0022-538X/80/10-0353/08$02.00/0
Intracellular
Forms
of
Simian Virus 40
Nucleoprotein
Complexes
III. Study
of Histone
Modifications
M. COCA-PRADOS, G. VIDALI,'AND M.-T. HSU* TheRockefellerUniversity, New York, New York10021
Themodification patterns of histones present in variousfonrns of intracellular
simian virus 40 nucleoprotein complexes were analyzed by acetic
acid-urea-polyacrylamide gelelectrophoresis. The results showed thatdifferent viral nu-cleoproteincomplexes contain different histonepatterns. Simian virus 40
chro-matin, which contains the activities for the synthesis of viral RNA and DNA,
exhibits a histone modification pattern similar to that ofthe host chromatin.
However, virion assembly internediates and mature virions contain highly mod-ified histones. Pulse-chase experiments with
[3H]lysine
showed that the newlyincorporated histones in the virionassembly
internediates
were alreadyhighly
modified. The majority of in vivo acetylation activity of histones occurred on the
70S simian virus 40 chromatin as analyzed by pulse-labeling with [3H]acetate.
These results and our previous analysis of the virion assembly pathway suggest that three stages are involved in the packaging of simian virus 40 chromatin into
thematurevirion: (i) modification of histones, (ii) accumulation of capsid protein
around the chromatin with highly modified histones, and (iii) organization of capsid proteins into salt-resistant shells. The role of histone modification in virion
assembly isdiscussed.
Insimianvirus 40 (SV40)andpolyoma virus,
viral DNAsareassociated with histones derived
from the host cells (6). The histones in the
mature virion, however, show both qualitative
andquantitative differences from thehistonesin
thehost chromatin. Besides the absence ofHi
histones,the histonesin mature SV40 and
pol-yoma virus were found to be acetylated to a
much higher degree than the cellular histones
(2-4, 9, 11, 17). Recently, the histones in the
intracellular viral nucleoprotein complexes (NPC)werealsofoundtobehighlymodified (2, 3).However,theviral NPCused in these studies
were obtained byextraction from the infected
cellswith Tritonbuffer,which wehave shown
to causedisruptionofmaturevirions and virion
assembly
intermediates.Since virions and virionassemblyintermediates accumulate late in infec-tion,the 70S NPC studied by previous
investi-gatorsare probably
composed
ofboth thegen-uine70S viral chromatin andthe disrupted
vir-ions. Thus, it is rather unclear whether the
highly modified histone patterns observedin the
70S NPC represent the true70S viral
chroma-tins.
In our earlier report we developed a
gentle
extraction method for
isolating
intactintracel-lular formsof viral NPC(7).Three formsof viral
NPC could be obtained by this extraction
method:(i)maturevirus(V), (ii)virion
assembly
intermediate (NPII), and (iii) the 70S viral chro-matin (NPI) which is the precursor to NPII complex and contains the major activities for synthesizing viral DNA and RNA. The purpose
ofthepresentcommunication was to
reinvesti-gate the histone modification patterns in the
various forms of intracellular SV40 NPC
ex-tracted by the gentle procedure. The results
showed that the histones in mature virions and
virionassembly intermediates (NPII) were
mod-ifiedto amuchhigherextentthanthose ofSV40
and cellular chromatins. Furthermore, we
per-formedpulse-labeling experiments and showed
that extensive modification of histones in virion assemblyintermediatesoccurred before the
ac-cumulation of capsid proteins around SV40
chromatin.
MATERIALS AND METHODS
Cells and growth of SV40 virus. SV40 virus strain SVS was grown on CV-1 cells as described
previously(7).
Radioactive pulse-labeling of SV40-infected cells.Allpulse-labelingexperimentswereperformed
in a 370C room. Forpulse-labeling with [3H]lysine,
cells were first incubated for 10 min in lysine-free medium(GIBCOLaboratories)supplementedwith2%
dialyzedfetal calfserum. Themediumwasthen re-placed by a medium containing 200 to 400 !LCi of
[3H]lysine (60to80 Ci/mmol;Amersham Corp.)per ml for the period desired. At the end of the pulse 353
on November 10, 2019 by guest
http://jvi.asm.org/
354 COCA-PRADOS, VIDALI, AND HSU
period, thepetri dishwasimmediatelyplacedonice,
and theradioactive mediumwasreplaced byice-cold Tris-buffered saline. For chase experiments with
ly-sine, theradioactive mediumwaswithdrawn,and the cells were washedoncewith10mlof medium contain-ing15mgofunlabeledlysine-HClpermland 2% fetal calf serum and overlaid with thesamemedium for the chaseperioddesired.
Cellswerepulse-labeledwith 5 mCi of[3H]acetate
(2to 5Ci/mmol;AmershamCorp.)perml for 10or30 min.Procedures forterminating pulse-labeling follow
thosedescribed for[3H]lysinepulse-labeling. Extraction ofintracellular SV40 NPC. SV40 NPCwereextractedfrom the nuclei ofSV40-infected cells,asdescribedpreviously(7).Briefly,infected cells werewashed withTris-bufferedsaline andlysedwith 0.5%Nonidet P-40. The nucleiweresuspendedinTD buffer (25 mM Tris-hydrochloride, pH 7.4, 0.136 M NaCl,7mMKC1,0.7mMNa2HPO4)andhomogenized
in a tight-fitting Dounce homogenizerfor about 30 strokes. Cell nuclei were pelleted by centrifugation
(1,000xg),and theextract waslayeredon a5 to40%
sucrosegradient containing10mMTris,pH7.4-0.5%
phenylmethylsulfonyl fluoride and spun in anSW40
rotorat37,000 rpmfor70min.
Gelelectrophoresisanalysisof histone. Discon-tinuous sodiumdodecylsulfate-gelelectrophoresiswas
performed bythemethod of Laemmli(10). Polyacryl-amidegels,12%(13by22cmby1.5mm),wereusedto
analyze histones present inSV40NPC. Forstudying
the histonemodificationpattern, the acid-urea-poly-acrylamide gel electrophoresissystemasmodifiedby
Shmatchenko andVarshavsky wasused (15). SV40 NPCweredissolved in9Murea-3%
mercaptoethanol-0.9Maceticacid-0.5%cetyltrimethylammonium bro-mideandelectrophoresedina15%polyacrylanidegel
(13by33 cmby1.5mm)at 17V for34hat4°C.Gels
werefixed in 50%trichloroacetic acidovernightat4°C
and stained with0.1%Coomassieblue in50% trichlo-roacetic acid. Destaining was achieved by repeated washings withasolutioncontaining5%methanol and 7% aceticacid. Gelsweresplicedinto 1-mmslices for radioactivecounting.
RESULTS
Electrophoretic patterns of histones in SV40 NPC. SV40 NPC were isolated from
SV40-infectedCV-1 cells at48hpostinfectionas
described previously(7). Theextractwasfurther
fractionated in a 5 to 40%sucrose gradient into three subspecies of viral NPC: (i) SV40 chro-matin (70S), (ii) replication intermediates (1OOS), and (iii) virion andvirion assembly
in-termediates(150 to210S). The histones present
in these various fractions of intracellular NPC
were analyzed by acetic
acid-urea-polyacryl-amide gel electrophoresis as described by
Pan-yimnandChalkley(12). This gel system is
partic-ularly useful in analyzing histone modification
patterns in which modified species migrate as
slower bands (8, 12). Such analysis reveals a
rather distinctive modification pattem of
his-tonespresentinSV40chromatinand in mature
virus purifiedfromCsClgradients (Fig. 1Band
C). The histonespresentinSV40chromatinare
lessmodified and resemble cellularhistone
pro-file(Fig. 1A and B), whereaspurifiedviruslacks
0 0
4-0_
I..
x
a-0
A H20
H2b
It
ItI
ItI
-1
H 1 i
1~~~I
11
a4Ho
SHE
I
II IIH2
liii
1 II"
I,II 1
I
lIl
I1 1'
B III 1l1i1i
Il It I
6~ ~~i II
3-~
~
I t li2
0I
tlO 10 20 30 40 50 60 7O
[image:2.499.268.439.145.596.2]Fraction number
FIG. 1. Acid-urea-polyacrylamide slab gel
electro-phoresis analysis of cellular histones (A, scan of stained gel)andof
[3H]lysine-labeled
histonespres-entinSV40chromatin(B)andCsClgradient-purified SV40 virions (C). For labeled samples, gels wre
slicedinto 1-mm slices and treated with 0.8 ml of Protosolasdescribed in the text before counting in scintillationfluid.
on November 10, 2019 by guest
http://jvi.asm.org/
355
theHisubfractionand contains highly modified
histones. The most prominent differences
be-tweenthe histones in mature virus and those in viral chromatin are the presence of highly
ace-tylatedforms of histone H4 subfractions and the
relative reduction of the H2a-H2b peak in
ma-ture virus (Fig. 10).The reduction of the H2a-H2b peak in mature virus is not due to the
deficiency of these histones in mature virus,
sincesodiumdodecyl sulfate-polyacrylamide gel
analysis failedto reveal anyquantitative
differ-ences of these histones between mature virus
and viral chromatin (unpublished data). Thus,
theseresults indicatethathistones H2a or H2b
inSV40virion, or both, are modified to a higher extentthanthose in viral orcellularchromatins.
Virion assemblyintermediatesisolated from
su-crosegradientscontain thesame histonepattern
asthat inmature virus except that histone Hi
is present in these complexes. The extensive
modification pattern of virion assembly
inter-mediates as compared with that of viral
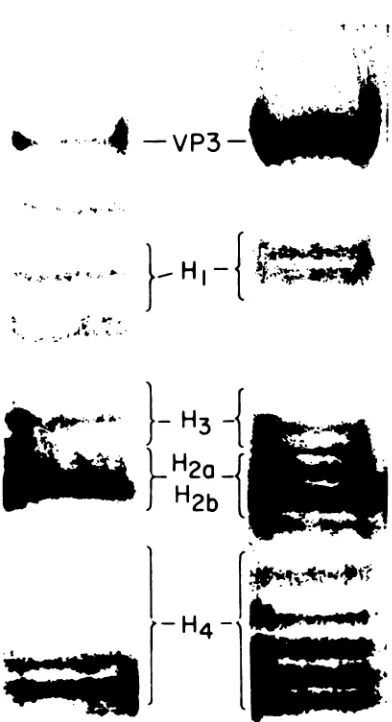
chro-matincanbeseenin Fig.2.
Thehigh level of histone acetylation in SV40
virion and virion assembly intermediates
rela-tive tothe viral chromatin wasalso confirmed
by the analysis of thecontent ofE-acetyllysine
inthesecomplexes. Thecontentofe-acetyllysine
inSV40 virions and virion assembly
intermedi-atesis aboutfive times higher than that in SV40
chromatin (Table 1).
Thus, theseresults demonstrate that thereare
twohistone modificationpatterns in
intracellu-larSV40 NPC: (i) aless modifiedpattern
asso-ciated with viral chromatin which is involvedin
thebiosynthesisof viral RNA and DNA(7),and (ii)anextensivelymodifiedpatterninthe virions
and virionassemblyintermediates.
Electrophoretic pattern ofnewly incor-porated histones in SV40NPC. The results
described above show that the histones inSV40
chromatin and virion
assembly
intermediatesaremodifiedtodifferentextents.
Previously
wehave shown that virionassemblyintermediates
arederived from SV40 chromatin
(4,
7). Thus,
the conversion ofSV40 chromatin into virion
assembly intermediates is associated with the
modificationof histones.
However,
it isnotclearwhetherthechangein histonemodification
pat-tem occursearly duringtheassemblyorduring
thematurationofvirions. Toanalyzethis
prob-lem we pulse-labeled SV40-infected CV-1 cells at 48hpostinfectionwith[3H]lysinefor 90 min.
During thisrelativelyshortpulse
period,
labeledhistones wereobserved onlyin
early
virionas-semblyintermediates(4),asmostof thelabeled
protein couldbedisassociated fromSV40DNA
in theCsClgradient.The histonespresentinthe
.'
-VP3-}_HI
s*# ..4 s.
I f P- .
... ,i4-. .
>-
H3
}
H20
H2b
L
H4-.t~~~~~~~~~~~~~~~~
S_
-I
_0
*ta
1'
Ie
FIG. 2. Acid-urea-polyacrylamide gel
electropho-resis of histonemodificationpatternsin SV40 chro-matin (left column) and SV40 virions and virion precursors (right column) isolatedfrom a sucrose
gradientasdescribedpreviously (6).
pulse-labeled SV40chromatin and virion
assem-blyintermediateswere
analyzed
inacid-ureagel.
The results are shown in Fig. 3. The labeled
histones which just become detectable in the
newlyformed virion
assembly
intermediate(150
to 180S in the sucrose gradient) are already
extensively modifiedas inthe maturevirus. In
a separate
experimental
protocol,
the histonesinSV40chromatinwerebriefly pulsedandthen
chased into virionassemblyintermediates. Dur-ing a short pulse (2 to 5 min) labeled histones
were observed in the positions of SV40
chro-matin andreplicationinternediates(5),butno
radioactivityin the positionofvirionassembly
internediates couldbedetected(Fig.4AandB).
During a brief pulse for 2 miin followed
by
achase witha 100-foldexcess ofunlabeled
lysine
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.256.452.68.429.2]TABLE 1. e-Acetyllysinecontentin SV40virion,
virionassemblyintermediates,and viralchromatin'
H3, H2., H2b, e-Acetyl- Lysineas Complex H4(input lysine e-acetyl-cpm) (cpm) lysine
708 chromatin 1.70x 105 2,310 1.36 Assembly interme- 3.84x104 2,737 7.12
diates
Virion 1.98 x104 1,203 6.08
aSV40nucleoproteincomplexeswerelabeled for24
h with[3H]lysineandfractionated in thesucrose gra-dientasdescribedpreviously (7). Analysisof
e-acetyl-lysinewasperformed asdescribedby La Bella et al. (9).
for1h,labeledhistonesbegantobe observed in
the virion assembly intermediates (complete
conversion ofnewlyformedSV40 chromatin into
virionassembly intermediaterequiresatleast4
h of chase, depending on the period of viral
infection). Again, the newly incorporated
his-tones presentin thevirionassembly
intermedi-ates were highly modified as compared with
SV40chromatin (Fig.
4C,
and C3) which issim-ilartothesteady-state pattern shown inFig.1.
The pulse-chase experiment also revealed a
subtle change of histone pattern in the newly
synthesized SV40 chromatin. Histone H4 is
more highly acetylated in the briefly pulsed
chromatin than in the chased steady-state
la-beled chromatin (Fig. 4A1-A3). On the other
hand, histone H3 is less modified in thenewly
synthesizedchromatin than inthe chased
chro-matin. These results are similar to those
ob-served incellular chromatin(8).
Analysisof rate of acetylation of histones
in SV40 NPC. The results described above
showthatnewlyincorporated histones inyoung
virionsarealready highly modified. These data,
however, donottelluswhen themodification of
histones in virion assembly intermediates
oc-curs. Modification could occurin afraction of
SV40 chromatin before the accumulation of
cap-sid proteins onto these modified chromatins.
Alternatively,
modification of histones couldhappen after the fornation of virion assembly
intermediates. To distinguish these possibilities
we studied the kinetics of one of the major
histone modifications, acetylation, by
pulse-la-beling with
[3H]acetate
(apulse-labeling studyofhistone phosphorylation was notcarriedout
becauseofthe longequilibration time of the
3P-labeled pool). A short pulse with
[3H]acetate
enablesone tomeasure the rate of acetylation
without concerning theaccumulation of
acety-latedhistones. If the majority of histone
acety-lationoccurs and accumulates in the virion
as-sembly intermediates, then a short pulse with
cY
x
E
0a
u
[image:4.499.57.251.82.185.2]10 20 30 40
FIG. 3. Acid-urea-polyacrylamide slab gel
analy-sisofhistonespresent inSV40NPCpulse-labeledin vivo with 0.2mCiof[3HJlysinepermlfor90min. The viral NCP wereextracted,fractionatedina sucrose
gradient, andanalyzedin a15%
acid-urea-polyacryl-amide slabgel.(a)SV40virion precursorsedimenting
between 180 and210S; (b) 70SSV40chromatin.
[3H]acetate should label the complex. On the
other hand, ifacetylation of histone occurs in
the 70Schromatin and thenaccumulates in the
virion assembly intermediates, then only 70S
chromatin will be preferentially labeled during
ashortpulsewith[3H]acetate. When
SV40-in-fected CV-1 cells were pulsed briefly with [3H]acetate for10 min,themajorityof histone
acetylationwasobserved in the70SSV40
chro-matin fraction instead ofinthe virionassembly
intermediates (Fig. 5).Alongerlabelingperiod
results in the accumulation of label in virion
assembly intermediates (unpublished data).
Theseresultssuggestthatthe hyperacetylation
of histones in the SV40 virion probably takes
place in SV40 chromatin before the
accumula-tion of capsid proteins. The distribution of
[3H]acetateinthepulse-labeled histonesinSV40
chromatinwasalso analyzedin
acid-urea-poly-acrylamide gel. The result isshown in Fig. 6. As
expected, histone Hi is not labeled. Since
Hi
on November 10, 2019 by guest
http://jvi.asm.org/
0
'1~~~
t14{rIiii B
iIilSi1X
B10 20 304 0203 0 tO 2 04
6
-i
li 1I ilIBzB3 o haedfo hafera5-inpus with iiifcodyine (C IC3,adtevrlNCwrII
2-~~~~~~~~~~~10
2
II~~~~~~~~~~~2C
FI.w.Pus-caeanalysis of hitn mdfcainprfls fS40NCiacid-urea-plarlmdgelaecie slabntetx.A ftesmls.Sere4ru
simultaneouslywith the addedcellularhistone markers inparallellanes. Thesecellularhistonestandards are shown on top of eachcolumnfor theidentification of individual histone species. Gels were sliced into 1-mmslices and treated with 0.8 ml of Protosol before counting inscintillation fluid.
hasanacetylgroup atits amino terminiadded
during its synthesis in the cytoplasm (13), this
result indicates that themajority oflabels
ob-served in Fig. 5 and 6 are due to the internal
lysine acetylation. The data inFig.5and6show
that histones H3, H4, and H2a in 70S chromatin
arethemajorhistonespeciesacetylatedduring
ashortpulse.
DISCUSSION
Wehavepresented evidencethatthe histone
modification patterns in SV40 chromatin are
strikingly different from those ofSV40 virion
andvirionassemblyintennediates. Whereasthe
histonespresent in thelatter NCPareunusually
highly modified, the SV40 chromatin contains
essentially unmodifiedhistones. Insomeofthe
previous
reports (2, 3), histonespresent in the70S SV40 chromatin extracted by the Triton
methodwerefound tobehighlymodified. The
discrepancy between the present data and the previousreportismostlikelyduetothe
contam-ination of 70SSV40 chromatin by the disrupted
virions or virion assembly intermediates when
SV40 complexes were prepared by Triton
ex-traction.Disruptionof virionsduringTriton
ex-tractionwasshownbyourlaboratorypreviously
(7). Thus, the presentresults show that there
are twodistinct histonemodificationpatterns in the intracellular SV40 NCP during lytic
infec-tion of CV-1 cells, an essentially unmodified
histone pattern in thebulkofSV40 chromatin
and a very highly modified pattem in SV40
virionsand virionassembly internediates.
Since we have shown in the previousreports
(4,7) thatSV40virion assembly intermediates and mature virions are derived from the 70S
SV40 chromatin, the differencesin the histone
on November 10, 2019 by guest
http://jvi.asm.org/
358 COCA-PRADOS, VIDALI, AND HSU
a
b
c
H.
-I--hi
ll
[image:6.499.140.382.78.435.2]H2a
FIG. 5. Autoradiographofhistones inSV40NPCpulse-labeledat 48hpostinfectionfor10min in vivo with
2.5mCiof[3HJacetateper ml.Pulse-labeledproteinsinSV40 NPCwereanalyzedina12%sodiumdodecyl
sulfate-polyacrylamide gelandfluorographed for1monthbythe methodofBonner andLaskey (1). (a) 70S
SV40chromatin; (b) 100Sreplicative intermediate; (c)180to210S virion and virion precursors. Theprotein ontopofeach lanerepresentsanunknownproteinspeciesthatcomigrateswithviralcapsid proteinVP-I.
modificationpatternsbetween thesecomplexes suggest thatchangesof histonemodification
pat-ternsaccompanythetransitionfromSV40 chro-matintovirionassembly intermediates. Our
re-sults with [3H]lysine pulse and chase suggest
that thechanges in histonepatternsoccurearly
during thegenesis of SV40virion.Pulse-labeling experiments with [3H]acetate indicatethatthere
is no major acetylation activity of histones in
thevirionassembly intermediate. Instead,most
of the acetylation activity wasdetected in the
70S SV40 chromatin fraction. This result
sug-gests that the hyperacetylation of histones in virionassembly intermediatesoccursbefore the
accumulation of capsid proteins. Although we
cannot address the question of when histone
phosphorylation mightoccurbecause of
experi-mental difficulties, ourdataof thesteady-state
32P-labeling patternof histones in thedifferent
fractions ofSV40 NCP do not show a
signifi-cantlyhigher degree ofphosphorylation of
his-tonein virionascomparedwithSV40 chromatin (unpublisheddata). These results andour
pre-vious analysis of the SV40 virion maturation
pathway indicate thattransformation of SV40
chromatin into mature virus involves at least
three stages in the following order: (i)
hyper-modification of histones, (ii) accumulation of
capsid proteins, and (iii) organization of capsid
proteinsinto asalt-resistant shell.
The biological function of the unusually
ex-tensive modification of histones in the SV40
on November 10, 2019 by guest
http://jvi.asm.org/
AND HISTONES
H20 Hi H3 H2b H4
I I 1 a.. .
N
0
x
E
10 20 30 40
FIG. 6. Acid-ureagel electrophoresis of
[HJace-tate-pulse-labeled histones in SV40 NPC isolated fromasucrosegradient. Histonesuerepulsed for 10 min with 2.5 mCiof[3HJacetateperml andanalyzed in an acid-urea gel as described in the text. (a)
Assemblyintermediates; (b) 70S SV40chromatin.
virion isunknown. Thepresenceofveryhighly
modified histonesinSV40 virionand virion
as-sembly intermediates butnotinSV40 chromatin
suggests thatmodification of histones is
neces-saryfor theefficient interactionbetween
nucleo-somes and capsid proteins for packaging SV40
DNA-histonecomplexes. Thismaybe the
mech-anism by which the pool of SV40 chromatin
destined for metabolic processes (synthesis of
RNA and DNA) and the pool of chromatin for
the production ofprogenyviruscan be
distin-guished. Thesetwopoolsofchromatinare
pre-sumably equilibrated by the action of histone
modification anddemodification enzymes.The
hypothesisthathypermodification of histones is
necessary for the efficient packaging of SV40 chromatin issupported by the following
obser-vations: (i)nucleosome invirionassembly
inter-mediates remained associated withcapsid struc-turesafter micrococcal nucleasedigestion,
sug-gesting strong interaction between the highly
modified nucleosome and capsid proteins; (ii)
preliminary results of in vitro assembly using
native 70S SV40 chromatin and the 70S
mini-chromosome derived frommaturevirus showed that the latter ismoreefficient inpartial
reas-semblywithcapsids(Coca-PradosandHsu,
un-published data); (iii) host range mutants ofa
relatedvirus, polyoma,aredeficient in modified
histones (14), and the efficiency of virus produc-tion is also suppressed, in spite of apparently normal synthesis of capsid proteins (16). Further
studies are necessary tocharacterize the role of
histone modifications in the interaction between
theSV40 chromatin and capsid proteins for the
formation of SV40 virion.
ACKNOWLEDGMENTS
We thank M.Wilsonand E. M.Johnson for helpful discus-sions and J. Cozzitorto and H.-Y. Yuforexcellenttechnical
asistance.
This work was supported byPublicHealthServicegrant CA-19073 from the National CancerInstitute to M.-T.H. and
bygrantPCM 7907104 from the National Science Foundation toG.V.M.C-P. is a Fellow of the Spanish NationalCouncilfor Research.
ADDENDUM
After thecompletionof thismanuscriptF. LaBella andC. Vesco(J. Virol. 33:1138-1150, 1980) published similar results.
LITERATURE CITED
1. Bonner,W.KL,andR. A.Laskey.1974. Afilm detection
methodfortritium-labeledprotein andnucleic acidsin
polyacrylamide gels.Eur.J. Biochem.46:83488.
2. Chen,Y.-IL,J. P.MacGregor,D. A.Goldstein,and
KLR.Hall.1979. Histonemodifications insimianvirus 40and innucleoprotein complexescontaining super-coiledviral DNA. J.Virol.30:218-224.
3. Chestier, A., and K. Yaniv. 1979.Rapid turnover of
acetylgroups in the four core histones of simian virus 40minichromosomes. Proc.Natl. Acad.Sci. U.S.A.76:
46-50.
4. Coca-Prados, M., andM.-T. Hsu. 1979. Intracellular formsof simian virus 40nucleoprotein complexes. IL.
Biochemical and electron microscopicanalysis ofsimian
virus 40virion assembly. J.Virol. 31:199-208. 5. Cremisi,C.,A.Chestier, andKLYaniv.1977.
Prefer-ential associationofnewlysynthesizedhistones with
replicating SV40DNA. Cell 12:947-951.
6.Fareed, G. E., andD.DavaL1977.Molecularbiologyof
papovaviruses.Annu. Rev.Biochem.46:471-522. 7. Fernandez-Munoz, R., M. Coca-Prados, and M.-T.
Hsu. 1979.Intracellularfonns of simian virus 40 nu-cleoprotein complexes.L.Methods ofisolationand char-acterization inCV-1cells. J. Virol.29:612-623. 8. Jackson, V., A. Shires, N. Tanphaichitr, and R.
Chal}ley.1976.Modificationstohistonesimmediately aftersynthesis. J. Mol. Biol. 104:474-483.
9.LaBela, F., G. Vidali, and C. Vesco. 1979. Histone acetylationinCV-1cellsinfectedwithsimianvirus40.
Virology96:564-575.
10. Laemmli, U. K. 1970. Cleavage ofstructural proteins duringtheasemblyof thehead ofbacteriophageT4. Nature(London)227:680-685.
11.MacGregor,J.P.,Y.-ILChen,D. A.Goldstein,and
KLR.Hall.1978.Propertiesof the histones associated with simian virus40 replicative form nucleoprotein complex. Biochem. Biophys. Res. Commun. 83:814-821.
12. Panyim, S8,and R. Chalkley. 1969.High resolution
polyacrylamide gel electrophoresisof histones. Arch. Biochem.Biophys.30:330-337.
13. Phillip,D. K P.1963.Thepresence ofacetylgroups in histones.Biochem.J.87:258-263.
14. chafhausen,B.S.,and T.IL Benjamin. 1976. Defi-ciencyinhistoneacetylationinnontransforminghost
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.499.56.232.72.301.2]rangemutantof polyomavirua Proc. Natl. Acad. Sci. 16. Staneloni,R.J.,M. M.Fluck,and T. LBenjamin. U.S.A.73:1092-1096. 1977.Hostrangeselection oftransfonnation-deficient
15.Shmatcheko,V. V.,and A. J.Varshavaky. 1978.A hr-t mutantsofpolyoma virus. Virology 77:598-609.
technique oflow-pH gelelectrophoresisofchromosomal 17. Tan, K. B.1977.Histonesmetabolismin simian virus 40
proteinswhich doesnotrequirepreliminaryremovalof infected cells andincorporationintovirions.Proc. Natl. DNA.Anal. Biochem.85:42-46. Acad.Sci.U.S.A. 74:2805-2809.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 1.phoresisstainedentscintillationSV40slicedProtosol Acid-urea-polyacrylamide slab gel electro- analysis of cellular histones (A, scan of gel) and of [3H]lysine-labeled histones pres- in SV40chromatin (B) and CsClgradient-purified virions (C)](https://thumb-us.123doks.com/thumbv2/123dok_us/1488464.101516/2.499.268.439.145.596/phoresisstainedentscintillationsv-slicedprotosol-polyacrylamide-histones-histones-chromatin-csclgradient-purified.webp)