0022-538X/08/$08.00⫹0 doi:10.1128/JVI.00951-08
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Nuclear Pore Composition and Gating in Herpes Simplex
Virus-Infected Cells
䌤
Helmut Hofemeister† and Peter O’Hare*
Marie Curie Research Institute, The Chart, Oxted, Surrey RH8 0TL, United Kingdom
Received 7 May 2008/Accepted 11 June 2008
The mechanism by which herpes simplex virus (HSV) exits the nucleus remains a matter of controversy. The generally accepted route proposes that capsids exit via primary envelopment at the inner nuclear membrane and subsequent fusion of this primary particle with the outer nuclear membrane to gain capsid entry to the cytoplasm. However, recent observations indicate that HSV may induce gross morphological alterations of nuclear pores, resulting in the loss of normal pores and the appearance of dilated gaps in the nuclear membrane of up to several 100 nm. On this basis, it was proposed that a main route of capsid exit from the nucleus is directly through these altered pores. Here, we examine the biochemical composition of some of the major nuclear pore components in uninfected and HSV-infected cells. We show that total levels of major nucleoporins and their sedimentation patterns in density gradients remain largely unchanged up to 18 h after HSV infection. Some alteration in modification of one nucleoporin, Nup358/RanBP2, was observed during enrichment with anti-nucleoporin antibody and probing for O glycosylation. In addition, we examine functional gating within the nucleus in live cells, using microinjection of labeled dextran beads and a recombinant virus expressing GFP-VP16 to track the progress of infection. The nuclear permeability barrier for molecules bigger than 70 kDa remained intact throughout infection. Thus, in a functional assay in live cells, we find no evidence for gross perturbation to the gating of nuclear pores, although this might not exclude a small population of modified pores.
Herpes simplex virus (HSV), like all herpesviruses, repli-cates and packages its genome into newly formed capsids in-side the nucleus of infected cells. The nucleus is bounded by a nuclear envelope, a double lipid bilayer composed of the outer and inner nuclear membrane (ONM and INM, respectively), which is underpinned on the nucleoplasmic side by the nuclear lamina, a dense meshwork of intermediate filaments formed from interlaced dimers of the lamins A/C and B. Transport of molecules between the nucleus and cytoplasm normally takes place via nuclear pores embedded within and connecting the INM and ONM. However, progeny nucleocapsids, with a size of 100 nm are too large to pass through normal nuclear pores, which have a gating mechanism for soluble proteins and as-semblies through aqueous channels with a diameter of about 10 nm (reviewed in references 1, 35, 46, and 47). It has been generally accepted therefore that a primary pathway of exit is via nucleocapsid attachment to the INM and subsequent bud-ding into the luminal space, thereby acquiring a primary lipid envelope (reviewed in references 3, 9, 26, and 43). However, the mechanism by which HSV exits the nucleus remains a matter of controversy (27, 49). An alternative pathway of cap-sid exit from the nucleus to the cytoplasm was proposed on the basis of high-resolution scanning and transmission electron microscopy of both HSV- and bovine herpesvirus-infected cells (22, 50). These analyses indicated that infection resulted in
gross enlargement of nuclear pores through which nuclear content, including capsids, was transported to the cytoplasm. It was concluded that capsids use impaired nuclear pores as gate-ways to gain access to the cytoplasmic matrix. Although the weight of evidence strongly favors nuclear exit by the deenvel-opment-reenvelopment model, many issues remain to be re-solved (27, 49).
We have previously reported changes at the INM after HSV infection, exemplified by altered diffusional mobility of the lamin B receptor; dissociation of a population of lamin A/C from the lamina (40); and hyperphosphorylation of a major nuclear membrane protein, emerin (29). Alterations in nuclear structure, the nuclear lamina, and the INM were also reported in subsequent studies (2, 20, 33, 37, 41, 42) and have also been observed during cytomegalovirus replication (30). These alter-ations are likely to underpin major conformational changes in the nuclear envelope associated with access of the emerging capsid to the INM and the relevant virus-host interactions, in particular involving the essential proteins UL34 and UL31 (12, 16, 17, 38, 39, 41).
In the present study, expanding on comparative analysis of nuclear components, we examine some of the major nuclear pore constituents in infected versus uninfected cells and un-dertake a functional analysis of nuclear gating in live infected cells. We found no major perturbations in the total levels of major nucleoporins after infection or gross effects on sedimen-tation profiles in density gradients. Although other subtle changes may occur, we found no evidence for impairment or alteration in gating function of nuclear pores, as evidenced by the exclusion of different-sized dextran beads in live infected cells. Although qualifications may always be necessary, and local loss of nuclear pores could occur without overall affect on
* Corresponding author. Mailing address: Marie Curie Research Institute, The Chart, Oxted, Surrey RH8 0TL, United Kingdom. Phone: 44 01883 722 306. Fax: 44 01883 714 375. E-mail: P.OHare @mcri.ac.uk.
† Present address: BIOTEC, TU-Dresden, Germany.
䌤Published ahead of print on 18 June 2008.
8392
on November 8, 2019 by guest
http://jvi.asm.org/
gating function, these results indicate that there is no discern-ible alteration in pore function that would be expected from broad dismantling and loss of pores from the membrane.
MATERIALS AND METHODS
Tissue culture.Vero and Hep2 cells were maintained in Dulbecco modified Eagle medium (DMEM) supplemented with 100 U of penicillin/ml, 100g of streptomycin/ml, and 10% newborn calf serum (NBCS) at 37°C in a 5% CO2
environment. Infections were routinely performed in 35-mm dishes (2⫻105
cells/well) using HSV-1[17] at a multiplicity of infection (MOI) of 10 PFU/cell in DMEM containing 2% NBCS. Mock-infected cells were incubated in DMEM containing 2% NBCS without virus. After adsorption of virus for 1 h at 37°C, the medium was replaced with DMEM containing 10% NBCS, and the cells were processed at the times indicated.
Immunolocalization.For immunolocalization studies, cells were plated and infected on 16-mm borosilicate glass coverslips (BDH) placed in 35-mm cell culture dishes (Falcon). Cells were fixed in an ice-cold mixture of equal volumes of ethanol and acetone for 15 min, rinsed, and permeabilized in phosphate-buffered saline (PBS) containing 50 mM NH4Cl, 0.5% Triton X-100, and 0.1%
sodium dodecyl sulfate (SDS) for 10 min. The coverslips were further blocked in PBS containing 10% goat serum for 20 min and incubated for 20 min with anti-nucleoporin antibody MAb414 (Covance) diluted 1:2,000. After a wash in PBS, bound antibodies were detected using fluorescent-labeled secondary anti-body (Molecular Probes). Coverslips were mounted in Mowiol (Sigma). Images were routinely acquired by using a Zeiss LSM 410 confocal microscope with a Plan-Apochromat⫻63 oil immersion objective lens (NA 1.4) and zoom factors ranging from 1 to 8 of the LSM 410 acquisition software.
Microinjection and live cell analysis.Monitoring the permeability of the nuclear envelope during infection was performed as described in previous studies on NPC gating during the cell cycle (21). Vero cells were plated in 35-mm plastic dishes which contained a gridded, glass insert, enabling the localization and recovery of individual cells. Cells were infected with an 1 strain, HSV-1[V41], expressing green fluorescent protein (GFP)-VP16 (19) at an MOI of 5. Prior to injection, TRITC (tetramethyl rhodamine isothiocyanate)-labeled dex-tran beads (Molecular probes), with molecular masses of 10 or 70 kDa, were diluted to 1 mg/ml in injection buffer (100 mM glutamate, 10 mM HEPES [pH 7.6]) and centrifuged at 10,000⫻gfor 30 min (21). Using a semiautomatic microinjector 5170 (Eppendorf) fitted to a Zeiss Axiovert 200 microscope, the beads were injected into the cytoplasm of individual uninfected cells or cells 6 h after infection with HSV[V41]. For monitoring the progress of infection and imaging, the dishes were transferred to a heated stage of the inverted LSM410 confocal microscope and monitored at different times thereafter. The intracel-lular localization of the TRITC-labeled dextran beads and GFP-VP16 expression in individual cells could then be recorded as described above.
Sample preparation and Western blotting.Mock-infected or HSV-1-infected cells (plated in 60-mm dishes) were washed in ice-cold PBS; harvested in 1 ml of ice-cold PBS buffer containing 2 mM EDTA, 1 mM Na3VO4, 1 mM
phenyl-methylsulfonyl fluoride, 1 mM dithiothreitol (DTT), and Complete protease inhibitor mix (Roche Diagnostics); and pelleted at 2,000 rpm for 5 min at 4°C. The cell pellets were subsequently lysed in 200l of radioimmunoprecipitation assay (RIPA) buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 1.0% NP-40, 0.5% sodium deoxycholate, 2 mM EDTA, 1 mM Na3VO4, 1 mM
phenylmethyl-sulfonyl fluoride, 1 mM DTT, 10 nM okadaic acid, and Complete protease inhibitor mix). Homogenates were incubated on ice for 5 min and then briefly sonicated and centrifuged at 10,000 rpm for 5 min at 4°C. SDS-polyacrylamide gel electrophoresis (PAGE) and Western blotting were performed according to standard methods. The samples were denatured in SDS sample buffer at 45°C for 15 min and generally separated on Tris-acetate gradient gels using 4 to 8% polyacrylamide (Invitrogen). After electrophoresis, the proteins were transferred onto Immobilon-P membranes (Millipore Corp.), which were blocked by incu-bation in methanol according to the manufacturer’s protocol and then dried for 15 min at room temperature prior to incubation with primary antibody in PBST (PBS with 0.1% Tween 20) with 0.5% bovine serum albumin. For immunode-tection, the membranes were incubated overnight with the following primary antibodies in PBST with 0.5% bovine serum albumin diluted as follows: MAb414, 1:5,000; anti-O-GlcNAc monoclonal antibody (Covance), 1:5,000; and anti-VP5 antibody (Virusys), 1:2,000. After a washing step, the membranes were incubated in PBST containing horseradish peroxidase-conjugated anti-mouse or anti-rabbit immunoglobulin G secondary antibodies (Bio-Rad), and proteins were detected by using the ECL West Pico reagent (Pierce).
Immunoprecipitation.Clarified cell homogenates prepared as described above were diluted in ice-cold RIPA buffer supplemented with 0.5l of antibody MAb414 or 20l of wheat germ agglutinin (WGA)-Sepharose (Vector Shield) and then incubated overnight at 4°C. The samples containing MAb414 or control samples were supplemented with 20l of protein G-Sepharose and further incubated for 2 h at 4°C with gentle rotation. Protein complexes were isolated by centrifugation 2,000 rpm for 5 min, washed three times in RIPA buffer, and analyzed by SDS-PAGE and Western blotting.
Nuclear envelope sucrose gradient fractionation.Mock-infected and infected (MOI⫽10) Vero cells were harvested at 16 h postinfection (hpi), washed, and centrifuged at 2,000 rpm for 5 min. The cells were then suspended in 1 ml of ice-cold hypotonic buffer (50 mM HEPES [pH 8], 5 mM MgCl2, 2 mM DTT,
0.025% NP-40, and Complete protease inhibitor mix), incubated for 15 min on ice, and homogenized by 30 strokes in a glass Dounce homogenizer. Isolation of nuclear envelopes was performed according to the method of Georgatos and Blobel (13) with modifications as described previously (24). After preparation, the samples were further treated with DNase I for 1 h at 4°C and subsequently centrifuged at 10,000⫻gfor 20 min. The pellet containing nuclear envelopes was resuspended in 700l of gradient buffer (50 mM HEPES [pH 8], 20% sucrose, 150 mM NaCl, 2 mM MgCl2, 0.1 mM EGTA) and transferred to centrifugation
tubes preloaded with equal volumes of gradient buffer containing 70, 50, and 30% sucrose. Gradients were subject to ultracentrifugation at 100,000⫻gfor 8 h in a Beckman AH650 rotor. After centrifugation, the gradients were fractionated from top to bottom, and aliquots of single fractions were analyzed by Western blotting with the appropriate anti-nucleoporin antibodies.
RESULTS
Examination of nucleoporins in virus-infected cells. Al-though recent work from several laboratories has begun to address alteration to the nuclear membrane and lamina during infection, there has been no work exploring biochemical com-position and gating at the nuclear pore during infection. To begin addressing these issues, we wanted to examine whether HSV infection induced any gross compositional changes to some of the major constituents of nuclear pore complexes (NPCs). The NPC is a dynamic, multiprotein complex prised of different nucleoporins, together with additional com-ponents assembled into a very large complex. The NPC spans, and is anchored to, the nuclear membrane and attaches to cytosolically disposed filaments and a nucleoplasmic basket (8, 10, 11, 15, 23, 35, 46, 51). Many of the core nucleoporins are composed of FG-repeats, clusters of hydrophobic phenylala-nine-glycine amino acid repeats, which are thought to form a permeability barrier and enable selective gating through the nuclear pore (11, 34, 45, 48). The FG-repeat containing nucleoporins may also function as adaptor proteins for differ-ent specific transport pathways. Although the NPC has been regarded as a static channel through which dynamic transport of cargos takes place, more recent evidence indicates the pore complex is itself a dynamic complex whose composition may be regulated.
To begin to probe for alterations in the major NPC constit-uents, we made use of a well-characterized nucleoporin-spe-cific antibody MAb414 (5), together with additional compo-nent-specific antibodies. The MAb414 antibody specifically recognizes a subset of the more prominent FG-repeat contain-ing nucleoporins, includcontain-ing nucleoporin 358 (Nup358), local-ized on the cytoplasmic face, Nup153 and Nup214 on the nucleoplasmic face, and Nup62 (p62), which is present in the central channel of the NPC. We also examined NPC compo-sition by using antibodies to the membrane spanning proteins pom121 and gp210 and a separate specific antibody to Nup62. In the first series of experiments, we analyzed the
on November 8, 2019 by guest
http://jvi.asm.org/
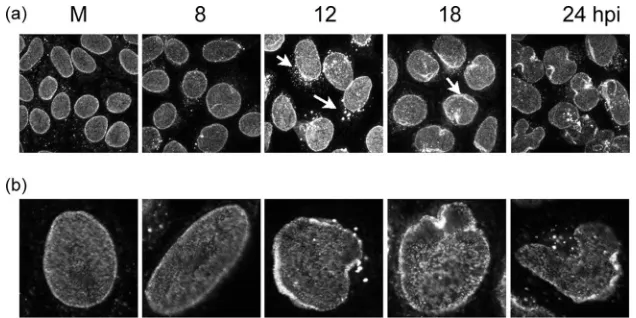
sive effect of HSV infection on NPCs by immunofluorescence and Western blotting. Cells infected by HSV-1 were fixed at different times after infection and probed with MAb414 (Fig. 1). In uninfected cells we observed a dense nuclear pattern enriched around and lining the whole nuclear envelope (Fig. 1, M). During infection, while the nucleus alters in shape and size, we observed a modest but distinct change in the pattern of staining of MAb414, with what appeared to be microclusters around the nuclear rim between 12 and 18 h. We also observed in many cells a minor but distinct speckled cytoplasmic stain-ing, although this was also occasionally observed in uninfected cells. Curiously we also observed a slight increase in intensity of overall staining under identical conditions, although this was lost at very late times (Fig. 1, 12, 18 and 24 hpi). We do not know the reason for this effect, which could be due to epitope unmasking or to a real increase, but we did not observe any increase in the abundance of the nucleoporins by Western blotting (see below). Taking into account the increase of the nuclear volume and the deformation of the nuclear envelope induced by HSV infection, localization of nucleoporins by MAb414 showed no gross morphological change or loss at least until very late times during infection.
In parallel, we examined the profile of the major nucleopor-ins by Western blotting with MAb414. We also wanted to examine the O-glycosylation status of nucleoporins, since this is one of the major posttranslational modifications of the nucleoporins (6, 14, 28, 32, 44) and indeed antibody to N -acetylglucosamine (O-GlcNAc) detects certain nucleoporins that are not as readily detected by MAb414. The results dem-onstrate that, during infection, monitored in this case by the progressive accumulation in VP5, no major difference in abun-dance of the major nucleoporins was detected with MAb414 (Fig. 2, left panel). Interestingly, using the anti-O-GlcNAc antibody, an increase in modification could be detected from 12 to 18 hpi. Although the precise identity of all of the proteins detected by this antibody are not known, many have been shown to be nucleoporins (6, 14, 28, 44). Given the stable levels of the nucleoporins detected by MAb414, one explanation for this result could be increased modification by O glycosylation,
either generally or specifically of nucleoporins. Either way we certainly did not observe any obvious loss of these components.
Analysis of isolated nuclear envelopes.Although total levels of NPC components appear constant during the main course of infection, the composition and integrity of the NPC was ana-lyzed in more detail by sucrose gradient centrifugation. Nu-clear envelope preparations were made from cells 16 h after infection or mock infection as described in Materials and Methods and loaded onto a 5 to 70% sucrose gradient. After ultracentrifugation the gradients were fractionated and ana-lyzed by Western blotting. Nuclear envelope fractions were first analyzed for the presence and distribution of nucleoporins with the MAb414 (Fig. 3). Nucleoporins Nup358, Nup214, and Nup153 cosedimented in the gradient, with fractionations from
[image:3.585.133.451.67.226.2]FIG. 1. (a) Immunofluorescence of nuclear pore proteins at different time points after infection with HSV-1. Cells (HEp-2 cells) were infected with HSV-1 at an MOI of 10 or mock infected and then washed fixed and stained with the nucleoporin-specific antibody MAb414 at the times indicated. Arrows indicate alterations in the distribution of the staining pattern in infected cells as discussed in the text. (b) Higher magnifications of typical images of single cells are shown in row b.
FIG. 2. Analysis of major nucleoporins during HSV1 infection. Cells were infected with HSV-1 at an MOI of 10, and samples were taken at different time points as indicated. Aliquots of the extracts were denatured immediately in SDS sample buffer and separated on 4 to 8% gradient SDS-PAGE gels, transferred to polyvinylidene difluo-ride membranes, and stained with MAb414 to detect the nucleoporins Nup358, Nup214, and Nup153 proteins. Parallel samples were probed with O-GlcNAc specific antibody. In parallel, samples were probed with antibody to the major capsid protein VP5 to monitor virus infec-tion.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.311.521.488.630.2]uninfected and from infected cells resulting in very similar distribution patterns (Fig. 3a and b).
Peak fractions from uninfected and infected cells were also analyzed for the presence of other nuclear pore components, the membrane-spanning nucleoporins pom121, gp210, and the central channel protein Nup62 (Fig. 3c). No significant changes in the abundance or distribution of these components were observed (Fig. 3c and data not shown).
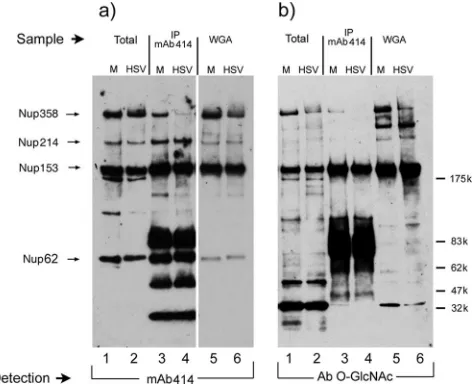
Immunoprecipitation of nucleoporins.Continuing to probe for potential differences in levels or association of nucleoporins during HSV infection, we analyzed the nucleoporins by the immunoprecipitation and subsequent Western blotting of the associated proteins. These analyses were performed with ei-ther antibody MAb414 or WGA-coupled agarose. WGA selec-tively enriches for proteins modified by N-acetylglucosamine and is frequently used as a probe for the O-GlcNAc-modified nucleoporins, as described above.
The results show parallel samples from mock-infected (Fig. 4, lanes M) and infected cells (Fig. 4, lanes HSV) as follows: total extracts (lanes 1 and 2), enriched MAb414-precipitated samples (lanes 3 and 4), and WGA-enriched samples, detected first by Western blotting with MAb414 (Fig. 4a). In the total samples, Nup358, Nup214, Nup153, and p62 were seen with little alteration as observed earlier (cf. lanes 1 and 2). In MAb414 immunoprecipitates, the ratios of the nucleoporins changed somewhat with, for example, Nup153 and p62 appear-ing relatively enriched (cf. lanes 1 and 3). Additional bands in the precipitate represented antibody species in the primary immunoprecipitates. The alteration in detection in the precip-itate versus the total sample likely represents differences in ratios of these components in different subcomplexes present in the extract and precipitated by the antibody. Nevertheless, for the nucleoporins detected, again there was no major change in uninfected versus infected extract (cf. lanes 3 and 4). We noted one possible exception to this in that Nup358, while present in the total infected cell extract, was selectively under-represented in the infected cell immunoprecipitates compared to the other nucleoporins (cf. Nup358 in lanes 3 and 4 versus Nup214 or Nup153 in lanes 3 and 4). This relative reduction in Nup358 immunoprecipitates was also observed when
[image:4.585.133.450.68.208.2]compo-nents were enriched with WGA, followed by probing for nucleoporins with MAb414 (Fig. 4, lanes 5 and 6). In addition, we probed the same samples, i.e., total extracts, MAb414 im-munoprecipitates, and WGA-enriched fractions with antibody to O-GlcNAc (Fig. 4b). In the total samples, not all bands are detected with the same efficiency as when probing with MAb414. However, the results indicate that while the total levels of Nup358 in uninfected and infected cells were similar (Fig. 4a, lanes 1 and 2), the levels of O-GlcNAc-modified
FIG. 3. Gradient fractionation of nucleoporins from isolated nuclear envelopes from mock-infected (M) (a) or HSV-infected (b) cells. Cells (HEp-2 cells) were infected at an MOI of 10, and at 16 hpi nuclear envelope preparations were made as described in Materials and Methods. The extracts were fractionated by sucrose density ultracentrifugation, fractions were harvested from top to bottom, and aliquots were analyzed by Western blotting with MAb414. Fractions numbers are indicated at the top of the Western blots. (c) The peak fraction 8 in each case, probed with MAb414 for the major nucleoporins or separately with different antibodies for the presence of additional nucleoporins as indicated.
FIG. 4. Immunoprecipitation of nucleoporins. The composition of the NPC was further compared between mock-infected (M; lanes 1, 3, and 5) and HSV-1-infected (HSV; lanes 2, 4, and 6) cells by immuno-precipitation (IP). Cells (HEp-2 cells) were infected as described in the text and harvested at 18 hpi. Aliquots of total cell lysates used for the immunoprecipitation are in lanes 1 and 2 (input). Nucleoporins were affinity purified either by antibody MAb414 (lanes 3 and 4) or by WGA-Sepharose in (lanes 5 and 6). Aliquots of the enriched fractions were analyzed by Western blotting. Nucleoporins were detected by antibody Ab414 (a) or by an antibody to O-GlcNAc modified proteins (b). The sizes of the molecular weight markers and the positions of the corresponding nucleoporins are indicated on the right and left sides, respectively.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.305.541.413.605.2]Nup358 were reduced in infected cells Fig. 4b, lanes 1 and 2). This relative reduction was again observed in the WGA-en-riched fraction, when probed with the O-GlcNAc antibody (Fig. 4b, lanes 5 and 6). One explanation consistent with these results is that while HSV does not induce major changes to these major pore constituents; nevertheless, changes in the levels of O-glycosylated Nup358 (Fig. 4b, lanes 1 and 2) and in the recruitment of a population of Nup358 into MAb414-precipitable complexes (Fig. 4a, lanes 3 and 4) may occur. Overall, these results indicate that HSV infection does not result in major changes in the composition of these major components of NPCs nor in their levels or association. How-ever, specific modifications to selected proteins of the NPC, exemplified by underglycosylation of Nup358, may occur, and this could have bearing on NPC function, gating, or selectivity.
Permeability barrier of the nuclear envelope during HSV-1 infection.The transport of large proteins and assemblies be-tween the cytoplasm and nucleoplasm is regulated by the gat-ing properties of the NPCs, which restricts molecules larger than 10 nm or 45 kDa from passive diffusion between the compartments. The nucleocytoplasmic transport of molecules beyond the limit for passive diffusion therefore requires an active transport mechanism, dependent upon specific signals and adaptors and catalyzed by specific import-export machin-ery. Many previous studies on the localization of herpesvirus components have included parallel examination of the steady-state location of certain host cell nuclear components. Pro-found reorganization of the nucleus and of bulk nuclear com-ponents such as host cell chromatin have been noted in previous studies (4, 7, 18, 25, 36, 42). However, while the appearance of nuclear proteins, such as INM proteins, within cytoplasmic membranes has been observed, major dislocation of nuclear proteins to the cytoplasm has not been reported. Nevertheless, given recent results from electron microscopy on major structural alteration to NPCs and the proposal for direct exit of capsids of 100-nm diameter through modified pores, we sought to examine the functional gating of NPCs in live in-fected cells.
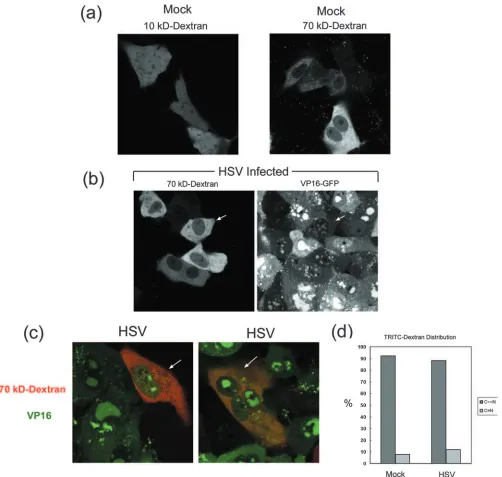
One route to examine the permeability of NPCs in vivo is by microinjection of inert fluorescently tagged dextran beads of defined size (21). Beads of defined size are injected into the cytoplasm and, if below the threshold for passive diffusion (e.g., 10 kDa), rapidly equilibrate in the nucleus and through-out the cell. Beads above the gating threshold (e.g., 70 kDa) remain excluded from the nucleoplasm. To establish the sys-tem, we first analyzed the distribution of TRITC-labeled 10-and 70-kDa beads after microinjection into the cytoplasm of uninfected Vero cells. Concentrations of beads and microin-jection procedures were designed based on previous work (21) and are detailed in Materials and Methods. After injection, cells were incubated and monitored for the distribution of the beads. Typical results showing the distribution of the different-sized beads are shown in Fig. 5a. All cells injected with the 10-kDa dextran beads showed homogeneous distribution throughout the cell. In contrast, the majority of the 70-kDa beads remained in the cytoplasm, although minor amounts were observed in the nucleus, and occasionally (ca. 8%) cells injected with the 70-kDa beads exhibited a diffuse distribution throughout the cell. This latter feature may be due to inadver-tent nuclear injection or pressure during injection, or possibly
division, but did not affect the main conclusions from the study (see below).
To pursue the effects of infection on gating we used an HSV-1 strain, HSV[V41], expressing GFP-VP16. To obviate any potential effect of microinjection on infection itself, our approach was to first infect cells and then, 6 h after infection, to randomly inject the infected cells with fluorescent TRITC-labeled 10- or 70-kDa beads. The localization of the injected fluorescent beads was then analyzed in parallel with the ex-pression and localization of GFP-VP16 at different time points. Yields of HSV[V41], as with general HSV replication, are observed by 10 hpi and are maximal by approximately 20 hpi (19). The bead distribution was observed over a corresponding time course. Initially, at least 50 microinjected cells were ex-amined for each bead size. As expected, the 10-kDa beads were distributed equally between the cytosol and nucleoplasm, and this distribution did not change during the time course from 8 to 24 hpi, showing no restriction, e.g., from diffusion within the nuclear viral replication centers (data not shown). The distribution of the 70-kDa beads in typical fields is shown in Fig. 5b and c. In Fig. 5b, each of the cells in the central patch contains cytoplasmic-restricted 70-kDa beads (left panel), and each of these cells exhibits pronounced VP16 expression typ-ified by VP16 recruitment into globular nuclear replication compartments (right panel). The distribution of the 70-kDa beads in these cells is indistinguishable from that seen in un-infected cells (Fig. 5a, right panel).
Additional fields of microinjected cells are shown in Fig. 5c, in this case showing the merged images of TRITC-labeled 70-kDa beads and VP16-GFP in the same fields. Again, in cells well in advanced stages with VP16 recruited into coalescing replication compartments and in a cytoplasmic vesicular pat-tern as described previously (19), TRITC-labeled 70-kDa beads remained largely excluded from the nucleus. A summary of evaluation of the distribution of the 70-kDa TRITC-labeled dextran beads in mock-infected versus infected cells expressing GFP-VP16 is shown in Fig. 5c. The distribution was scored as the ratio of cytoplasmic (C) to nuclear (N) fluorescence, with C⬎⬎N being typical of the selective partitioning of the 70-kDa beads, and C⫽N reflecting no selective partitioning (similar to the distribution of the 10-kDa beads). Although some ex-perimental variation was observed in the ratio between exper-iments and there was a modest increase in the C⫽N distri-bution in infected cells, the vast majority of the infected cells retained effective gating (at least for up to 18 hpi). We inter-pret these results to indicate that, at least at this level of resolution, the nuclear envelope permeability barrier remained predominantly intact for particles of 70 kDa (36 nm) or larger size throughout the time course of normal HSV infection.
DISCUSSION
Herpesvirus nucleocapsids, 100 nm in diameter, are too large to pass through normal nuclear pores, which have aque-ous channels incorporating a gating mechanism for selective transport of soluble proteins and nucleoprotein assemblies (re-viewed in reference 46). It has been generally accepted there-fore that in the initiation of infection, HSV capsids do not enter the nucleus intact but rather undergo some structural rearrangement that results in capsid breakdown and DNA (or
on November 8, 2019 by guest
http://jvi.asm.org/
nucleoprotein complex) entry to the nucleus. Similarly, it is generally accepted that the primary pathway of nuclear exit is via capsid attachment to the INM and subsequent budding into the luminal space, thereby acquiring a primary lipid envelope (reviewed in references 3, 9, 26, and 43). However, the
[image:6.585.44.546.66.543.2]mech-anism by which HSV exits the nucleus remains a matter of controversy (27, 49), and an alternative pathway of capsid exit from the nucleus to the cytoplasm has been proposed on the basis of high-resolution electron microscopy (22, 50). These analyses indicated that infection resulted in gross enlargement
FIG. 5. Analysis of the permeability of the nuclear envelope during HSV-1 infection. (a) Uninfected Vero cells were microinjected with 10- or 70-kDa TRITC-labeled dextran beads as indicated. After injection, live cells were analyzed by confocal microscopy for the distribution of the fluorescent beads. Typical results from numerous fields are illustrated. (b) Cells were infected with HSV[V41] expressing GFP-labeled VP16 and 6 h later were microinjected with the 70-kDa TRITC-labeled dextran beads. Monolayers were incubated at 37°C and monitored for the progression of infection and localization of VP16-GFP and in parallel in the same cells for the localization of the 70-kDa TRITC-labeled dextran beads at different time points after infection. A typical field showing the separate distribution of VP16-GFP and TRITC-labeled dextran at 16 hpi is shown. (c) As described for panel b, with two merged fields showing TRITC and VP16 in the same field. These fields were evaluated at 16 hpi and are typical of many fields analyzed. (d) Summary of evaluation of the distribution of the 70-kDa TRITC-labeled dextran beads in mock-infected versus infected cells expressing GFP-VP16. The distribution in infected cells was from a total of 83 cells evaluated at 14 and 18 hpi. Approximately 110 mock-infected cells were scored. The distribution was scored as C⬎⬎N, typical of the selective partitioning of the 70-kDa beads, or C⫽N when the distribution resembled the diffuse distribution of the 10-kDa beads. Overall, the vast majority of the infected cells retained effective gating.
on November 8, 2019 by guest
http://jvi.asm.org/
of nuclear pores to between 300 and 500 nm, and up to 1,000 nm, through which nuclear content including capsids were transported to the cytoplasm.
Several reports have now documented significant changes to nuclear structure, specifically the nuclear membrane and un-derlying lamina, after HSV infection (2, 37, 40–42). A recent report also indicated that HSV-1 infection induces the recruit-ment of certain kinases to the nuclear rim, resulting in the phosphorylation of lamina components (33). As an extension to these observations, it could be that significant modifications are made to nuclear pore structure and function, but currently there is little direct analysis.
The main aim of the present study was to examine gating in live HSV-infected cells, since it would be reasonably predicted that gross alteration of the sort described above, sufficient to allow transport of a macromolecular assembly such as capsids, would have a significant effect on nuclear transport and gating controls. Cumulative observations on the selective accumula-tion of regulatory factors, DNA replicaaccumula-tion components, etc., within the nucleus would not of themselves provide evidence that gating remained intact since there is the possibility, if not the likelihood, of selective retention of nuclear components or assemblies. We set out to perform an analysis that has not been previously reported upon, namely, the examination of gating in live infected cells. We used an approach similar to that used in the examination of the breakdown of the nuclear envelope in mitosis (21), microinjecting fluorescently tagged dextran beads of different sizes and identifying and monitoring infection us-ing a GFP-expressus-ing virus. We observed selective retention of the 70-kDa beads in the cytoplasm, (as opposed to free diffu-sion of 10-kDa beads) but could find no significant difference in the distribution of the 70-kDa beads during infection when late replication compartments were formed, monitoring up to 18 h after infection.
Several possibilities may be put forward in the attempt to reconcile these observations. For example, it could be that within the profoundly rearranged nuclear pores, normal gating was either retained or indeed the altered pores were impene-trable for normal soluble-phase transport. We consider this unlikely. What might be more plausible is that the pore dis-ruption that was reported is relatively rare and localized and does not register as a change in the total diffusional capacity of the nucleus, such as our assay examines. If this were the case, it would require a spatially restricted mechanism to induce pore alteration and likely means that any exit through such altered pores was also spatially restricted.
However, while there was little evidence for alteration in functional gating there was evidence for modification of some sort to the pore. In their original work, Leuzinger et al. (22) examined NPC using anti-Nup153 antibody and reported the increasing presence of Nup153 in the cytoplasm of infected cells, similar to that observed here with MAb414. Similar re-sults were also recently reported by Nagel et al. (31), who observed cytoplasmic staining, occasionally in foci, a finding consistent with our observations. These authors also reported the clustering of nucleoporins and the appearance of large gaps in the otherwise continuous rim pattern of staining. These gaps were not necessarily associated with the pore alteration, as observed by electron microscopy, and the authors also pointed out that in the majority of cells with cytoplasmic
cap-sids, the nuclear pore architecture as observed in their studies was unchanged (31). Although we did not observe major dis-ruption in biochemical sedimentation analysis, two modifica-tions merit further comment. We reproducibly observed an increase in the profile of high-molecular-weight O-glycosylated proteins, among which nucleoporins are known to be the major constituents. Although we do not know the identity of the major species involved, it could be that such modification plays a role in pore function in infection. For example, considering the efficiency once in the cytoplasm with which an infecting capsid must dock at the pore, it remains unclear how capsids after egress do not reattach to pores, engendering a type of futile cycle. The relative efficiency of docking of capsids to pores of infected cells has not been examined, but it is possible that pore alterations that did not affect gating could affect virus-pore interaction. The other alteration that we observed was in reduced amounts of Nup358 in immunoprecipitated complexes with other nucleporins. Although modest, this was a reproducible finding with two- to threefold reductions not ob-served for other nucleoporins. Nup358 is also known as RanBP2 and is a Sumo-specific E3 ligase involved in the proper targeting of other pore proteins such as RanGap1. Further investigation should help reveal specific alteration to Nup358/RanBP2 function and location and the possible in-volvement in such modifications to altered trafficking of pro-teins or complexes at the pore.
In summary, functional analysis in living cells demonstrated no significant difference in partitioning or nuclear pore gating in HSV-infected cells. The results are consistent with the pro-posal that egress in infected cells is via an envelopment-deen-velopment route and that disrupted pores are not a major route of capsid exit to the cytoplasm.
ACKNOWLEDGMENTS
This study was funded by Marie Curie Cancer Care and by a Well-come Trust International Fellowship to H.H.
We thank Ulrike Kutay, ETH Zurich, for the generous supply of pom121 and gb210 antibodies.
REFERENCES
1.Bednenko, J., G. Cingolani, and L. Gerace.2003. Nucleocytoplasmic trans-port: navigating the channel. Traffic4:127–135.
2.Bjerke, S. L., and R. J. Roller.2006. Roles for herpes simplex virus type 1 UL34 and US3 proteins in disrupting the nuclear lamina during herpes simplex virus type 1 egress. Virology347:261–276.
3.Browne, H., S. Bell, T. Minson, and D. W. Wilson.1996. An endoplasmic reticulum-retained herpes simplex virus glycoprotein H is absent from se-creted virions: evidence for reenvelopment during egress. J. Virol.70:4311– 4316.
4.Burch, A. D., and S. K. Weller. 2004. Nuclear sequestration of cellular chaperone and proteasomal machinery during herpes simplex virus type 1 infection. J. Virol.78:7175–7185.
5.Davis, L. I., and G. Blobel.1986. Identification and characterization of a nuclear pore complex protein. Cell45:699–709.
6.Davis, L. I., and G. Blobel.1987. Nuclear pore complex contains a family of glycoproteins that includes p62: glycosylation through a previously uniden-tified cellular pathway. Proc. Natl. Acad. Sci. USA84:7552–7556. 7.de Bruyn Kops, A., and D. M. Knipe.1994. Preexisting nuclear architecture
defines the intranuclear location of herpesvirus DNA replication structures. J. Virol.68:3512–3526.
8.Drummond, S., and T. Allen.2004. Structure, function, and assembly of the nuclear pore complex. Symp. Soc. Exp. Biol.2004:89–114.
9.Enquist, L. W., P. J. Husak, B. W. Banfield, and G. A. Smith.1998. Infection and spread of alphaherpesviruses in the nervous system. Adv. Virus Res.
51:237–347.
10.Fahrenkrog, B., D. Stoffler, and U. Aebi.2001. Nuclear pore complex archi-tecture and functional dynamics. Curr. Top. Microbiol. Immunol.259:95– 117.
on November 8, 2019 by guest
http://jvi.asm.org/
11.Frey, S., R. P. Richter, and D. Gorlich.2006. FG-rich repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties. Science314:815–817.
12.Fuchs, W., B. G. Klupp, H. Granzow, N. Osterrieder, and T. C. Mettenleiter.
2002. The interacting UL31 and UL34 gene products of pseudorabies virus are involved in egress from the host-cell nucleus and represent components of primary enveloped but not mature virions. J. Virol.76:364–378. 13.Georgatos, S. D., and G. Blobel, and B. Lamin.1987 constitutes an
inter-mediate filament attachment site at the nuclear envelope. J. Cell Biol.105:
117–125.
14.Hart, G. W.1997. Dynamic O-linked glycosylation of nuclear and cytoskel-etal proteins. Annu. Rev. Biochem.66:315–335.
15.Isgro, T. A., and K. Schulten.2007. Association of nuclear pore FG-repeat domains to NTF2 import and export complexes. J. Mol. Biol.366:330–345. 16.Klupp, B. G., H. Granzow, and T. C. Mettenleiter. 2001. Effect of the pseudorabies virus US3 protein on nuclear membrane localization of the UL34 protein and virus egress from the nucleus. J. Gen. Virol.82:2363–2371. 17.Klupp, B. G., H. Granzow, and T. C. Mettenleiter.2000. Primary envelop-ment of pseudorabies virus at the nuclear membrane requires the UL34 gene product. J. Virol.74:10063–10073.
18.Knipe, D. M., D. Senechek, S. A. Rice, and J. L. Smith.1987. Stages in the nuclear association of the herpes simplex virus transcriptional activator pro-tein ICP4. J. Virol.61:276–284.
19.La Boissiere, S., A. Izeta, S. Malcomber, and P. O’Hare.2004. Compart-mentalization of VP16 in cells infected with recombinant herpes simplex virus expressing VP16-green fluorescent protein fusion proteins. J. Virol.
78:8002–8014.
20.Leach, N., S. L. Bjerke, D. K. Christensen, J. M. Bouchard, F. Mou, R. Park, J. Baines, T. Haraguchi, and R. J. Roller.2007. Emerin is hyperphosphory-lated and redistributed in herpes simplex virus type 1-infected cells in a manner dependent on both UL34 and US3. J. Virol.81:10792–10803. 21.Lenart, P., and J. Ellenberg.2006. Monitoring the permeability of the
nu-clear envelope during the cell cycle. Methods38:17–24.
22.Leuzinger, H., U. Ziegler, E. M. Schraner, C. Fraefel, D. L. Glauser, I. Heid, M. Ackermann, M. Mueller, and P. Wild. 2005. Herpes simplex virus 1 envelopment follows two diverse pathways. J. Virol.79:13047–13059. 23.Lim, R. Y., B. Fahrenkrog, J. Koser, K. Schwarz-Herion, J. Deng, and U.
Aebi.2007. Nanomechanical basis of selective gating by the nuclear pore complex. Science318:640–643.
24.Makatsori, D., N. Kourmouli, H. Polioudaki, L. D. Shultz, K. McLean, P. A. Theodoropoulos, B. M. Singh, and S. D. Georgatos.2004. The inner nuclear membrane protein lamin B receptor forms distinct microdomains and links epigenetically marked chromatin to the nuclear envelope. J. Biol. Chem.
279:25567–25573.
25.Maul, G. G., and R. D. Everett.1994. The nuclear location of PML, a cellular member of the C3HC4 zinc-binding domain protein family, is rearranged during herpes simplex virus infection by the C3HC4 viral protein ICP0. J. Gen. Virol.75:1223–1233.
26.Mettenleiter, T.2002. Herpesvirus assembly and egress. J. Virol.76:1537– 1547.
27.Mettenleiter, T. C., and T. Minson.2006. The egress of herpesviruses from cells: the unanswered questions. J. Virol.80:6718–6719.
28.Miller, M. W., M. R. Caracciolo, W. K. Berlin, and J. A. Hanover.1999. Phosphorylation and glycosylation of nucleoporins. Arch. Biochem. Biophys.
367:51–60.
29.Morris, J. B., H. Hofemeister, and P. O’Hare.2007. Herpes simplex virus infection induces phosphorylation and delocalization of emerin, a key inner nuclear membrane protein. J. Virol.81:4429–4437.
30.Muranyi, W., J. Haas, M. Wagner, G. Krohne, and U. H. Koszinowski.2002. Cytomegalovirus recruitment of cellular kinases to dissolve the nuclear lam-ina. Science297:854–857.
31.Nagel, C. H., K. Dohner, M. Fathollahy, T. Strive, E. M. Borst, M. Messerle,
and B. Sodeik.2008. Nuclear egress and envelopment of herpes simplex virus capsids analyzed with dual-color fluorescence HSV1(17⫹). J. Virol.82:3109– 3124.
32.Pante, N., and U. Aebi.1994. Toward the molecular details of the nuclear pore complex. J. Struct. Biol.113:179–189.
33.Park, R., and J. D. Baines.2006. Herpes simplex virus type 1 infection induces activation and recruitment of protein kinase C to the nuclear mem-brane and increased phosphorylation of lamin B. J. Virol.80:494–504. 34.Patel, S. S., B. J. Belmont, J. M. Sante, and M. F. Rexach.2007. Natively
unfolded nucleoporins gate protein diffusion across the nuclear pore com-plex. Cell129:83–96.
35.Rabut, G., P. Lenart, and J. Ellenberg.2004. Dynamics of nuclear pore complex organization through the cell cycle. Curr. Opin. Cell Biol.16:314– 321.
36.Randall, R. E., and N. Dinwoodie.1986. Intranuclear localization of herpes simplex virus immediate-early and delayed-early proteins: evidence that ICP 4 is associated with progeny virus DNA. J. Gen. Virol.67:2163–2177. 37.Reynolds, A. E., L. Liang, and J. D. Baines.2004. Conformational changes in
the nuclear lamina induced by herpes simplex virus type 1 require genes UL31 and UL34. J. Virol.78:5564–5575.
38.Reynolds, A. E., B. J. Ryckman, J. D. Baines, Y. Zhou, L. Liang, and R. J. Roller.2001. U(L)31 and U(L)34 proteins of herpes simplex virus type 1 form a complex that accumulates at the nuclear rim and is required for envelopment of nucleocapsids. J. Virol.75:8803–8817.
39.Reynolds, A. E., E. G. Wills, R. J. Roller, B. J. Ryckman, and J. D. Baines.
2002. Ultrastructural localization of the herpes simplex virus type 1 UL31, UL34, and US3 proteins suggests specific roles in primary envelopment and egress of nucleocapsids. J. Virol.76:8939–8952.
40.Scott, E. S., and P. O’Hare.2001. Fate of the inner nuclear membrane protein lamin B receptor and nuclear lamins in herpes simplex virus type 1 infection. J. Virol.75:8818–8830.
41.Simpson-Holley, M., J. Baines, R. Roller, and D. M. Knipe.2004. Herpes simplex virus 1 UL31 and UL34 gene products promote the late maturation of viral replication compartments to the nuclear periphery. J. Virol.78:5591– 5600.
42.Simpson-Holley, M., R. C. Colgrove, G. Nalepa, J. W. Harper, and D. M. Knipe.2005. Identification and functional evaluation of cellular and viral factors involved in the alteration of nuclear architecture during herpes sim-plex virus 1 infection. J. Virol.79:12840–12851.
43.Skepper, J. N., A. Whiteley, H. Browne, and A. Minson.2001. Herpes simplex virus nucleocapsids mature to progeny virions by an envelopment3 deen-velopment3reenvelopment pathway. J. Virol.75:5697–5702.
44.Snow, C. M., A. Senior, and L. Gerace.1987. Monoclonal antibodies identify a group of nuclear pore complex glycoproteins. J. Cell Biol.104:1143–1156. 45.Terry, L. J., and S. R. Wente.2007. Nuclear mRNA export requires specific FG nucleoporins for translocation through the nuclear pore complex. J. Cell Biol.178:1121–1132.
46.Tran, E. J., and S. R. Wente.2006. Dynamic nuclear pore complexes: life on the edge. Cell125:1041–1053.
47.Vasu, S. K., and D. J. Forbes.2001. Nuclear pores and nuclear assembly. Curr. Opin. Cell Biol.13:363–375.
48.Weis, K.2007. The nuclear pore complex: oily spaghetti or gummy bear? Cell
130:405–407.
49.Wild, P.2006. The egress of herpesviruses from cells: the unanswered ques-tions. J. Virol.80:6717–6718.
50.Wild, P., M. Engels, C. Senn, K. Tobler, U. Ziegler, E. M. Schraner, E. Loepfe, M. Ackermann, M. Mueller, and P. Walther.2005. Impairment of nuclear pores in bovine herpesvirus 1-infected MDBK cells. J. Virol.79:
1071–1083.
51.Wozniak, R. W., and C. P. Lusk.2003. Nuclear pore complexes. Curr. Biol.
13:R169.