0022-538X/91/126749-12$02.00/0
Copyright © 1991, American Society for Microbiology
Regulation of
Herpes
Simplex
Virus
True Late
Gene Expression:
Sequences Downstream from the US11 TATA Box Inhibit
Expression from
an
Unreplicated Template
PHILIP K. KIBLER, JOANNE DUNCAN, BRUCE D. KEITH, THOMAS HUPEL,
ANDJAMES R. SMILEY*
Molecular Virology and ImmunologyProgram, PathologyDepartment,
McMaster
University,
Hamilton, Ontario,
Canada L8N3Z5
Received 11March 1991/Accepted 18 September 1991
Thetruelategenesof herpes simplex virus type1 (HSV-1)areexpressed only after theonsetof viral DNA replication. Previous studies demonstrated that latepromoters lack elementsupstreamof the TATA box and suggested that only a subset of TATA elements can function in the context of true late promoters. We determined which structural features oftruelatepromotersareresponsible for the stringent requirement for viral DNA replication by insertingaseries of simple modelconstructsinto the HSV-1genome in place ofone of the two promoters ofthe UL24gene.Anoligonucleotide consisting of 19 nucleotides spanning the TATA box of theHSV-1 true late US)) genedrove barely detectable levels of expression; bycontrast,the corresponding regions of the Adenovirus type2major latepromoterand the HSV-1 truelate glycoprotein Cpromoterwere much moreactive. Transcripts driven from all of these minimal TATA box promotersaccumulated without viral DNA replication. The activityof the US)) TATA boxwasstimulated by addingupstream Spl-binding sitesorplacing the US)) orrabbit
13-globin
cap/leader region (-11to+39) downstream. TheSpl-TATA and TATA-j8-globin cap/leader constructs remained replication independent, while the TATA-USII cap/leader promoter displayedtrue late regulation. These results demonstrate thatsequences located within the US)) cap/leader region imposeastrict requirement for viral DNA replicationon a minimal TATA boxpromoter.DNA replication is thought to play a central role in the
activation and repression ofgene expression during
devel-opment (7, 83), yet little is known about the mechanisms
involved.DNAviruses provide useful models for
investigat-ingthe link between transcription and replication, because
expression ofaspecificsubsetofviral genes (the late genes)
requires viral DNA replication. In adenovirus and herpes
simplexvirus type 1 (HSV-1), replication appears to induce a cis-acting modification of the template that renders it
permissive for late gene expression (14, 49, 76). To gain
further insight into the mechanisms that control late gene
expression, we examined the cis-acting requirements for regulated late promoter activity during HSV-1 infection.
HSV-1 is a large enveloped virus with a 152-kb
double-stranded DNA genome
bearing
atleast 70 genes(51).Shortlyafterinfection, theviralgenomeis delivered to the nucleus,
where it is transcribed by the host RNA polymerase II. HSV-1genes fallinto atleastfour distinct temporal classes
that are regulated in a cascade fashion (35; reviewed in
references 21 and 80). Fiveimmediate-early (IE or a) genes aretranscribed first, and thesearetheonly HSV genes that are expressed in the absence of de novo viral protein synthesis. The IE proteins, in turn, activate the early (E or
,B)geneswhichencode the viral DNAreplicative machinery.
Theleaky late (L or yl) genes are induced soon after the E
genes; they are first expressed
prior
to the onset of viralDNAreplication,andexpression increasesatlatertimes ina
replication-dependent fashion. Finally, the true L (or -y2)
genes are activated only after DNA replication has begun.
Each HSV gene bears itsown promoter-regulatory region,
and promoter transplant and nuclear run-on transcription
*Correspondingauthor.
experimentsindicate thattemporal regulationoccurslargely atthe transcriptionallevel(27, 59, 66, 82).
Coordinate activation of HSV IE gene expression is
mediated through a class-specific consensus sequence, TA
ATGARAT, thatis found in one or morecopies ineach of
the IEcontrolregions (13, 25, 46, 47, 60, 78). IE
transcrip-tionis activatedbyanabundant structural componentofthe
virion, Vmw65 (also known as VP16, oa-TIF, and ICP25),
thatis delivered into cells upon infection(2, 10, 59).Vmw65
formsacomplex with cellularfactors, including OCT-1,and
theresultingactivatorcomplex bindstothe TAATGARAT
sequence, in part through the DNA-binding domain of
OCT-1(3, 26, 43, 44,52, 56, 61). Fourof thefiveIEproteins (ICPO, ICP4, ICP22, and ICP27), in turn, contribute to
activation oftheviral E and L genes(reviewedin reference
21).
The mechanisms of activation of E and L promoters
remain unclear.EandLpromotersdonotcontainobviously
conserved class-specific sequences analogous to TAATGA
RAT,andextensive mutationalanalyseshavenotuncovered
cis-actingsequencesthataresolely and specifically
required
for activation by IE
polypeptides.
Rather, the E and Lpromoter elements that are required for activation by IE
proteinsarethesame asthose needed for basal-levelactivity
in uninfected cells (12, 15, 17, 18, 33, 39, 65; reviewed in
references 21 and 54). These data have been
interpreted
tosuggest that IEproteinsinduceEandLgene
expression by
alteringtheactivity ofone or morecomponentsof thecore
transcriptionapparatus(21,54). Consistent with this hypoth-esis, the HSV IE proteins display relatively relaxed target
specificityandare abletoactivateavarietyof
heterologous
genes that have been newly introduced into cells
by
trans-fection or as part ofan infecting viral genome
(18-20, 22,
55-57, 73, 77). Some evidence suggests that TATA box
6749
on November 10, 2019 by guest
http://jvi.asm.org/
factor TFIID serves as a critical target of the IE proteins:
linker scanning mutations of the TATA box of the E
thymi-dine kinasegene(tk) result in a more severephenotype when
assayed under conditions ofIE trans induction than when
assayed under basal conditions (12), and changes in the
TATAbox sequence can markedlyaffect the responseofthe
simianvirus 40 early promoter to HSV IEpolypeptides (22).
Inaddition,certain constructs containing only aTATAbox
cap site region are efficiently induced by IE proteins in
transfection assays (65). Direct evidence for a functional
interaction between IE proteins and TFIID comes from
studies showing that the pseudorabies virus IE protein (a
homolog of HSV-1 ICP4) activates transcription invitro by
increasing the rate ofbinding of TFIID to the TATA
se-quence(1).
HSV true L promoters appear to haveasimpler structure
than E promoters, and some evidence suggests that this
difference playsanimportant regulatory roleduring infection
(33, 34, 39). The E promoters that have been examined
consist ofanear upstreamregioncomposedofbinding sites for cellular transcriptionfactors linked to a TATA
box/cap
site (18, 53, 54). In contrast, the promoters ofthe true L
US]], glycoproteinC (gC), and UL38 genes lack elements
upstreamof the TATAbox (24, 33, 34, 39). Inthe gCgene,
a 15-nucleotide (nt) segment spanning the TATA box was
reported to be sufficient for true L temporal regulation
during lytic virus infection (33). Johnson and Everett (39) first noted that these results suggest a correlation between
promoter complexity and temporal regulation during
infec-tion. Oneinterpretation of these observationsis that true L
regulation represents the default state of a minimal core
promoter. According to this view, the upstream promoter
elements found in E promoters allow expression from an
unreplicated template while DNAreplication is required to
activateaminimalpromoterconsisting ofonlyaTATAbox.
Consistent with this hypothesis, Johnson and Everett (39)
found that a transfected construct bearing the intact E
glycoproteinD(gD) promoter was expressedin the absence
of DNA replication, while a derivative lacking sequences
upstream of the TATA box was active only when the
template was allowed to replicate. Conversely, Homaetal.
(33) were able to relieve the requirementfor DNA
replica-tionforexpressionof a true L gene byaddingthe upstream
regionof the Etkpromoter tothe gC TATA box. However,
severalobservationssuggest that thesituationismore
com-plex and provide evidence that true L promoters contain
specialized cis-acting sequences that actively contribute to
true Lcontrol. (i) Imbalzano et al. (37) found thata mutant
tk genelackingallknown promoter elements upstreamofthe
TATA box remained under E control, implying that
up-streamelementsare notalwaysrequiredfor expression from
anunreplicatedtemplate. (ii)Homaetal. (33)foundthat the
tk TATA sequence was unable to function when it was
transplanted to the gC locus, leading to the hypothesis that
true L promoter activity requires a specific type of TATA
box sequence. (iii) Mavromara-Nazos and Roizman (50)
observed that sequences downstream of the TATA box of
thetrueL-y42gene canconfer some aspects of L regulation
on anE promoter.
We investigated the structural features that contribute to
temporalregulation of HSV-1 true L promoters bystudying
aseriesofsimple model promoters placed in the intact viral
genome. We confirmed that a short region spanning the gC
TATA box was highly active when removed from the
contextof the intactgC promoter, while thecorresponding region of the tk promoter was inert. However, the US]]
TATA boxwasbarely
active, indicating
that the functionaldifferences between TATA elements revealed
by
this type ofassaydonotstrictlycorrelate with thetemporalclass of the
promoters from which
they
are derived. In addition,tran-scripts driven fromall of the active minimal TATA promot-ersaccumulatedin the absenceof viral DNAreplication.We
found that sequences located downstream of the US]]
TATA boxcontributed topromoter activity and imposed a
strict requirement for viral DNA replication on minimal
TATA box promoters that were otherwise relatively
repli-cation independent. These data demonstrate that sequences
downstream ofthe US]] TATA box play an active role in
specifying
true Ltemporal
regulation.
MATERIALS ANDMETHODS
Viruses and cells. HSV-1 KOS PAAr5 (29) was used
throughoutthese studies. Virus stocks weregrown and their
titersweredeterminedonVerocells maintained inaminimal
essential medium (GIBCO) supplemented with5% fetal calf
serum.
RNAextraction. Verocells wereinfected at amultiplicity
of infection of 10 PFU per cell, and where indicated,
aphidicolin was added at 10,ug/ml. Cytoplasmic RNA from
infected cellswaspreparedbythemethod of Berk andSharp
(4).
Primer extension analysis. Primer extension analysis was
performed as previously described (73). The following
syn-thetic primers were purchased from the Central Facility of the Institute for Molecular Biology and Biotechnology,
McMaster University: (i) gD, 5'-CCCCATACCGGAACG
CACCACACAA-3'; (ii) HSV-1 UL24, 5'-CGAGACAATC
GCGAACATCTACACC-3'; and (iii) 7SL RNA, 5'-AACT
TAGTGCGGACACCCGATCGGC-3'.
Construction of model promoters. Simplemodel promoters
were assembled from synthetic oligonucleotides purchased
from the Central Facility of the Institute for Molecular
Biology and Biotechnology, McMaster University, byusing
standard methods (48). All of the promoter constructs were
inserted into the unique BamHI site of pTKSB (79) in the
UL24 orientation. pTKSB contains the HSV-1 tk gene
bearing a 200-nt deletion extending from +480 to +680; the
deletion is spanned by a BamHI linker.
Promoters consisting of the TATA boxes of the
adenovi-rus type 2 (Ad2) major late promoter (MLP) and the HSV-1
US]] and tk promoters were assembled from
double-stranded 25-mers containing 19 nts spanning the TATA box
flanked by linker sequences and 5' GATC protruding ends.
Theseduplexes regenerateaunique BamHI site downstream
of theinsert following insertion into pTKSB. The sequences
of the top strands of the oligonucleotides used were as
follows:
Ad2 MLP: 5'-gatctcgagctcGGGGGGCTATAAAAGGGG-3' HSV-1 US1: 5'-gatctcgagctcTGAGATCAATAAAAGGGG-3' HSV-1 tk: 5'-gatctcgagctcACTTCGCATATTAAGGTG-3'
Uppercase type indicates sequences derived from TATA
regions, and lowercase type indicates flanking linker DNA.
A promoter bearing the HSV-1 gC TATA box was
gener-ated as follows. A pTKSB derivative bearing the Ad2 MLP
TATA insert was digested with SstI and BamHI to release
the Ad2 sequence. Adouble-stranded 19-mer bearing the gC
sequence and terminating in SstI and BamHI cohesive ends
was then cloned in its place. Thesequence of the topstrands
of the oligonucleotides used to generate the gC TATA
sequence was5'-cGCCCGGGTATAAATTCCG-3'.
on November 10, 2019 by guest
http://jvi.asm.org/
ers bearing point mutations in the Ad2 and USJI TATA
sequences were constructed by the same strategy. The
sequencesof the upperstrands of the oligonucleotides used to generate thepoint mutations were as follows:
Ad2 MLP: 5'-cGGGGGGCAATAAAAGGGG-3'
USJJ: 5'-cTGAGATCTATAAAAGGGG-3' A promoter bearing two tandem Spl sites linked to the
USJJ TATA sequence was constructed by cleaving the
pTKSB derivative bearing the USJI TATA sequence with
SstI and BamHI andthen ligating adouble-stranded 47-mer
bearing the Spl-TATA construct and SstI and BamHI
pro-truding ends. The sequence of the upper strand of the
inserted oligonucleotide was 5'-gatctcgGGGGCGGGGCTG
GGGCGGGGCCTGAGATCAATAAAAGGGG-3'. A
pro-moterbearing mutated Spl sites linkedtothe USJJ TATA
box was constructed in the same manner, aside from the
changes in the Spl sequences noted (see Fig. 6).
Promoters bearing the USJJ cap/leader sequence
down-stream of the TATA box were constructed by cloning a
double-stranded 55-mer bearing the USJJ cap/leader and
BamHI cohesive ends into the BamHI site immediately
downstream of the TATA box. The sequence of the top
strand of the USII cap/leadersequenceoligonucleotidewas
5'gatcGAGGACCGGGAGGCGGCCAGAACCGCCGTG
CACGACCCGGAGCGTCCCCTG-3'. The promoter
bear-ing the rabbit,-globin cap/leader sequence wasconstructed
in the same manner. The sequence of the upper strands of
the oligonucleotides use to generate the rabbit ,-globin
cap/leader sequencewas 5'gatcAGGGCAGCTGCTGCTTA
CACTTGCTTTTGACACAACTGTGTTTACTTGCAAT
CCG-3'.
All of the constructs were verified by DNA sequence
analysis using thedideoxy-chain termination method (63).
Marker rescue. Viralrecombinants bearing inserts of the
promoterconstructs were produced by in vivo
recombina-tion following cotransfection of Vero cells with infectious
viral DNA and plasmids carrying the appropriate promoter
sequences (71, 72). The resulting tk-deficient viral mutants
wereisolated by plaque purification in thepresenceof 100 ,ug
of5-bromodeoxycytidine perml. Recombinants bearing the
desired insertions were identified by Southern blot
hybrid-ization(74).
RESULTS
Functional heterogeneity of TATA box elements during
HSV-1 infection.Previous work suggested that thepromoters
of the HSV-1 true L USJJ and gC genes lack required
upstream regions, leading to the hypothesis that a TATA
box/cap site regioncanserve as anefficientpromoterduring
thelateststagesof infection (33, 34, 39). Inaddition, the data
of Homaetal. (33) indicated thata 15-nt segment spanning
the TATAregion of the gCpromoterwassufficient fortrue
L regulation while the TATA box of the E tk gene was
unable to substitute. As afirst step in exploring structural
features thatcontribute to true L regulation during HSV-1
infection, wecomparedthe activities ofavariety of TATA
box elements all placed precisely in the same sequence
context within the HSV-1 genome. In this assay, we
re-placed thepromoterforatranscript of the UL24gene with
oligonucleotides bearingtestTATA sequences.
Twotranscripts of the UL24geneoverlap the tkgeneand
are transcribed from the opposite DNA strand (32, 51, 62,
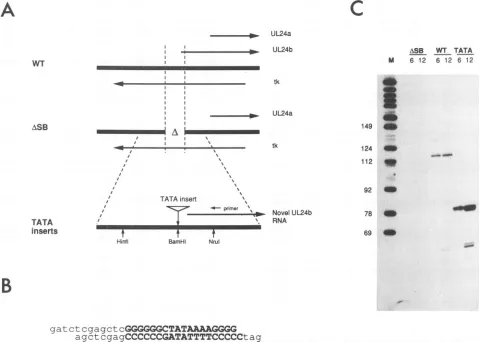
85). We took advantage ofapreviously described deletion of
tk coding sequences (ASB; reference 79) to link various
TATA box elements to UL24 transcribed sequences. The
ASB deletion extends from -172 to +24 relative to the initiationsite of thelarger UL24
transcript
(UL24b; Fig. 1A)
and isbridged by
aBamHI linker. As shown inFig. 1C,
aviralmutant bearingtheASB mutation failedtoaccumulate
detectable levels of UL24b
RNA,
indicating
that this dele-tioninactivatesthe UL24bpromoter. Weannealedcomple-mentary
pairs
ofoligonucleotides
toproduce duplexes
con-taining
various TATA box sequences(Fig. 1B).
Theseduplexes, bearing
5' GATCprotruding ends,
wereinsertedintothe BamHI sitethat spans the ASB deletion
endpoints,
in effect
replacing
the natural UL24bpromoterwith thetestTATA element. The
resulting
tk-deficientdeletion-substitu-tion mutadeletion-substitu-tions were thentransferred into theintact HSV-1
genome
by homologous
in vivo recombination.Following
selection of tk-deficientprogeny
by plaque
purification
in thepresence of 100 ,ug of
5-bromodeoxycytidine
perml,
virusisolates
bearing
the TATA insertswereidentifiedby
South-ern blot
analysis (74).
The activities ofthe various TATAboxes were then assessed
by
scoring
forproduction
ofUL24b RNAs
during
lytic
infection of Vero cells. On thebasis of the structures of the constructs,
transcripts
pro-moted
by
the inserted TATAboxeswerepredicted
toinitiateatca. +39relativeto the
wild-type
UL24b capsite,
withinsequences that
ordinarily
comprise
thenontranslatedleaderofthe UL24b
transcript.
Firstwe studiedthe
activity
ofanoligonucleotide bearing
19 residues
spanning
the consensus TATA box ofthe Ad2MLP. Verocellswereinfected with
wild-type HSV-1,
ASB,
and a recombinant virus
bearing
the MLP TATA boxin-sertedacrosstheASB
endpoints,
andcytoplasmic
RNAwasexamined
by
primer
extensionusing
aprimer
complemen-tarytoresidues +116to +92 of thenative UL24b
transcript.
As
expected, wild-type
transcripts
gave rise to a ca. 116-ntprimer
extensionproduct
while ASB gave nosignal
(Fig.
1C). In contrast, RNAobtained fromcells infectedwiththe
construct
bearing
the TATA insert gave rise to a novelextension
product
ofca. 78nts(Fig.
1C).Thesedataindicatethat the Ad2 MLP TATA box induced the formation ofa
novel UL24
transcript
thatinitiated atthepredicted
siteca.30 nts downstream of the first T of the TATA sequence.
Confirming
thisinterpretation, Si
nucleaseanalysis
using
a5'-labelled
single-stranded fragment extending
between the NruI andHinfl
sitesdiagrammed
inFig.
1A(labelled
atthe NruIsite)
gave rise to thepredicted
ca. 70-ntprotected
fragment
(data
notshown).
This noveltranscript
accumu-lated to
higher
levels thanwild-type
UL24b RNA andincreased in abundance between 6 and 12 h
postinfection
(Fig.
1C). Accumulation oftranscripts
wascompletely
pre-vented when
synthesis
of IEproteins
was blocked withcycloheximide
(data
notshown).
These data are consistentwith the
suggestion
ofHomaet al.(33)
that certain TATAboxes are able to serve as
relatively
efficient promotersduring
the latephase
of HSVinfection; however,
the resultsleave open the
possibility
that theactivity
ofthe insertedTATA sequence
depends
on elementsprovided by
theflanking
HSVsequences.
We next examined
analogous
viral strainsbearing
the TATA boxregions
of the HSV-1 true LUSJJ
andgC
promoters, and the HSV-1 E tk promoter embedded in
precisely
thesamesequencecontextasthe Ad2 MLP TATAbox
(Fig. 2).
Alloftheconstructsdisplayed
similarlevels ofgD mRNA
(Fig. 2B),
confirming
that themultiplicities
ofinfection used were
comparable.
In contrast,large
differ-ences were observed between the levels of UL24b RNA
(Fig. 2A):
thegC
TATA sequencewas almost as activeason November 10, 2019 by guest
http://jvi.asm.org/
A
UL24b
WT
ASB
tk
UL24a
tk
TATAinsert
- primer %. TATA
inserts
Hinft BamHl Nrul
-.,,-- Novel UL24b ' RNA
ASB WT TATA
M 6 12 6 12 6 12
149 _
124
-112 q.p
92 _
78 _
69 _
gatctcgagctcGGGGGGCTATAAAAGGGG agctcgagCCCCCCGATATTTTCCCCCtag
FIG. 1. (A) Experimental design. Oligonucleotides bearingtestTATAboxsequences wereinserted intothe HSV-1genomeacrossthe
ASBdeletion inthe UL24 orientation, and then infected cell RNAwasscored for thepresenceof the predicted novel UL24btranscript by primer extension and Si nuclease analysis. The transcriptionmapof the HSV-1 tk locus isdisplayed, andtheextentof the ASB deletion is
indicated. ASBremoves200ntsof tk codingsequences, aswellasthe UL24bpromoter,and is spanned byaBamHI linker.(B) Sequence
ofthe Ad2 MLPTATA boxinsert.Boldfacetype,TATAsequence;lowercasetype,flanking linker DNA. (C) Analysis of UL24b transcripts induced by the Ad2 MLP TATA box. A 20-,ug sample of cytoplasmic RNA harvested 6or12 hpostinfection with 10 PFUof the indicated
virus strainpercellwasanalyzed byprimer extension usingaprimer complementarytoresidues 116to92 of the native UL24b transcript (A). Thewild-type(WT) strainwasHSV-1 KOS PAAr5. Markers (M)were3'-labelled HpaII fragments of pBR322DNA. The numberstotheleft
aresizes in nucleotides.
the Ad2 MLP sequence, while the USJJ TATA box was
muchless active and the tk TATA boxwascompletelyinert
inthisassay.Each of the active TATA elementsgaveriseto
a transcript that initiated at the same position within the
flanking UL24 sequences, so that the resulting RNAs all
have the same primary nucleotide sequence. It therefore
seems likely that the large differences in UL24b transcript
abundance reflect relative promoter strength rather than
differences in RNAprocessing, transport,orstability. These
results confirmed previous suggestions that the gC and tk
TATA sequences differ markedly in the ability to drive
transcription when they are removed from the context of
theirintactpromoters(33). However, the low activity of the
USJJ TATAelement suggested that these functional
differ-encesdonotstrictly correlate with the temporal class of the
promotersfrom which the TATA elementswere derived.
TheintactHSV-1 US]] andgCgenes aresubjecttotrueL
regulation, and their expression is almostcompletely
depen-dent on viral DNA replication (30, 40). To determine
whether the US]J and gC TATA regions residing at the
UL24 locusdisplayedthesameregulatory propertiesastheir
parentalpromoters,weassayed UL24b RNA levelsat6and
12 h postinfection with and without a block of viral DNA
replication imposed by aphidicolin. Transcriptsdrivenfrom
the Ad2 MLP, USJJ, andgC TATA box regions
accumu-lated without viral DNA replication, althoughthe amounts
were somewhatreduced (Fig. 2A).Thisresponseis
charac-teristic ofleakyrather thantrueLregulation. One interpre-tation of this result isthat the USJJ andgCTATAregions fall under theinfluence oftemporalcontrolsignalspresent in
the flanking DNA sequences when they are placed at the
UL24 locus. Consistentwiththis hypothesis, the wild-type UL24btranscript is also subjecttoleakyL regulation (data
notshown). Anotherpossibilityis that theseconstructslack
true Lregulatory elements thatare present in theintactgC
and USJJ promoters. Data supportingthis latterpossibility for the USJJ promoterarepresentedbelow.
Structural basis forthe lowactivityof the USJJ TATAbox. We examined the Ad2 MLP and HSV-1 USJJ TATA
se-quences for features that might account for their very
B
UL24a
I
I
A
'k
I I
I
I I
I
I
I
I I
r---
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.65.547.84.426.2]A
IK
z a- CL nsCL~~~~~~~~~~~O C!LC
Mca s a m a +
M
92 am
78 __ _
69 W
47 26 100 26 7.4 4.7 6.5 3.8 49 17 65 16 - %max.
abs.
Ad2 T -> A:
Ad2 MLP:
US 11 TATA:
US11 A -> T:
GGGGGGCAATAAAAGGGG 4
GGGGGGCTATAAAAGGGG
* * ** *
TGAGATCAATAAAAGGGG TGAGATCTATAAAAGGGG
B
Ad2 Usll Ad2-A Usll-T
M 6 9 12 6 9 12 6 912 6 9 12
124 _
I12 4
92 m
78 _ _
B s
C. Ad2; MLP GGGGGGCTATAAAAGGG ESV1 USL1 TG.AGATCAATAAAAGGGG
HSV1 gO
22 86100 988.15.0 16 14 15 24 36 30 %mox abs.
[image:5.612.351.543.78.392.2]GCCCGGGTATAAATTCCG HSV1 tk ACTTCGC.ATATTAAGGTG
FIG. 2. Activities of various TATA elements. (A) Vero cells
wereinfected with recombinant viruses containing the Ad2 MLP,
USII, gC, or tk TATA sequence with and without 10 ju.g of
aphidocolin (aph)perml, and 10-,ug aliquots of cytoplasmic RNA
harvested at 6 and 12 h postinfection were scored for UL24
transcripts by primer extensionasdescribed in the legendtoFig. 1. Relativesignal intensitieswerequantified by microdensitometry and are indicated below the lanes (strongest signal, 100%). Molecular weight markers (lane M) were 3'-labelled HpaII fragments of
pBR322 DNA. Marker fragment sizes (in nucleotides)areshownon
the left. max. abs., maximum absorbance normalized to100% for the Ad2 MLP 12-h timepoint. (B) Levels of HSV-1 glycoproteinD
(gD) transcripts. Twenty-microgram aliquots of RNA from thesame
time points used for panel A were assayed for levels of gD
transcripts by primer extension. (C) Sequences of the Ad2 MLP, HSV-1 USIH, HSV-1 gC, and HSV-1 tk TATA inserts.
different activities. The oligonucleotides bearing the MLP and USJJ TATAregionsdiffer in sequence atfivepositions
(Fig. 3A): the first residueof theAT-rich TATAcore is aT
in the MLP sequence and an A in the USJI element; in
addition,four G-C base pairspresentin the MLP sequence
immediatelyupstreamoftheTATAboxareA-Tpairsatthe
corresponding positionsof the USJJ sequence. Wechanged
the first residue of eachTATA core to the sequence ofthe other(Fig. 3A) and assayedthe resulting constructs during lytic infection of Vero cells (Fig. 3B). A mutation that
changedthe first Tof the MLP TATAcoretothe Afound in
the USJJ sequence reduced its activity six- to sevenfold;
however, the resulting promoter was still approximately
threefoldmoreactive thanthe USJJ construct. Conversely,
changingthe first Aofthe USJJ TATAcoretoaTincreased
promoter strength sixfold but this construct was still less active than the Ad2sequence. These resultssuggestthat the
AT-richTATAcoreand5'-flankingresiduesboth contribute
to the differences observed between the Ad2 and USJJ
TATAregions.
C
_m
_ii
_gD
FIG. 3. Effects ofpoint mutations onthe activities ofthe Ad2
MLP andHSV-1USIITATAsequences. (A)Structures of mutant
TATA elements. Asterisks denote differences between the Ad2
MLP and HSV-1 USII TATAsequences. (B) Activities ofmutant
TATA elements. Cellswereinfected with viralstrainsbearingthe
indicated TATA insertsattheUL24 locus. RNAwasharvestedat6, 9, and 12 h postinfection, and 10-sg aliquotswereassayed by primer
extensionanalysisasdescribed in thelegendtoFig. 1.Thearrowto
the right indicates the location of the expected primer extension product. Relative signal intensities are shown below the lanes
(strongest signal, 100%). Molecularweight markers(lane M) were
3'-labelledHpaII fragments of pBR322 DNA. Marker fragment sizes (in nucleotides)areshowntothe left.max. abs., maximum absorb-ance normalized to 100% for the Ad2 MLP 12-h time point. (C) LevelsofgDmRNA.Ten-microgram aliquotsof the RNAsamples used forpanel BwerescoredforgD transcripts by primerextension.
Although we have not systematically defined the
struc-tural requirements for efficient TATA activity in the
se-quencesettingofourassay,the datapresentedinFig. 2and 3 are consistent with the hypothesis that aclear T-A-T-A motifflanked byGC-richsequences is required. In support of thisinterpretation, wefound that several otherconstructs
bearing these features were as active as the Ad2 MLP
sequence (datanot shown).
Effect of theUSIHcap/leader region.The resultspresented above indicated that the USJJ TATAregion extendingfrom -33 to -14 served as a relatively inefficientpromoterthat
was subjectto leakyLregulationwhen itwasplacedatthe
UL24locus in the intact viralgenome. Incontrast, Johnson and Everett(39)found thataDNAsegmentextendingfrom -31 to +40 relative to the USJJ cap sites drove efficient
replication-dependent expression in a transienttransfection
assay,inafashion thatcloselymimicked true Lregulationof A.
Ad2 MLP US1' gC
go
69 _
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.71.300.79.360.2]HSV genes. Together, theirresults andourdatasuggestthat
sequences downstream of the USJJ TATA box play an
important role in regulating USJJ expression. We tested this
possibility by adding sequences extending from -11 to +40
relativeto the USJJ capsitestoconstructsbearing the USJJ
and Ad2 MLP TATA boxes. A double-stranded 55-mer
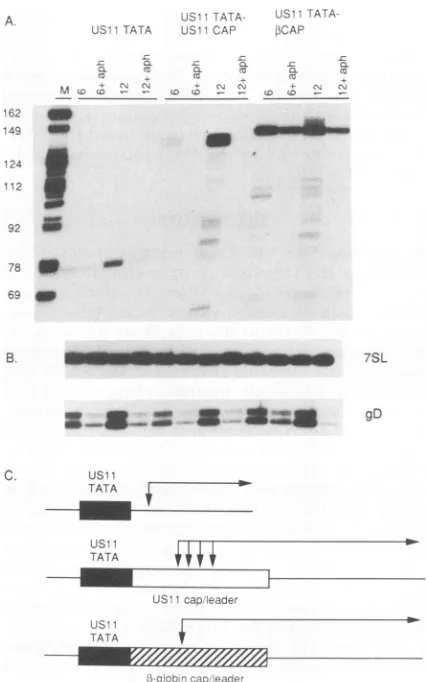
bearing the USJJ cap/leader region and BamHI cohesive ends was cloned into the BamHI site at the 3' end of the TATA box sequences, and the resulting constructs were transferred into the UL24 gene in the intact viralgenomeas described above. With the construct bearing the USJJ TATA box, this manipulation regenerated the correct se-quence of the HSV-1 USJJ promoter region from -33 to +40, asidefrom three changes at -14 to -12. Addition of the USJJ cap/leader region is predicted to increase the length of the UL24 transcript driven from the test TATA sequences by ca. 55 nts relative to that produced by con-structsbearing only aTATA region (Fig. 4B).
Vero cells were infected with viral strains bearing the Ad2 MLP and USJJ TATAboxes linkedto the USJJ cap/leader region, and RNAwas harvested at6and 12 h postinfection with and withoutaphidicolin blockadeof viral DNA synthe-sis (Fig. 4). Controlexperiments(Fig. 4B) demonstrated that the RNA samplesusedcontained equivalentamountsof7SL RNA, a cellular cytoplasmic RNA that does not change in abundance afterinfection (57a), and that eachconstructgave rise to similar levels of gD mRNA. Addition of the USIJ
cap/leaderregionto the USJJ TATA box had three effects.
(i) UL24b-related transcripts derived from the cap/leader promoter displayedthecharacteristic spread oftranscription start sites over a ca. 6-nt interval previously described for the native USJJ gene (38, 40; data not shown), whereas transcripts arising from the USJJ TATA box alone displayed a single start site. Further analysis revealed that transcripts driven from the USJJ cap/leader promoter initiated at pre-cisely the same sites as those arising from the native USJJ
promoter and that the largest primer extension product
correspondedtothe residuedesignated +3onthebasis ofSi
nuclease analysis (39; data not shown). (ii) The levels of
RNA arisingfrom the USJJ TATAboxcap/leaderpromoter
were boostedeightfold by 12h postinfection relative tothe levels of RNA arising from the construct bearing only the
USJJ TATA box. Lastly, in contrast to the promoter
con-sistingof only the USJJ TATA box, accumulation of UL24
RNA was prevented by blocking viral DNA replication. Thus, the construct bearing the USJJ cap/leader region was
regulated like the intact USJJ gene. Addition of the USJJ
cap/leader did not increase the levels of RNA arising from
the Ad2MLP TATAbox; however, in this case as well,the
resultingconstruct displayed increaseddependence on viral
DNA replication. These datasuggestthat sequenceslocated
downstream of theTATA box contribute to USJJ promoter activity and play a role in the temporal regulation of USJJ gene expression.
Effect of a heterologous cap/leader sequence. The
con-structs bearing isolated TATAelements linkedto the UL24
gene initiate transcription within sequences that do not
ordinarily serveasatranscription startsite. Incontrast, the
TATA-USJJ cap/leaderpromoters drive initiation from the normal USJJ cap sites. To determine whether theincreased activity and replication dependence of the TATA-USIJ cap/leaderconstructsresulted from provision ofabona fide
initiator element, we joined the USJJ TATA box to an
oligonucleotide bearing the corresponding portion of the
rabbit f3-globin gene (-16 to +40), such that transcription initiatedat theglobin RNAcapsite (Fig.SC).Addition of the
A. MLPlAlA
MLP TATA US.-CAP
CL < n
a u aL 0
u t1 CI 4
M c cD C2 N + N N
162 _
I49 _
a
124 _ 1-12
-92 _
78 _
69 _
US11TATA
US1 TATA US! 1 CAP aL a a Cl
S0 - CZ L
-U_tD CO
*E -uA.- b
B.
C usil
TATA
US11 capleader
Ad2MLP
TATA
JS1 capleader
FIG. 4. Effect of the USJI cap/leader region on the Ad2 MLP andUSHJTATAsequences.(A) Verocellswereinfectedat 10 PFU
percellwith virusescontaining eitherthe Ad2MLPorUSJI TATA sequencealoneorincombination withthe USJHcap/leaderregion.
RNAwas harvestedat6and 12hpostinfection, and 10-,ug aliquots
were assayed for UL24-related transcripts by primer extension as described in thelegendtoFig. 1.Duplicateinfections wereassayed
in thepresenceof 10
,pg
ofaphidicolinperml (lanes labelled +aph).Molecular weight markers (lane M) were 3'-labelled HpaII
frag-ments of pBR322 DNA. Markerfragment sizes (innucleotides) are shownontheleft. (B) Levelsof 7SLandglycoprotein D(gD)RNAs.
Aliquotsof the RNA samples usedfor panel Awere scored for7SL and gDtranscripts by primer extension. Theassaysfor 7SLandgD
RNAs used 0.5 and 10
,ug
ofRNA, respectively. (C) Structures of theTATA-USII
cap/leader promoters. The Ad2 MLP and USJITATAsequences were ligatedtoadouble-strandedoligonucleotide
bearing the US11 sequence extending from -11 to +39(see Mate-rials andMethods).
globin cap/leader region also increased the levels of RNA arising from the USJJ TATA box; however, the resulting
transcript accumulatedto high levels intheabsence of viral DNA replication (Fig. 5A). Thus, although a heterologous
initiator element increased the activity of the USJJ TATA box, only the USJJ sequence imposed a strict requirement forviral DNA replication. The slight differencebetweenthe sizes of the primer extension products arising from the globin and USJJ cap/leader promoters reflects the fact that thelongest USJJ extension product corresponds to residue +3 in thenumbering scheme of Johnson and Everett (39).
7SL
gD
do. woma i& -- -.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.315.547.73.432.2]US-
-ATA--S'' TATA usI CAP A
UjS'- TATA-,3CAP
H _
H < _
I<
H D- CL) < FL
_cn D Cn)
E *-- E
Mv L/) ( 0
c-m Cl) Z
-l)
0L C/
92
78
7SL
- a
69
z- -= a
a
doo 4
-m-
-C. usl1
TATA
B.
SP1 S!te GGGGCGGGGCT
Dp- SP s,e:GGTTCGGGGCT
-SA-A.A
[image:7.612.81.295.77.418.2]SP, SP, 7A-A F_ 1
FIG. 5. Effect of the rabbit ,-globin cap/leader region on the
USJI TATA box. (A) RNAwasharvestedfrom Vero cellsat6and 12 h postinfection with viruses containing the USHI TATA
se-quence,the US]I TATAsequencelinkedtothe USJI cap/leader,or
the USII TATA sequencelinkedtothe rabbit ,B-globin cap/leader insertedatthe UL24 locus. Duplicate infectionswereassayed in the
presence of 10 ,ug of aphidicolin per ml (lanes marked +aph).
UL24-relatedtranscript levelswereassayed by primerextension of
10 j±gof RNAasdescribed in the legendtoFig. 1. Molecular weight
markers (lane M) were 3'-labelled HpaII fragments of pBR322
DNA. Markerfragment sizes (innucleotides)areshownontheleft. (B) Levels of 7SL and glycoprotein D(gD) RNAs. Aliquots (0.5 and 10 ,ug) of the RNA samples used for panel Awere scoredfor 7SL andgD RNAs, respectively, by primer extension. (C) Structures of thepromotersusedfor panel A.
Effect of theUSIH cap-leaderregiononthe regulationofan
Spl-TATApromoter.Theresultspresentedinthepreceding
sectionssuggested that the USJJ cap/leader region imposed
an increased requirement for viral DNA replication on a minimal TATA box promoter. We tested whether the cap/ leader region was able to exert a similar effect on a more
complexpromoterinwhich the TATA boxwasactivatedby
an upstream promoterelement.
First we constructed a promoter bearing an artificial upstream region consisting of two tandem consensus
Spl-bindingsites linkedtothe USJJ TATA box. TheSpl-binding
sites hadthe samesequence asthe singlesitepresentinthe Ad2Elbpromoter andwerearrangedrelativetoeach other
toreproducethe spacingfound in the simian virus 40early
promoter. This upstream region was joined to the USJJ
FIG. 6. Effect of tandem Spl-binding sites on levels of RNA
driven fromtheUSI1 TATAsequence.(A)Vero cellswereinfected
at 10 PFU per cell with viruses containing the USJI TATA
se-quence, the USHI TATA sequence linked to two tandem Spl-binding sites, orthe USII TATA sequencelinked to two tandem mutated Spl-binding sites inserted at the UL24 locus. RNA was
harvested at 6 and 12 h postinfection, and 10-,ug samples were
assayed by primer extension analysisasdescribed in thelegendto
Fig. 1. Molecular weightmarkers(lane M) were3'-labelled HpaII fragmentsofpBR322DNA. Markerfragmentsizes(in nucleotides)
areshownonthe left.(B)Structures of the promoters used forpanel
A. Thesequencesof thewild-typeandmutantSpl-bindingsitesare
displayed.
TATA box such that the distance between the TATA se-quenceand theproximalSplsitewasthesame asin the Ad2
Elb promoter. As a control, we constructed another
pro-moterin which theSpl-bindingsiteswereinactivatedbytwo
pointmutations previously shown toeliminate Spl binding
(41; Fig. 6B). These two promoters were then fused to
UL24b transcribed sequences, transferred into the viral
genome, and examined during lytic infection of Vero cells. Addition ofSpl-binding sites greatly stimulated accumula-tion of RNA driven from the USJJ TATA box (averageof 17-foldoverthree experiments), whereas the mutated
Spl-bindingsites had no effect(Fig. 6A).
We thencompared the regulation of the Spl-TATA
pro-moterwith that ofaderivativebearingthe USJJ cap/leader A.
62 _ *,49_
241
112
92 IN
78 a
69 _
B.
gD
US,
TATA
tif
US''caD
eace-US, TATA
we
a Z::
ML zl: C- -a
7: ct
M co L-D S. -_, LO :.r-l 1-- 11 G --
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.372.528.87.437.2]A
co-infection (CD
oo
H-EL i
on or,
149
TATA-CAP -_
n-TATA-iii- a
TATA --w
1.24
112
92
78
69
B.
us,
P. SP! TATA 1
--- 1F'1 2us_
UlS ce:.DCi:.c-'
FIG. 7. Effect ofthe USI1 cap/leader regiononan Spl-TATA promoter. (A)Vero cells werecoinfected with viral strainsbearing Spl-TATA and Spl-TATA-USII cap/leaderpromoters inserted at
the UL24locus(5 PFUof each virus strainpercell).Samples (10 ,ug) ofRNA harvested at 3, 6, 9, and 12 h postinfection were then assayed for UL24-related transcripts by primer extension as de-scribed in thelegendtoFig.1.Whereindicated,10,ugofaphidicolin (aph)per ml wasincluded.The twolanesontheright containedcells singly infected witheach virus strain(MOI, 10PFU percell).The arrows on theleftindicate the expected sizes ofprimer extension products for promoters with and without the
USJJ
cap/leader region. Molecular weightmarkers (lane M)were3'-labelledHpaII fragmentsof pBR322 DNA. Markerfragment sizes(in nucleotides) are shownon the right. (B) Structures ofthe promoters used for panelA.region downstream (Fig. 7B). Vero cells were coinfected
with viral strains bearing the Spl-TATA and
Spl-TATA-USJJ cap/leader constructs, and RNA was harvested at 6
and 12 h postinfection with and without an aphidicolin
blockade of viral DNAreplication. The UL24-related
tran-scripts arising from the Spl-TATA and Spl-TATA-USII cap/leaderpromoters gave rise to distinct primer extension
products, allowing visualization of each transcript in mix-tures containing both. Addition of the
USJJ
cap/leader region to the Spl-TATA promoter resulted in increaseddependence on viral DNA replication: in the experiment depicted in Fig. 7, the levels of RNA arising from the
Spl-TATApromoter werereduced twofold by blockade of DNA replication while the RNA levels resulting from the
Spl-TATA-USII cap/leader
construct were inhibitedsix-fold.These data indicate that the USI Icap/leaderaltered the
regulation of the Spl-TATApromoter; however, the
result-ing construct was significantly less dependent on DNA
replication than was theTATA-USIJ cap/leaderpromoters
lackingupstreamSplsitesdescribedearlier. Thus,upstream
Spl sites partially alleviated the requirementforDNA
rep-licationthatisimposed bythe USJJ cap/leader regionon a
minimal TATApromoter.
DISCUSSION
We examined the structural features required for the
characteristic DNA replication dependence of the HSV-1
trueL US] Ipromoterby comparing the activitiesofaseries
of simple model promoters in the intact HSV-1 genome.
Previous work had led to the hypothesis that HSV true L
geneexpression requires viralDNAreplication becausetrue
Lpromoters lack elements upstreamof the TATA box(33,
39). Accordingtothisview,upstreamelements suchasthose
foundin Epromotersallowexpressionfromanunreplicated
template while DNA replication is necessary for activation
ofaminimal TATA box promoter. In addition, Homaetal.
(33)found that only asubset of TATA elements are able to
serveas trueLpromoters. On the basisofthesedata, itwas
proposed that true L promoters are characterized by a
specializedtype of TATA sequence that canfunction in the
absence of other promoter elements. Our results confirm
that certainpromoterslackingupstreamelementsaresubject
to trueLcontrolduring HSV-1 infection; however, wefound
that thisregulatory responserequires specialized sequences
located downstream of the TATAbox.
In agreement with earlier findings, ourdata indicate that
TATA elements differ markedly in the ability to drive
transcription when they are removed from the context of
their native promoters: the Ad2 MLP and gC TATA
se-quences werehighly active, the US]] sequencedrovebarely
detectable levels of expression, and the tk element was
completely inert. The results obtained with the gC and tk
TATA elements paralleled those obtained by Homa et al.
(33) in an entirely different sequence setting (the gClocus),
reinforcing the conclusion that these elements are
function-ally distinct. However, the low activity of the US]J TATA sequence demonstrated that the behavior of a TATA
ele-mentin thistype ofassaydoesnotstrictly correlate withthe
temporal class of the promoter from which it was derived.
Taken in combination, our data are consistent with the
hypothesis that efficient activity in the sequence setting of
our assay requires a clear T-A-T-A motif embedded in
GC-rich flanking sequences. Similar conclusions have
emerged from saturation mutagenesis of the gC TATA
element (62a). It is interesting that these features are
char-acteristic of the "consensus" TATA sequence derived by
comparing avariety of cellular and viral elements (6, 8).
The TATA box binds transcription factor TFIID as the
firststepintheassemblyof the RNA polymerase II initiation
complex (9, 23). Therefore, the heterogeneity of TATA
elements revealed by our assay presumably reflects
func-tionally distinct interactions with TFIID. Functional
differ-ences between TATA elements have been previously
ob-served in other systems (67, 68, 75, 81): forexample, onlya
subset ofTATAboxes can serve astargets for transactiva-tion by the adenovirus Ela protein (67, 68). Inasmuch as current evidence suggests that yeast and mammalian cells
haveasingle geneencoding TFIID (11, 16, 28, 31, 36, 42, 58,
64),the differences thatwe haveobserved can be explained
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.65.280.66.423.2]in two ways. The TATA elements may bind TFIID with
different affinities, directly leading to the observed
differ-encesin promoteractivity. Accordingtothishypothesis, the
Ad2 MLP TATA sequence would have the highest affinity
while the tk TATA box would have the lowest.
Alterna-tively, as previously suggested (81), TFIID may bind the
various TATA boxes with similar affinities but adopt
func-tionally distinctstates,dependingontheprecisesequenceof
the TATA element. Indeed, recent data demonstrate that
TFIIDundergoes aconformational change upon bindingto
the TATA element (45). Accordingtothis latterhypothesis,
only a subset of TATA boxes would result in a TFIID
conformation that is susceptible to transactivationby HSV
IEproteinsin the absence of other promoter elements.
We found that TATA elements were subject to leaky
rather thantrue Lregulation whenthey wereplaced atthe
UL24 locus. In
principle,
thisfinding
may either reflect aregulatory influence ofthe UL24 sequences that flank the
TATAinsert orindicate that theintactgC and USJJ genes
contain regulatory elements in addition to the TATA box
thatarerequiredfortrueLcontrol. While the firstpossibility
cannot be
entirely
discounted, our data strongly argue forthe second hypothesis in the case ofthe
USJJ
promoter.Thus, sequences spanning the
USJJ
transcription initiationsites andaportionof the nontranslated leader increased the
levelsof RNA driven from the
USJJ
TATAbox andimposeda
stringent
requirementfor viral DNAreplicationonminimalTATAbox promoters. The latterregulatoryeffectappeared
to be specific in that the corresponding region ofthe rabbit
,-globin
gene did not impose a replication requirement.These data confirm Johnson and Everett's conclusion (39)
that sequences extending from -31 to +40 relative to the
USJJ capsitesaresufficientfortrueLregulationand further
suggest that sequences downstream ofthe TATA boxplay
anactiveregulatoryrole. The simplest interpretation isthat
the
USJJ
downstream sequences block expression from aminimal TATA box promoter and DNA replication over-comes this
negative
effect. Several previous reports havesuggested
a role for sequencesdirectly
downstreamof theTATA sequenceinregulatinglevelsofHSV-1transcripts(5,
12, 24, 84).Inaddition,Mavromara-Nazos andRoizman (50)
found that the nontranslated leader of the true L-y42 gene
enhanced tk expression at late times postinfection when it
was placed downstream of the tk TATA box. While these
studies suggest that sequences downstream of the TATA box can
play
aregulatory
roleduring infection,
sequencesresponsible
forthereplication dependenceoftruelate genes have notbeenpreviously
identified.Because the
USJJ
downstream sequence renderedexpres-sion fromaminimal TATA promoter stringently dependent
onDNA
replication,
wetested theability
of this sequencetoimpose
a replication requirement on a more complexpro-moter in which the
USJJ
TATA box was activated byupstream Spl-bindingsites. Whilean
Spl-TATA-USJJ
cap/leader promoter was moredependentonviral DNA
replica-tion thanan
Spl-TATA
promoter,significant expression
wasobserved without viral DNA
replication.
Thus, upstreamSpl
sites were able to relieve the replication dependenceimposed by
theUSJJ
downstream sequencespartially.
These dataarein agreement withpreviouswork
demonstrat-ing
that upstream promoter elementscaninducereplication-independent
expressionfrom TATAbox/cap
regionsthatareotherwise
subject
to true L control(33, 39).
Inaddition,
these results illustrate howa range of
regulatory
responsescanbeachieved
by
various combinatorial arrangements ofalimited numberof
cis-acting
elements.The USJJ cap/leader region increased the levels of RNA
arisingfrom the USJJ TATA box. We consider itlikelythat
this effect results fromprovision of bonafide transcription
start sites at the
appropriate
distance from the TATA ele-ment. Consistentwiththisview,theanalogous regionof therabbit
P-globin
genehadasimilareffect;inaddition,prelim-inary data indicate that a linker-scanning mutation of the
US]] cap sites
strongly
inhibits promoter activity(unpub-lished data). ManyRNApolymeraseII promotersdisplay a
conserved sequence atthe
transcription
start site(6).
Thisinitiator elementplaysa role in promoter function(69) and,
in some cases, can support
TFIID-dependent transcription
in the absence ofa TATA box (70). The sequence of the
USIJ start sites bears little resemblance to the initiator
consensus,and USJJ
transcription
initiatesatmultiple
sitesspread over a ca. 6-nt interval (38, 40). However, the
characteristic
spread
of US]] initiation sites ispreserved
when the
USJJ cap/leader
region is linkedtothe Ad2 MLPTATAbox
(Fig.
4 anddatanotshown),
suggesting
that theUSII cap
region plays
a role in start site selection. Ittherefore seems
likely
that the USJJ promoter bears avariant initiator element. TheUSJI
cap/leader region
didnotincrease RNA levels when it was linked to the Ad2 MLP TATAbox.
Perhaps
thehighly
active Ad2 element doesnotbenefit from
linkage
tothe US]]initiator because this TATAcore sequence is sufficient to form a
productive complex
with TFIID.
How does the US]I
cap/leader region impose
anincreasedrequirement
for DNAreplication
on aminimal TATApro-moter? The
ability
of upstreamSpl-binding
sitestoalleviatethe effect
partially
argues that the blockimposed
by
theUSJJ
sequence isprobably
at the level oftranscription.
Assuming
that this is the case, two distinct classes ofmechanisms can be
proposed.
It ispossible
that the USJJ cap/leader region binds viral orcellularproteins that blocktranscription
initiationorelongation. According
tothisview,
replication
forkspassing through
theregion might dislodge
the repressor(s),relieving
theinhibitory
effect.Alterna-tively,
itispossible
that the sequencesatthe USJJtranscrip-tion start sites are
permissive
for initiation driven from aminimal TATA box
only
after conformationalchanges
areinduced byDNA
replication.
Thishypothesis
proposesthatthe US]]
cap/leader region
cannot interact with the basaltranscription
machinery
toformaproductive
initiationcom-plex
on anunreplicated template.
The available evidence doesnotdistinguish
between thesepossibilities,
and bothareconsistent with the observation that DNA
replication
isrequired
incis forexpression
oftrue L genes(49).
We aretrying
tomapmoreprecisely
theregulatory
sequencesin theUS]J
cap/leader region
asthefirst step towardsdistinguish-ing
between these alternatives.It will be of considerable interest to determine what
proportion
of HSV-1 true L promoters bearregulatory
sequences downstreamofthe TATAbox. Previous studies
suggestthat thetrue L
gC
gene doesnotcontainregulatory
sequencesinthe
cap/leader region (33), implying
thatnotalltrue L genes are controlled in the same way as USJJ.
Consistent with this
view,
thegC
and US]] promotersbehave very
differently
whenthey
are linked to reportergenesand
assayed
in transienttransfectionexperiments:
theUSJJ
promoterstringently
requires replication
in cis(38),
while the gC promoter drives efficient
expression
from anunreplicated template (65).
These observations raise thepossibility
that sequences located outside thegC
promoterregion
mayberequired
fortrueLregulation
ofthis gene. Theexperiments
presented
inthis report demonstrate thaton November 10, 2019 by guest
http://jvi.asm.org/
asegmentoftheUSJIJ promoterextendingfrom-11to +40
is sufficient toconfer true Lregulationon aminimalTATA
boxpromoter. Further study of the mechanismsinvolved is
likely to increase our understanding of the link between
DNAreplication and transcription.
ACKNOWLEDGMENTS
We thank Margaret Howes for technical assistance and F. L.
Graham for critical reviews of the manuscript.
Thisworkwassupported bya grantfrom theMedical Research Council of Canada. J.R.S. is aTerry Fox Senior Scientist of the
NationalCancer Institute of Canada, andT.H.wastherecipient of
astudentship from the National Science and Engineering Research
Council of Canada.
REFERENCES
1. Abmayr, S.M., J. L. Workman, and R. G. Roeder. 1988. The pseudorabies immediate early protein stimulates in vitro
tran-scription by facilitating TFIID:promoter interactions. Genes
Dev. 2:542-553.
2. Batterson, W., and B. Roizman. 1983. Characterizationof the herpes simplex virion-associated factor responsible for the induction ofagenes.J. Virol.46:371-377.
3. Baumruker, T., R. Strum, and W. Herr. 1988. OBP100 binds remarkably degenerateoctamermotifsthrough specific interac-tionswith flanking sequences. Genes Dev. 2:1400-1413.
4. Berk,A.J., and P.A.Sharp.1977. Sizing and mapping of early adenovirus mRNAs by gel electrophoresis ofS1 endonuclease digested hybrids. Cell 12:721-732.
5. Blair, E. D., C. C. Blair, and E. K. Wagner. 1987. Herpes simplex virus virion stimulatory protein mRNA leader contains
sequenceelements whichincrease both virus-induced
transcrip-tion and mRNA stability. J. Virol. 61:2499-2508.
6. Breathnach, R., and P. Chambon. 1981. Organization and expression of eukaryotic splitgenescoding for proteins. Annu.
Rev. Biochem. 50:349-383.
7. Brown,D. 1984. Theroleof stable complexes thatrepressand activate eukaryoticgenes. Cell 37:359-365.
8. Bucher, P., and E. N. Trifinov. 1986.Compilationandanalysis ofeukaryotic pol II promoter sequences. Nucleic Acids Res.
14:10009-10026.
9. Buratowski, S., S.Hahn, L.Guarente, and P.Sharp. 1989.Five
intermediate complexes in transcription initiation by RNA
poly-meraseII. Cell56:549-561.
10. Campbell, M. E. M., J. W. Palfreyman, and C. M. Preston.
1984. Identification of herpes simplex virus DNA sequences
which encodeatrans-acting polypeptide responsible for
stimu-lation of immediate early transcription. J. Mol. Biol. 180:1-19. 11. Cavallini, B., I.Faus, H. Matthes,J. M. Chipoulet, B.Winsor,
J. M.Egly, and P.Chambon.1989. Cloning of thegeneencoding the yeastprotein BTF1Y, which cansubstitute for the human TATAbox-binding factor. Proc. Natl. Acad. Sci. USA 86:9803-9807.
12. Coen, D. M., S. P. Weinheimer, and S. L. McKnight. 1986. A genetic approachtopromoterrecognitionduringtransinduction ofviralgeneexpression. Science 234:53-59.
13. Cordingley, M., M. Campbell, andC. M. Preston. 1983.
Func-tional analysis of a HSV-1 promoter: identification of
far-upstream regulatory sequences. Nucleic Acids Res.
11:2347-2365.
14. Crossland, L. D., and H. J. Raskas. 1983. Identification of adenovirus genes that require template replication for expres-sion. J. Virol. 46:737-748.
15. Eisenberg, S. P., D. M. Coen, and S. L. McKnight. 1985. Promoter domains required for expression of plasmid-borne copies of the herpes simplex virus thymidine kinase gene in
virus-infectedmousefibroblasts andmicroinjected frogoocytes.
Mol.Cell Biol. 5:1940-1947.
16. Eisenmann,D. M.,C.Dollard,and F.Winston.1989.SPT15, the
gene encoding the yeast TATA binding factor TFIID, is
re-quired for normal transcription initiation in vivo. Cell 58:1183-1191.
17. Everett, R. D. 1983. DNA sequences required for regulated expression of the HSV-1glycoproteinDgene lie within 83bp.of the RNAcap sites. Nucleic Acids Res. 11:6647-6666. 18. Everett, R. D. 1984. A detailed analysis of an HSV-1 early
promoter:sequencesinvolvedintrans-activationbyviral imme-diate-early gene productsare notearly gene specific. Nucleic Acids Res. 12:3037-3056.
19. Everett, R.D.1984.Transactivation oftranscriptionby
herpes-virus products: requirement for two HSV1 immediate-early polypeptidesfor maximumactivity.EMBOJ. 3:3135-3141. 20. Everett, R. D. 1985. Activation of cellular promoters during
herpes simplex virus infection of biochemically transformed cells. EMBO J. 4:1973-1980.
21. Everett,R. D.1987.Theregulationoftranscriptionof viral and cellulargenes by herpesvirus immediate-early gene products. AnticancerRes. 7:589-604.
22. Everett, R. D. 1988. Promoter sequence and cell type can
dramatically affect the efficiency of transcriptional activation inducedbyherpessimplexvirus type 1 and itsimmediate-early
geneproducts Vmwl75andVmwllO.J. Mol. Biol. 203:739-751. 23. Fire, A., M. Samuels, and P. A. Sharp. 1984. Interactions between RNApolymerase II, factors,andtemplateleadingto
accuratetranscription.J. Biol. Chem. 259:2509-2516. 24. Flanagan, W.M.,A.G.Papavassiliou,M.Rice,L. B.Hecht,S.
Silverstein, and E. K. Wagner. 1991. Analysis ofthe herpes simplex virus type 1 promoter controlling the expression of U138, a truelate gene involved incapsid assembly. J. Virol. 65:769-786.
25. Gaffney, D. F., J. McLauchlan, J. L. Whitton, and J. B.
Clements. 1985. A modularsystem for the assay oftranscription regulatory elements: the sequence TAATGARAT is required for herpes simplex immediate-early gene activation. Nucleic AcidsRes. 13:7847-7863.
26. Gerster, T., and R. G. Roeder. 1988. A herpesvirus
trans-activatingproteininteractswithtranscriptionfactorOTF-1 and othercellularproteins. Proc. Natl. Acad. Sci. USA 85:6347-6351.
27. Godowski,P.J.,and D. M.Knipe.1986.Transcriptionalcontrol
of herpesvirus gene expression: gene functions required for
positive andnegative regulation. Proc. Natl. Acad. Sci. USA
83:256-260.
28. Hahn, S.,S. Buratowski, P. A.Sharp, andL. Guarente. 1989. Isolation ofthegeneencodingthe yeast TATAbinding protein
TFIID:ageneidenticaltotheSPT15suppressorofTyelement insertions. Cell58:1173-1181.
29. Hall,J. D.,D. M.Coen, B.L.Fisher,M.Weisslitz,S.Randall,
R. E.Almy,P. T.Gelep,and P. A.Schaffer.1984.Generationof
geneticdiversity in herpes simplex virus: an antimutator
phe-notype mapstothe DNApolymeraselocus.Virology132:26-37. 30. Hall,L.M., K.G. Draper,R.J.Frink,R. H.Costa,and E.K. Wagner. 1982. HerpessimplexvirusmRNAspeciesmappingin EcoRIfragment1. J.Virol. 43:594-607.
31. Hoey,T., B. D. Dynlacht,M. G. Peterson, B. F. Pugh,andR. Tjian. 1990. Isolation and characterization of the Drosophila
gene encoding the TATA box binding protein TFIID. Cell
61:1179-1186.
32. Holland, L. E., R. M. Sandri-Goldin, A. L. Goldin, J. C. Glorioso, and M. Levine. 1984. Transcriptional and genetic
analysesoftheherpessimplexvirustype 1 genome:coordinates
0.29to0.45. J. Virol. 49:947-959.
33. Homa, F. L., J. C. Glorioso, and M. Levine. 1988. Aspecific
15-bpTATAboxpromoter element isrequiredforexpressionof
aherpes simplexvirus type 1 late gene. Genes Dev. 2:40-53. 34. Homa, F. L., T.M. Otal, J. C. Glorioso, and M. Levine. 1986.
Transcriptional control signalsofaherpes simplexvirustype 1
late (-y2) gene lie within bases -34 to +124relative to the 5' terminus of the mRNA. Mol. Cell. Biol. 6:3652-3666. 35. Honess,R.W.,andB.Roizman. 1974.Regulationofherpesvirus
macromolecularsynthesis. I. Cascaderegulationof the
synthe-sis ofthree groups ofviral proteins. J. Virol. 14:8-19. 36. Horikoshi, M.,C.K.Wang,H.Fujii,J.A.Cromlish, P. A.Weil,
and R. G. Roeder. 1989. Cloningand structure ofayeastgene
encoding ageneral transcription factorTFIIDthat bindstothe
on November 10, 2019 by guest
http://jvi.asm.org/
TATA box. Nature (London) 341:299-303.
37. Imbalzano, A. N., D. M. Coen, and N. A. DeLuca. 1991. Herpes simplex virus transactivator ICP4 operationally substitutes for the cellular transcription factor Spl for efficient expression of the viral thymidine kinase gene. J. Virol. 65:565-574.
38. Johnson, P. A., and R. D. Everett. 1986. DNA replication is required for abundant expression of aplasmid-borne lateUS11 gene of herpes simplex virus type 1. Nucleic Acids Res. 14:3609-3625.
39. Johnson, P. A., and R. D. Everett. 1986. The control of herpes simplex virus type 1 late gene expression: a TATAbox/cap-site region is sufficient for fully efficient regulated activity. Nucleic Acids Res. 14:8247-8264.
40. Johnson, P. A., C. MacLean, H. S. Marsden, R. G.Dalziel,and R. D. Everett. 1986. The product of gene US11 of herpes simplex virus type 1 is expressed as a true late gene. J. Gen. Virol. 67:871-883.
41. Jones, K. A., J. T. Kadonaga, P. A. Luciw, andR.Tjian. 1986. Activation of the AIDS retrovirus promoter by the cellular transcription factor, Spl. Science 232:755-759.
42. Kao, C. C., P. M. Lieberman, M. C. Schmidt, Q. Zhou, R.Pei, and A. J. Berk. 1990. Cloning of a transcriptionally active human TATA binding factor. Science 248:1646-1650.
43. Kristie, T. M., and B. Roizman. 1984. Separation ofsequences defining basal expression from those conferring a gene recogni-tion within the regulatory domains ofherpessimplexvirustype 1 a genes. Proc. Natl. Acad. Sci. USA 81:4065-4069. 44. Kristie, T. M., and B. Roizman.1987. Hostcellproteins bindto
the cis-acting site required for virion-mediated induction of herpes simplex 1a genes. Proc. Natl. Acad. Sci.USA84:71-75. 45. Lieberman, P. M., M. C. Schmidt,C. C. Kao, and A.J. Berk. 1991. Two distinct domains in the yeast transcription factorIID and evidence for a TATAbox-induced conformationalchange. Mol. Cell. Biol. 11:63-74.
46. Mackem, S., and B. Roizman. 1982. Structural features ofthe herpes simplex virus a gene 4, 0, and 27 promoter-regulatory regions which conferaxregulation on chimericthymidinekinase genes. J. Virol. 44:939-949.
47. Mackem, S., and B. Roizman. 1982. Differentiation between promoter and regulatory regions of herpessimplexvirus 1:the functional domains and sequence of a moveable a regulator. Proc. Natl. Acad. Sci. USA 79:4917-4921.
48. Maniatis, T., E. F. Fritsch,and J. Sambrook. 1982. Molecular cloning: a laboratorymanual.ColdSpring HarborLaboratory, Cold Spring Harbor,N.Y.
49. Mavromara-Nazos, P., and B. Roizman. 1987. Activation of herpes simplex virus 1 -y2 genes by viral DNA replication. Virology 161:593-598.
50. Mavromara-Nazos, P., and B. Roizman. 1989. Delineation of regulatory domains ofearly(1)andlate(y2)genes by construc-tion of chimeric genes expressed in herpes simplex virus 1 genomes. Proc. Natl.Acad. Sci. USA86:4071-4075.
51. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M.C.Frame,D.McNab, L. J. Perry, J. E. Scott, and P. Taylor. 1988. Thecomplete DNA sequence of the unique region in the genome of herpessimplexvirus type 1. J. Gen. Virol. 69:1531-1574.
52. McKnight, J. L.,T. M.Kristie, and B.Roizman. 1987. Binding of the virion protein mediating a gene induction in herpes simplex1-infectedcells to its cis site requires cellularproteins. Proc.Natl. Acad. Sci. USA84:7061-7065.
53. McKnight, S. L., and R. Kingsbury. 1982. Transcriptional control signals of a eukaryotic protein-coding gene. Science 217:316-324.
54. McKnight, S. L., and R. Tjian. 1986.Transcriptional selectivity of viral genes inmammaliancells. Cell46:795-805.
55. McKusker, C.T., and S. Bacchetti. 1988. Theresponsiveness of human papillomavirus upstream regulatory regions to herpes simpleximmediate-early proteins. Virus Res. 11:199-207. 56. O'Hare, P., and C. R. Gooding. 1988. Herpes simplex virus
regulatory elements and the immunoglobulin octamer domain bind acommon factor and are both targets for virion transacti-vation. Cell52:435-445.
57. Panning,B.,andJ. R.Smiley.1989.Regulationofcellular genes transducedbyherpessimplexvirus. J. Virol. 63:1929-1937. 57a.Panning,B.,andJ.R.Smiley. Unpublisheddata.
58. Peterson, M. G., N. Tanese, B. F. Pugh, and R. Tjian. 1990. Functional domains and upstream activation properties of cloned human TATAbindingprotein. Science248:1625-1630. 59. Post,L.E., S.Mackem, andB. Roizman.1981.Regulationofa
genes of herpes simplex virus: expression ofchimeric genes producedby fusion ofthymidinekinase withagenepromoters. Cell24:555-565.
60. Preston, C.M.,M.G.Cordingly,and N. D.Stow.1984.Analysis
of DNA sequenceswhichregulatethetranscriptionofaherpes simplex virus immediategene.J.Virol. 50:708-716.
61. Preston,C.M.,M. C.Frame,and M.E. M.Campbell. 1988. A complex formed between cellcomponents and an HSV
struc-turalpolypeptidebindsto aviral immediate-earlygene
regula-tory DNA sequence. Cell 52:425-434.
62. Read,G.S., J.A.Sharp,and W.C. Summers. 1984. In vitro and in vivotranscriptioninitiation sitesontheTK-encodingBamHI Qfragment of HSV-1 DNA. Virology 138:368-372.
62a.Roof,L.,and F.Homa. Personalcommunication.
63. Sanger, F.,S.Nicklen,andA. R.Coulson. 1977. DNA
sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
USA74:5463-5467.
64. Schmidt,M.C.,C. C.Kao,R.Pei,and A.J.Berk. 1989. Yeast TATA-box transcription factor gene. Proc. Natl. Acad. Sci. USA 86:7785-7789.
65. Shapira,M., F. L.Homa,J. C. Glorioso,andM.Levine. 1987.
Regulation ofthe herpes simplexvirus type 1 late (-y2)
glyco-protein C gene: sequences between base pairs -34 to +29 controltransientexpressionandresponsivenessto transactiva-tion by the products of immediate early (a) 4 and 0 genes. NucleicAcidsRes. 15:3097-3111.
66. Silver,S.,andB.Roizman. 1985. -y2-thymidinekinasechimeras
are identically transcribed butregulated as-y2genes inherpes
simplex virus genomes and as 1 genes in cell genomes. Mol. Cell. Biol. 5:518-528.
67. Simon,M.C.,T.M.Fisch,B.J.Bemecke,J.R.Nevins,andN.
Heintz. 1988.Definition ofmultiple,functionallydistinctTATA elements,oneof which isatarget inthehsp70promoter forElA
regulation. Cell52:723-729.
68. Simon,M. C., R.J.Rooney,T. M.Fisch, andN. Heintz. 1990.
Ela-dependent transactivation of the c-fos promoter
requires
the TATAA sequence. Proc.Natl. Acad. Sci. USA 87:513-517. 69. Smale, S. T., and D. Baltimore. 1989. The "initiator" as a
transcriptioncontrol element. Cell 57:103-113.
70. Smale,S.T.,M.C.Schmidt,A.J.Berk,andD.Baltimore.1990.
TranscriptionalactivationbySplasdirected
through
TATAorinitiator: specificrequirementformammalian
transcription
fac-torIID. Proc. Natl. Acad. Sci. USA87:4509-4513.71. Smiley, J. R. 1980. Construction in vitro and rescue of a
thymidine kinase-deficient deletionmutation ofherpes
simplex
virus. Nature(London) 285:333-335.
72. Smiley, J.R.,B. Fong,andW.-C.Leung. 1981. Construction of
adouble-jointed herpes simplex virus DNAmolecule: inverted
repeats are required for segment inversion and direct repeats
promote deletion. Virology 113:345-362.
73. Smiley, J.R.,C.Smibert,and R. D. Everett.1987.
Expression
of acellulargeneclonedinherpessimplexvirus: rabbitbeta-globin
is regulated as an early viral gene in infected fibroblasts. J.
Virol. 61:2368-2377.
74. Southern, E. M. 1975. Detection of
specific
sequences amongDNAfragmentsseparated by gel
electrophoresis.
J. Mol. Biol.98:503-517.
75. Struhl, K. 1986. Constitutive and inducible Saccharomyces
cerevisiaepromoters:evidence fortwodistinctmolecular mech-anisms. Mol. Cell. Biol. 6:3847-3853.
76. Thomas,G.P.,and M. B. Mathews.1980. DNA
replication
andtheearlytolate transition inadenovirus infection. Cell
22:523-533.
77. Tremblay, M. L., S.-P.Yee, R. H. Persson, S. Bacchetti, J.R. Smiley, andP. E. Branton. 1985. Activation and inhibition of
expressionofthe72,000-Daearly
protein
of adenovirustype5inon November 10, 2019 by guest
http://jvi.asm.org/
mouse cells constitutively expressinganimmediate early
pro-tein of herpes simplex virustype1. Virology144:35-45.
78. Triezenberg,S. J., S. L. LaMarco, and S. L. McKnight. 1988.
Evidence of DNA:protein interactions that mediate HSV-1 immediate early gene activation by VP16. Genes Dev.
2:730-742.
79. Varmuza, S.L., and J. R.Smiley. 1985.Signals for site-specific cleavage of HSV DNA: maturation involves two separate
cleavageevents atsites distaltotherecognitionsequences.Cell 41:793-802.
80. Wagner, E.K. 1985. Individual HSV transcripts,p.45-104. In
B.Roizman (ed.), The herpesviruses, vol. 3. Plenum Publishing Corp., New York.
81. Wefald, F. C., B. H. Devlin, and R. S. Williams. 1990. Func-tional heterogeneity of mammalian TATA-box sequences
re-vealed by interaction with a cell-specific enhancer. Nature (London) 344:260-262.
82. Weinheimer, S. P., and S. L. McKnight. 1987. Transcriptional andpost-transcriptionalcontrols establish the cascade ofherpes simplex virusprotein synthesis. J. Mol. Biol. 195:819-833. 83. Weintraub, H. 1985. Assembly and propagation of repressed
andderepressed chromosomalstates.Cell42:705-711. 84. Weir, J. P., and P. R. Narayanan. 1990. Expression of the
herpes simplex virus type 1 glycoprotein C gene requires
sequences in the 5' noncoding region of the gene. J. Virol.
64:445-449.
85. Wilkie, N.M., R. P.Elgin, P. G. Sanders, and J. B. Clements. 1980. Theassociation of herpes simplex virus with squamous carcinoma of the cervix, and studies of the virus thymidine kinasegene.Proc. R. Soc. London B Biol. Sci. 210:411-421.