Four
Major Sequence
Elements
of
Simian
Virus
40 Large
T
Antigen
Coordinate Its
Specific and Nonspecific
DNA
Binding
DANIELT. SIMMONS,l 2* GERHARD LOEBER,2t AND PETERTEGTMEYER2
Schoolof Life and Health Sciences, University ofDelaware, Newark, Delaware 19716,1* andDepartment of
Microbiology, State University ofNew York, Stony Brook, New York 117942
Received 17October 1989/Accepted 23January 1990
Bymutational analysis, wehave identifiedamotif critical tothe properrecognitionandbinding ofsimian
virus40 large tumorantigen (Tantigen)tovirus DNAsequencesattheorigin of DNA replication. This motif
is tripartiteandconsistsoftwoelements (termed Al and B2) thatarenecessaryfor sequence-specific binding
ofthe originandacentral element (Bl) which is required fornonspecificDNA-binding activity. Certain amino
acids inelements Al (residues 152 to 155) and B2 (203to207) may make directcontactwith the GAGGC
pentanucleotide sequences in binding sites I and II on the DNA. Alternatively, these two elements could
determinetheproperstructureof the DNA-binding domain, although foranumberofreasons wefavor thefirst
possibility. In contrast,elementBi (183to 187) ismostlikely important for recognizinga generalstructural
featureofDNA. Elements Al and B2arenearlyidentical in all known papovavirus T antigens, whereas Bl is
identical onlyin the closely related papovaviruses simian virus 40, BK virus, and JC virus. In additiontothese three elements,afourth(B3; residues215to219)isnecessaryforthebindingof T antigentositeIIbutnot to siteI.WeproposethatadditionalcontactsitesonTantigenareinvolvedin theinteractionwith site IItoinitiate thereplication oftheviral DNA.
The simian virus40(SV40) largetumorantigen(Tantigen)
is a 708-amino-acid phosphoprotein that is involved in the initiation and elongation stages ofSV40 DNA synthesis in infected monkey cells. Its role in DNAreplication is begin-ningtobeunderstood. ItappearsthatTantigen first bindsto
the pentanucleotide sequences GAGGC in sites I and II closeto thereplication origin (10, 12, 20, 47,48) and melts theDNA intheproximity ofsiteII (4). AfterDNAsynthesis
has begun, Tantigen mayfunctionas ahelicase tounwind
theparentalDNAstrands (8, 14, 43, 49).
Themechanism bywhichpapovavirus Tantigens bindto the sequences at the replication origin is not known. Over
thelast fewyears,thedomainresponsiblefor DNAbinding
hasbeenidentifiedbygenetic (7, 32, 34)andbiochemical(39) approaches. However, little orno similarityexists between
thisdomain and that of well-studiedDNA-binding proteins,
so noobvious modelpresentlyexists toexplain thebinding
to DNA.
There is considerable evidence which implicates the
re-gionbetween residues 140 and 260 inorigin binding (1, 40, 46). A Zn2+ finger motif between residues 301 and 321
appears to contribute to the stability of the DNA-protein complex (1, 40), althoughitisnotrequiredfor DNAbinding perse.
Severalgroupshavegeneratedmutantsof Tantigenwith altered binding activity for the origin. With one notable
exception(32), however, their approach has been to study only one or a few mutants within this region, and conse-quently there is very little information about the overall
organizationof theDNA-bindingdomain. We have starteda
systematicandthorough mutagenesisof this domain in order to identify the sequences thatare important forori-binding
activity. The aim here is twofold. One goal is to identify
aminoacid residues that makecontactwithsequencesatthe
* Corresponding author.
t Present address: Ernst Boehringer Institute, Bender-AG, Dr.
Boehringergasse 5-11,A-1121Vienna, Austria.
origin, and thesecond is tofind theresidues thatarecrucial
for the proper structure (and therefore function) of this
domain.With thisinformation,amodel of DNAbindingmay emerge.
Oursurveyhasrevealed that theDNA-bindingdomain has
acomplex organization. By testing the effectsofsingle-site
mutations onvirus replicationand by performinga number
of DNA-binding assays on replication-negative T-antigen mutants, we have identified several important sequence
elementswithinthisdomain. This information has been used todevelopabasicmodelthatdescribes the interaction of the
domain withorigin andnonorigin DNA. MATERIALS AND METHODS
Plasmids. pBS-SV40 contains the entire SV40 genome
inserted into the BamHI site of Bluescript (Stratagene). pSKATis a plasmidderived in our lab from pIA#4 (a gift
from Y. Gluzman). It contains the SV40 T-antigen gene
insertedbetween adenovirus type 5 map units 0 to 1.4 and
the major late promoter of adenovirus type 2. Additional adenovirus sequences (map units 9 to 15.5) are present to allow for recombination with homologous sequences in a
large adenovirus fragment (map units 4 to 100). pSVO+ contains thewild-type SV40 origin consisting of sitesI and
II; pOS1contains site I only,and pSVOdl3 contains site II
only (44).
Mutagenesis protocol. Mutations were generated in pBS-SV40orpSKAT by annealing oligonucleotides withasingle
mismatchtoauridine-containing single-strandedDNA
tem-plate aspreviously described (23).The oligonucleotide was
extended with T4 DNApolymerase,and the resulting dou-ble-stranded DNAwas used to transform Escherichia coli BMH 71-18 (International Biotechnologies, Inc.).
Single-stranded DNA was sequenced by the dideoxy procedure (37). In somecasesthe mutantTantigengenewasrecloned from pBS-SV40to pSKAT by standard recombinant DNA
procedures.
Virus replication assays. pBS-SV40 harboring a mutation 1973
on November 10, 2019 by guest
http://jvi.asm.org/
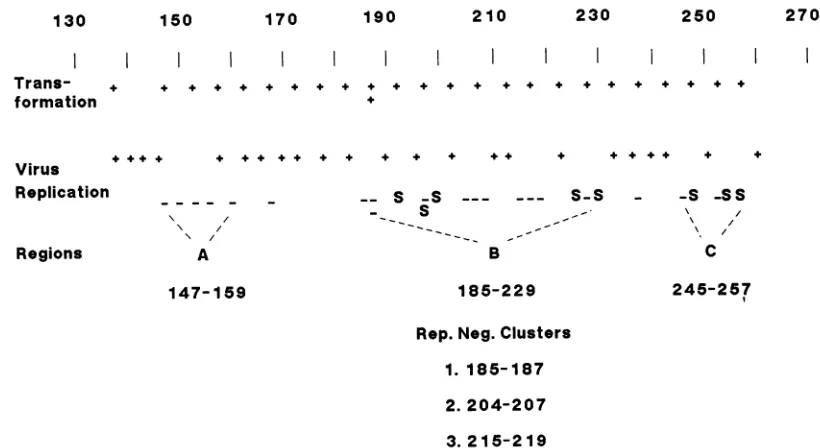
130 150 170 190 210 230 250 270
Trans- + + + 4 + + + + + + 4 4 + + + + 4 4 + + + + 4 +
formation +
.4.4.4. 4. +4.4.4. + 4. 4. 4. 4. 4.4.
-- S -S -_
--_ S
B 185-229
S-S _ -S -SS
\ /
\ / \~/
C 245-257
Rep.
Neg.Clusters 1. 185-187 2. 204-207 3.215-219FIG. 1. Plaque formation and transformationassaysofT-antigenmutants.Single-pointsubstitution mutations in theDNA-bindingdomain
weretested for their effectsonvirus multiplication in monkey cells. Symbols:+,mutations that resulted in replication like that of wild-type SV40; S, mutations thatgave rise to smallplaques; -, mutations that resulted in noreplication. Mutations that had an effecton virus replicationareshownon aseparateline from those that hadnoeffect. Mutants with altered replication propertieswerearrangedinto three
groups(A, B, andC), with the centralgroup(B) containing three clusters ofreplication-negative (rep. neg.)mutants. Some of the mutants
werealso tested for the abilitytotransform primary mouse embryo cells (transformation +).Insome cases, wetestedtwomutantswith different amino acidsubstitutionsatthesamesite.
inthegeneforTantigenwascleavedwith BamHItorelease
the mutant genomic DNA. The DNA was ligated at low
DNAconcentrationstofavor the formation of circularDNA and thentransfected into monkey cells (CV-1 orBSC-1) by
the DEAE-dextran procedure (27) as previously described
(23). Plaques were counted 10 to 30 days posttransfection, dependingupon plaque size. Plates which didnothave any plaqueswere incubated for aminimum of 30 days to make surethat smallplaques did notappear.
Transformation assays.Ligated circular SV40 DNAs
con-taining point mutations in the DNA-binding domain were
tested for their transforming activities on primary mouse
embryocellsas described previously (23).
Adenovirus recombinants.Adenovirus-SV40 recombinants containing thegeneforTantigen (mutantorwildtype)were
generated by cotransfection ofKpnI-linearized pSKATand XbaIfragment A ofadenovirustype5d1309 (21) (map units
4 to 100) in adenovirus-transformed 293 cells as described
previously (3, 25, 42, 45). Transfectionswerecarriedoutby using a CaPO4 precipitation technique (6). Plaques were
picked, and the virus was grown out and screened for T-antigen expression byimmunofluorescence. Virus stocks giving rise to 50 to 100% T-antigen-positive cells were
usually used for T-antigen production. Recombinant viruses
wereobtained for all replication-negative mutations butone
(185ST).
Preparation ofmutant T antigen. 293 cells were infected
with recombinant adenovirusesatamultiplicity of about 10
PFUpercell. At 20h, the cellswerelysedand Tantigenwas
recoveredfrom the lysate by immunoprecipitation (41) with
PAb416 monoclonal antibody. In the case of the 185ST
mutation, sufficient Tantigen for biochemical analysis was
obtained by transfection of the linearized pSKAT into 293
cells by using the same protocol as that for generating
recombinant adenovirus. The binding of thismutantT
anti-gentoDNA wascomparedwith that ofwild-typeTantigen preparedin the sameway.
DNA-bindingassays. In most experiments, DNA-binding activity wasmeasured aspreviouslydescribed(41), usinga modification of the assay ofMcKay (28). All binding reac-tions were performed at T-antigen excess. DNA-binding activity under replication conditions was measuredby the
method described by Deb and Tegtmeyer (11). When present, competitorDNAwas added from the beginningof
the reaction.
RESULTS
Generation of mutations in the DNA-binding domain. We
generated single-point mutants at regular intervals in the
DNA-bindingdomain of Tantigen. Mutations wereinitially
made atevery5 amino acid residues. Additional mutations
were made in a second round of mutagenesis near sites
which hadan effect on virusreplication. The substitutions werechosenso astomakeconservativechanges.Wherever
possible, the same substitution was made for one amino
acid;however,differentsubstitutionsweresometimes made
whendifferent codonswereinvolvedbecause, for efficiency, wewantedtolimit the mutation in the DNAtoasinglebase
pair. Our aim inmaking the "softest" possible amino acid substitutionswastokeepstructural alterations in theprotein
to a minimum. This, we reasoned, would optimize our
chances ofidentifying residues that make contactwith the GAGGC pentanucleotidesattheorigin.
Mutations were generated in a plasmid containing the entire SV40 genome (pBS-SV40) (23). The mutagenesis protocolwas similartotheonedevisedby Kunkel(22)and isdescribed in detail elsewhere(23).MutantSV40DNAwas
excised from the plasmid and then tested forinfectivity in
monkey cells. Virus replication results for 51 mutants that Virus
Replication
Regions
\ /
A 147-159
on November 10, 2019 by guest
http://jvi.asm.org/
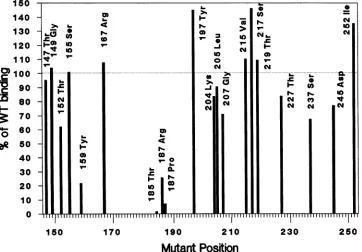
[image:2.612.102.512.73.297.2]150 140 130 120 110
> 100
D 90
M
80 70O 60
be
5040 30 20 10
0
150 170 190 210 230 250
[image:3.612.129.491.82.334.2]Mutant Position
FIG. 2. Binding ofmutantT-antigen proteinstoplasmid DNA. Mutant T antigenswereimmunoprecipitatedwithPAb416and testedfor
binding toasmall(400-base-pair)labeled plasmid fragment(TaqIfragmentDofpSVO+).Resultsareshownas apercentageof wild-type
T-antigen-bindingactivity underidentical conditions (competitor calf thymus DNAwasnotadded); 100%orepresentsabout 10,000cpmof
labeled DNAbound. Resultsarealsoshown in Table 1.
weremade in theDNA-binding domainareshownin Fig. 1.
About 45% of the mutants replicated like wild-type virus, 15% replicatedatless thanwild-typerates asjudged by the
appearanceof smallplaques (S), and the remaining 40% did
notreplicateatall even athigh input DNA concentrations. Figure 1 also shows thatallof the mutantsthatweretested
transformed primary mouse embryo cells. There was no
significant qualitativeorquantitativedifference in the
trans-forming properties of these mutants relative to that of wild-type virus, indicating that gross structural alterations
didnotoccurin themutant Tantigens.
We found it useful togroup the mutants thatshowed an
effectonvirusreplication into three classes correspondingto
regions in the DNA-binding domain, which we term A, B, andC. (Fig. 1). These regions mapped from residue 147to
159, 185 to 229, and 245 to 257, respectively. Only one
replication-negative mutation (at residue 167)waslocalized
betweenregions A and B, and another (at residue 237)was between regions B and C. The residue 237 mutant was
actually unimpaired in DNA binding (see below), which makes the division between regions B and C somewhat clearer. The large central region contained several clusters ofreplication-negative mutants mapping at residues 185to
187, 203to 207, and 215to219.
Generation ofmutant proteins. The mutants that did not
replicate in monkey cells are likely to be the ones with
altered domain function. T antigens harboring these
muta-tions wereprepared for biochemical analysis. For this
pur-pose, corresponding-base-pair mutations were made in an
adenovirus-SV40hybrid virus whichwasusedas an expres-sion vector. Mutant T-antigen proteins were prepared by
immunoprecipitation (41) from infected 293 cells by using PAb416 (18) monoclonalantibody. TheamountsofTantigen ineachsamplewereestimatedby Coomassie blue stainingof
acrylamide gelsfollowedby densitometry. Equalamountsof each Tantigenwereused in bindingassays.
DNA-bindingassaysofmutantTantigens. Wehave previ-ously adopted a rapid and quantitative assay for DNA
binding using T antigen bound to Staphylococcus aureus
(41). This assay was used for comparing the
DNA-binding
activities ofmutantproteinswiththat ofwild-typeT
antigen.
Several different labeled substrate DNAs andtwo different conditions were used. First, to measure binding ofmutant
proteins to any double-stranded DNA, we used a small
(about 400-base-pair) fragment containing plasmid se-quences.Bindingwith thisDNAwasperformed withoutany
added competitor DNA (Fig. 2 and Table 1). All but four
mutantproteins demonstratedbinding activity close to that of wild type. The four mutations mapped at residues 159,
185, and 187 (HistoArg and HistoPro), and the protein had
activitywhichwas25%orless of that of the wildtype
(Table
1 contains acomplete list of amino acid substitutions inthe replication-negative mutants). The four mutant T antigens
areclassifiedas poornonspecific DNAbinders.
Wenext testedthe activities of themutantTantigensfor bindingtoalabeled DNAfragment containing thewild-type SV40DNAorigin region (sites I andII)(Fig. 3A and Band Table 1). The assay was performed in the absence or
presence of unlabeled competitor (1,000-fold excess) calf
thymus DNA. In the first case, both ori- and nonspecific-binding activities were measured, and in the second, only
ori-specificbindingwas measured (34, 41, and unpublished
results) because the calfthymus DNA competes with
non-specificbinding. When the binding of eachmutantprotein to the orifragmentwascompared in the absence andpresence
ofcompetitor DNA, two major differences appeared (Fig.
3A and B and Table 1). The activities of the proteins with mutations at residues 155 and 204 dropped significantly in
U)~ ~ ~ ~ ~ N
10~~~~U
Co~~~~~~~~~~~~~~~~~1
CoN~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~
a~~~~~~P
N-N)
e (0
-1)
-4W) U)
") N N
v- N N 30
...o...N...N...N... N...
U)
NM
Cl* Co~~~~~~~~C
v-~~~~~~~~~~~I
1-
h.~~~~~~~~v
0.--C N
1-v- ~ rr rri WT ~rr 1 FF
on November 10, 2019 by guest
http://jvi.asm.org/
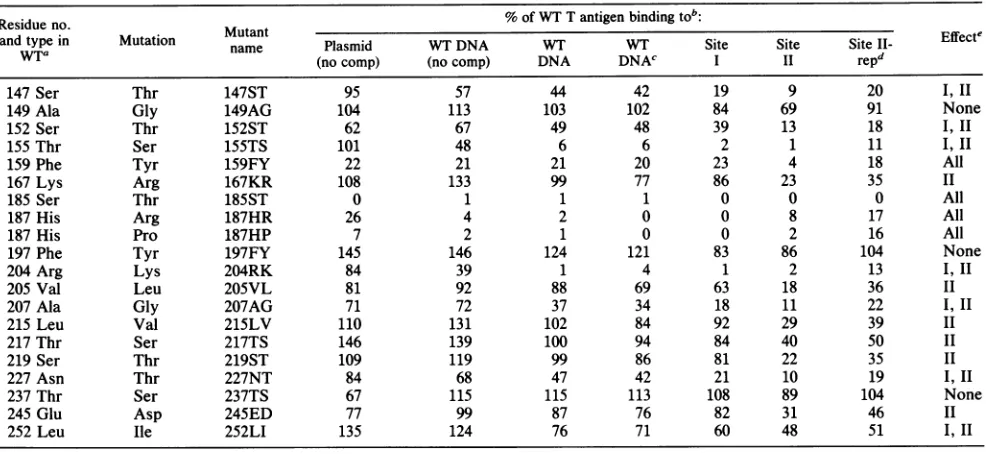
TABLE 1. DNA-bindingresults ofreplication-negativeT-antigenmutants
Residueno. % of WT Tantigen bindingtob:
and
WTaldue
typein Mutation MutantEfetno. utaton name Plasmid WTDNA WT WT Site Site Site l- Effecte
wtra
(nocomp) (nocomp) DNA DNAc I II repd147Ser Thr 147ST 95 57 44 42 19 9 20 I,II
149 Ala Gly 149AG 104 113 103 102 84 69 91 None
152Ser Thr 152ST 62 67 49 48 39 13 18 I, II
155Thr Ser 155TS 101 48 6 6 2 1 11 I,II
159 Phe Tyr 159FY 22 21 21 20 23 4 18 All
167Lys Arg 167KR 108 133 99 77 86 23 35 II
185 Ser Thr 185ST 0 1 1 1 0 0 0 All
187His Arg 187HR 26 4 2 0 0 8 17 All
187His Pro 187HP 7 2 1 0 0 2 16 All
197Phe Tyr 197FY 145 146 124 121 83 86 104 None
204 Arg Lys 204RK 84 39 1 4 1 2 13 1,11
205 Val Leu 205VL 81 92 88 69 63 18 36 II
207 Ala Gly 207AG 71 72 37 34 18 11 22 I,II
215 Leu Val 215LV 110 131 102 84 92 29 39 II
217 Thr Ser 217TS 146 139 100 94 84 40 50 II
219Ser Thr 219ST 109 119 99 86 81 22 35 II
227 Asn Thr 227NT 84 68 47 42 21 10 19 I, II
237Thr Ser 237TS 67 115 115 113 108 89 104 None
245 Glu Asp 245ED 77 99 87 76 82 31 46 II
252 Leu Ile 252LI 135 124 76 71 60 48 51 I, II
aWT, Wildtype.
bMutantTantigenswereisolatedbyimmunoprecipitationfrominfectedor (forThr-185) transfected293 cells. Theproteins werethen tested inDNA-binding
reactionsasdescribedin Materials andMethods. Various substrateDNAs were used in the binding reactions asindicated. Calf thymus DNAwasincluded
(1,000-foldexcess) as acompetitorexcept whereindicated (no comp).
cThis assay with wild-typeDNA wasperformedunder the sameconditionsas the assayswithsite I and site IIDNAs.
dBindingtositeII DNAunderreplication conditions.
Summary of effectson DNAbindingtovarioussubstrate DNAs.I,IImeansthat binding to bothsitesI and II wereaffected,and all means thatbindingto
allsiteswasaffected.Theeffects ofthemutationat site 252 weresmall.
the presence of competitorDNA, demonstrating that these
twosites (Thr-155 and Lys-204 in the wild-type protein) are
important for origin-specificrecognition or for the stability of the ori DNA-protein complex. The binding activity of the
mutant with a change at residue 207 also dropped in the presence of competitor DNA, but the effect was not as
dramatic. Itshould be noted that the activity of the protein
with a mutation at residue 159 did not change from its
original level of 20% (ofthat of wild type) (Fig. 2) when it
wastested with ori DNA either with or without competitor
DNA
(Fig. 3 and Table 1).Binding ofmutant Tantigens to DNA sitesIandII.We next tested thebinding activities ofthe mutantproteins to sepa-rated sites I and II. This is important since, under the
conditions used, most of the binding to the origin is for site I sequences (17, 47, 48; unpublished results). The binding reactions were performed with DNA fragments containing
deletions insites IIandI, respectively(44,46). Thebinding
activities ofthese mutantproteins to wild-type ori (Fig. 4A),
to site I (Fig. 4B), and to site II (Fig. 4C) under identical conditions in the presence of competitor DNA are compared in Fig. 4 and Table 1. Several of the mutant proteins were
placedinonegroup onthebasis that they bound ori DNA at levels somewhat less than that of wild type and on the fact thattheydemonstrated a progressively lower activity as the substratewaschanged from complete ori DNA to site I and then to site II. These are the five mutant proteins with
changes at residues 147, 152, 207, 227, and 252. We can
categorizethese mutants asbeing affected in binding to sites I and II, although not all were affected equally. A second class of mutants demonstrated lowered binding to site II,
mostly. The mutants in this group had changes at residues 167, 205, 215, 217, 219, and 245. Third, there were mutations
(at
residues 155, 159, 185, 187 [Arg and Pro], and 204) whichresulted in low activity with all three substrates used in this
experiment.This category includes the four poor nonspecific DNAbinders(Fig. 2) as well as the two (at residues 155 and 204)which did not bind specifically to sequences at the origin (Fig. 3). In the fourth class, three mutations (at residues 149, 197, and 237) resulted in near-wild-type levels of activity withall three DNAs.
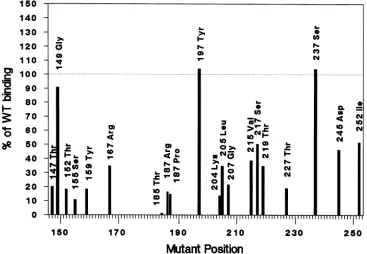
Todemonstrate that the three mutants in the fourth class could bind to site II DNA under conditions that would
permitthereplication of the DNA in vitro, we repeated the binding under replication conditions (5, 11). Under these
conditions, which include ATP and incubation at 37°C,
bindingtosite II is enhanced compared with binding without ATP (5, 11).These three mutant proteins (with mutations at residues 149, 197, and 237, respectively) had wild-type levels
of binding by this criterion (Fig. 5). All other mutant T
antigens bound site II less well. The three mutant proteins with wild-type levels of DNA binding were deficient in helicase activity, which explains the replication defects of the mutant viruses (K. Wun-Kim and D. T. Simmons, unpublished data).
Table 1summarizes the results of all DNA-binding assays. Also shown is asynopsis of the effects of each mutation on thebinding to various substrate DNAs.
DISCUSSION
Theeffect of amino acid substitutions on DNA binding can be due to twodifferent types of changes: achange inprotein conformation or a change in a residue that is involved in function. It has been documented that changes in structure often affect the stability of proteins (31). Thus, for a protein whose structure is not known, it can be generally assumed thatmutations which affect activity without having an effect
on November 10, 2019 by guest
http://jvi.asm.org/
0.
._
I_
Mo
9-150
140 130 120
110
0
100
90
80
70
60 80 40 30 20
10
0
150
140 130 120 110
100
90 80 70 60 80 40 30 20
10
0
150 170 190 210 230
Mutant Position
[image:5.612.127.490.81.537.2]250
FIG. 3. BindingofmutantTantigenstowild-typeSV40 origin. ImmunoprecipitatedmutantTantigensweretestedinabindingassaywith
alabeledfragment containing the wild-type SV40 origin(TaqI fragmentEof pSVO+). Reactionswereperformed in the absence (A) or
presence(B) of1,000-foldmass excessof unlabeled sheared calf thymus DNAas acompetitorofnon-sequence-specificbinding.
on overall stability are infunctional rather than structural
sites(31). Although this isnotahard-and-fast rule,wehave
madeuseof ittodevelopaworkingmodel ofTantigen-DNA
interactions. Our mutantT antigens do not appearto have widely different stabilities, asjudged by the facts that the mutants that were tested gave rise to an equal number of
transformed cellfoci intransformation assays (Fig. 1) and that about equal amounts of T antigen weregenerated by
transfection in 293 cells (not shown). However, we do not want toexclude thepossibility that, insome cases, changes
in DNA-binding activity are due primarily to structural
alterations.
The interpretation of our data also rests in part on the assumptionthat ifseveralneighboring amino acidsarefound
tobe importantforactivity, theregion itself isfunctionally
involved. Ourreasoning is that if theregionisimportantonly
fordetermining the proper structure (i.e., the active site is
elsewhere), anumberof amino acid substitutions would be toleratedand, inouranalysis,mutations resultingin pheno-typic changes would not cluster within it. By making
con-servative amino acid changes, we have attempted to mini-mize signalsduetoconformationalchanges. Thecontention thataregionservesadistinctfunctioncanbe strengthenedif all mutations in the regionlead tosimilar phenotypes, i.e.,
the presence ofone activity but notanother. We have also considered the magnitude ofthe change in DNAbindingto
evaluate thepossibilitythatany one aminoacidispartofan
active site.
I-v-
V0
lNY
0Ncm
I-~~~~~~~~~~~~~~~~~~~~~~~Imc
CM CM
00.
I- N~~~~~~r-VN
I-U)~~~~~~~~r
on November 10, 2019 by guest
http://jvi.asm.org/
18
130
20
100
90
80
70
60
40
30
20
10 0
0)
150
*-
140130
C120
100
90
F
80~
7060
50
_-
40
0
30
20
8
18
150
140
130
120
110
1
80
70
60
50
40
30
20
10
0
I-6
&.U)150
170
190
210
230
250
[image:6.612.130.492.78.569.2]Mutant Position
FIG. 4. Binding ofmutantTantigenstosites I andIIatthereplication origin. ImmunoprecipitatedmutantTantigensweretestedina
bindingassaywithalabeledfragment containing the wild-type SV40 origin (A), site I DNA(B) (TaqI fragment E ofpOS1),orsiteIIDNA (C) (TaqI fragment E of pSVOdl3). All reactionswereperformed witha 1,000-foldexcessofcompetitor DNA. Results arepresentedasa
percentageofwild-typeT-antigen binding under eachcondition.
Theresultsofourexperiments implicate multiple regions
within the DNA-binding domain ofSV40 T antigen in the
efficient binding to various substrates. By the criteria
described above, two regions appear to be of primary importanceinbindingtoDNAatsites I and II ina
sequence-specific fashion. These regions are illustratedby the
DNA-bindingproperties oftheproteins with mutationsmappingat residues155and204 (andtoasmallerextent atresidue 207).
Of alltheproteins tested, thefirsttwomutantsshowed the
greatest percent inhibition of binding to ori DNA when
testedin thepresenceofcompetitorcalfthymusDNA(Fig.
3 and Table1). The boundaries of thesetworegions canbe deduced from the effects ofneighboring mutations on ori binding (Fig. 3 and 4), fromthe pattern ofreplicating and
nonreplicatingmutants(Fig. 1),fromDNA-bindingdata for
T-antigen mutantsdescribed in the literature (2, 24, 34)and from the comparison of amino acid sequences of various
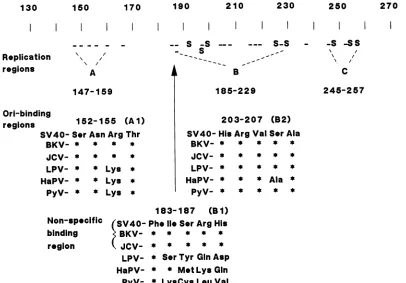
papovavirusTantigens(compiled byJ.Pipas).Thefirstsuch
region, whichwecall Al (Fig. 6), mostlikelyextends from residues 152 to 155 (Ser-Asn-Arg-Thr). This 4-amino-acid
sequence is strongly conserved among all papovavirus T
antigens (Fig. 6).This issignificantbecauseatleastsomeof
,1,* *
I
|*|W|w*||||*|vW|
.1. ... ... ...0 0:00
()-q~j ~~%.0N 1
cm
1~~~4 ~ ~ ~ ~ J>NN N~~~c
cm~~~~~~.
I- (0 o~~~~VmIrY-r
TIIIIIIIIIIII II II ? II II II III II -IIIIII
2- a.
on November 10, 2019 by guest
http://jvi.asm.org/
140 130 120
110
tm
100D 90
}580
g
70O 60
be 50
40 30 20 10
0
150 170 190 210 230 250
Mutant Position
FIG. 5. Binding ofmutantTantigenstositeIIDNAunder replication conditions. ImmunoprecipitatedmutantTantigensweretestedin
abindingassaywithlabeledsiteIIDNAunder replication conditions. All reactionsincluded ATP and competitor calf thymus DNA andwere
performedat37°C.
those proteins (those from SV40, BK virus, JC virus, and polyomavirus)canbindtothesameviralorigin (15, 33, 35), and all probably recognize the same pentanucleotide
se-quenceGAGGC (12, 13, 33). Two of the four residues (152
and155) inthisregion have been mutated inourstudy, and
thesemutantsaredeficient in bindingtosites I andII(Fig. 4 andTable1) butnot toplasmidDNA(Fig. 2).Athird residue (153) has been implicated in DNA binding by Prives et al. (34). Furthermore, this sequence lies withina larger region
(from about residue 144 to 158) identified by Pauchaet al. (32) which appeared to be important in sequence-specific binding. Wecantakeagoodguessatthe limits of this region
because sequence homology among various papovavirus T antigens is considerably weaker immediately downstream
andto someextentupstream ofAl. Second, a mutation at residue 149 hadnoeffectonoribinding (Fig. 3 and 4), anda
mutation at residue 157 had no effect on virus replication
(Fig. 1). It is worthnoting that regionAlcontainsapotential
N-glycosylation site.
Bysimilarreasoning, asecondregion (called B2) appears to be critical for recognition of the origin. This second sequenceextendsfrom residue 203to207(Fig. 6) and is just
asimportant for ori bindingasregionAl. The 5-amino-acid sequenceofregionB2is alsohighly conservedamongall six
papovaviruses (Fig. 6). Sequence similarity drops signifi-cantly outside of this region, and mutations atresidues 202 and 209 hadnoeffect onvirusreplication (Fig. 1). Three of
thefive residues(204, 205, and 207) in B2were mutated in
thisstudy andappearto beimportant forbinding to site II
(205) or to both sites I and II (204and 207) (Fig. 3 and 4,
Table 1). (Subsequent analysis of the residue 205 mutant showed it to be partially defective in binding to site I as
well.) Furthermore, a residue 203 (His to Gln) mutant is
defective inorigin-specific binding (24).
It is important to note that not all amino acids within
regions Al and B2 appear to have equal importance in
binding to the origin. For example, in region B2 of the
wild-type protein, Arg-204 may be much more important
thanAla-207orVal-205, although mutationsatall threesites showaneffect. Certain residueswithin elementsAland B2,
and in particular Thr-155 and Arg-204, may therefore
con-tributemoretothe directcontactwith DNAsequencesatthe
origin. Additional sites are mostlikely involved in making thepropercontactswith theorigin. Saturation mutagenesis
of these regions coupled with the substitution of various aminoacidsateach site shouldprovideuseful information in thisregard.
Athirdregion,termed Bi, is also crucial forthe interac-tion with DNA (Fig. 6). This region, unlike Al and B2, appears tobeimportant in the general recognitionof DNA andnot strictlyinsequence-specific binding. Therearetwo
major reasons for believing that this is the case. First, a
mutation at residue 185 or 187 seriously reduces
DNA-binding activity to all substrates tested, including plasmid
DNA.Second, regionBi (residues 183to187)isidentical in the T antigens of SV40, BK virus, and JC virus, but the
corresponding sequence is strikingly different in the other
papovaviruses except at residue 183 (Fig. 6). This implies
that Bi ispresentin thisregion onlyin the first three virus
proteins. In the otherthree, (B-lymphotropic virus,hamster
virus,andpolyomavirus)Bi iseither absentoritsfunctional
equivalentispresent elsewhere in theDNA-bindingdomain. The limitsof thisregionwereplacedat183 and 187 because mutants with mutations at residues 182 and 189 replicate (Fig. 1)and residue 188 isnotconserved inSV40, BKvirus,
andJC virus. Since themutants withchangesin thisregion
were unable to bind to site I or II in addition to being
defective in nonspecific DNA binding (Fig. 3 and 4), it suggeststhat TantigenmustbindDNAnonspecifically first before itcan efficiently interact with originsequences.
Wehypothesize, therefore,thatthe critical sequencesfor
origin binding makeup amotif of three majorelements and
a-CY
a.
~~~~~~~~
~~~~~~0N
0) C4~~~~~Ua-4. L.0
-.0c00~~~~~~~a
v- C4-*
IIIIrTiiiIIIIIHII
.TT1IfTrI;1TrTIrTrT
TT1rT.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.124.491.76.330.2]130 150 170 190 210 230 250 270
Replication
\\
/,regions A
147-159 Or-binding
regions 152-155 (Al)
SV40- Ser Asn Arg Thr
BKV-* * * *
JCV-* * * *
LPV- * * Lys *
HaPV- * * Lys *
PyV- * * Lys *
--
s
-S --- --- S-S- S
B
185-229
203-207 (B2) SV40- HisArg Val Ser Ala
BKV-* * * * *
JCv-* * * * *
LPV- * * * * *
HaPV- * *
PyV- * *
* Ala *
* * *
183-187 (B1)
Non-specific SV40-PhelIe SerArgHis
binding BKV- * * * * *
region Jcv- * * * * *
[image:8.612.109.515.76.359.2]LPV- * Ser Tyr GinAsp HaPV- * * Met Lys Gln PyV- * LysCys LeuVal
FIG. 6. Sequenceelementsoftheori-bindingmotif. Theori-binding motifisthoughttobecomposedof threesequenceelements. Two of
thethree elements(Aland B2)are necessaryfortheproperrecognitionandbindingtosites I and IIattheorigin.Thethird element(Bi)is important fornonspecific bindingtoDNA. Shownaretheamino acidsequencesthatcorrespondtothesethree elementsforSV40,BK virus (BKV), JCvirus(JCV), lymphotropic papovavirus (LPV),hamsterpapovavirus (HaPV),andpolyomavirus (PyV)Tantigens. Notethatthe central element B1 is presentin thefirst three virus T antigens only. Theequivalentelement in LPV, HaPV, andPyV may bepresent elsewherein theprotein.
that the central element (Bi) is necessary for nonspecific
DNA interactions. An appealing possibility is that some
residues in Bi (as well asother sequences) areinvolved in
making the initial contact withtheDNA. Anchored inpart
through Bi, theDNA slides alonguntilthe GAGGC
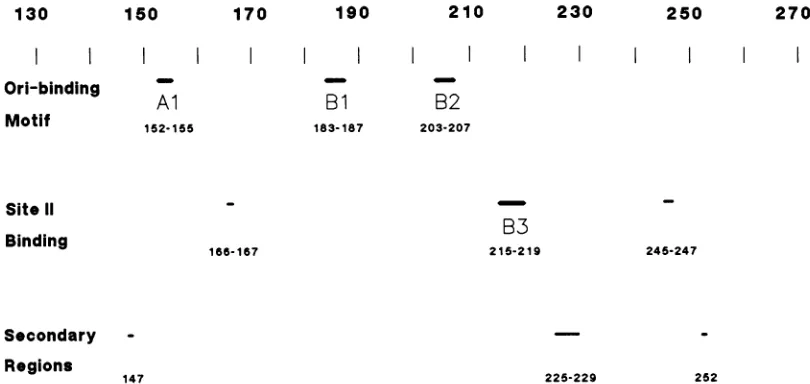
penta-nucleotide sequences appear, at which point elements Al and B2(or certain residues within them) alsomakecontact. Inadditiontothe three motif elements described above,at leastoneregion(region B3 [residues 215to219]) is important
for theproper recognition of site II sequences. Three mu-tantswith mutations within this region show alteredbinding
tositeII DNAonly (Fig. 3). Other potential regions involve sequences surrounding residues 167(166 and 167?) and 245 (245to247?) (Fig. 7).At themoment,it isdifficulttoexplain
howcertainproteinsequencescould be important in binding
to site II but not siteI DNA, since both sitescontain very
similarpentanucleotide sequencesthatarerecognized by T
antigen (12). Perhaps the organization of the
pentanucle-otidesorthegeneralstructureof the DNA itself is important.
Site I contains twoGAGGC pentanucleotide segments
ori-entedtowards theearly region, and siteIIcontains four such
segments, twoorientedone wayandtwoorientedthe other
way(12). Thestructureofthe DNAatthesetwosites could also be a determining factor, since the DNAappears tobe bentatsite1(36)butnot atthe fourpentanucleotides of site
II,although bendingoccursin theATtractadjacenttothem (9). Another difference is the fact that T antigen probably makes additional contacts within the inverted repeat se-quencesin siteII(4), aDNA elementthat is not presentin
siteI.Whatever thereasons,itappearsthatfurther
protein-DNA contactsareimportantfor theproperbindingtositeII, which in turn is required for the initiation ofvirus DNA
replication (29, 38). Our data suggestthat element B3 in T
antigen(Fig. 7)andperhapsother sitesaswell(residues167 and245)areinvolved in theseinteractions.
The fourregions described above appeartobe the most
important for proper recognition and binding to various
originsequences,buttheyarecertainlynottheonly regions
involved. It islikelythatresidues 147, 159, 225 to229,and 252 make some contribution to the proper structure and function of theori-bindingdomain(Fig.7). Unlike the amino acids ineach of the fourmajorelements describedabove, it isnotpossibletopredictwhether theseresiduesare
impor-tantforproperconformationorwhether theyhavea direct
functional role. This is either because the mutations are
isolated or because the magnitude of the effect on DNA
bindingwassmall. On the basis of thephenotypiceffectsof its mutation, residue 159could, like elementBi, be
impor-tantfornonspecificDNAbinding.
Our results may explain the phenotype ofa mutant de-scribed by Margolskee and Nathans (26), which had a
changeatresidue 157(AlatoLeu).Thismutantwasisolated
as asecond-siterevertantofanothermutation inoneof the four GAGGC pentanucleotides in site II. The mutant T
antigendisplayedarelaxedspecificityfor theorigininthat it
recognizedthewild-typeaswellasthealteredorigins. Given
its proximity to element Al, it is conceivable that the mutationatresidue 157 altered the local conformation and
indirectlyaffected the function of this elementorperhaps of
--S -S
\ / \
\2 2 C
245-257
on November 10, 2019 by guest
http://jvi.asm.org/
130 150 170 190 210 230 250 270
Ori-binding Motif
Site II
Binding
147
Bi
Al
152-165
B2
183-187 203-207
B3
215-219
166-167 245-247
Secondary
Regions
225-229 252
FIG. 7. Mapof important regions in the DNA-binding domain. Three categories of elements are shown. At the top is the tripartite motif importanttoorigin binding, inthe middle are three regions that appear to be required for the proper binding to siteII,and at the bottom are severalsecondaryregions whichhave a lesser influence on the activity of the domain.
element B2(iftheyare close together in space). However,
only three-dimensional datacandirectlyaddress this issue.
Our analysis most likely underestimates the number of important sites, since notall positions have been mutated, and thepossibility existsthat, atsomesites wheremutations didnotresult in aphenotypicchange,adifferentamino acid
substitution may show an effect. Furthermore, since our
DNA-binding assays wereperformed atprotein excess and were not under equilibrium conditions, a greater effect on DNAbindingmayhave been observed in some cases. On the whole, however, it seems likely that the overall pattern of
Al 1 52- 155
GAGGC
203-20 183-187
B2 B1
Al 1 52- 155
?_ GAGGCC
215-21 9 203-20 183187
83
SITE
I
SITE
11
FIG. 8. Basic model of DNA interactions. Thismodel describes the involvement of varioussequenceelements inbindingtosites I andIIattheorigin. At siteI,themajor elementsareAl, Bl, andB2. Aland B2interact with the GAGGCpentanucleotides, whereas Bi recognizes a general structural feature of DNA. The arrows on
eitherside of the GAGGC sequence signifythat theorientation of
theDNArelativetotheDNA-bindingdomain is unknown. At site
II,thesamethree elements areinvolved,butatleastone(B3)and possibly two others are also required for binding. The sites of
contactof these latter elementsonthe DNAarenot known.
contributing sites is not very different from that shown in
Fig. 1 as regionsA, B, andC, although thisfiguregives no
indications of the relative importance of each residue to
domainfunction.
The properstructures of the regions directly involved in DNA bindingare ofobvious importance.Thereis evidence
that the structure ofelement Bi is crucial, since a mutant
which maps at residue 186 (Arg to Thr) is
temperature-sensitive (19), implying that a structural change in the
protein whichmakes itincapableof recognizingDNAatthe nonpermissive temperature has occurred. Secondary struc-ture programs based on the methods of Garnieret al. (16)
andNovotny and Auffray (30) predictthat the sequence in
Bi (183to 187)orpartofit formsan-turn, analogoustothe ,-turn in thehelix-turn-helix motifs of certain
DNA-binding
proteins. However,noclearpredictionscanbe made for the
secondary structuresAl and B2, the sequence-specific
ele-ments. Predictions from various programs are compatible
with a p-sheet, a ,-turn, or an extended conformation. Perhapstheonlyconsistencyis the fact thatnonepredictan
a-helixfor either AlorB2.
Ourmajor pointsareillustrated inasimple workingmodel thatshows the involvementof varioussequenceelements in
bindingtosites I and II(Fig. 8). AtsiteI, a
tripartite
motif consisting of elementsAl,
Bi,
and B2 isassociated withthe DNA. Al and B2 make sequence-specificcontactswith the GAGGCpentanucleotides. Since thesetwoelements recog-nize thesameDNA sequences,theycould be closetogetherin three-dimensional space. Region Bl may consist of a
3-turn
andbindstoDNAnonspecifically.AtsiteII,thesamemotifis involved, but inaddition, element B3 and
possibly
others makecontact withsomeunknown DNA sequences. Notshown, butimplied,is the fact thatthesesites ofcontact areonlypossiblein the properstructural
background
of theDNA-binding domain.
ACKNOWLEDGMENTS
WethankMaryAnderson,WilliamYoung,and HemaMallickfor experttechnical assistanceand PeterKisselandJacquesPene for preparing theoligonucleotides. We alsoacknowledge PatHearing
andhis students for helping us set upthe adenovirus expression
/
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.104.509.77.269.2] [image:9.612.67.298.446.636.2]systemand Alan SmithforpointingouttheN-glycosylation sitein
regionAl.
ThisworkwassupportedbyanRSA(CA08466) from the Public HealthServicetoD.T.S. andby Public Health Servicegrants CA 36118, CA 18808,andCA28146 fromtheNationalCancer Institute. This work wasinitiated while D.T.S. was on sabbatical leave at
StonyBrook.
LITERATURE CITED
1. Arthur, A.K., A. Hoss, and E. Fanning. 1988. Expression of simian virus 40 T antigen in Escherichia coli: localization of T-antigen origin DNA-binding domain to within 129 amino acids.J.Virol. 62:1999-2006.
2. Auborn, K., M. Guo, and C. Prives. 1989. Helicase,
DNA-binding, and immunological properties ofreplication-defective
simian virus40mutantTantigens.J.Virol.63:912-918. 3. Berkner,K.L.,and P.A.Sharp. 1983. Generation of adenovirus
bytransfection ofplasmids. Nucleic AcidsRes. 11:6003-6020. 4. Borowiec, J., and J. Hurwitz. 1988. Localized melting and
structuralchangesin theSV40originofreplicationinducedby
T-antigen. EMBO J.7:3149-3158.
5. Borowiec, J.,andJ.Hurwitz.1988.ATPstimulates thebinding
of simian virus 40(SV40)largetumorantigentotheSV40origin
ofreplication. Proc. Natl. Acad. Sci. USA85:64-68.
6. Chen, C., andH. Okayama. 1987. High-efficiency transforma-tion of mammalian cells by plasmid DNA. Mol. Cell. Biol. 7:2745-2752.
7. Clark, R., K. Peden, J. M. Pipas, D. Nathans, and R. Tjian. 1983. Biochemical activities ofT-antigen encoded by simian virus40 A genedeletionmutants. Mol. Cell. Biol.3:220-228. 8. Dean, F. B.,P. Bullock, Y. Murakami, R.Wobble, L.
Weiss-bach, and J. Hurwitz. 1987. Simian virus 40 (SV40) DNA
replication: SV40largeTantigenunwindsDNAcontainingthe SV40 origin of replication. Proc. Natl. Acad. Sci. USA 84: 16-20.
9. Deb, S.,A. L.DeLucia,A.Koff,S.Tsui,and P.Tegtmeyer. 1986. Theadenine-thyminedomain of the simian virus40coreorigin
directsDNAbendingandcoordinately regulatesDNA
replica-tion. Mol. Cell. Biol. 6:4578-4584.
10. Deb, S., S. Tsui, A. Koff,A. L. DeLucia, R. Parsons, and P. Tegtmeyer. 1987. The T-antigen-binding domain of the simian
virus40coreorigin of replication. J. Virol. 61:2143-2149. 11. Deb, S.P.,andP. Tegtmeyer. 1987. ATPenhancesthebinding
of simianvirus40largeTantigentotheorigin ofreplication.J. Virol.61:3649-3654.
12. DeLucia,A.L.,B. A.Lewton,R.Tjian,and P.Tegtmeyer. 1983.
Topography of simian virus 40 A protein-DNA complexes: arrangementofpentanucleotide interactionsitesattheoriginof
replication.J.Virol. 46:143-150.
13. Dilworth, S.,A. Cowie,R.Kamen,and B. Griffin.1984.
DNA-binding activity of polyomavirus large tumor antigen. Proc. Natl. Acad. Sci. USA 81:1941-1945.
14. Dodson,M.,F. B.Dean,P.Bullock, H.Echols,andJ. Hurwitz. 1987. Unwinding of duplex DNA from the SV40 origin of
replicationbyTantigen. Science 238:964-967.
15. Frisque,R.J. 1983.Regulatorysequencesandvirus-cell inter-actionsofJCvirus,p. 41-59.In J.L.Sever andD. L.Madden
(ed.),Polyomavirusesand humanneurological disease.Alan R. Liss, Inc., NewYork.
16.
Garmier,
J.,D.J.Osguthorpe,and B.Robson.1978.Analysisof the accuracy and implication of simplemethodsforpredicting the secondary structure of globular proteins. J. Mol. Biol. 120:97-120.17. Gottlieb,P., M. S. Nasoff, E. F. Fisher, A. M. Walsh, and M. H. Caruthers. 1985. Binding studies of SV40 T-antigen to SV40
bindingsiteII. Nucleic AcidsRes. 13:6621-6634.
18. Harlow,E., L. V. Crawford,D.C.Pim, and N. M. Williamson. 1981.Monoclonalantibodiesspecific for simianvirus 40 tumor
antigen.J. Virol.39:861-869.
19. Hutchinson,N. I., L.-S. Chang, M. M. Pater, N.Bouck, T. E.
Shenk, and G. DiMayorca. 1985. Characterization of a new simian virus40mutant, tsA3900, isolated from deletionmutant tsA1499. J. Virol. 53:814-821.
20. Jones, K. A., and R. Tjian. 1984. Essential contact residues withinSV40 largeTantigen binding sitesI andII identifiedby
alkylation-interference. Cell 36:155-162.
21. Jones, N., and T. Shenk. 1979. Isolation of adenovirus type5 host rangedeletionmutantsdefective for transformation ofrat
embryocells. Cell17:683-689.
22. Kunkel,T. A.1985.Rapidandefficientsite-specificmutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
23. Loeber, G.,R.Parsons,and P.Tegtmeyer.1989. The zincfinger regionof simian virus 40largeTantigen.J. Virol. 63:94-100. 24. Manos,M.M.,and Y. Gluzman. 1985. Genetic andbiochemical
analysis of transformation-competent, replication-defective simianvirus 40largeTantigenmutants.J. Virol. 53:120-127. 25. Mansour,S. L.,T.Grodzicker,andR.Tjian. 1985. An
adeno-virus vector system used to express polyoma virus tumor antigens.Proc. Natl. Acad. Sci. USA 82:1359-1363.
26. Margolskee,R.F.,and D. Nathans.1984. Simian virus 40 mutant Tantigenswithrelaxedspecificityfor the nucleotide sequence atthe viral DNAoriginofreplication. J. Virol. 49:386-393. 27. McCutchan, J. H.,andJ.S.Pagano.1968. Enhancement of the
infectivityof simian vius 40deoxyribonucleicacid with diethy-laminoethyldextran. J. Natl.Cancer Inst.41:351-357. 28. McKay, R. 1981. Bindingof simian virus 40 T antigen-related
protein toDNA.J. Mol. Biol. 145:471-488.
29. Myers, R. M., andR. Tjian. 1980.Construction andanalysisof simian virus40origins defective intumorantigenbinding and DNAreplication. Proc. Natl. Acad. Sci. USA77:6491-6495. 30. Novotny, J., and C. Auffray. 1984. A programforprediction of
protein secondary structure from nucleotide sequence data: application tohistocompatibility antigens. Nucleic Acids Res. 12:243-255.
31. Pakula,A.A.,and R. T.Sauer.1989. Geneticanalysisofprotein stability and function. Annu. Rev.Genet. 23:289-310. 32. Paucha, E.,D.Kalderon,R. W.Harvey,andA. E. Smith. 1986.
Simian virus40originDNA-binding domainonlargeTantigen. J.Virol. 57:50-64.
33. Pomeranz, B., and J. Hassel. 1984. Polyomavirus and simian virus 40 large Tantigens bind to common DNA sequences. J. Virol. 49:925-937.
34. Prives, C., L. Covey, A. Scheller, and Y. Gluzman. 1983. DNA-binding properties of simian virus 40T-antigen mutants defective in viralDNAreplication. Mol. Cell. Biol. 3:1958-1966. 35. Ryder, K., A. L.DeLucia, and P. Tegtmeyer. 1983. Binding of
SV40 A protein to the BK virus origin ofDNA replication. Virology 129:239-245.
36. Ryder, K., S. Silver, A. L. DeLucia, E. Fanning, and P. Tegtmeyer. 1986. An altered DNAconformation in origin region Iisadeterminant for the binding ofSV40largeTantigen.Cell 44:719-725.
37. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
38. Shortle, D., and D. Nathans. 1979.Regulatorymutantsof simian virus 40: constructed mutants with base substitutions at the origin ofDNAreplication. J. Mol. Biol. 131:801-817.
39. Simmons,D. T.1986.DNA-binding region of the simian virus40 tumorantigen. J.Virol. 57:776-785.
40. Simmons, D. T. 1988. Geometry of the simian virus 40 large tumorantigen-DNA complex as probedby protease digestion. Proc. Natl. Acad. Sci. USA85:2086-2090.
41. Simmons, D. T., W. Chou, and K. Rodgers. 1986. Phosphoryla-tiondown regulates the DNA-binding activity ofsimian virus 40 Tantigen. J.Virol. 60:888-894.
42. Solnick,D. 1983.Shuffling adenovirus promoters: a viral recom-binant with early region 1A underlatetranscriptional control. EMBO J. 2:845-851.
43. Stahl, H., P. Droege, and R. Knippers. 1986. DNA helicase activity ofSV40 largetumorantigen. EMBOJ.5:1939-1944. 44. Stillman, B., R. D. Gerard, R. A. Guggenheimer, and Y.
Gluzman. 1985. Tantigenandtemplaterequirements for SV40 DNAreplication invitro. EMBO J. 4:2933-2939.
45. Stow, N. D. 1981.Cloning ofa DNAfragment from the left-hand
on November 10, 2019 by guest
http://jvi.asm.org/
terminus of the adenovirus 2 and its in site-directed mutagenesis. J. Virol. 37:171-180.
46. Strauss, M., P. Argani, I. J. Mohr, and Y. Gluzman. 1987. Studies onthe origin-specific DNA-binding domain of simian
virus 40 large Tantigen. J. Virol. 61:3326-3330.
48. Tjian, R. 1978.Protein-DNA interactionsatthe origin ofsimian virus 40 DNA replication. Cold Spring Harbor Symp. Quant.
Biol. 43:655-662.
48. Tjian, R. 1978. Thebinding site of SV40 DNA foraT antigen-related protein. Cell 13:165-179.
49. Wold, M.S., J. J. Li, and T. J. Kelly. 1987.Initiationofsimian virus 40 DNA replication in vitro: large-tumor-antigen and origin-dependent unwinding of the template. Proc.Natl.Acad. Sci. USA 84:3643-3647.