0022-538X/89/031275-09$02.00/0
Copyright © 1989,American SocietyforMicrobiology
Functional Mapping
and
DNA
Sequence
of
anEquine Herpesvirus
1
Origin
of
Replication
RAYMOND P. BAUMANN, V. RAMANA R. YALAMANCHILI, AND DENNISJ. O'CALLAGHAN*
DepartmentofMicrobiology and Immunology, Louisiana State University MedicalCenter,
Shreveport,
Louisiana 71130-3932Received 9August 1988/Accepted6 December 1988
Thegenomeofequine herpesvirus1(EHV-1) defective interfering(DI) particle DNA originates fromdiscrete regions withinthe standard (STD)EHV-1genome: theleft terminus(0.0to0.04mapunits) and the inverted repeats (0.78to0.79 and 0.83to0.87mapunits ofthe internalinvertedrepeat;0.91to0.95 and 0.99to1.00
map units ofthe terminal inverted repeat). Since DI DNA must containcis-acting DNA sequences, suchas
replicationorigins, whichcannotbe supplied intransby theSTDEHV-1 virus, regions of the EHV-1genome
showntobe in DI DNA wereassayedfor thepresenceofa viral originofDNA replication. Specifically,STD
EHV-1 DNA fragments encompassing the genomic regionspresentin DI particle DNAwereinserted into the vector pAT153, and individual clones were tested by transfection assays for the ability to support the amplificationandreplication of plasmid DNA in EHV-1-infected cells.TheSma-1 subfragment ofthe internal inverted repeat sequence (0.83 to 0.85 map units) was shown to contain origin of replication activity. Subcloningand BAL 31 deletion analysis of the 2.35-kilobase-pair (kbp) Sma-1 fragment delineated a200-bp
fragmentthat contained origin activity. The origin activities of all EHV-1 cloneswhich werepositive by the
transfection assay were confirmed by methylation analysis by using the methylation-sensitive restriction enzymes DpnI and MboI. DNA sequencing ofthe 200-bp fragment which contained an EHV-1 origin of
replication indicatedthatthis region has significant homologytopreviouslycharacterized origins of replication of human herpesviruses. Furthermore, comparison of known origin sequences demonstrated that a 9-bp sequence,CGTTCGCAC, which isconservedamongalloriginsof replicationof human lytic herpesviruses and which is contained within the 18-bp region in herpes simplex virus type 1 origins shown by others to be protected byan origin-binding protein(P.Elias,M. E. O'Donnell,E. S. Mocarski, andI.R. Lehman, Proc.
Natl. Acad. Sci. USA 83:6322-6326)is also conserved acrossspeciesin the EHV-1 originofreplication.
Equine herpesvirus 1(EHV-1) isanatural infectiousagent ofhorsesandisanimportantcauseofspontaneousabortions in pregnant mares (35-37, 39). Serial undiluted passage of
standard (STD; infectious) EHV-1 results inthe generation of defectiveinterfering (DI) particles which displayallofthe properties which characterize classic DIparticles (19, 20, 37, 38). EHV-1 DIparticles havebeengenerated both intissue culture inL-M suspension cells (19) and invivo in suckling Syrianhamsters(6). Infectionofpermissivehamsterembryo cells withEHV-1 viralpreparationsenrichedfor DIparticles results in theestablishment of oncogenic transformationand persistent infection (10, 37, 45, 49). Cell lines established in thismannerrelease both infectious EHV-1 and DIparticles and have remainedpersistently infected afterover 10years
inculture (10;R. Hartyand D.J. O'Callaghan, unpublished observation). Ourlaboratory has continued to characterize the genetic structure and molecular organization of both STD and DIparticle DNA with the aim ofelucidating how DI particles participate in the establishment of persistent infection.
The EHV-1 genome is adouble-stranded DNAmolecule with a molecular size of 94 megadaltons, (MDa) which consists oftwo components: a long (L) region (73 MDa)
covalently linked to a short (S) region (21 MDa). The S regioncontainstwo identical invertedrepeats (IRs;8MDa) which bracketashortunique region (Us; 5 MDa)andwhich enable the entire Sregionto invert in orientation relative to the fixed Lregion (4, 21, 50, 64). Recentanalysisof EHV-1 DIparticle DNA has revealedthat DI DNAoriginatesfrom
* Correspondingauthor.
the leftterminus (0.0to0.04 mapunits)andtheIRS (0.78to 0.79 and0.83to0.87mapunits;0.91to0.95and 0.99to1.00
mapunits) of the STD EHV-1genome(1-3). Since DI DNA
must contain cis-acting regions (e.g., replication origins) which cannotbe supplied in transby the helper virus, it is probable that an EHV-1 originiscontained within the STD
genome leftterminus and/or within the IRsequences known
to bepresent in EHV-1 DI DNA.
The existence of replication origins in herpes simplex virus (HSV) DNAwasinitially suggested by electron micro-scopeexamination ofreplicatingviralgenomes(14, 22).The location of HSV origins was inferred fromearly studieson
the DNAcompositionofHSV DIparticles (forareview,see
reference 14). Subsequent studies demonstrated conclu-sively that DI DNA contains a replication origin located within the c component of the IRS (33, 54, 55, 62). Five distinct human herpesvirus lytic originsofreplication have
nowbeencharacterized andsequenced:twoin HSV-1
(oris,
[56, 57, 59]andoriL2[17, 63]), twoinHSV-2
(oriS2
[65]andoriL2[28]),andoneinvaricella-zostervirus(VZV;
oris
[58]). Inaddition, analysisof clonedpseudorabies virus DI DNA has suggested that two origins ofreplication arepresent at different locations in the unique long section of thepseu-dorabies virusgenome(66). Ingeneral the human
herpesvi-rus origins ofreplication share extensive homology, andall
containnearly perfect palindromeswhichcanbearrangedin hypothetical stem-and-loop structures. In addition, all five origins contain a conserved 11-base-pair (bp) motif (CGT TCGCACTT [58]) which has been shown by nuclease pro-tection studies oforis, to be bound by aprotein present in
HSV-1-infectedcells (13). Recentmutagenesis studies have 1275
on November 10, 2019 by guest
http://jvi.asm.org/
suggested
that the leftarmof theoris,
palindrome
containing
the conserved sequence is essential for
origin
activity
whereas the
right
arm can be deleted withoutsignificantly
inhibiting
replication
(11).
Thisstudy
also suggests that theability
toformacruciformstructureisnotessential fororis,
activity. Lastly,
transfection studies have indicated thatHSV-1
origin-mediated plasmid
amplification requires
only
seven viralgene
products
(9, 32, 67).
Inthispaper,wedescribethe
mapping
and DNAsequenc-ing
ofan EHV-1origin
ofreplication
within STD viral IR sequencespreviously
shown to bepresent
in EHV-1 DIparticle
DNA.The EHV-1origin
is showntopossesssignif-icant DNA
homology
acrossspecies
with all fivepreviously
sequenced
humanherpesvirus origins, including
a9-bp
motifnoted
by
others(58)
to behighly
conserved inherpesvirus
origins
andwhich hasrecently
been showntobeprotected
in the HSVoris, by
anorigin-binding protein
(13).
Unlike otherherpesvirus origins,
the EHV-1origin
does not contain anear
perfect palindrome
andis therefore limited in itsability
to form
hypothetical stem-and-loop
structures.MATERIALS ANDMETHODS
Cellsandviruses. EHV-1
(Kentucky
Astrain)
wasgrown in L-Msuspension
cell cultures asdescribed elsewhere(36,
39,
40).
Passage
ofEHV-1wasperformed
atlowmultiplicity
of infection
(0.005
PFU percell).
Virus waspurified by
polyethylene glycol
6000precipitation
ofinfected cell-freesupernatants, followed
by
severalcycles
ofrate-velocity
centrifugation
in Dextran 10gradients
(40).
L-M TK- cellswere obtained from the American
Type
Culture Collection(ATCC
CCL1.3)
and were maintained in the absence ofbromodeoxyuridine.
Isolationof viral DNA. Purified EHV-1 viral
particles
weresuspended
inTE
buffer(0.01
MTrishydrochloride,
pH
7.4,
0.001 M
EDTA)
and treated with sodiumdodecyl
sulfate(SDS;
finalconcentration,
1%,
wt/volandproteinase
K(100
,ug/ml)
for2 hat37°C
(19).
Following
incubation,
DNAwas extractedonce with80%phenol
inTEbuffer and twice withchloroform-isoamyl
alcohol(24:1,
vol/vol).
DNA waspre-cipitated by
the additionof 2.5 volumes of95% ethanol anda 1/20volume of4 M sodium acetate and was incubatedat
-70°C
for 1 h. DNA was collectedby centrifugation
at31,000
x gfor30 min andsuspended
in 0.1x TEbuffer.Plasmid construction. The
cloning
of the STD EHV-1 genome andofDI DNA hasbeen describedpreviously
(2-4,
46-48).
Allcloneswereinsertedinto thevectorpAT153
(60),
which lacks vector sequences known to
poison
the simianvirus 40
origin
ofreplication
(30),
or were insertedinto ourderivative, pAT12,
which contains themultiple cloning
siteof
pUC12
(61)
inserted between the EcoRI-HindIII sites ofpAT153
(pAT153
was agift
of RobertPerry). Cloning
of thedesired DNA
fragments
wasperformed
by
established pro-cedures(31).
When necessary,specific
DNAfragments
weresliced as individual bands from
low-melting-point
agarose(SeaPlaque;
FMCCorp.,
MarineColloids,
Div., Rockland,
Maine)
and isolatedby
extraction. Aftertransformation,
bacteria
(Escherichia
coliHB101)
were grown in thepres-enceofthe
appropriate
antibiotic,
andplasmid
DNAswereisolated from individual colonies
by
theminilysate
proce-dure
(23).
DNAswerescreened for insertsby
minisubmers-ible
gel
electrophoresis
through
0.8 to2.0%
agarose gels,following
restriction endonucleasedigestion.
Individualclones were
expanded
intolarge
bacterialgrowths,
and theplasmid
DNAwas isolatedby
thelysis-by-boiling
technique
(23)
and ethidiumbromide-CsCldibuoyant
density
gradientcentrifugation.
Clones pl-121 dll, d16, d17, and d18 were generatedby
BAL 31digestion of clone pl-121. pl-121was linearized
by
digestion with XbaI and treated for various times with 1.25 U of BAL 31(New EnglandBioLabs,Beverly,Mass.)at30°C. Phosphorylated XbaI linkers(New England BioLabs) were ligated to the appropriate BAL 31-digested samples (T4
DNA ligase; Bethesda Research Laboratories, Gaithers-burg, Md.), and samples weredigested with XbaI, ethanol precipitated, and religated overnight at 4°C. Individual clones were screened for inserts as described above, and specific insertswerefreed andligatedback into intact
pAT12
to ensure that all clones contained identical vector se-quences. Theextentof deleted sequenceswasestimated
by
gel electrophoresis of restriction enzyme digests: d16 (-200), d17 (-400), d18 (-600), and dll(-625).
Transfection assays. EHV-1 DNAfragments were tested for origin activity by using the transfection assay of Stow and McMonagle (59). L-M TK- cells were selected for transfectionanalysis becausetheyarepermissivefor EHV-1 replication, can readily be transfected, and will survive EHV-1 infection at ahigh multiplicity of infection (>5) for greaterthan 16 h(unpublished observation). Briefly, subcon-fluent monolayers of L-M cells in25-cm2 flaskswere trans-fected with equimolar amounts ofplasmid DNAs
(approxi-mately 200 ng) by the calcium phosphate technique (16).
DNAprecipitate(1 ml)wasadded to 10 ml of Eagle minimal essential medium EMEM (5% fetal bovine serum) in each 25-cm2 flask and incubated at37°C. Cells were shocked at 2 to 4 h posttransfection by the addition of 1 ml of 10% dimethyl sulfoxide-20% glucose for 5minat room tempera-ture(5), rinsed, and refed. At 6 h posttransfection, cellswere infected with EHV-1 at a multiplicity of infection of 5 or greater,andat20to24 hpostinfection, cells were harvested byscraping into 2 ml of 1xTE-0.6% SDScontaining 10to20 Fg/ml of proteinase K (Boerhinger MannheimBiochemicals,
Indianapolis, Ind.). Flasks were incubated overnightat37°C. DNAwasextracted twice withphenol saturated with 1 x TE buffer and once with chloroform-isoamyl alcohol (24:1, vol/vol). Following ethanol precipitation, DNA samples
wereresuspendedin 0.5 ml of distilledwaterandquantitated
spectrophotometricallyandvisually by ethidium bromidegel
electrophoresis.
Restriction enzymedigestion and Southern blot
hybridiza-tion. DNAs(10 to 15 ,ug or about 1/20 of the total DNAyield)
weredigestedtocompletion with a three- to fivefoldexcess of the desired restriction endonuclease under conditions recommendedby the supplier (Bethesda Research
Labora-tories). The resultant DNA digests were electrophoresed
through 0.8% agarose, stained with ethidium bromide (0.5 ,ug/ml), andphotographed in shortwave UV light (52). Gels
containing viral DNA fragments were alkali denatured, neutralized, transferredto hybridization membranes
(nitro-cellulose; Schleicher & Schuell, Keene, N.H.) in lOx SSC
(lx SSC is 0.15MNaClplus0.015 Msodium citrate) by the method of Southern (53), and immobilized by baking in vacuo at 80°C for 2 h. Filters were prehybridized in 6x SSC-0.5% SDS-5x Denhardt solution (lx Denhardt solu-tion is0.02% eachbovine serumalbumin,
polyvinylpyrroli-done, and Ficoll [Pharmacia Fine Chemicals, Piscataway,
N.J.])-200 jig
ofsalmon spermDNAper ml for 4 h at65°C. Probe DNA was 32P labeled by nick translation (43),dena-tured, andhybridized (2.0 x 106cpm/ml) with filters in the samemixture overnightat65°C.
Filterswererinsedoncewith 2x SSCatroomtemperature and then washed for 1 h in 2x SSC-0.5% SDS at 65°C,
followed by a 1-h wash in 0.lx SSC-0.1% SDS at 65°C.
on November 10, 2019 by guest
http://jvi.asm.org/
UL --~~~~~~~~~u - DI
~
UL L { 5 ; STD
S
~~~~~~~~2.35
kbp NSm-Hp Sp C H N B
115 117b
| I ~~~~~~118
- e 119 120 121 * 121-d16
1 121-d17
* 121-dl8
B-BamHI
C-CIaI
H-Hind III
Hp-HpaI
N-NruI
S-SmaI
Sp-SpeI
0.530
I
o*4
*635
121-dll 136 135 134
127 131 128 133
132
0.045
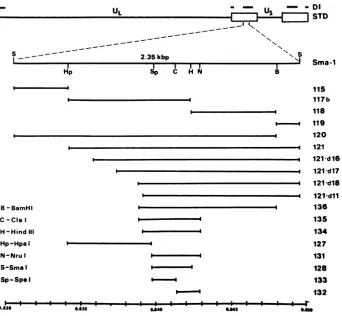
FIG. 1. Restrictionenzyme cleavage map and subcloning of the Sma-1 fragment. On the top lines are shown the DNA sequences present inEHV-1 DI DNAand the structure of the STD EHV-1 genome. IRs are shown as the boxed areas. The Sma-1 DNA fragment is shown in enlarged form below the STD EHV-1 genome. Subclones of the Sma-1 fragment are shown and are listed by number at the right. The constructionofspecific clones is described inMaterials and Methods. The abbreviationsfortherestriction endonucleases are given at bottom left. The bottom linegivesmapunits fortheSma-1 fragment.
Filterswereairdried and allowedtoexposeKodakX-Omat ARfilmat -70°C for 1 to 7 days in the presence of Cronex
Lightning-Plus (Du Pont Co., Wilmington, Del.).
intensify-ing screens.
DNAsequencing. The Sangerdideoxy technique was
em-ployed for DNA sequencing (51). A kit purchased from
American Bionetics (Hayward, Calif.) containing deaza dGTPwasroutinely usedforDNA sequencing. The 203-bp fragmentbetween the SpeIand ClaIrestriction sites (clone
pl-133) was sequencedinM13 mpl8 andmpl9 aspart ofa
generalprojecttosequence theentire 2.35-kbp Sma-1
frag-ment (Yalamanchili et al., manuscriptin preparation). The EHV-1 pl-133 DNA sequence was compared with the
pub-lished sequencesofthe known humanherpesvirusoriginsof
replication (28,34, 58, 63,65)by usingtheBIONETnational
computerresourceformolecular
biology
(26).RESULTS
Selection and cloning of STD EHV-1 DNA fragments. Previous studiesfrom thislaboratory demonstrated that DI
particle DNA originates from the left terminus (0.0to0.04 mapunits)and IRs(0.78 to0.79 and 0.83to0.87 map
units;
0.91to0.95 and 0.99to1.00 map
units)
of the EHV-1 STD genome.Analysis
ofDIparticle
DNA in otherherpesvirus
systems indicated that
cis-acting
sequences, such as cleav-ageencapsidationsignalsandreplication
origins,
arelocatedin DI DNA(15).Therefore,wechose fourSTD EHV-1 DNA clones for replication origin analysis which together con-tained virtually all of the DNA sequences known to be presentin EHV-1 DI DNA. These clonedwereBglIIN(0.00 to 0.03 map units), Sma-5 (0.78 to0.79 map units), Sma-1
(0.83to0.85 mapunits),and Sma-2(0.85to0.865 mapunits) (Table 1 and Fig. 1). As a precaution, each insert was recloned into the vector pAT153, which lacks DNA se-quences showntopoisonthereplication ofthesimian virus 40origin (60).
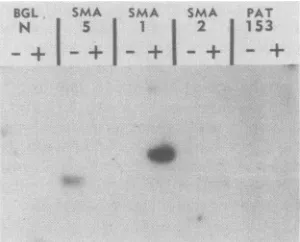
Individual EHV-1 clones in pAT153 were assayed for
originactivity bythemethod ofStow and
McMonagle
(59).Plasmid DNAs inequimolaramounts
(equivalent
to200 ngof pAT153)weretransfected intoduplicate flasks ofL-Mcells,and the transfected cells were either mock infected or infected(multiplicity of
infection,
5 orgreater)with EHV-1 STD virus.CellDNAswereextracted,
digested
withEcoRI,
and analyzed by Southern blot hybridization byusing 32p_
labeledpAT153DNA as a
probe (Fig.
2).EcoRIcleavesonly
once in each
plasmid
construct and therefore shouldyield
onlyasingle detectable band in infected-cell lanes in which
plasmid amplification has occurred in
linkage
to anEHV-1originof
replication.
Only
the Sma-1 clone showedamplifi-cation ofvectorsequencesin the lane with the infected cells
(Fig. 2).Thebandseenin theSma-5 mock-infected-celllane representsweak
homology
between theprobe
andcell DNAv
i w
---v
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.134.476.71.387.2]TABLE 1. Plasmidconstructions used fororigin analysis
Clone' ~ Size
MpuisOrigin
Clonea
(sbipz)(b
Map units activitycBglII N 2,700 0.000-0.030
-Sma-5 1,200 0.780-0.790
Sma-2 2,100 0.850-0.865
Sma-1 2,350 0.830-0.850 +
pl-115 450 0.830-0.834
-pl-117bd 1,000 0.834-0.843 +
pl-118 700 0.834-0.848
-pl-119 200 0.848-0.850
-pl-120 2,150 0.830-0.848 +
pl-121 1,900 0.834-0.850 +
d16e 1,700 0.836-0.850 +
d17 1,500 0.837-0.850 +
d18 1,300 0.839-0.850 +
dll 1,275 0.839-0.850 +
pl-127 700 0.834-0.840
-pl-128 300 0.840-0.842 +
pl-131 370 0.840-0.843 +
pl-132 167 0.841-0.843
-pl-133 203 0.840-0.841 +
pl-134 445 0.839-0.843 +
pl-135 470 0.839-0.843 +
pl-136 1,100 0.839-0.848 +
a All clones were inserted intothe vectorpAT153orits derivativepAT12
(see Materials and Methods).
bSizesweredetermined by DNAsequencingorestimatedbycomparison
toelectrophoretically separated restriction enzyme digestsofpBR322 and lambda bacteriophageDNA.
cOriginactivitywasdeterminedby transfection assay and and restriction enzymemethylation analysis (see Materials and Methods).
d Clone pl-117wasfoundtocontaintwocopies oftheinsert;asecond clone containing onlyasingleinsert(pl-117b)wasderived.
eClones pl-121d16, d17, d18,anddllwerederivedbyBAL 31deletion of clone pl-121.
sequences and was visible in all lanes upon prolonged
exposure but was not observed reproducibly in different Southernblots. Noamplification ofvector sequencesalone orofotherSTDEHV-1clones(Fig.2)wasdemonstrated. In
fact, overthe course of theseexperiments, amplificationof vectorsequences alone or EHV-1 DNA sequences outside ofSma-1 was never observed, even when conditions were
intentionally varied and excessive amounts of DNA were usedfortransfection. Thesestudiessuggestedthat an EHV-1 origin of replication exists within STD EHV-1 IR se-quences (Sma-1 fragment) known to be present in EHV-1 DNAparticleDNA.
Toconfirmthatthe Sma-1 plasmid is actually replicating in
EHV-1-infected cells, methylation analysis was conducted on cell DNApreparations. E. coli
Dam'
strains methylateadenine residues within their DNA; in contrast eucaryotic
cellsneithermethylate their DNAatadenine residues nor do
theypossess anadenine demethylase (27, 42). Thus,
restric-tionenzymes which aresensitive to the methylation state of the DNA can be used to differentiate between DNA repli-cated inbacteria and DNA which has replicated in eucary-otic cells. Two such enzymes are DpnI, which cleaves the sequenceGATC only when the A residue is methylated, and
MboI,
which cleaves at GATC only when the A residue is unmethylated. Plasmid DNA which has replicated in eucary-oticcells is DpnI resistant but MboI sensitive.DpnI-MboI
analysis ofamplified Sma-1 DNA (Fig. 3B)indicated that it was DpnI resistant (Fig.
3B,
lane ED) butMboI
sensitive(Fig. 3B, lane EM). When DpnI-sensitive plasmid DNA was
mixed withcell DNAbefore digestion (Fig. 3B, lane *ED), the exogenousplasmidwascleavedbyDpnI
(low-molecular-weight
smear). Hence,
lackofDpnI
cleavage
in lane ED isnotdueto
generalized
inhibition ofDpnI cleavage by
the cellDNA
sample.
Fine
mapping
oftheEHV-1origin.
The EHV-1oriS
wasdelineated further
by
subcloning
of the Sma-1fragment.
Aseriesof sixdifferent subclonesof Sma-1werederived in the vectors
pAT153
andpAT12
and includedpl-115,
pl-117b,
pl-118,
pl-119,
pl-120,
andpl-121 (Table
1 andFig. 1).
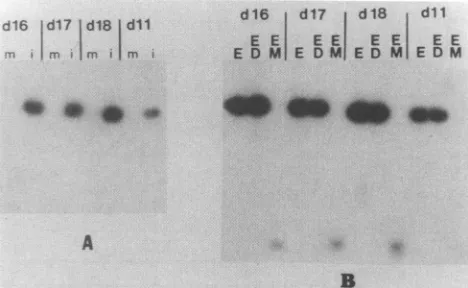
Transfection
analysis
of the six clonesindicated that clonespl-117b,
pl-120,
andpl-121
possessed origin activity
but clonespl-115,
pl-118,
andpl-119
wereorigin negative (Fig.
3A and
4A).
DpnI-MboI
analysis
oforigin-positive
clonesindicated that
amplified
sequences wereDpnI
resistantandMboI
sensitive,
a pattern indicative ofunmethylated
DNAwhich has
replicated
ineucaryotic
cells(Fig.
3B and4B).
Interestingly,
our initial isolate ofpl-117
contained twocopies
of theHpaI-HindIII
fragment
(Fig.
1)
anddemon-strated
only
minimalamplification
in theorigin
assay. Asingle-copy
clone(pl-117B
[lanes
with*])
which wassub-sequently
isolateddisplayed
readily
detectableorigin
activ-ity
(Fig.
3A).
Perhaps
tandemormultiply
origin
sequences indirect
linkage
areinhibitory
toorigin
function.The extentof
amplification
and theability
to detectinput
DNA varied among
transfections,
but theability
to bereplicated
wasalways
consistent for individual viral DNAclones. For
example,
inFig.
4A constructspl-120
andpl-121
underwent markedamplification
in infectedcells,
whereas the
amplification
seenfortheparental
Sma-1clonewas muchmore modest.
DpnI-MboI
analysis
indicatedthatplasmid
DNA in the mock-infected-cell lane(Fig.
4B,
pl-117B,
lanem)
wasunreplicated
(i.e.,
DpnI
sensitive andMboI
resistant),
eventhough
input pl-117B
DNA wasde-tected in the mock-infected-celllane
(Fig. 4A).
Incontrast,plasmid
DNA in the infectedsample (DpnI
resistant andMboI
sensitive)
hadreplicated
[Fig.
4B,
pl-117B
(i)].
Thesestudies indicate that EHV-1
replication origin
sequences were contained in the1-kbp
fragment
pl-117B
which wasbordered on the left
by
aHpaI
site and on theright by
aHindlllsite
(Fig.
1).
Clone
pl-121
showedoptimal
origin activity
inourtrans-fection assay. Since
pl-117B (the
smallestorigin-positive
fragment)
represents the left 1.0 kb ofpl-121
(Fig.
1),
theBGL SMA SMA SMA PAT
N 5 1 2 153
FIG. 2. Origin analysis of STD EHV-1 clones homologous to
sequences present in DI DNA. Individualfragments wereinserted into thevectorpAT153 (orpAT12)andanalyzedfororigin activity
by transfection into L-M cells. All cell DNAs (10to 15 ,ug) were
digestedwithEcoRItolinearize transfectedDNAs,electrophoresed
through0.8%agarose, and blottedtonitrocellulose. Nitrocellulose filterswereprobedwith pAT153DNAwhichhadbeen
32p
labeledbynick translation. The individualclones usedareshownatthe top.
Duplicateflasks ofcellswereused whichwereeither mock infected (-)orvirus infected(+).The mapunits ofSma-1are0.83to0.85.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.368.518.505.626.2]-; 1X- lV- 1t1 119 120 121 . ._
:w- r'~ t8r m rln -tyl ril Vc
117B7 1_.J.. 12 .Is 12 SMA
E EE E E E E E F E E
EDMDEDMDEDMDEDMC dlb1 ld17 dIB dil
T m m m
d16 d17 d 18 dll
E E EE E E E E
E D M E DMI E D M E DM
..
a~
0 40 .,w
.0 ,
[image:5.612.55.298.71.195.2]A B
FIG. 3. Origin analysis of subclones of the Sma-1 DNA frag-ment.(A) Transfected-cell DNAs (10 to 15 ,ug) were digested with EcoRI, electrophoresed in 0.8% agarose, and transferred to nitro-cellulose. The filter was probed with vector DNA which had been 32P labeled by nick translation. The individual clones used for transfection are shown at the top. Lanes are designated m for mock infected or i for infected-cell monolayers. Clone pl-117B is desig-nated by a star (*). pl-117 contains a two-copy insert. pl-117B contains a single-copy insert of the same fragment. v, EHV-1 genomic DNA digested with EcoRI; c, positive control from an
infected-cell lysate which was transfected with Sma-1. (B) DpnI-MboI analysis of Sma-1 subclones. Cell DNAs from infected-cejl lanes (i) in panel A which were positive fororigin activity were digested withEcoRI(E),EcoRIandDpnI(ED), orEcoRIandMboI (EM).Inlanes ED marked bystars (*) exogenous vector DNA was mixed with cell DNAs before digestion as a control to ensure that DpnI was active. Total-cell DNAs were analyzed by Southern blot analysisasin panelA.
exonuclease BAL 31 was used to delete sequences from the HpaI site located at the left end of pl-121. Four clones pl-121dl6, dl7, d18,and dll werecharacterized (Fig. 1and Table 1). All four clones were origin positive (Fig. SA),
including pl-121 dll, which has approximately 625 bp
de-leted from the left end. DpnI-MboI analysis (Fig. SB)
con-firmedthat all clones hadreplicated.Thesestudiessuggested
that EHV-1 origin activity could be localized to the
right-ward 400 bp ofclone pl-117B. Therefore, a new series of
_14
so
so
A * ,.
B
FIG. 5. Origin analysis of BAL 31 deletion constructs ofpl-121. Deletions were constructedinpl-121 by using BAL 31 as described in Materials and Methods. (A) After transfection, cell DNAs (10 to 15 ,ug) were digested with EcoRI, electrophoresed through 0.8% agarosegels,andblottedtonitrocellulose. Filters were probedwith pAT153 which had been 32P labeled by nick translation. m, Mock infected; i, infected. (B) Cell DNAs from infected-cell (i) lanes were digested with EcoRI (E), EcoRI and DpnI (ED), or EcoRI and MboI (EM) and then weresubjected toSouthern blot analysis as in panel A.
cloneswasdevelopedwhichcenteredonthis400-bp region. These clones included 127, 128, 131, 133, pl-134, pl-135, andpl-136. As shown in Fig. 6 (also see Fig. 1 and Table 1), the smallest cloneshowing origin activity was the203-bpclone pl-133 (0.840to0.841mapunits) which is bordered onthe leftbyanSpeI site and on the right by a ClaI site. The origin-positive clones 128, 131, 134, pl-135, and pl-136 all contain clone pl-133 sequences as a subsetoftheir total DNAsequences. InFig. 6, cell DNAsin all lanes(includingthemock-infected-cell lanes)were double digested with EcoRI and DpnI. Only total-cell DNAs iso-lated from EHV-1-infected cells (Fig. 6, lanes i) contained DpnI-resistant plasmid DNA (i.e., DNA which has repli-cated ineucaryotic cells).
DNA sequenceanalysis of theEHV-1
oris.
The EHV-1STDDNAfragmentcorrespondingtothe minimaloriginof
repli-pUJC pAT
Vr'fla Sitia1 117B 120 121
tl1i T.n m m V
A
117B rmi 1175..EE 120 121
[image:5.612.317.551.73.217.2] [image:5.612.56.297.493.610.2] [image:5.612.326.545.504.660.2]EE .__ E..._.W....-_..._._ ... ...
E E E E E E Ef-'
E D MWE DM E D M: E D
\^-B
134 135 136 dlB
n m m m
131 132 133 128
m m m m
_'.
_
00
.0 FIG. 4. Origin analysis of subclones pl-117B, pl-120, and pl-121.(A) Total-cellDNAswereisolated aftertransfection,and10to
15 ,ug was digested with EcoRI, electrophoresed through 0.8% agarose, and blottedtonitrocellulose.Thefilterwashybridizedwith pAT153 which had been32plabeledbynicktranslation. Clones used for transfectionarelistedatthe top ofeachsetoflanes.Sma-1was
cloned into eitherpAT153(Sma-1pAT)orpUC12 (Sma-1pUC).V, EHV-1 genomic DNA digested with EcoRI; mock infected; i, infected. (B) CellDNAs frompanelAinamountsof 10to15jigwere
digestedwithEcoRI(E), EcoRIandDpnI(ED),orEcoRIandMboI (EM). CellDNAswereanalyzedbySouthernblothybridizationas
described above.
B
A
I..
FIG. 6. Origin analysisof Sma-1 subclones. The200-bppl-133 clonehas origin activity. Aftertransfection, cell DNAs were iso-latedand10to15,ugweredigestedwithEcoRIandDpnI. Samples
wereelectrophoresedina0.8% agarosegelandblottedto nitrocel-lulose. FilterswerehybridizedwithpAT153 DNAwhichhad been
32p
labeledby nick translation. m, Mockinfected;i, infected.on November 10, 2019 by guest
http://jvi.asm.org/
A
ATCGATTATCGGACGAAAATTGGAAACGCGTCCCGTGGCACAAATCCTGCACCCTGATTG 60 GCCCAGAGGCCCGTTCGCACCAATCACCAATAAGTTTTAATAATAATTATTGCAACAAAG 120
t
TGCGAACACTACGTGTTCGCACTTCTTATCCGTTCCACGCCCCCACCCCCCATCTCGGGA 180
GCGCGCATGGCACCGTGCCAACTAGT
6350
Hind III
B
SSmaI
IRs
206
8200 CIa I
R
il
11250bp Sma I
Jr
Il
USI-AATAAA IE 6.0 kb TATAA ORI VE
1.2 kb
FIG. 7. DNAsequenceand genomic location of the EHV-1origin. (A) TheDNA sequenceof theClaI-SpeI pl-133clonewasdetermined by the dideoxy-chaintermination method. Arrows show the beginning and end of themajorregionofhomologybetween theEHV-1origin andthe human herpesvirus origins (Fig. 8). The conserved 9-bp sequence CGTTCGCAC, also identified in the human lytic origins of replication(Fig. 8), is indicated. (B)Schematicdiagram showing theorganization of theinternal EHV-1 IR. The single6.0-kilobase (kb) EHV-1 immediate early (IE) gene is shown. The putative transcriptional start andpolyadenylation sites have been identified by DNA sequencing(Grundyetal.,manuscript inpreparation).Thelocationof the veryearlygene(VE)and thereplicationoriginarealsogiven.An areaofrepeats(R)identified byDNAsequencing (Yalamanchilietal.,inpreparation)isshowntotherightoftheoriginofreplication.Base pairs startingat theleftward Sma-1 site, whichmarks(approximately) thebeginning oftheIR,areshownatthetop.
cation (pl-133)wascloned into M13, andboth strands were sequenced(Fig.7A). The EHV-1 pl-133 DNA sequence was
comparedwith thepublished sequences of the known human herpesvirus origins of replication (23, 34, 58, 63, 65) using the BIONET national computer resource for molecular
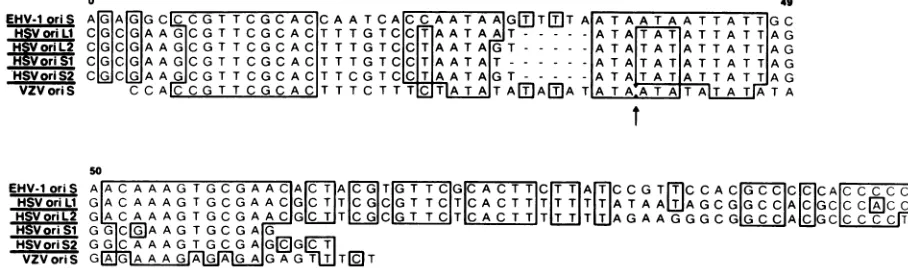
biology (26). Significant homology was demonstrated be-tween allfive humanherpesvirusoriginsand the EHV-1
oris
between bp65 and bp 170 (Fig. 8). The percent homology overthis 100-bp region ranged from 50% with VZV
oriS
to60% with the HSV origins. Interestingly, alignment of ho-mologous regions (Fig. 8) revealed that the 9-bp sequence
(CGTTCGCAC) which is conserved in all five herpesvirus origins is present in the EHV-1 ori sequence. This 9-bp
sequence is a subset of the 11-bp sequence (CGTTCG CACTT) noted by Stow and Davison(58) to beconserved in
all human herpesvirus origins. It is also within the region
shown to be protected in HSV
oris,
by an origin-bindingprotein(13). Apparently, this9-bp sequenceisrequisite for
origin function and is conserved even acrossherpesviruses
ofdifferent species, horseto human.
Unlike otherherpesvirus origins, the EHV-1 origin does not contain a near-perfect palindrome. Within the area of homology between the EHV-1 origin and the human herpes-virus origins,apartial stem-and-loopstructure canbedrawn (Fig. 9). Whether this imperfect cruciform structure exists in vivo remains to be proven.
DISCUSSION
We have described the functional mapping and DNA sequencing of an EHV-1 origin of replication. Previous studies from this laboratory demonstrated that EHV-1 DI DNA originates from within the left terminus (0.00 to 0.04 map units) and the IRs (0.78 to 0.79 and 0.83 to 0.87 map units; 0.91 to 0.95 and 0.99 to 1.00 map units) of the STD EHV-1 genome (1-3). STD EHV-1 plasmid clones
contain-0 49
EHV-1 oriS AG A G GCOO G T TCG CAC CAATCA
CCAATAAAG3TIf]T
AATAATAATTA TT GC HSVoriLI CGC G AAGOG TTCGCAC TTTGTC CAATAAT- TA TAT ATATA TTAG HSoriL2 CIGICGIAA GCGT TC GCACTTTGTOCICTIAATA GT- - ATATATATTATTAGHSori ICGICGAAAGCGTTCGCACTTTGTCOCITIAATAIT--- ATAITATIATTATTIAG
HSVoriS2 COJAGAAGCOGT TCGCACTTCGTC C TAATAG T-- ATA T A TAT T TAG VZVoriS CCALCOG T TCGCACTTTCTT T TATA
AE)AA
TAATA TAT A TAATA50 110
EHV-1 oriS A AC AAAGTGCGAACA[T A TGTTCG ACTTT A TO GTCCAC GCCC CACCCCCCATc HSVoriUL GACAAAGTGCGAACOGTT GC GTTC TCA CTT TTT T TTATAAUTAGCGGCCA
OGCCCE)CCGG
CT HSVoriL2 GACAAAGT GC GAAC G T TC CGTTCT CACTTT TWTTAGAAG
GGOC G ACCCG T HSVoriS1 GGCDAAGTGCGA GHSVoriS2 GGC AAA G TGC G AGG l
VZV oriS GAG A AA G AGAGAGAGT QTT
FIG. 8. Comparison of homology among the herpesvirus lytic origins of replication. DNA sequences were taken from Murchie and McGeoch (34;
oris,),
Whitton and Clements (65;oris2),Weller et al. (63;oriL,),
Lockshon and Galloway(28;oriL2),
andStow and Davison (58;VZVoris).Homologieswereidentified by using the BIONET computer system. Boxed regions indicate areas of homology. Horizontal linesindicategaps that were introduced to give optimal alignment. Arrows designate a removal of 9 bp (TATATATAT) from the VZVoriS
sequence to permit maximal alignment.
IE
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.94.534.80.240.2] [image:6.612.70.524.536.671.2]5'
C C A AC A c GG T T C G C A C C C A A T A A T T T T A
I I I I I I I I I I I I I I I A
3* c A C A A G C G T G A A AC A C G T T A T T A
AT,A
A T129
FIG. 9. Hypothetical stem-and-loop structure for EHV-1 oriS. Shown is the stem-and-loop structurethat canbe drawn in thearea of
maximalhomology between theEHV-1orisand the human herpesvirus lytic origins of replication: bp 71to129.Verticallines designateareas ofpotential basepairing.
ing DNA fragments from regions shown to be present in EHV-1 DI DNA were assayed for origin activity in
transfec-tion assays. Of these clones, only a clone from the central
portion oftheIRs(Sma-1; 0.83 to 0.85 map units) was shown to have origin activity. A series of subclones of Sma-1 was generated and was used in transfection studies to demon-strate thatoriginactivity is contained within a 200-bp frag-ment (0.840 to 0.841 map units). Comparison of the DNA sequenceof this200-bpfragment to those of human herpes-virus origins demonstrated that substantial homology exists among them, including a 9-bp sequence (CGTTCGCAC)
previouslynoted tobe conserved among the human herpes-virus origins of replication (58). To our knowledge, this work represents the first description of the location and DNA sequenceofaherpesvirus replication origin other than those
of a human herpesvirus. Thus, it appears that between
herpesviruses ofdifferent species there is significant
conser-vation of DNA sequences in viral origins of DNA
replica-tion. Structurally,thesimilarities between the EHV-1 origin and the human herpesvirus origins appear restricted, in that the EHV-1 origin does not contain a near-perfect palin-drome. Although, a stem-and-loop structure can be pro-posed for the EHV-1 origin, the propro-posed structure has a 7-bp region in its center which does not have
complemen-tarity forthe formation of a stem-loop structure; thus, one would presume that the formation of this hypothetical EHV-1stem-and-loop structure would not be as favored as that of the near-perfect stem-and-loop structures proposed for the human herpesviruses. Interestingly, a recent study reports that anHSV-1
oriS
clone from which the right side of thepalindrome was deleted retains origin activity (11). These results suggest that the ability to form a stem-and-loop structure maynotbe essential toorigin activity.
Electronmicroscope visualization ofreplicatingHSV ge-nomes (14, 22) and molecular analysis of HSV DIparticles
(15) support thefunctional importance of the HSV
oris
andoriL
inproductive infections. However, recent studies haveindicated that the oriL andat leastonecopy of the
oris
aredispensable forHSV-1 biological activity (29, 41). Further-more,deletion oftheoriL,wasfoundnot toaffect theability of HSV-1 to establish latency in the murine eye model.
Interestingly, the HSV
oris
hasbeenshowntobecontainedin a transcribed open reading frame (24), an observation
which opens the possibility that trans-acting regulatory
factors encoded by the origin
region
may act to modulateorigin activity. The importanceof interaction between
rep-lication origins and transcriptional elements in regulating
viralgrowth, although postulatedforHSV,hasbeenclearly substantiated in other viral systems (7, 12).
Indeed,
others have noted some homology amongthe HSVorigin
and the adenovirus and papovavirus origins (25). Of course, thelocations oftheHSVoriLand
oris
sequencesaresuggestiveof a role in transcriptional regulation. The HSV
oriL
is flanked by the coding regions for the major DNA-binding protein (ICP8) and the DNA polymerase, while theoriS
is bracketedby the IE genes for ICP4 andICP22/47.Similarly, theEHV-1oris
is located in the centerof theIRS
where the single EHV-1 IE gene and the EHV-1 very early gene have been mapped (Fig. 7B) (8, 18, 44). Furtherinvestigation will be necessary to define the exact functional and regulatory properties of the herpesvirus origins.The studies described here are part of our overall en-deavor to characterize the genetic elementsthat constitute EHV-1 DI DNA. Ultimately, we hope that understanding STD and defective EHV-1 DNA at the molecular level will enable us to decipher how DI particles participate in the establishment ofapersistent infection of permissive cellsby STD EHV-1 (10, 37). Certainly, regulation of replication origin activity might beamajorcomponent in the establish-mentof apersistent infection. Currently, we are cloning and
sequencing potentialorigins from cloned EHV-1DI DNA to determine iftheyarecomplementary to the known sequence of the STD EHV-1
oriS
(Yalamanchilietal.,inpreparation).ACKNOWLEDGMENTS
We thankAngela Houston andCynthia Harrison for typing the manuscript. We thanktoFrankGrundy and Jackie Fetherstonfor helpfulsuggestions.
Support for this investigation wasobtained from Public Health Service research grant Al 22001 from the National Institutes of Health,aGrayson FoundationInc.researchgrant, a grantfrom the Louisiana State University System Biotechnology Institute, and grant 86-CRCR-2257 from the U.S. Department of Agriculture Biotechnology Program. R.P.B. is a recipient of a postdoctoral fellowship fromthe American Heart Association-Louisiana Chap-ter, Inc.
LITERATURE CITED
1. Baumann, R. P., S. A. Dauenhauer, G. B. Caughman, J. Staczek, and D. J. O'Callaghan. 1984. Structure and genetic complexityof the genomes ofherpesvirusdefectiveinterfering particles associatedwithoncogenic transformationand persist-entinfection. J.Virol. 50:13-21.
2. Baumann, R. P., J. Staczek, and D. J. O'Callaghan. 1987. Equine herpesvirus type 1 defective-interfering (DI) particle DNA structure: the central region of the inverted repeat is deleted from DI DNA.Virology 159:137-146.
3. Baumann, R. P., J. Staczek, and D. J. O'Callaghan. 1986. Cloningand finemappingthe DNA ofequineherpesvirustype
onedefective interfering particles. Virology153:188-200.
4. Baumann, R. P., D.C. Sullivan, J. Staczek, andD.J. O'Cal-laghan. 1986. Genetic relatedness and colinearity of the
ge-nomes of equine herpesvirus type one and three. J. Virol. 57:816-825.
5. Cameron,I. R.,N. M. Wilkie, andJ. M. MacNab. 1983. The infectivityofherpes simplex virus DNA inratembryocells is
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.94.527.109.159.2]enhanced synergistically byDMSO and glucose. J.Virol. Meth-ods 6:183-191.
6. Campbell, D. E., M. C. Kemp, M. L. Perdue, C. C. Randall, and G. A. Gentry. 1976. Equine herpesvirus in vivo: cyclic produc-tion of a DNA density variant with repetitive sequences. Virol-ogy 69:737-750.
7. Campbell,J. L. 1986. Eukaryotic DNA replication. Annu. Rev. Biochem. 55:733-771.
8. Caughman, G. B., A. T. Robertson, W. L. Gray, D. C. Sullivan, and D. J. O'Callaghan. 1988. Characterization ofequine herpes-virus type 1 immediate early proteins. Virology 163:563-571. 9. Challberg,M. D. 1986. A method foridentifying the viral genes
required for herpesvirus DNA replication. Proc. Natl. Acad. Sci. USA 83:9094-9098.
10. Dauenhauer, S. A., R. A. Robinson, and D. J. O'Callaghan. 1982. Chronic production of defective-interfering particles by hamster embryo cultures of herpesvirus persistently infected andoncogenically transformed cells. J. Gen. Virol. 60:1-14. 11. Deb, S., and M. Doelberg. 1988. A 67-base-pair segment from
theOri-S region of herpes simplex virus type 1 encodes origin function.J. Virol. 62:2516-2519.
12. DePamphilis, M. L. 1988. Transcriptional elements as compo-nents of eukaryotic origins of replication. Cell 52:635-638. 13. Elias,P., M. E. O'Donnell, E. S. Mocarski, andI. R. Lehman.
1986.ADNA binding protein specific for an origin of replication ofherpes simplex virus type one. Proc. Natl. Acad. Sci. USA 83:6322-6326.
14. Freidmann, A., J. Shlomai, and Y. Becker. 1977. Electron microscopy of herpes simplex virus DNA molecules isolated frominfectedcells by centrifugation in CsCl density gradients. J. Gen. Virol. 34:507-522.
15. Frenkel, N. 1980. Defective interfering herpesviruses, p. 91-120. In A. J.Nahmias, W. R. Dowdle, and R. F. Schinazi (ed.), The human herpesviruses. Elsevier Science Publishing, Inc., New York.
16. Graham,R. L., and A. J. van der Eb. 1973. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 5:456-462.
17. Gray,C. P., and H. C. Kaerner. 1984. Sequence of the putative originofreplication in the UL region of herpes simplex virus type1 ANGDNA.J. Gen. Virol. 65:2109-2119.
18. Gray, W. L., R. P. Baumann, A. T. Robertson, D. J. O'Cal-laghan, and J. Staczek. 1987. Characteristics and mapping of equine herpesvirus type 1 immediate early, early and late transcripts. Virus. Res. 8:223-244.
19. Henry,B. E., W. W. Newcomb, and D. J. O'Callaghan. 1979. Biological and biochemical properties of defective interfering particlesofequine herpesvirus 1. Virology 92:495-506. 20. Henry, B. E., W. W. Newcomb, and D. J. O'Callaghan. 1980.
Alterationsin viral protein synthesis and capsid production in infection with DI particles of herpesviruses. J. Gen. Virol. 47:343-353.
21. Henry,B. E., R. A. Robinson, S. A. Dauenhauer, S. S. Atherton, G. S. Hayward, and D. J. O'Callaghan. 1981. Structure of the genomeofequineherpesvirus type 1. Virology 115:97-114. 22. Hirsch,I., G. Cabral, M. Patterson, and N. Biswal. 1977. Studies
on the intracellular replicating DNA of herpes simplex virus type 1.Virology81:48-61.
23. Holmes,D. S., and M.Quigley. 1981. A rapid boiling method for the preparation of bacterial plasmids. Anal. Biochem. 114: 193-200.
24. Hubenthal-Voss,J., L. Starr, and B. Roizman. 1987. The herpes simplex originsofDNA synthesis in the S component are each contained in a transcribed open reading frame. J. Virol. 61: 3349-3355.
25. Knopf, C. W., B. Spies, and H. C. Kaerner. 1986. The DNA replication origins of herpes simplex virus type 1 strain An-gelotti DNA. NucleicAcids Res. 14:8655-8667.
26. Kristofferson, D. 1987. The BIONET electronic network. Na-ture(London)325:555-556.
27. Lacks, S., and B.Greenberg. 1977. Complementary specificity ofrestriction endonucleases of Diplococcus pneumoniae with respecttoDNA methylation. J. Mol. Biol. 114:153-168.
28. Lockshon, D., and D. A. Galloway. 1986. Cloning and charac-terization of
ori,.2,
a largepalindromic DNAreplicationorigin of herpes simplex virus type 2. J. Virol. 58:513-521.29. Longnecker, R., and B. Roizman. 1986.Generationofan invert-ing herpes simplex virus 1 mutant lacking the L-Sjunction a sequences, an origin of DNA synthesis, and several genes including those specifying glycoprotein E and thea47 gene. J. Virol. 58:583-591.
30. Lusky, M., and M. Botchan. 1981. Inhibition of
SV40
replication in simian cells by specific pBR322 DNA sequences. Nature (London) 293:79-81.31. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a laboratory manual. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.
32. McGeoch, D. J., M. A. Dalrymple, A. Dolan, D. McNab, L. J. Perry, P. Taylor, and M. D.
Challberg.
1988. Structures of herpes simplex virus type 1 genes required for replication of virus DNA. J. Virol. 62:444-453.33. Mocarski, E. S., and B. Roizman. 1982. Herpesvirus-dependent amplification and inversion of cell-associated viral thymidine kinase gene flanked by viral A sequences and linked to anorigin of viral DNA replication. Proc. Natl. Acad. Sci. USA 79: 5626-5630.
34. Murchie, J. J., and D. J. McGeoch. 1982. DNA sequence analysis of an immediate-early gene region of the herpessimplex virus type 1 genome. J. Gen. Virol. 62:1-15.
35. O'Callaghan, D. J., G. P. Allen, and C. C. Randall. 1978. The equine herpesviruses, p. 1-32. In J. T. Bryan and H. Gerber (ed.), Equine infectious diseases, vol. 4. Veterinary Publica-tions, Princeton, N.J.
36. O'Callaghan, D. J., W. P. Cheevers, G. A. Gentry, and C. C. Randall. 1968. Kinetics of cellular and viral DNA synthesis in equine abortion (herpes)virus infection of L-M cells. Virology 36:104-114.
37. O'Callaghan, D. J., G. A. Gentry, and C. C. Randall. 1983. The equine herpesviruses, p. 215-318. In H. Fraenkel-Conrat and R. R. Wagner (ed.), The herpesviruses, vol. 2. Plenum Publish-ing Corp., New York.
38. O'Callaghan, D. J., B. E. Henry, J. H. Wharton, S. A. Dauen-hauer, R. B. Vance, J. Staczek, and R. A. Robinson. 1981. Equine herpesviruses: biochemical studies on genomic struc-ture, DI particles, oncogenic transformation, and persistent infection, p. 387-418. In Y. Becker (ed.), Developments in molecular virology, vol. 1. Herpesvirus DNA. Martinus Nijhoff Publishers, The Hague, The Netherlands.
39. O'Callaghan, D. J., J. M. Hyde, G. A. Gentry, and C. C. Randall. 1968. Kinetics of viral deoxyribonucleic acid, protein, and infectious particle production and alterations in host mac-romolecular synthesis in equine abortion (herpes) virus-infected cells. J. Virol. 2:793-804.
40. Perdue, M. L., M. C. Kemp, C. C. Randall, and D. J. O'Cal-laghan. 1974. Studies of the molecular anatomy of the L-M cell strain of equine herpesvirus type 1: proteins of the nucleocapsid and intact virion. Virology 59:201-216.
41. Polvino-Bodnar, M., P. K. Orbert, and P. A. Schaffer. 1987. Herpes simplex virus type 1 oriL is not required for virus replication or for the establishment and reactivation of latent infection in mice. J. Virol. 61:3528-3535.
42. Razin, A., H. Cedar, and A. D. Riggs (ed.). 1984. DNA methyl-ation: biochemistry and biological significance, p. 1-10. Spring-er-Verlag, New York.
43. Rigby, P. W., M. Dieckmann, C. Rhodes, and P. Berg. 1977. Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase 1. J. Mol. Biol. 113:237-251.
44. Robertson, A. T., G. B. Caughman, W. L. Gray, R. P. Baumann, and D. J. O'Callaghan. 1988. Analysis of the in vitro translation products of the immediate early RNA of equine herpesvirus type 1. Virology 106:451-462.
45. Robinson, R. A., B. E. Henry, R. G. Duff, and D. J. O'Cal-laghan. 1980. Oncogenic transformation by equine herpesvi-ruses (EHV). I.Properties of hamster embryo cells transformed by UV-irradiated EHV-1. Virology 101:335-362.
on November 10, 2019 by guest
http://jvi.asm.org/
46. Robinson, R. A., and D. J. O'Callaghan. 1981. The organization ofintegratedherpesvirus DNA sequences in equine herpesvirus type 1 transformedand tumorcells, p. 419-436. In Y. Becker (ed.), Developments inmolecular virology, vol. 1. Herpesvirus DNA. Martinus Nijhoff Publishers, The Hague, The Nether-lands.
47. Robinson, R. A., and D. J. O'Callaghan. 1983. A specific viral DNAsequence is stably integrated in herpesvirus oncogenically transformed cells. Cell 32:569-578.
48. Robinson, R. A., P. W. Tucker, S. A. Dauenhauer, and D. J. O'Caflaghan. 1981. Molecular cloning of equine herpesvirus type 1 DNA: analysis of standard and defective viral genomes andviral sequences in oncogenically transformed cells. Proc. Natl. Acad.Sci. USA78:6684-6688.
49. Robinson, R. A., R. B. Vance, and D. J. O'Callaghan. 1980b. Oncogenictransformation by equine herpesviruses. II. Coestab-lishment of persistent infectionand oncogenictransformation of hamster embryocells byequineherpesvirus type 1 preparations enriched fordefective interfering particles.J.Virol. 36:204-219. 50. Ruyechan, W. T., S. A. Dauenhauer, and D. J. O'Callaghan. 1982. Electron microscopicstudy ofequine herpesvirus type 1 DNA. J.Virol.42:297-300.
51. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
52. Sharp, P. A., B. Sugden, andJ.Sambrook. 1973. Detectionof two restrictionendonuclease activities in Haemophilus parain-fluenzae using analytical agarose-ethidium bromide
electropho-resis. Biochemistry12:3055-3063.
53. Southern, E. M. 1975. Detection ofspecific sequences among DNAfragments separated bygelelectrophoresis. J.Mol. Biol. 98:503-517.
54. Spaete, R. R., and N. Frenkel. 1982. Theherpes simplex virus amplicon: anew eucaryotic defective-virus cloning-amplifying vector.Cell 30:295-304.
55. Spaete, R. R., and N. Frenkel. 1985. Theherpes simplexvirus amplicon: analyses of cis-acting replication functions. Proc. Natl.Acad.Sci. USA82:694-698.
56. Stow, N. D. 1982.Localization ofanorigin ofDNAreplication within the
TRA/IRS
repeatedregion of the herpessimplexvirustype1genome. EMBOJ. 1:863-867.
57. Stow, N. D. 1985. Mutagenesis of a herpes simplex virusorigin of DNAreplication anditseffectonviralinterference. J. Gen. Virol. 66:31-42.
58. Stow, N.D.,andJ.Davison. 1986. Identification ofa varicella-zoster virus origin of DNA replication and its activation by herpes simplex type 1 gene products. J. Gen. Virol. 67:1613-1623.
59. Stow, N. D.,and E. C. McMonagle. 1983. Characterizationof theTRS/IRSorigin of DNA replication of herpes simplex virus type1.Virology130:427-438.
60. Twigg, A.J., and D. Sheratt. 1980.Trans-complementable copy number mutants of plasmid Col El. Nature (London) 283: 216-218.
61. Vieira, J.,andJ. Messing. 1982.ThepUC plasmids,aM13mp7 derived system for insertion mutagenesis andsequencing and syntheticuniversalprimers. Gene 19:259-268.
62. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes simplexvirus DNA: localization of replication recognition sig-nals within defective virus genomes. Proc. Natl. Acad. Sci. USA 78:742-746.
63. Weller, S.K.,A.Spadaro, J.E.Schaffer,A.W. Murray,A. M. Maxam, and P. A. Schaffer. 1985. Cloning, sequencing, and functionalanalysis of ori-L,aherpessimplex virustype1origin of DNA synthesis. Mol. Cell. Biol. 5:930-942.
64. Whalley, J. M., G. R. Robertson, and A. J. Davison. 1981. Analyses of thegenomeof equineherpesvirustype 1: arrange-mentofcleavageforrestriction endonucleasesEcoRI,BglIland BamHI. J. Gen. Virol. 57:307-323.
65. Whitton, J. L., and J. B. Clements. 1984. Replication origins and asequenceinvolved in coordinate induction of the immediate earlygenefamilyareconserved inanintergenic region of herpes simplex virus. Nucleic Acids Res. 12:2061-2079.
66. Wu, C. A., L. Harper and T. BenPorat. 1986. Cis functions involved inreplication andcleavage-encapsidationof pseudora-bies virus. J. Virol.59:318-327.
67. Wu, C. A., N.J.Nelson,D.J. McGeoch,and M. D.Challberg. 1988. Identification ofherpes simplex virus type 1 genes re-quired fororigin-dependent DNA synthesis. J. Virol. 62:435-443.