 467VD. A summary of the differentbinding
patterns
exhibitedbythedimeriza-tion-positiveEBNA-1 mutants is given in Table 2.
The mobility shift assays using individually translated mutant EBNA-1polypeptides and mutant oligomer-binding sites provided importantadditional evidence to support the
conclusion that the DNA domain of EBNA-1 is located between aa 459 and 487. Furthermore, the unique binding
patterns
demonstrated by 467VD and 477A-SR stronglyimplicate aa467, 468, and477 asbeing directly involved in DNArecognition.
DISCUSSION
EBNA-1is essential forreplicationof the latent episomal form of the
Epstein-Barr
virus genome and is one of severalproteins involved in the regulation of transcription fromthe latency C and Fpromoters. Further, EBNA-1 is the only viral protein expressed in the tumor cells of Epstein-Barr
virus-positive Burkitt's lymphoma (27). This observation raises the possibility that EBNA-1 may contribute to the
tumorigenicphenotype of these cells. The exact mechanism
bywhich EBNA-1performsany of itsbiologicalfunctions is
incompletelyunderstood. EBNA-1 does not possess any of theenzymaticactivities associated withreplicationproteins
such assimian virus 40 T antigen (1, 7, 20), and a
specific
VOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.317.557.93.424.2]4882 CHEN ET AL.
Uf)
In
co
a:)<O a cs
cn
a: r- ( ,) Q- a--: u) i >-< , t 0<t qr) U)
V)0 >- K
a0 > >C
OD N D at
aD r- wo
t
Costu4-CO
'4--t o
>O
fD
#20:
TGGATAGCAT #74
TGGATAGTAT
#74:
TGGATAGCTAT
Cl: >-::
< > >CL)
u) r- r- co
_oC. CD
#104:
TGGATAGCGC
#96:
TGGATTGCAT
.61
#33 TGIATAGCAT
#104
TGGATAGCGC
#96
TGGATlGCAT
#71 TGGAAAGCAT
#36 TGGATCGCAT
FIG. 6. Differential binding of individuallytranslatedwild-type
andmutantEBNA-1polypeptidestooligomer-bindingsitescarrying
symmetric basesubstitutions. DNAbindingwasassessedby
mobil-ityshift assays. To facilitate comparison, only theupper, shifted
DNA bandsarepresented. Thecoresequencesof themutant30-mer
probesare shown,with the altered bases in bold type and
under-lined. Theoligomer numbersarethose used inaprevious study (2).
An equal amount of EBNA-1 protein was used in each binding
reaction as judged by SDS-PAGE analysis of the radiolabeled
polypeptides.
transcriptionalactivation domain hasnotbeen documented. One property that is clearly an integral part of both its replication and transcriptional regulatory functions is the ability to bind specifically to consensus sequences within oniP and at the Q locus downstream of the latency F promoter(2, 15, 24). Proteinslackinganintact DNAbinding
domainareincapable ofsupportingeither transactivationor
replicationfunctions (22, 40).
EBNA-1 binds to a 16-bp palindromic recognition
se-quenceasadimer(1,8).Anearlieranalysisofanamino-and
carboxy-terminal deletion series locatedboth DNAbinding anddimerizationfunctionswithin theregionbetweenaa459
and607(1).TheavailabilityoftheEBNA.OTlx monoclonal antibodyandthe determination ofitsepitope location
pro-vided us with an opportunity to use
heterodimerization-immunoprecipitation as aprimary assaytoexamine dimer-ization ability. Confirmatory evidence for dimerizationwas
also obtained in the electrophoretic mobility shift assays using cotranslated EBNA(459-641) and the constructions spanningaa408to641(Fig. 4).Thepresenceof intermediate
#102:
TGGATAGCAC#73_
#73: TGGATAACAT
#33: *
'*06
S
a
#71:
TGGAAAGCAT
#197:
TGGATAGGAT
#196: TGCATAGCAT
FIG. 7. Comparison of bindingof group II(485D and511VY) and group IV (477A-SR and467VD)polypeptides to mutant oligomer probes. Mobility shift assayswereperformed asdescribed inthe legendtoFig.6.Onlytheupper,shifted DNA bandsarepresented.
mobility complexes in these assays is indicative of het-erodimer formation. For those mutants that were DNA-bindingnegative, dimerizationwasconfirmedby glutaralde-hyde cross-linking (data not shown). When applied to a series of mutant EBNA-1 polypeptides, these approaches led to theidentificationoftwosegments,aa501to532andaa 554to598, that contained motifsrequiredfor dimerization.
Mobility shift assays performed with this same series of EBNA-1 mutantsidentified components ofaDNAbinding domain between aa 459 and487, upstreamof the dimeriza-tion domains. The relative positioning and structures of these domains do not obviously conform to those of any previously defined families ofDNA-binding proteins, such asthehelix-turn-helix, helix-loop-helix,basic,coiled:coil,or zincfinger proteins (3, 4, 10, 11, 16, 17, 19).
The results of previous protease digestion experiments showed that theentiresegmentspanningaa459to607of the polypeptide adopts a highly ordered, protease-resistant structurewhen EBNA-1is boundtoDNA(32).This obser-vation is relevantto an understandingof the data obtained withthemutantEBNA-1 polypeptidesbecause the proper-ties ofanumber of them mayresult from the introductionof constraints on polypeptide structure. For example, muta-tions in theregion from eitheraa501to532or aa554to598
destroyeddimerization. Both domains maydirectly
partici-pateindimerization,or oneof theseregionsmay berequired to establish a conformation that allows the dimerization motifs located in the other domaintointeract. The glycine-proline-richsegmentbetweenaa537 and553wasspecifically susceptible to cleavage with high levels of protease (32), suggestingthat this segment of the DNA-boundproteinmay beexposedorhavealess-ordered conformation. The A537-553 variantwas notcapable of homodimerization in cross-J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
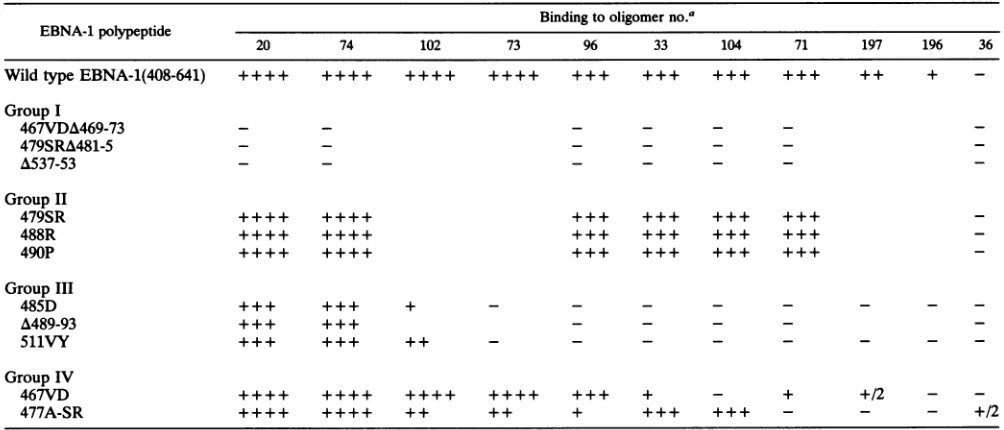
[image:8.612.311.552.67.325.2] [image:8.612.95.268.68.394.2]EBNA-1 DNARECOGNITION AND DIMERIZATION 4883 TABLE 2. Relative binding ofdimerization-positive EBNA-1 polypeptides to mutant oligomers
Binding tooligomer no.a
EBNA-1polypeptide
20 74 102 73 96 33 104 71 197 196 36
WildtypeEBNA-1(408-641) ++++ ++++ ++++ ++++ +++ +++ +++ +++ ++ +
-GroupI
467VDA469-73 - - -
-479SRA481-5 -
-A537-53 - - _ _ _ _
GroupII
479SR ++++ ++++ +++ +++ +++ +++
488R ++++ ++++ +++ +++ +++ +++
490P ++++ ++++ +++ +++ +++ +++
GroupIII
485D +++ +++ + - - -
-A489-93 +++ +++ _ _ _ _
511VY +++ +++ ++ - - -
-GroupIV
467VD ++++ ++++ ++++ ++++ +++ + - + +/2 -
-477A-SR ++++ ++++ ++ ++ + +++ +++ - - - +/2
a +++ , + + +++, and +/2indicate relative(high to low) levels ofEBNA-1binding; - indicatesnobinding;combinationsshowingneither+nor
-symbolswere nottested.
linking experiments (32) but was able to dimerize with a betweenaa459 and487(Fig. 8). Further,twoof the group III
wild-type aa-408-to-641 polypeptide in both coprecipitation mutants werelocatedadjacent to theright-handboundary of
assays (Fig. 1) and cross-linking experiments (data not this domain. Theleucine-to-aspartate change at aa485 and
shown). This deletionvariant was the single example of a the aa 489 to 493 deletion boundonly as heterodimers when
dimerization-positive,
DNA-binding-negative mutant that cotranslated with wild-typeEBNA-1(459-641), and althoughmapped outside oftheregion ofEBNA-1 fromaa459to487. they bound normally to wild-type probe when individually Inview of itsprotease susceptibility, itseemsunlikelythat translated, they failed to bind to any of the intermediate-theaa537to553region contributes directlytoDNAbinding. affinity mutant probes. The loss of binding to these latter
The lack ofDNA-binding abilitymaybe areflection of the probes was unrelated to the position of the mutated
nucle-destabilizationin dimerizationfunctionthat is apparent from otide withinthe binding site and hence is unlikely to involve theinabilityofthispolypeptidetoform homodimers. Prece- direct DNAcontacts. Leucine 485 and aa 489 to 493 may be dence for such a phenotype comes from studies of E47 in contributing to a conformation that favors DNAinteractions which a variant that could form heterodimers but not ho- with the recognitionamino acids. Protease digestion exper-modimers also lacked DNA-binding ability in the het- imentsindicated that the region spanning aa 459-607 under-erodimer form (37). Dimerization structures involvingboth goes a conformational change on binding (32). This struc-a-helical (6, 10, 23, 30) and
n-sheet
(12) interactions have turalrearrangement maybeessential to maximizecontacts been described. There arepredicted
n-sheet structures between thepolypeptide
and DNA interfaces. If the group within both of the EBNA-1 dimerization segments and a IIImutations interfere with
theability
tomakethis
confor-predicted
ahelix in the secondaa 554-598region
(Fig. 8).
mational
change,
then the DNA contacts madeby
these The loss of DNAbinding
demonstratedby
three of thepolypeptides might
besufficiently
destabilized thatbinding
deletionmutantsservedtolocateaDNA
recognition
domain could occuronly
to high-affinity-binding sites. The othergroup III mutant, 511VY, lies 18 aadownstream and con-verts two glycine residues to hydrophobicvaline and
ty-EBNA-1
I-
-sis DNAbindn
641 rosine residues. It should be noted that polypeptidescon-taining other nonconservative amino acid mutations in the samevicinity, namely leucine 488toarginineand alanine 490
459
00to
proline bind the mutant oligomers indistinguishably fromPredictedStructure - - -- - the
wild-type polypeptide.
Dimerizaion a The behavior of the group IVmutantsis also
particularly
DNABinding 501 S32 554 598
interesting
because itimplies
that theseparticular
aminoI59
487 acids may be involveddirectly
in DNArecognition. [image:9.612.63.565.82.297.2]Conver-RKKGGWFGKHRGQGGSNPKFENIAEGLRA sion of aa 467 and 468 from lysine and histidine to valine and FIG. 8. Location of the dimerization andDNA-bindingdomains
aspartate
(467VD)
and aa477 fromlysine
toalanine (477A-FIG. 8...Loato of th.ieiainad N-idnoan SR) resulted in EBNA-1 polypeptides that were able to of EBNA-1. The predicted secondary structure (5) of the region . . . . .,spanningaa459 to607 is shown. The amino acidsequence of the
discriminate
betweendifferent
intermediate-affinity
DNA DNArecognitiondomain is alsoprovided alongwith thepositionsof probes. Acomparisonof relativebindingshowed that while the deletions that abolish DNA binding (stippled bars) and the 467VD bound to themajorityof the mutantoligomerswitha substitutionsthataffect DNArecognition (*). higher affinitythan 477A-SR(Fig. 7;Table2),thereweretwo VOL. 67,19931
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.66.305.573.662.2]4884 CHEN ET AL.
notable
exceptions; oligomers
33 and104,
whichhave mu-tationsatposition
8 and atpositions
1 and2, respectively.
Anexaminationof the
binding
of 477A-SR(Fig. 6;
Table2)
showed that this
polypeptide,
which bound aswell as thewild-type
protein
tooligomers
33 and104,
boundonly
weakly
tooligomer
96anddidnotbindatalltooligomer
71.Oligomer
96carriesanucleotidechange
atposition
5 ofthebinding site,
andoligomer
71 is altered atposition
6.Fur-thermore,
the aa-477mutant wasabletobind,
albeitweakly,
toan
oligomer (no. 36) carrying
amutationatposition
5thatwas not bound
by
thewild-type protein.
EBNA-1-binding
site
recognition
ismostlikely
mediatedby
acombinationof contactswith bases in themajor
groove(2, 18)
andcontacts with thephosphate
backbone. Thecontributionto recogni-tion ofthelysine
and histidine at aa 467 and 468 and thelysine
ataa477canonly
bespeculated
uponatthistime.The mostdeleteriouschanges
forbinding
of 467VDwerelocated atdisparate positions
inthebinding site,
positions 1, 2,
and8,
and the interactionsinvolving
these amino acids would appear to becomplex.
Because the mutations that moststrongly
affectedbinding
of 477A-SRwere each located at nucleotide5 or6 of thebinding
site,
the contacts madeby
lysine-477
maytherefore involve thesepositions.
A
large
number ofDNA-binding
proteins
utilize a DNArecognition
motifthat is analpha
helix. The E2transcrip-tional
regulator
ofpapillomavirus
and the GCN4bZIPpro-tein contain such a motif
(6, 12)
as do the helix-turn-helixproteins.
Examples
of thislattergroupincludemanyof theprokaryotic transcriptional regulatory proteins
and theeu-karyotic
homeodomainproteins
(reviewed
in references 8 and11).
Ontheotherhand,
both the yeast met repressor andthe
TFIID
TATA-boxbinding protein
utilizeanti-parallel
13 strandsor1 ribbons asthe DNArecognition
motif(21, 34).
The EBNA-1 DNA
binding
domain identified in thisstudy
overlaps
withapredicted
ahelix,
buttheexactnatureof the EBNA-1 DNArecognition
motifremainstobeelucidated.ThebHLH groupof
proteins
containabasic DNArecog-nitiondomain andan
adjacent
dimerization domainconsist-ing
oftwoamphipathic alpha
helicesseparated by
avariableloop
that is 9 to 13 aainlength
(4, 10, 37). Comparison
ofproteins
within this group has identified a consensus se-quence for the bHLH motif(4). Using
computeranalysis,
Inoue et al.
(13)
matched this consensus to aregion
ofEBNA-1betweenaa465 and 587 and
suggested
onthe basis of this amino acidalignment
that EBNA-1might belong
to the bHLHclass.Thedegree
ofmatchtothe bHLHconsen-sus was
intriguing
butimperfect.
Theproposed
basicregion
of EBNA-1 contained a three of six match to the bHLH consensus,andthematches for helix 1 and helix 2werethree
ofsix and fiveof
eight, respectively,
whiletheloop region
wouldcontainanextended 83aa.The behaviorofanumber of the EBNA-1 variants does not appearto be
compatible
with the bHLH model. This isparticularly
true of the EBNA-1 mutants in theregion
(aa
477to493)
ofpredicted
helix 1.Deletion offiveaa
(aa
481to485)
from thecenterof the helixremoves two ofthe consensushydrophobic
resi-dues anddestroys
the 4:3:4hydrophobic pattern
which would create thehydrophobic
interface between the pre-dictedalpha
helices.Nonetheless,
theA481-485mutantwas dimerizationcompetent. We convertedtwooftheconsensushydrophobic
residues tocharged
residues(Leu-485
toAsp and Leu-488toArg).
Mutationof theequivalent
residues in the bHLHprotein
E47 fromhydrophobic
tocharged
abol-isheddimerizationof E47(37),
but these mutations hadno effect on dimerization of EBNA-1. We also introduced ahelix-breaking proline
residue into thepredicted
helix 1at aa490 and foundthat this mutation had no effect on dimeriza-tion. Further, mutations in the loop region of a bHLH protein have been shown not to affect dimerization (37), whereas two separate grouped mutations (499SR505HSR
and 517AAASS) in the predicted loop of EBNA-1 each abolished dimerization. The DNA recognition
region
mapped in our experiments, aa 459 to 487, does in fact
overlap with the basic domain predicted by the sequence comparison, and the second dimerizationdomain,aa554to 598, overlaps with the predicted helix 2. However, the
experimental data is not compatible with a dimerization domainbeing located in the region of predicted helix 1.Since
thespacing between the basic DNA-binding domain and the firsthelix of the dimerization domain is constant in bHLH proteins, this finding would argue against the inclusion of EBNA-1 within thisprotein family. The partial amino acid match to the bHLH consensus within the EBNA-1 DNA recognition domain and the second dimerization domain may represent convergent evolutionary selection of particular
amino acidcombinations that favor thetype of interactions involved in DNAbinding and protein-protein contacts.
ACKNOWLEDGMENTS
We thank Mabel Chiu and J.-C. Zong for excellent technical assistance and SarahHeaggans for manuscript preparation.
This workwas supported by Public Health Service grantsRO1 CA42245andRO1 AI31454 from the National Institutes of Health.
REFERENCES
1. Ambinder,R.F.,M.Mullen, Y. N. Chang, G.S. Hayward, and S. D.Hayward. 1991.Functional domains ofEpstein-Barr virus nuclearantigen EBNA-1. J. Virol. 65:1466-1478.
2. Ambinder,R.F.,W.A.Shah,D. R. Rawlins, G.S.Hayward, and S. D. Hayward. 1990. Definition of the sequence require-ments for binding of the EBNA-1 protein to its palindromic targetsites inEpstein-Barr virus DNA. J. Virol. 64:2369-2379. 3. Berg, J. M. 1988. Proposed structure for the zinc-binding domains from transcription factor IIIA and related proteins. Proc. Natl. Acad.Sci. USA 85:99-102.
4. Blackwood,E.M.,and R. N.Eisenman. 1991. Max:a helix-loop-helixzipper protein that formsasequencespecific DNA binding complexwithmyc.Science251:1211-1217.
5. Chou, P. Y.,and G. D. Fasman. 1974. Prediction of protein conformation.Biochemistry13:222-245.
6. Ellenberger,T.E., C. J. Brandl,K.Struhl,and S. C.Harrison. 1992. The GCN4basicregion leucinezipper binds DNAasa dimer ofuninterrupted ahelices: crystalstructure of the pro-tein-DNAcomplex. Cell 71:1223-1237.
7. Frappier, L.,and M. O'Donnell. 1991.Overproduction, purifi-cation, and characterization of EBNA-1, the origin binding protein ofEBV. J.Biol.Chem. 266:7819-7826.
8. Frappier, L., and M. O'Donnell. 1991. Epstein-Barrvirus nu-clearantigen1mediates aDNAloop within the latent replica-tionorigin ofEpstein-Barrvirus.Proc. Natl. Acad. Sci. USA 88:10875-10879.
9. Gahn, T. A., andC. L. Schildkraut. 1989. The Epstein-Barr virus origin of plasmid replication, ori-P, contains both the initiationandtermination sites of DNAreplication. Cell 58:527-535.
10. Harrison, S. C. 1991.AstructuraltaxonomyofDNA-binding domains.Nature(London) 353:715-719.
11. Harrison,S.C.,and A. K.Aggarwal.1990.DNArecognition by proteins with the helix-turn-helix motif. Annu. Rev. Biochem. 59:933-969.
12. Hegde,R.S.,S. R.Grossman,L. A.Laimins, and P. B.Sigler. 1992.Crystalstructure at1.7 A of thebovinepapillomavirus-1 El DNA-binding domain bound to its DNA target. Nature (London)359:505-512.
13. Inoue,N., S. Harada, T. Honma, T. Kitamura, and K. Yanagi. 1991. The domain of Epstein-Barr virus nuclear antigen 1 J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
EBNA-1 DNA RECOGNITION AND DIMERIZATION 4885 essential forbindingtooriPregion hasasequencefitted for the
hypothetical basic-helix-loopstructure.Virology 182:84-93. 14. Jankelevich, S., J. L. Kolman, J.W. Bodnar, and G. Miller.
1992. Anuclear matrix attachmentregionorganizes the Epstein-Barr viral plasmid in Raji cells into a single DNA domain. EMBO J. 11:1165-1176.
15. Jones,C.H.,S.D.Hayward,and D. R. Rawlins.1989. Interac-tion oflymphocyte-derived Epstein-Barr virus nuclear antigen EBNA-1withits DNAbinding sites.J.Virol. 63:101-110. 16. Kaptein, R. 1991. Zinc-finger structures. Curr. Opin. Struct.
Biol. 2:109-115.
17. Kerppola, T. K., and T. Curran. 1991. Transcription factor interactions: basicsonzippers. Curr. Opin.Struct. Biol. 1:71-79.
18. Kimball, A. S., G. Milman, and T. D. Tullius. 1989. High resolution footprints of theDNA-binding domain of Epstein-Barr virus nuclearantigen-1. Mol. Cell Biol. 9:2738-2742. 19. Landshulz,W.H., P. F. Johnson, andS.L.McKnight.1988. The
leucinezipper:ahypotheticalstructurecommon to a newclass ofDNAbindingproteins. Science 240:1759-1764.
19a.Middeldorp, J. M., M.-R Chen,and S.D. Hayward. Unpub-lished data.
20. Middleton, T., and B. Sugden. 1992. EBNA-1 can link the enhancer elementto the initiator element of theEpstein-Barr virusplasmid origin ofDNAreplication.J.Virol. 66:489-495. 21. Nikolov, D.B., S.-H. Hu,J. Lin,A. Gasch, A.Hoffmnann, M.
Horikoshi, N.-H.Chua,R.G. Roeder,andS.K.Burley.1992. CrystalstructureofTFIID TATA-boxbinding protein. Nature (London)360:40-46.
22. Polvino-Bodnar, M., and P. A. Schaffer. 1992. DNA binding activity is required forEBNA-1-dependent transcriptional acti-vationand DNAreplication. Virology 187:591-603.
23. Rasmussen,R., D. Benvegnu, E. K.O'Shea,P.S.Kim, and T. Alber.1991.X-rayscattering indicates that the leucinezippers is acoiled coil.Proc.Natl. Acad. Sci. USA88:561-564. 24. Rawlins, D. R., G.Milman, S.D.Hayward,andG.S.Hayward.
1985. Sequence-specificDNAbinding ofEBNA-1toclustered sites in theplasmidmaintenanceregion. Cell 42:859-868. 25. Reisman, D., and B. Sugden. 1986. trans activation of an
Epstein-Barr viraltranscriptionalenhancerbytheEpstein-Barr viral nuclearantigen 1. Mol. Cell Biol. 6:3838-3846.
26. Reisman, D.,J. Yates,and B.Sugden. 1985.Aputative origin of replication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol. 5:1822-1832.
27. Rowe, M., D. T. Rowe, C. D. Gregory, L. S. Young, P. J. Farrell, H.Rupani,and A. B. Rickinson. 1987. Differencesin B cell growth phenotype reflect novel patterns of Epstein-Barr viruslatentgene expressionin Burkitt'slymphoma. EMBO J. 6:2743-2751.
28. Sample, J., L. Brooks, C. Sample, L. Young, M. Rowe, C. Gregory, A. Rickinson, and E. Kieff. 1991. Restricted EBV
protein expression in Burkitt lymphoma is due to a different Epstein-Barr nuclear antigen 1 transcriptional initiation site. Proc.Natl. Acad.Sci. USA 88:6343-6347.
29. Sample, J., E. B. D. Henson, and C. Sample. 1992. The Epstein-Barr virus nuclear protein1 promoteractivein type 1latency is autoregulated. J.Virol.66:4654-4661.
30. Sauer, R. T., R. Jordan, andC.0.Pabo. 1990. X Repressor:a modelsystemforunderstanding protein-DNAinteractions and proteinstability. Adv. Protein Chem.40:1-61.
31. Schaefer, B. C., M.Woisetschlaeger,J. L. Strominger, and S. H. Speck. 1991.Exclusiveexpression of EBV nuclear antigen1in Burkitt lymphoma arises from athirdpromoter,distinct from the promotors used in latently infected lymphocytes. Proc. Natl. Acad. Sci. USA 88:6550-6554.
32. Shah, W. A., R. F. Ambinder, G. S. Hayward, and S. D. Hayward.1992. Binding of EBNA-1toDNA creates a protease-resistant domain that encompasses the DNArecognition and dimerizationfunctions. J. Virol. 66:3355-3362.
33. Smith, P. R., and B. E. Griffin. 1992. Transcription of the Epstein-Barrvirus gene, EBNA-1from differentpromoters in nasopharyngeal carcinoma andB-lymphoblastoid cells. J. Virol. 66:706-714.
34. Somers,W.S., and S. E. V.Phillips.1992.Crystalstructureof themetrepressor-operatorcomplexat2.8Aresolution reveals DNArecognition by 3-strands.Nature(London) 359:387-393. 35. Su, W.,T. Middleton, B. Sugden,and H. Echols. 1991. DNA
looping between theorigin of replication of Epstein-Barr virus and its enhancer site: stabilization of anorigin complex with Epstein-Barr nuclear antigen 1. Proc. Natl. Acad. Sci. USA 88:10870-10874.
36. Sugden,B., and N. Warren. 1989.A promoterofEpstein-Barr virus that canfunction duringlatent infectioncanbe transacti-vated by EBNA-1, a viral protein required for viral DNA replication during latent infection. J.Virol.63:2644-2649. 37. Voronova, A., and D. Baltimore. 1990. Mutations that disrupt
DNAbinding and dimer formation in the E47 helix-loop-helix proteinmap todistinct domains. Proc. Natl. Acad. Sci. USA 87:4722-4726.
38. Wysokenski, D. A., and J.L. Yates. 1989. Multiple EBNA1-binding sites are required to form an EBNA1-dependent en-hancerand toactivateaminimal replicative origin withinoriP of Epstein-Barr virus. J. Virol. 63:2657-2666.
39. Yates, J., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci.USA81:3806-3810. 40. Yates, J. L., and S. M. Camiolo. 1988. Dissection of DNA
replication and enhancer activation functions of Epstein-Barr virusnuclear antigen1. CancerCells 6:197-205.
41. Yates, J. L., N.Warren, and B. Sugden.1985. Stable replication of plasmids derived from Epstein-Barr virus in a variety of mammalian cells. Nature(London) 313:812-815.
VOL.67,1993