Copyright ©1967 American SocietyforMicrobiologv Printedin U.S.A.
Amber
Mutants of
Bacteriophages T3 and T7
Defective in
Phage-directed
Deoxyribonucleic
Acid Synthesis
R. HAUSMANN AND BEATRIZ GOMEZ
DivisionofBiology,SouthwestCenter forAdvancedStudies, Dallas, Texas 75230
Received for publication 3 June 1967
Amber mutants of the related phages T3 and T7 were isolated and tested for their ability to restore-as the wild type does-thymidine incorporation in ultra-violet
(UV)-irradiated,
UV-sensitive, nonpermissive host bacteria (Escherichia coliBS-1).
Most amber mutants had thisability. However, in both T3 and T7, mutants unable topromotethymidine incorporation underthese conditionswere found and classified into two well-defined complementation groups: T3DO-A andT3DO-B,
T7DO-Aand T7DO-B.
Infection
ofB,-,
cells with representatives of groups DO-A had the following characteristics: (i) phage-directed uridine uptake inUV-irradiatedcells was reduced to less than
20%
ofnormal; (ii) breakdown of host deoxyribo-nucleic acid(DNA)
was delayed and incomplete; (iii) no serum-blocking antigens appeared; (iv) no cell lysis occurred; (v) the ability to exclude the heterologous wild type was impaired. Amber mutants ofthe DO-B groups, infecting B5 l, were able to: (i) promote an efficient phage-directed uridine uptake in UV-irradiated cells; (ii) bringaboutrapidbreakdown of host DNA; (iii) synthesize serum-block-ingantigens;(iv) lyse
the host cells, generally after the normal latent period; (v) exclude efficiently the heterologous wild type. Although physiological similarities betweentherespective
DO-AmutantsorDO-Bmutantsof T3 and T7 wereevident,no
physiological
cross-complementation occurred,
and genetic crosses gave no evidence ofgenetic
homologies between groups of T3 andT7.Coliphages T3and T7are closelyrelated.
They
are
morphologically
identical(34), they
cross-react
serologically
(1, 19),
and theirdeoxyribonu-cleic
acids(DNA)
canbehybridized efficiently,
by
melting
andannealing
(29).
Geneticrecombi-nation
betweenT3 and T7 has also beenreported
(19).
Notwithstanding
theirsimilarities,
upon simultaneous mixed infection ofEscherichia coliwith
thesetwophages,
mutual exclusion occurs;i.e.,
inmore than90%
of the hostcells,
only T3,
oronly T7,
isproduced (19, 20).
Thereasonsforthis
areunknown,
but so far we have ruled outmutual interference
withadsorption
andinjection
(unpublished data).
Togather
moreinformation
relevant to an
understanding
of the exclusionmechanism,
weinvestigated
moreprecisely
thesimilarities
and differences between T3 andT7,
as far as the functions necessary forphage-di-rected
DNAsynthesis
are concerned. Theap-proach
was to isolate amber mutants of these phages which are defective inphage-directed
DNA
synthesis,
to characterize these mutantsgenetically
andphysiologically,
andtoinvestigate
their
behavior
inregard
to thephenomenon
ofmutual
exclusion.
Anabstract of part of this work has been published (Bacteriol. Proc., p. 154,1967).
MATERIALS AND MEUHODS
Bacteria. Our
permissive
host, used for obtaining alllysates, was E.coliBBw/1l.
Wederived it by two mutational steps from an E. coli BB strain which is permissive for certain amber mutants and was kindlysent to us from R. S. Edgar's laboratory. E. coli
BBw/l
is more suitable as an indicator strain for colormutants(19)
ofT3 andT7 and is resistant to phageT1. Asnonpermissivehosts, either E. coliB/5orE.coli
B,_1
wereused;the formeralsostemsfromR. S. Edgar's laboratory, the latter-an ultraviolet-sensitive strain (30)-is from W. Harm's collection.
E.coliK-12thy- str8 TDNA,amutantin which DNA
synthesis istemperature-sensitive, was isolated byF.
Bonhoeffer (2) andkindly givento usbyH. Schuster.
Phlage.
Thecrigin and characteristics ofour refer-ence types of T3 and T7, here called T3am+ and T7am+,have beenpreviously reported (18).Isolation ofamber mutants.Wild-typelysates grown
on E. coli B/5 were treated with nitrous acid, hy-droxylamine, or 2-aminopurine; in general, pro-cedures as outlined by Edgar and Lielausis
(11)
or 779on November 11, 2019 by guest
http://jvi.asm.org/
HAUSMANN AND GOMEZ
Freese (16) were followed. The mutagen-treated lysates were grown foronecycle inBBw/1 to elimi-nate mutational heterozygotes. Phage from these secondary lysates were allowedtoadsorb for 5to 10 min to concentrated cultures of BBw/l at multi-plicities of less than 101, andwere diluted into indi-cator mixtures containing BBw/I and B/5 in the proportions of1:3 (forT3) or 1 :1 (forT7). Platings weremadeondried color agarplates (4)byuseof the spreading method. Ambermutants wererecognizedby their small and irregular plaques. They were picked and platedonBBw/l. Two orthreerandomplaques
grown onthis hostwerethen retested for the amber character byplatingonBBw/l,onB/5,andonmixed indicator (BBw/l plus B/5). If the mutants showed low levels of leakiness and low reversionfrequencies, high-titer lysates inBBw/l were madefrom asingle
plaque. The letter prefixes A, H, and N were used,
respectively,todesignateambermutantsisolated from lysates treated withaminopurine,hydroxylamine,and nitrousacid.
Spot tests. Complementation studies were carried
out byspot-testingall amber mutants (15). A lysate tobetestedwasdiluted,mixed withaculture ofB/5,
and plated byspreading. Immediatelyafter the agar surface had dried, drops of diluted lysates of other ambermutantswereplacedonthesameplate,which was incubated overnight. The tests were performed
with different concentrations ofphage,ranging from 107 to109particles/ml.Two ambermutants were
con-sideredtobelongtodifferentcomplementationgroups ifonespot inatleastoneconcentration showed
com-pletelysis, while theselfingcontrolsatthesame
con-centrationsclearlyshowednolysis.
Growth conditions. M9medium supplemented with Casamino Acids
(0.5%1c)
was used (30); in crosses, nutrient broth wasused. Unless otherwisestated, all experiments wereperformed at 37 C, withexponen-tiallygrowingbacteria (about3 X 108cells/ml),on a
rotatoryshaker.
Ultraviolet (UV) irradiation. Samples (10 ml) ofa
B.-1
culturewereplaced
in apetri
dish andexposed
under continuousshakingtodoses of 1,200 or 8,000ergs/mm2 ofUVfroma15-wGeneralElectric germi-cidallamp.Thelowerorhigherdoses weregiven, re-spectively, when thymidine or uridine incorporation wasmeasured. The cells wereincubated for about 10 minto bringbacterial DNA (30) or DNAand ribo-nucleic acid (RNA) synthesis to a halt. 3H-labeled
thymidine or uridine, and phage, were then added.
Burstsize and latentperiod of wild-type phages were
notinfluencedby irradiation of the host with the lower UV dose, but the higher dose markedly reduced the
phage-producing capacityof the cells.
Conditionsof infection. Phage was generally added
at amultiplicity ofatleast 10particles per cell. This assuredinfectionofabout
90%0
of thecells by at leastonephage particle within 1 min, andofvirtually all cells after 3 min. This was measured by viable cell
countsinsamples treated with antiphage serum. When mixedinfections withT3 and T7wereperformed, T3
was added at a multiplicity three times higher than T7,tocompensate for the lower adsorption rate of T3.
Thiymidinie
or uridineincorporation. When indicated,1 Muc of 3H-thymidine or 0.05 MAc of 3H-uridine (Schwarz BioResearch Inc., Orangeburg, N.Y.) was added per ml of bacterial culture. As suggested by Boyce and Setlow (3), 250 ,ug (per ml) of deoxyadenosine was added for improved thymidine incorporation. At intervals, 1-ml samples werechilled with equal volumes ofice-cold 10%trichloroacetic acid. The samples were filtered and washed with
5%lo
trichloroacetic acid. The radioactivity retained on the membrane filters wasmeasured in a Beckman scintillation counter. In-corporation of thymidine or uridine into acid-in-soluble materials was considered to be a measure of DNA synthesis or RNA synthesis, respectively (30).
Phage crosses. A sample (0.5 ml) of an exponen-tiallygrowing culture of permissive host bacteria (3 X
108cells/ml) was added to an equal volume of a mix-tureof two different amber mutants (5 X 109 particles of each type, per ml). The phage mixturecontained also 4 mm potassium cyanide. Five minutes was al-lowedforadsorption. The multiplicity of infection was about 10particles of each parental phage. The infected cells werediluted into brothcontainingspecific anti-phage serum (K 2),and 2mm potassium cyanide. Again after 5min, the cells were further diluted, bya
factor of 10-4, and allowed to lyse spontaneously. Platings were made onBBw/l and on B/5. Since the frequency of back-mutants was negligible, phages forming plaques on B/5 were considered to be wild-type recombinants. Their percentage relative to the
counts onBBw/I wasdoubled (to account for double-amberrecombinants), and then plotted on the linkage map.
Other measurements. DNA concentrations were measured by the method of Burton, as described by Epstein et al. (15). Cell densities were checked tur-bidimetrically with a Klett-Summerson colorimeter. Serum-blocking antigen was evaluated by the method of DeMars (6).
RESULTS
Identification of amber mutants defective in
phage-directedDNA synthesis. First, the 200-odd independentlyisolatedamber mutants of T3, and
about 150 ofT7, wereclassified into
complemen-tation groups by spot-testing. Thus, 18 and 20
such groups were individualized, respectively, in T3 and T7.To
identify
thoseblocked
in a function required for phage DNAsynthesis, one or more mutantsof each group were tested for their abil-ity to restore thymidine incorporation upon infec-tion ofUV-irradiated B8-1 cells. [In these cells, no bacterialDNAsynthesis takes place (30), and phage-directedDNAsynthesis canbe clearly dis-tinguished.] For this purpose, 3H-thymidine was added to aB.-,
culture which had beenirradi-atedabout 10minearlier.Samples of thisculture wereinoculatedwithphage and further incubated
for 25 min; radioactivity in the acid-insoluble
fraction was then measured. T7am+ determined
the incorporation of more than twice as much thymidineasdidT3am+ (Table 1). Five of theT3
780 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE 1. Phage-directed incorporation of 3H-thymidine in UV-irradiatedEscherichia
coli
B,_1a
T3 T7
Spot- Thymni- Spot-
Thymi-test Phage ine test Phage dine
group incorpo- group
~~~~incorpo-group ~ ~ group rated
No phage 425 No phage 499
am 3, 229 am+ 7,929
1 amNl9 388 1 anH280 437
2 amH5 432 2 amH30 431
3 amH13 488 3 amH131 514
4 amN22 336 4 amAII 7,566
S amNl04 465 5 amA21 8,970
6 amA47 3,095 6 amA24 6,609
7 amA59 1,188 7 amH5 4,286
8 amA69 1,145 8 amH6 7,252
9 amH4 3,923 9 amH10 7,366
10 amH26 2,127 10 amHl 1 8,567
11 amH34 2,524 11 amH13 3,508
12 amH42 3,486 12 amH20 6,767
13 amH44 5,277 13 amH51 7,322
14 amH63 2,200 14 amH71 6,202
15 amH84 3,682 15 amH80 7,161
16 amN76 1,458 16 amH90 9,272
17 amN90 3,087 17 amHI08 8,823
18
amNl58
3,140 18 amHlll 7,99119 amH127 7,525 20 amN72 6,175
aAn exponentially growing culture of E. coli
Be.1
(3 X108cells/ml) wasgiven 1,200 ergs/mm2of UV and incubated for 5 min, to bring bacterialDNA synthesis to a halt.
3H-Thymidine
(1oc/
ml) was added; immediately afterwards, samples
of the culture were infected with phage
(multi-plicity ofinfection, 10) and incubated at37 C for 25 min.Radioactivity of the fraction insolublein cold 5% trichloroacetic acid was then measured,
and is expressed as counts per minute per milli-liter ofculture.
amber
spot-test groupsand three oftheT7amber groups showed acomplete inability
to promotephage-directed thymidine incorporation.
[Follow-ing
thenomenclature
ofEpstein
et al.(15),
suchmutantswill henceforth bereferredtoasDO mu-tants
(DNA-zero).]
Levels ofincorporation
tovarying degrees
lower than normal were alsofound
with several grouprepresentatives,
butT3amH44
consistently
showedincorporation
levels 50 to
100%
higher
than wild type. Suchabnormal
levels were characteristic of certain whole groups, notonly
ofsomeof itsrepresenta-tive
mutants.Some mutants of T3 and T7 with such
anom-alous patternswere
investigated
inrespect
tothetime course of
thymidine incorporation
afterinfection of UV-irradiated
nonpermissive
bac-teria. T3wild-type
and several amber mutants ofT3 determined a sharp onset of thymidine incor-poration 5 to 6 min after infection, but differ-ent finallevels were reached (Fig.
IA).
The pre-viously identified DO mutants did not promote0 2 0
0
0
c
x 01
11
m
E m
Time afterinfection (Min)
FIG. 1. Time course of thymidine incorporation
after
infection of UV-irradiated Escherichia coli B,_1 with T3am+ or some amber mutantsof T3 (Fig.
JA),
andwith T7am+ or some amber mutants of T7 (Fig. IB). An exponentially growing culture of E. coli B,_1
(3 X 108 cells/ml) was given a UV dose of 1,200 ergs/mm2 and incubated for5 min at37 C. 3H-Thymi-dine (I
puc/ml)
wasadded and theincubation wascon-tinuedfor6mini. Samplesof this cultureweretheen
in-fected with phage (multiplicity of infection, 10). At intervals, J-mlsamples were withdrawnfor measuring
radioactivity in the fraction insoluble in cold
5%
tri-chloroacetic acid. The open circles coverpoints refer-ringtoallcurves. Black arrowsindicatetime of visible lysis ofa culture. The designationsDO-A and DO-B refer tocomplementationgroups,asdefinedinthe text.T7amH23 is one among several "leaky" mutants of group DO-A. Other mutants ofthis group, sUIch as
T7am280, did not promote thymidine incorporation
abovethebackgroundof theuninfected host.
781
P;.
6
-i
c
.E -,
4
Z;
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.460.26.217.62.465.2]782
~~~~HAUSMANN
AND
GOMEZJ.VR.
3 Initiation of
thymidine incorporation
inirradi-T3amH44 ated
B,-1
cells was shown todepend
onphage-A directed
protein synthesis,
asjudged by
the fact2
~~~~~~~~~~~that
it wastotally
inhibitedby
theaddition,
to-2
~~~~~~~~~~gether
with thephage inoculum,
of 30gg
(per
ml)
ofchloramphenicol.
Infection of
E. coi K-12TDNA
by
T3, T7,
andI0
T3am+ their DO-amber mutants. Besidesknocking
outt ~~~~host DNA
synthesis by UV,
anotherprocedure
{3amH13(DO-A) for
distinguishing
T3- or T7-directed DNAsyn-T3amN104(DO
-B)
thesis was used. It consisted ofinfecting
E.coli
0 0 2 N hg K-12
TD~NA,
anonpermissive
strain inwhich DNAE
~
~
~ ~ ~ ~~~~+synthesis
istemperature-sensitive (2),
at atern-perature
of 41 C. Under theseconditions,
DO-- B ~~~~~~T7amH90 amber mutants were not able to
promote
thy-midine
incorporation, although
thewild-type
a3
phages
and other ambermutants wereableto doso
(Fig. 2).
Itisevident, however,
that thisability
C
~~~~~~~~~~was
greatly
reduced in T3am+ ascompared
toi 2 T7am+. Under these
conditions,
the rate ofthy-mnidine
incorporation by
T3am~
wasonly
20%
that ofT7am+,
ascompared
to 50%
when UV-irradiated
B.,-
cellswereusedashosts.Patterns
of complementation
andgenetic
recoin-F7aimH280(DO -A) binationof
DO-ambermutants. Table 2 shows the 7~~~~~~am
H30(DO-B) classification of our DO-mutants intospot-test
0 tO 20 30 Nopae groups. The
aloainof
somemutants toaspe-Time after infection(Min) cific group was ambiguous, and three other
cri-FIG. 2. Time course
of thymidine incorporation
teria were used toclarify
this issue. These wereafter
infection
at 41C,
of
Escherichiaco/i
K-12rDNA
(i)
determination of average burst sizes per cellwith T3am+ or amber mutants
of
T3(Fig.
2A),
and when differentpairs
ofDO-amber mutants were withT7am+orambermutantsofT7(Fig. 2B).Bacteria crossed inthenonpermissive
hostB/5,
(ii)
meas-weregrownat30Ctoaconcentrationofabout5 X107 urements of
thymidine incorporation
undercon-cells/mI,
thenwarmedto41Candfurtherincubatedfor ditions ofmixed infection of UV-irradiatedB,,-iSmmn. Labeledthymidine
(1 pc/ml)
wasadded.Afewcells,
and(iii)
genetic mapping
of DO mutants.minutes later, phage was added. At
intervals,
J-ml Tables 3and4showforT3andT7,
respectively,
samples were withdrawn for measuringradioactivity
the average burst sizespernonpermissive,
andasinthefractioninsolubleincold5%trichloroacetic acid. a
control,
perpermissive
cell,
after mixed infec-Multiplicityof infection wasJOphagespercell,
except tion withtwoDOmutants. Undernonpermnissive
for the curve represented by black triangles (multi- cniin,tebrtszsi rse ftorpplicity
of
40). Blackarrowsindicate visiblelysisof
the cniin,tebrtszsi rse ftorpculture. resentativesof thesame
spot-test
groupweregen-erally
in the range of thebackground
of a selfincorporation
ofthymidine
at any time. After cross; crosses between mutants ofdifferentspot-infection with
T7am+, thymidine incorporation
test groupsyielded
burstsvarying
from 0.2to 80 was initiated after a similarlag period
of 5min;
particles. Bursts in thepermissive
hostranged
however, compared
to T3am+infection,
it pro- from 40to 130particles.
Burstsizes smaller thanceeded at about twice the rate
(Fig.
iB).
Most fiveparticles
pernonpermissive cell posed
theDNA-sythesizing
ambermutants ofT7were like question of whether functionalcomplementation
the wild type in this
respect.
But,
asinT3,
some had occurred ornot,
sincethey
were below the of themincorporated thymidine
at a lower rate burst sizes obtained with otherpairs
of mutantsthan did the wild
type,
although
thisincorpo-
by
afactor of 10ormore.ration started after the normal
lag.
InT7,
spot-
Measurements ofthymidine incorporation
testgroup 1contained,
besidesunambiguous
DO under conditions of mixed infection ofUV-irradi-mutants,
some"leaky"
mutants,
such as T7- atedB.-,
cellswereof furtherhelp
inestablishing
amH23,
which showed a residualthymidine
in- thedegree
of functionalcomplementation
be-corporation (up
to10%
of the normalrate).
The tween DO-amber mutants. Thepoorly
"coin-shape
of all thecurves wasnotaffectedby
chang-
plementing"
combinations ofspot-test
grouping
themultiplicity
ofinfection within the testedrepresentatives
ofT3, i.e.,
combinations withT3-rangeof 3to 30
particles
percell.amN19, T3amH5,
andT3amHl3
on onehand,
782
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
TABLE 2. Allocation ofDO-amber
mutanits
tocomplementationz
groupsPhage Complementation Spot-test Mutant stocks
group group
T3 T31DO-A 1 amNl9ga; amA34(?)'; amN69
2 amH5;amA34(?); amN108 3 amH13; amH79
T3DO-B 4 amN22; amA31; amH84; amN18 5 amHl04; amA31(?); amHlS; amNl8(?)
T7 T7DO-A 1 amH280; amH23; amH31;amH205; amH264; amH283;amH335;
amH358
T7DO-B 2 amH30; amA3; amH70; amH102; amH114; amN71
3
amH131;
amH70(?)
aThefirst mutant in each row is the arbitrarily chosen reference type against which all other mutants
were tested.
IInterrogation marks refer to dubious cases.
TABLE 3. Averagebuirstsizes pernzonipermissive or permissivecellaftermixedinlfectionl with
twoDO-amber mutal?tsofT3
Complementa- Spot-test tiongroup group
DO-A 1
DO-B
2 3
4
S
Phage
amNI9 amN69
amA34
amH5
amH13
amN22
amN18 amA31 amNI04
amH15
DO-A Spot-test
group 1 (amN19)
<0.05
(99)"
0.1 (108) <0.05 (77)0.3 (103)
0.2 (73) 21 (109)
20 (82)
23 (67) 25 (82) 28 (64)
Spot-test group 2 (amH5)
-b
0.8 (91) 0.3 (121) 0.1 (59)
0.5 (70)
27 (61) 25 (46) 38 (74)
29 (113) 36 (58)
Spot-test group 3 (amH13) 0.3 (128) 0.2 (105)
<0.05 (99)
19 (86)
14 (58)
23 (85) 20 (73)
27 (109)
DO-B Spot-test
group 4 (amN22)
27 (73) 18 (78)
<0.05 (128) <0.05 (103) <0.05 (77)
1.7 (60) 0.2 (57)
Spot-test group5
(amN104) 20 (95)
36 (126)
0.6 (66) 0.3 (70) <0.05 (59)
0.1 (93)
a Burst sizes below 0.05 could not be evaluated because of the background of unadsorbed input phageswhich had survived antiserum. Crosses wereperformed in Escherichia coli B/S (nonpermissive)
orE. coli BBw/1 (permissive). Burst sizes were obtained by dividing phage titers after spontaneous lysis by the titer ofplaque-forming units beforelysis. The burst sizes per permissive cell, infected in identical conditions, are given in parentheses.
bSee samecross in another box.
and T3amN22 with T3amNlO4 on the other, brought about no appreciable increase of thy-midine-incorporating ability. In other pairwise combinations, thespot-test grouprepresentatives of T3 restoredthymidine incorporation tovalues 30to 85% of normal (Table 5). In T7, values of
70 to
95%/
were found with representatives ofspot-test group 1 on one side and groups2or3 on the other. Between groups 2 and 3,
comple-mentationwasless than1O%ofnormal(Table 6).
For mapping purposes, crosses between DO mutants were performed in the permissive host BBw/l, and the progeny were platedon permis-sive and on nonpermissive (B/5) indicator bac-teria. Particles growing on B/5 were considered
am+ recombinants. The map distances shown in
Fig.
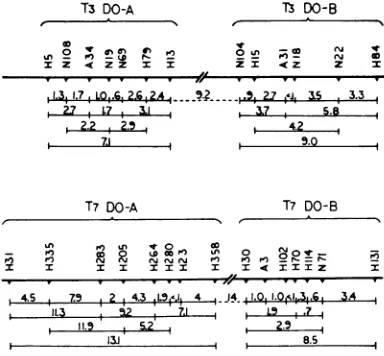
3refer
totwice the percentage ofam+
recom-binants relative to the total progeny, so as to account
for
thereciprocal (double-amber)
recom-binants. In T3,
representatives
of spot-test groups1, 2, and3mappedin the samesegment of theT3
genome,
extending
over 7 map units(1
unit de-fined as 1%c
recombination between twomark-ers);
representatives
of groups 4 and 5mapped
in another segment,
extending
over 9 units. Thedistance between the closest
representatives
ofeach segment was about 9
units,
and no repre-sentative of other spot-test groups have as yet been foundto lie in between. InT7,
a somewhatsimilar situation was found.
Representatives
of 783on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.460.18.422.74.205.2]HAUSMANN AND GOMEZ
TABLE4. Averageburstsizes per nonpermissive orpermissivecellafter infection with two DO-amber mutants ofT7a
DO-A DO-B
Complementation Spot-test Phage
group group Spot-testgroup 1 Spot-testgroup 2 Spot-testgroup3
(amH280) (amH30) (amH131)
DO-A I amH280 <0.05 (77)
-amH23 <0.05 (51) 35 (42) 45 (70)
amH205 0.1 (67) 29 (58) 36 (65)
DO-B 2 amH30 23 (53) <0.05 (54)
amN71 43 (59) 0.1 (45) 4.6 (64)
amHl14 30 (45) 0.2 (38) 3.8 (52)
3 amH131 27 (66) 3.5 (64) 0.2 (50)
aProcedures as in Table3.
TABLE 5. Complementationpatterni between DO-ambermutants of T3asmeasured by3H-thymidinie incorpora-tion in UV-irradiatedEscherichia coliB. la
DO-A DO-B
Group Phage am+
amN19 antH5 amnHl3 amN22 amN104
Wild type am+ 4,385 4,591 4,752 4,372 4,921 4,396
DO-A amNI9 303 261 161 1,570 1,966
amH5 189 343 2,814 3,439
amH13 228 1,464 3,077
amN22 250 187
DO-B amNl04 104
aTheprocedurewas asoutlined inTable I, exceptthat,when twophages were used, the multiplicity ofinfectionwas5of each. The resultsareexpressedascountsperminute per milliliter of culture. When
nophagewasadded,theactivitywas220 countspermin perml.
spottestgroup1
mapped
in onesegment, whereasrepresentatives
of groups 2 and 3mapped
inanother,
separated
from the formerby
14 mapunits
(Fig.
3).
The relativepositions
of these groupsof T3orT7with respecttorepresentatives
of
other spot-test groups have not yet been establishedunambiguously
because of looselinkage
relationships.
The data derived from these
mapping
experi-ments, as well as from burst size measurementsand stimulation of
3H-thymidine incorporation
in
nonpermissive
conditions, led
us to establishtwo
well-defined
complementation
groups whichare necessary for
phage-directed
DNAsynthesis
to occurinboth
phages,
T3andT7. As aworking
hypothesis, thepoorly
"complementing"
spot-testgroupswereconsideredto
belong
tothesamefunctional
unit,
since theirrepresentatives
mapped
at distant sites withinone map segment and the low levels of"complementation" might
be attributable to the formation ofwild-type
recombinants,
rather than to functionalcomple-mentation. The groups established were called T3DO-A and T3DO-B, T7DO-A and T7DO-B (without
implying
genetic homology betweenhomologously
named groups ofT3
and T7). Group T3DO-A comprises spot-test groups 1, 2, and 3, and spot-test groups 4 and 5 constitute group T3DO-B.Groups
T7DO-A and T7DO-Bcomprise
spot-test group 1 and spot-test groups2
and
3, respectively (Table 2).In the following, representative mutants of these groups will be further characterized in re-gard totheir physiology upon infecting nonper-missive cells.
Celllysis andsynthesis of serum-blocking
anti-gens. Allmutantsof groups T3DO-A and T7DO-Awereunabletolyse the nonpermissive host and didnotpromotesynthesis of serum-blocking anti-gens.Allmutantsof groups T3DO-B and
T7DO-B didlyse the nonpermissive host (although with
some mutants, lysiswasdelayed); they promoted synthesis of serum-blocking antigens within the
784 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE 6. Complementation pattern between DO-amber mutantsof T7 as measured by thymidine
incorporation in UV-irradiated Escherichia coli
Bs-1a
DO-A DO-B
Group Phage am+
amH280 amH30 amH131
Wild am- 8,273 8,154 7,052 8,957
type
DO-A amH280 226 5,867 7,823
DO-B amH30 215 776
amH131 263
aProcedureand definitions asoutlined inTable
5. Thebackground activity (no phageadded) was
308.
nonpermissive
cells,
although
in diminishedamounts
compared
tothe wild types.Postinfection
breakdownof
host DNA. Sincemostof the DNA ofT3 andT7 is built up from components
of
thehost
(21),
anextensive break-downof host DNA mustprecede
or accompanyphage-directed
DNAsynthesis.
This breakdown was detectedby
measuring
the amount ofacid-insoluble radioactive
material within labeled hostcells,
atvarious
times after infection withphage.
Nonpermissive
host cellswere grownin t*Wres-ence of
3H-thymidine
andcentrifuged
when theirconcentration reached
3 X 108cells/ml.
Thepellet
wasrinsed,
and thensuspended
inunlabeledmedium
at theoriginal
temperature(37
C)
and cellconcentration.
Theresuspended
culture wasdivided
insamples
which were infected either withwild-type
or witha DOmutantof either T3or T7. The
wild-type
phages brought
about asharp, albeit
limited,
solubilization
ofpreviously
incorporated
thymidine, starting
about 5 minafter infection
(Fig. 4).
Solubilization
leveled offafter
a few moreminutes,
and a slowreincor-poration of solubilized
material tookplace
until the cellslysed.
Weinterpret
these kinetics as acomposite
of twophenomena,
reflecting
thebreakdown
of hostDNA,
whichprovides
apool
of
components for thesubsequent synthesis
ofphage
DNA.The DO-amber mutants showed a different behavior. With
representatives
of groupsT3DO-B and T7DO-B, the onset of solubilization of
previously
incorporated
thymidine
occurred atthe same time as with the wild types, but it leveled off much later.
Only
about 40and15%c,
respectively,
of the host label remainedacid-precipitable
after infectionby
these ambermutants of T3 or
T7;
noreincorporation
ofsolubilized materialoccurred inthesecases. With
representatives ofgroupsT3DO-Aand
T7DO-A,
the onset of solubilizationof the host label wasdelayed and the rate of solubilization was less
than one-third as compared to the homologous representatives of group DO-B (Fig. 4). In all
cases, breakdown of host DNA after phage
in-fection could be prevented by chloramphenicol
(30
4g/ml),
added together with the phage. When similarexperimentsweredone, butwith-outremoving labeledthymidine before infection, the results obtained allowed additional
informa-tionaboutthebehavior of DOmutants (Fig. 5). Again, the representatives of groups T3DO-B
andT7DO-B brought aboutaneasily observable solubilization of previously incorporated thymi-dine aftera lagofabout 5 min. Upto this time,
however, incorporation of thymidine bythe host continued. This continued thymidine incorpora-tion in the (unirradiated) host was more
pro-nounced after infection with the representatives of groups T3DO-A and T7DO-A (in which
breakdown of host DNA was delayed). With
T3amH13,representinggroupDO-A,thereduced
phage-induced breakdown, as observed in the
experiment ofFig. 4, wascompletely masked by
the continuing incorporation of thymidine. We
consider theresidual incorporation ofthymidine by unirradiated
B.1
cells, after infectionbyDO-ambermutants, asrepresenting synthesis of
host-specific DNA. This view was taken considering
that thymidine incoiporation by this
UV-sensi-T3 DO-A
Z-o N
Z 4 ZZ
T3 DO-B
Z <Z Z
,13,1.7,LW,.G, 2.6.2... 9.2 2.7 *J. &5 3.3
27 , L7 11 ,7 5.8
2.2 2.54.2
71 ,- , 13.0
T7 DO-A T7 DO-B
I
Ln ,.) an .410 co
-K PI)I) l 0 N0Kcf) fIonfl Pe)°0 _-=_l_
-I X I :I I I I I I IZ
;7
s4.5 ,7' 2 4.L3 0.1 4II.iS.ViI.vIi5al. lb -.A i
W11.3 9.2 7.1 , L9.3
11.5 5.2 2.5
313 8.5
FIG. 3. Linzkage mapofsomeDO-ambermutantsof
T3 and of T7. Distances are given as twice the
per-centages of plaques on the nonpermissive indicator
relative toplaques ont the permissive one. The values
areaveragesoftwo tofourcrosses.
785
AA
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.460.231.423.415.593.2]HAUSMANN AND GOMEZ
0.05
,uc
of 3H-uridine was added per ml of culture15 No phage
(Fig. 7).
Uridineincorporation
by
the irradiated---
_-T3am+
cells wasresumed2to 3minaftertheaddition ofT3 amH5 wild-type phage and leveled offat about 10 min.
T3am
H13(
Contrary to phage-directed thymidine uptake(which was totally inhibited), the rate of uridine
l0
- incorporation was reduced by only about 30%by the addition of chloramphenicol (30
Aglml).
\\ arn
H122Mutants
ofgroupsT3DO-Band T7DO-BshowedN221 (DO-B) an incorporation rate quite similar to the wild
t^ 5
T3amNI-4J
types,
although
thelevel
ofincorporation
was o somewhat lower (Fig. 7). Uridine uptakedeter-34 B mined by representatives ofgroups T3DO-Aand
T7DO-A, however, was sharply reduced, as
com-,*|,2.| pared to the wild types and other DO-amber
0 10 20 30 mutants. In T7amH23, the most efficiently
.C incorporating representative of group T7DO-A,
E uridine
uptake
was limited to20%,,
that of wild5 No phage type; in
group
T3DO-A,
themaximum
was 5%* t -, ---.-- T7am+ thatofwild type (Fig.7). Inallcases,
appreci-C. %
2 c " I. \ T7amH280
10 T7amH2
~~~~~(DO-A)
3j
14:|\T7stmH2D_ ,2t A
No
phage
12
10
E5
~\z
8_c \ -
to
F \> .
~~~~~~T7am
N717
6-A
(D0-a)
N
T3omH13(D0-A)
H30 T3am+
0 10 20 30 22 -c
Time ofter infection (Min) T3amNIO4(DO-S)
FIG. 4. Breakdown of host DNA after infection O -10 0 10 20 30
with DO mutants ofT3 (Fig. 4A) or T7 (Fig. 4B). v
-Nonpermissive cells were grown in the presenceof N
tritiated thymidineforseveralgenerations, thenwashed z 2 B Nophoge
and suspended in unlabeled medium. A few minutes .
later, phage was addedata multiplicity of10 particles
per cell, andthe cells werefurther incubatedat 37 C.
-Atintervals,1-ml sampleswere addedtoequalamounts of ice-cold 10% trichloroacetic acid. Acid-insoluble 6
-radioactivity
was
plotted.
Black arrows indicate theS
*
T7am+timeofvisible
Iysis.
4--E
X2>^ w
~~~~~~T7
amH280(DO-A)tive host could be totally prevented with a small T7amH30(DO-B)
UV dose prior to infectionby DO-ambermutants. -10 0 tO 20 30
The data referring to thymidine incorporation Time afterinfection(Min)
after phage infectionwere corroborated by chem- FIG. 5. Time course of thymidine incorporation
ical determinations by Burton's method (15) of after infection of exponentially growing Escherichia
net DNA content of phage-infectedcells (Fig. 6). coli B,_1 cellswith T3am+ or some amber mutants of Phage-directed uridine uptake. The procedure T3 (Fig.5A) andwith T7am+ orsome ambermutants for these experiments was similar to those aimed ofT7 (Fig.SB).3H-thymidine(I pc) was addedwhen
at characterizing the DO mutants by their in- the bacteria had reached a concentration of2 X 108
ability
tabil(Fityto
to promotepromote
phage-directed thymidine
phagetdirethe
thymsidine
up-cells/ml.
After 14 min,samples
of
this culture wereup infected with phage (multiplicity of infection, 10).
take (Fig. 1), except that the UV-sensitive host At intervals, J-mlsamples withdrawnfor
measur-cells were given a higher dose of UV (8,000 ing radioactivity in the acid-insoluble fraction. Black
ergs/mm2)
and, instead of1,u c of3H-thymidine, arrowsindicatethetimeofvisiblelysis.786
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.460.48.236.65.400.2] [image:8.460.250.444.290.542.2]i
4-z
a
No phage
T3amH13(DO-A)
T3 am+
T3omN104(DO-B)
Nophage
T7amH280(0O-A)
TaOm+
+
T7amH30(DO-B)-10 0 10 20 30 40
Time after intection(Min)
FIG.6. Net DNA content ofculturesofEscherichia coli B_,, at various timesafterinfectionwith thewild
type or some amber mutants ofT3 (Fig. 6A) or T7 (Fig. 6B). Samplesof anexponentiallygrowingculture wereinfected withdifferent mutant strains of T3 or T7 at amultiplicity ofabout5phagespercellandfurther
incubated (37 C). At intervals, 8-ml samples were
chilled with 2ml of
70/cl
perchloric acid at 0 C, and their DNA content was evaluated by the method of Burton. Black arrowsindicate the time ofvisiblelysis ofaninfectedculture.able effect of
multiplicity of infection
wasde-tected.
Lack
of heterologous
complementation
between DO-amber mutants. Inprevious experiments
wehad
observed
thatsomeambermutantsof
T3and T7 werecapable of
cross-complementing
each
other,
at least to some extent, inspite
oftheir
mutualexclusion
from the progenyof
amixedly
infected
cell(to
bepublished).
Inheterologous
crossesbetween DO-ambermutants
of
T3andT7,
using
anonpermissive
host, we found no suchcross-complementation
in anycombination of T3and T7 mutants. Absence of
cross-complementa-tion
wasalsofoundatthe levelof DNAsynthesis
by
measuring
thymidine
incorporation
afterpair-wise
infection of
irradiatedrestrictive
cells withone T3 mutant and one T7 mutant
(Table
7).
Mutual exclusion betweenT3 and77,asaffected
by
DO-ambermutations. To test howDO-ambermutations
affect the exclusion ability (19, 20) ofthe corresponding
genomes, representatives of the groups DO-A and DO-B of T3 and of T7 were crossed with the heterologous wild types in the nonpermissive host. It was reasoned that mixed in-fection should be abortive in the fraction of cells in which the wild type was excluded by heterologousamber
mutant.(This
holds true only in the absenceu
-i
L-E E~
2
0 0
i,
-5 0 5 10 IS 20 Time after infection(Mini
FIG. 7. Time course ofphage-directed uridine
in-corporationafier infectionof UV-irradiated Escherichia
coli B,-, with T3am+ or some amber mutants ofT3 (Fig. 7A), and with T7am+ orsomeambermutantsof
T7 (Fig. 7B). An exponentiallycrowing culture (3 X 108cells/ml) wasgivena UV dose of 8,000
ergs/mm2
and incubated for 10 mini at 37 C. 3H-uridine (0.05,uc/ml) wasaddedandtheincubation was continued for
8 min. Samples of thisculture were theninfectedwith phage (multiplicity ofinfection, 10). At intervals,
1-ml
sampleswere withdrawnfor measuringradioactivityin
thefraction insoluble in cold
5%lc
trichloroacetic acid.on November 11, 2019 by guest
http://jvi.asm.org/
[image:9.460.33.222.67.334.2] [image:9.460.239.430.164.533.2]HAUSMANN AND GOMEZ
TABLE7. Lack of heterologolas complementation between DO-amnber muiltanlits of T3 and T7,asm1easuiredby
thymidine intcorporationzin UV-irrtadiatedEscherictia coliB,_l
DO-A DO-B
Group Phage Blank T7 asnl
T7amH280 T7amiH3() T7amzIH131
Blank 270 7,934 336 206 216
Wild type T3 am+ 4,686 5,935 4,260 1,513 1,685
DO-A T3 amNI9 205 8,707 288 303 241
T3 amH5 301 8,819 271 350 245
T3 anmHl3 128 7,324 240 216 191
DO-B T3 amnN22 305 4,978 185 266 218
T3 atmNl04 112 3,582 212 258 241
aTheprocedure wasasoutlined in
Table
1,
exceptthatT3 wasadded at amultiplicity of15, andT7at amultiplicityof5,tocompensate fordifferences in adsorptionrate. Under theseconditions, the off-springofaculture infected withT3anm+ andT7
anm+
wasabout50%c
ofeachtype.ofheterologous complementation. Platingson
per-missiveindicator afterlysisof themixedlyinfected cells indeed revealed the absence of the amber parental typeintheprogeny.) Onthe otherhand, if the function impaired by an amber mutation was essential to the exclusion mechanism, the growthofawild-type phagewould notbeaffected
by the presence of such a heterologous amber
mutant. Tables 8 and 9 show data pertinent to this type ofcross.The relativeinput multiplicities
of the parental types were so chosen (20) as to
allow about 80 to 90c% of the mixedly infected
cellstoexclude theheterologous wild type, ifthe same cross was performed as a control in the
permissive host. The mixedly infected cells were
plated before lysis, and the wild type-producing infectivecenterswerecounted. (On both
permis-sive and nonpermissive indicators, similar
num-bersof wildtype-producinginfectivecenterswere
counted.) Control experiments, in which cells were infected by the wild type only and plated similarly, indicated the number of wild
type-producing cells in the absence ofexclusion. The difference between the plaque counts in these
experiments indicated the number of cells in which the wild type had been excluded bythe
co-infecting heterologous ambermutant.
Representa-tivesofgroupsDO-Binbothphagesexcludedthe
heterologous wild-type phage nearly to the same extent inthe nonpermissiveand inthepermissive
hosts. The exclusioncapacityofrepresentatives of
groups DO-A, however, wasseverely impaired in the nonpermissive host.
Acorresponding observation was made atthe level of phage-directed thymidine incorporation
after mixed infection ofnonpermissive cells with
DO-amber mutants and heterologous wild type:
representatives of the DO-A groups were incap-able of appreciably reducing the amount of thymidine incorporated by co-infecting heter-ologous wild-type phage (Table7).This indicates that no exclusion of wild-type functions had occurred. On the other hand, representatives of the DO-B groups, upon mixed
infection
in theTABLE 8. Ability of T3 to exchude T7 am+, as affectedbyDO-ainbermutationst
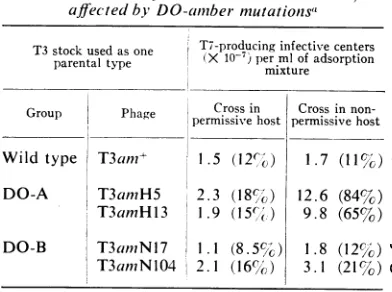
T3 stock used as one T7-producing infective centers parentaltype permixturemlofadsorption
Group Phage Crossin Crossin
non-Group Phage permissivehost permissive host Wild type T3am?i+ 1.5 (12,o) 1.7 (11%) DO-A T3amH5 2.3 (18c) 12.6 (84%) T3amH13 1.9
(15cl()
9.8 (65%) DO-B T3amNil7 1.1 (8. 5 ) 1.8 (12%)T3amNl04 2.1 (16%',) 3.1 (21%)
aHeterologous crosses, with samples of the
samemixtures of parental phages,weredoneboth
in Escherichia coli BBw/l (permissive host) and
E. coliB/5 (nonpermissivehost).Multiplicitiesof
infectionwereabout30 for T3and about 3 for T7.
The infected bacteria were plated on B/5 before lysis. Theinfective centers producingT7am+ were
counted and compared to thenumberofinfective
centers produced incontrol experiments inwhich the cells had been infected with T7amn+ only. The concentration of infective centers per ml of ad-sorption mixture in the control experiments was 1.3 X 108 for B/5 and 1.5 X 108 forBBw/l; these
are the
100%,V6
values, to which the percentages inparentheses refer.
788 J. VIROI,.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:10.460.242.438.366.512.2]AMBER MUTANTS OF PHAGES T3 AND T7
TABLE 9. Ability of T7 to exclude T3am+, as affected by DO-amber mutationsa
T7stock usedasone T3-producinginfectivecenters parentaltype (X
10-1)
permixturemlof adsorptionGroupGroup PhagePhage
~permnissive
erCrossinhost permissiveCross innon-hostWild type T7am+ 2.2 (17%) 3.0 (20%)
DO-A T7amH23 2.5 (19%) 10.8 (72%)
T7amH280 1.3 (10%) 8.9 (59%)
DO-B T7amH30 3.0 (23%) 4.5 (30%) T7amH131 1.8 (14%) 3.6 (24%)
aMultiplicities of infection were about 6 for T3and about 20 for T7. See alsolegendtoTable 8.
same
condition, depressed thymidine
incorpora-tion
by the heterologous
wildtypeby
40to50%
(T3am and
T7am+)
orby
about70%
(T7am
andT3am+).
DIscussioN
It
is
mostlikely
thatreinitiation of
thymidine
incorporation
5 to 6 min after infection ofUV-irradiated
E. coli B8 l cultures with T3 or T7is
due
tothesynthesis
of
phage-specific
DNA, since
Swenson
and
Setlow(30)
haveshown,
and wehave
confirmed,
that in that strain bacterial DNAsynthesis,
asmeasured
by thymidine
incorpora-tion,
stopsshortly
after UV irradiation andcan-not be
resumed.
It could beargued,
however,
that
the
infecting
phage
genomeintroduces
genetic
information for
arepair
mechanismwhich
would
allowhost DNAsynthesis
to berestored.
Thereis
evidence that
no such mechanismexists
in
either
T3 orT7,
since the UVsensitivity
of thesephages
isconsiderably
higher
whenplated
onE.coli
B8.1
ascompared
tothatonE.coli B.If these
phages
hadanefficientrepair
mechanism
of their
own[as
inT4,
for instance(17)],
survival
of
irradiated
phage
shouldbelargely
independent
of
thecharacteristics
of the host.Besides,
sinceanearly
normal
burst ofphage particles
resultsfrom
infection ofB,,l
cells irradiated with1,200
ergs/mm2
ofUV,
phage-specific
DNAsynthesis
must
obviously
occur.Thus, although
moredirect
evidence,
such ashybridization
data withphage-DNA,
is stilllacking,
weinterpret
thymi-dine
incorporation
after infection ofUV-irradi-ated E.coli
B%-
cells asreflecting exclusively
thesynthesis
ofphage-specific
DNA. In any event,however, the lack of
thymidine
incorporation
after
infection
with DO-ambermutants ofT3 or T7canbeattributed
toamber-determined
blocksof specific
genefunctions related
to DNAsyn-thesis.
These
functions
were also required forphage-directed
thymidine incorporation
at 41 C, uponinfection
of E. coli K-12TLNA,
a bacterial strainwhich
has been shown to have atemperature-sensitive DNA-synthesizing
machinery (2). Under theseconditions the wild
types of T3 and T7determined
anefficient
resumption
of thymidineincorporation,
followed
bycell
lysis
and phageproduction, whereas
DO mutants weretotally
inactive.
These results again argue against arepair function of the
DO genes.The
similarity of behavior of
T3 and T7 with respect tophage-directed
DNA synthesis was somewhatunexpected
inview of
theobservations
of Cox and
Yanofsky
(personal
communication)
that
T3clearly
induces
anincrease
in DNApolymerase
activity after infection, whereas
T7does
not,and
that T7 issusceptible
to themutagenic
effects
of
agenetically altered
DNApolymerase
of the host, and
T3is
not. But thefact that
inboth
phages, T3 and
T7, geneticfunctions
havehere
beenidentified
which are necessaryfor
phage-directed
DNA synthesis is not incontradiction
withthe
possibility
of hostDNA
polymerases
using
one orboth of
these genomes astemplates, since phage-specific
func-tions
might be
necessaryfor
making
the phage genomesavailable
to the host enzyme. In fact,only
thesingle-stranded
DNAof
the small phages
bX174 and S13 is known
tobe used
directly astemplate
for
some DNAsynthesis
by hostenzymes
(7,
31). (This
results
inthe formation
ofthe
replicative
form
of these
phages.)
Inphage
X,three
well-defined cistrons (genes N,
0,
and
P)are
required
for the initiation
of phage-specific
DNA-synthesis
(9), besides
afourth
recentlyidentified
genetic
factor
(14). Radding
(26)showed that
the N geneinfluences the
formation
of
theX-specific exonuclease,
although it is
notthe
structural
gene for this enzyme(27). Nothing
is known about thefunctions
of
genes 0and
P,but
it has beenshown
thatgenes N,0,
and Pdo
notinterfere
with theformation of
acircular
X DNAspecies from
the linearinfecting
DNA(28). InT4,
12genes necessary forphage-specific
DNAsynthesis
havebeen
reported
(12);
thefunctions
of3of these genes have been identified
(8,
32, 33). Of 18complementation
groups in T3, asdetermined
by
spot tests, 5 turned out to betotally
deficient inphage-directed
DNAsynthesis;
of
20 such groups inT7,
3 showed thischarac-teristic.
However, upontesting pairwise
combina-tions ofgrouprepresentatives
ofT3 or T7, only two clear-cutcomplementation
groups could befound
inthesephages by
the criterionof
restora-tion of
thymidine incorporation
inUV-irradiated
VOL. 1, 1967
on November 11, 2019 by guest
http://jvi.asm.org/
HAUSMANN AND GOMEZ
B8.1
cells. Burst-size measurements ofmixedly
infected
nonpermissive cells,
mapping
experi-ments, and physiological data supported this latter
finding.
This confirmed the observation of Edgar et al. (10) that spottests are morereliable when negative, since the formation ofintra-cistronic
wild-type
recombinants can besuffi-ciently frequent
tobring
aboutacomplete
clearing
of the spot. We thus divided ourDOmutantsof T3,aswellasthoseofT7,
intotwocomplementa-tion
groups, DO-A and DO-B. Thepossibility
cannot be
excluded, however,
that these groupsconsist of
morethanonecistron.Polarity
effects,
asdescribed by
Nakata and Stahl(23) for T4,
could
lower the level ofcomplementation
betweenadjacent
cistrons.The
exactrole of the
DOfunctions
in thesephages
is notknown,
but somephysiological
features
of
the infectious processby
these DO mutantsdeserve
some comment. DO-B mutantshad the
ability
toinitiate breakdown of the
host DNAafter the
same timeperiod
asfound
uponwild-type
infection,
i.e.,
about
5min,
at 37 C.Within
10min, DO-B mutants of T3solubilized
60%,and
thoseof
T7 upto90%, of
the 3H-thy-midinepreviously incorporated by
thehost
(Fig.
4). In contrast, DO-A mutants werefound
toinitiate
breakdown
later andat amuch lowerrate.In
both
cases,however, this breakdown
wasdependent
onprotein
synthesis
after
infection,
since it did
not occurin the
presenceof
chloram-phenicol.
This is notevidence,
however, for
thesynthesis of
phage-specific
nucleasesafter
infec-tion, since the
phage-induced
intracellular
alter-ations
might
have theeffect of
causing
the
bacterial
DNAtobecomeaccessible
tothealready
existing
host nucleases. Infact,
Pardee andWilliams
(25) reported
that noappreciable
in-crease in
deoxyribonuclease
activity
occurs afterT3
infection,
contrary to what isobserved with
the T-even
phages
and with X(24, 35).
A
strikingly distinctive difference
between DO-A and DO-Bmutants is the fact thatphage-directed uridine
uptake
(which
wasobservable
2 minafter infection ofheavily
UV-irradiatedB.,1
cells)
was not verymarkedly reduced
in DO-Bmutants; in DO-Amutants, only a maximum of 20% of the level of
incorporation
relative to the wild type wasfound. WithsomeDO-Amutants,incorporation
couldhardly bedistinguished
from thebackground
of theuninfectedirradiated
host.Wehavenotyetworked outfurther details about the absence of
phage-directed
uridine uptake byDO-A amber mutants of T3 or T7, but present ideas about the physiological processes of phage infection suggest that it reflects
interference
ofthe amber lesion with
synthesis
ofphage-specific messenger RNA. This could occur by severalmechanisms. For
example, sequential
transcrip-tion ofoperon-like
structures could be prevented by an amber-determined lack ofsynthesis
of aspecific
inducer. A more directinhibitory
action of the amber mutation could beenvisaged
ifoneconsiders
the still controversialinterdependence
oftranscription
andtranslation(5, 13).
It could also bethought
that DO-A ambermutations
affect the synthesis ofaprotein which is necessary forcompleting the injection of the phage genome, in analogy to what hasbeenshown
for phage T5 (22). However,this probably does
not occur in the presentcase, since phage-directed uridine
incorporation
by the wild
typeswasonlyslightly
affectedby
the presence ofchloramphenicol,
denoting efficient
injection
in presence of thedrug.
Anotherphysiological
distinction
between
different
typesof DO
mutantsis
thefact that,
in both T3 and T7, DO-A mutantsdo
notlysethe
nonpermissive
cellsand do
notpromotesynthesis
of
serum-blocking antigens,
whereas DO-Bmutants lyse these cells and
synthesize
serum-blocking antigens. Thus,
DO-B mutants show a behaviorquite different
from what isknown
for T4 (15) and X(9),
inwhich so-called
"latefunctions,"
like celllysis
andsynthesis
ofserum-blocking
power, are notexpressed
if DNAsyn-thesis
isgenetically impaired.
Since
T3 andT7 arerelated
phages,
andtheir
DO-Aand
DO-B mutants,respectively,
havequite similar
physiological characteristics,
thequestion
of the genetic homology between
these mutant groupsof
T3 andof
T7 arises. Geneticcrosses have not
been
informative
because noDO-amber
mutations
can as yet becrossedinto
a
heterologous
genome.Since functional
cross-complementation
has alsonotbeen
observed,
no indication of genetic homology between thesephysiologically similar
functions is as yet avail-able. Wehave, however, isolated
severalhybrids
among amber mutants (inaddition
to DOmutants)
of T3 andT7am+which
aregenetically
compatible
withT7(no exclusion)
but carrythe
T3amber site.
Thus, the question of genetic homology will be further pursued. Whethernon-complementation
is due to the fact that the heterologous geneproducts cannot be utilized by thephage-producing
machinery of either T3 or T7, or whether the mechanism of mutualexclu-sion prevents heterologous gene products from
being formed,
cannotyetbedecided.In regard to the mechanism of mutual
exclu-sion,
itseemsquite
clear that the DO-Afunction is required, whereas the influence of the DO-B function ofT3 orofT7is minimal or nonexistent. Thus, mutual exclusion could be thought of as arelatively early
event, in spite of the fact thatexpression
of the "early"sam+
gene in T3 (18)790
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
AMBER MUTANTS OF PHAGES T3 AND T7
does not seem to be specifically inhibited by the exclusion mechanism. Exclusion might occur at a stage of phage development between the syn-thesis of "early"proteins and the onset of phage-directed DNA synthesis. Further work will be oriented towards evaluating the importance to the exclusion mechanism of other genetic func-tions, whose expression might be affected by nonexpression of the DO-A function.
ACKNOWLEDGMENTS
Weexpressour appreciation to B.Moodyand K. LaRuefortechnical assistance.
Thisinvestigationwassupported byPublic Health Service grants 1 P01 GM13234 OIAI GMP and
1 RO GM14024-01 GEN from the National Institute of General Medical Sciences.
LITERATURE CITED
1. ADAMS,M.H.,ANDE. WADE.1954.Classification
of bacterial viruses: The relationship of two Serratia phages to coli-dysentery phages T3, T7, and D44. J. Bacteriol. 68:320-325. 2. BONHOEFFER, F. 1966. DNA transfer and DNA
synthesis during bacterial conjugation. Z. Vererbungslehre 98:141-149.
3. BOYCE, R.P.,ANDR. B.SETLOW. 1962.Asimple
method of increasing the incorporation of
thymidine into the deoxyribonucleic acid of Escherichia coli. Biochim. Biophys. Acta 61:618-620.
4. BRESCH, C. 1952. Unterscheidung verschiedener
Bakteriophagentypen durch
Farbindikator-Nahrboden. Zentr. Bakteriol. Parasitenk. Abt. I Orig. 159:47.
5. BYRNE,R., I. G. LEVIN, H. A.BLADEN, AND M.
W.NIRENBERG. 1964.Thein vitro formation of aDNA-ribosome complex. Proc. Natl. Acad. Sci. U.S. 52:140-148.
6. DEMARs, R. I. 1955. The production of
phage-relatedmaterials when bacteriophage develop-ment is interrupted by proflavin. Virology
1:83-99.
7. DENHARDT, D. T., AND R. L. SINSHEIMER. 1965. The process of infection with bacteriophage
cIX174.
IV. Replication of the viral DNA insynchronized infection. J. Mol. Biol. 12:647-662.
8. DE WAARD,A.,A. V. PAUL,AND I. R. LEHMAN. 1965. Thestructural gene
for
deoxyribonucleicacidpolymerasein bacteriophagesT4 and T5. Proc. Natl. Acad. Sci. U.S. 54:1241-1248. 9. DOVE,W.F. 1966.Action of the lambda
chromo-some I. Control of functions late in bacterio-phagedevelopment. J. Mol.Biol. 19:187-201. 10. EDGAR, R. S., G. H. DENHARDT, AND R. H.
EPSTEIN. 1964. A comparative study of condi-tional lethalmutations ofbacteriophage T4D.
Genetics49:635-648.
11. EDGAR,R. S.,AND I. LIELAUSIS. 1964.
Tempera-ture-sensitive mutants of bacteriophage T4D: Their isolation and genetic characterization.
Genetics 49:649-662.
12. EDGAR,R. S.,AND W. B.WOOD. 1966.
Morpho-genesis of bacteriophage T4 in extracts of
mutant-infected cells. Proc. Natl. Acad. Sci.
U.S. 55:498-505.
13. EDLIN,G., AND 0. MAAL0E. 1966. Synthesis and breakdownofmessengerRNA withoutprotein
synthesis. J. Mol. Biol. 15:428-434.
14. EISEN, H. A., C. R. FUERST, L. SIMLNOVITCH,R.,
THOMAS, L. LAMBERT, L. PEREIRA DA SILVA,
AND F. JACOB. 1966. Genetics and physiology of defective lysogeny in K12 (X): Studies of
earlymutants. Virology30:224-241.
15. EPSTEIN, R. H., A. BOLLE, C. M. STEINBERG, E.
KELLENBERGER, E. BoY DE LATOUR, R. CHE-VALLEY,R.S.EDGAR,M. SUSMAN,G.H. DEN-HARDT, AND A. LIELAUSIS. 1963. Physiological
studies of conditional lethal mutants of bac-teriophage T4D. Cold Spring Harbor Symp.
Quant. Biol. 28:375-394.
16. FREESE, E. 1963. Induced andspontaneous
muta-tions in bacteriophage, p. 3-18. In W. J.
Burdette [ed.], Methodology in basic genetics. Holden-Day, Inc., San Francisco.
17. HARM, W. 1963. Mutants of phage T4 with
in-creased sensitivity to ultraviolet. Virology 19:66-71.
18. HAUSMANN, R. 1967.Synthesis ofan
S-adenosyl-methionine-cleeMving
enzyme in T3-infectedEscherichia coli and its disturbance by co-infection with enzymatically- incompetent
bacteriophage.J.Virol. 1:57-63.
19. HAUSMANN, R. L., E. P. ALMEIDA MAGALHAES,
AND C. ARAUJO. 1961. Isolation and charac-terization of hybrids between bacteriophages T3 and T7. Anais Microbiol. Univ. Brasil 9:511-526.
20. HAUSMANN, R. L., E. P. ALMEIDA MAGALHAES,
AND C. ARAUJO. 1962. Intracellular
compati-bility of coliphages T3, T7 and Cro. Anais
Microbiol. Univ. Brasil 10:43-62.
21. LABAW, L.W. 1951.Theorigin of phosphorus in Escherichia coli bacteriophages. J. Bacteriol. 62:169-173.
22. LANNI, Y. T. 1965. DNA transfer from phageT5
to host cells: dependence on intercurrent
pro-tein synthesis. Proc. Natl. Acad. Sci. U.S. 53:969-973.
23. NAKATA, A., AND F. W. STAHL. 1967. Further evidence for polarity mutations in bacterio-phage T4. Genetics 55:585-590.
24. OLESON, A. E., AND J. F. KOERNER. 1964. A
deoxyribonuclease induced by infection with
bacteriophage T2. J. Biol. Chem. 239:2935-2943.
25. PARDEE, A. B., AND I. WILLIAMS. 1952. The
in-crease in
desoxyribonuclease
of virus-infectedE. coli. Arch. Biochem. Biophys. 40:222-223.
26. RADDING,C. M. 1964.Nucleaseactivityin defec-tive lysogens of phage X. Biochem. Biophys. Res. Commun. 15:8-12.
27. RADDING, C. M., J. SZPIRER, AND R. THOMAS. 1967. The structural gene for X exonuclease.
Proc. Natl. Acad. Sci. U.S.57:277-283.
28. SALZMAN, L. A., AND A. WEISSBACH. 1966.
Cir-cular DNA formation by lambda mutants.
Virology30:597-581.
29. SCHILDKRAUT, C. L., K. L. WIERZKOWSKI, J.,
791
VOL. 1, 1967on November 11, 2019 by guest
http://jvi.asm.org/
HAUSMANN AND GOMEZ
MARMUR, D. M. GREEN, AND P. DoTy. 1962.
A study ofthebasesequencehomologyamong
the T-series ofbacteriophages. Virology 18:43-55.
30. SWENSON, P. A.,ANDR. B.SETLOW. 1966. Effects
of ultraviolet radiation on macromolecular
synthesis in Escherichia coli. J. Mol. Biol.
15:201-219.
31. TESSMAN, E. S. 1966. Mutants ofbacteriophage S13 blocked in infectious DNA synthesis. J.
Mol. Biol. 17:218-236.
32. WIBERG,J.S. 1966. MutantsofbacteriophageT4
unable to cause breakdown of host DNA. Proc. Natl. Acad. Sci. U.S. 55:614-621.
33. WIBERG, J. S., M. L. DIRKSEN, R. H. EPSTEIN, S. E. LURIA, AND J. M. BUCHANAN. 1962. Early enzyme synthesis and its control in E. coliinfected withsomeambermutantsof
bac-teriophage T4. Proc. Natl. Acad. Sci. U.S. 48:293-302.
34. WILLIAMS, R. C., AND D. FRASER. 1953. Mor-phology of the seven T-bacteriophages. J. Bacteriol. 66:458-464.
35. WORMSER, E. H., AND A. B. PARDEE. 1957.
Deoxyribonuclease activity in Escherichia coli K12 (X) after bacteriophage induction with ultravioletirradiation.Virology 3:76-83.