0022-538X/91/010514-06$02.00/0
CopyrightC) 1991, AmericanSociety for Microbiology
Identification of Cellular Factors That Bind Specifically
to
the
Epstein-Barr Virus Origin of DNA Replication
SANG-JIN OH,THOMASCHITTENDEN, ANDARNOLD J. LEVINE* Department of Molecular Biology, Lewis ThomasLaboratory, Princeton University,
Princeton, New Jersey 08544-1014
Received 26 July 1990/Accepted 25 September 1990
Thespecific binding of HeLa cell factorstoDNAsequencesattheEpstein-Barrvirus(EBV) latent origin of
DNA replication was detected by gel shift experiments and DNase I footprinting analysis. These cellular
proteins protectedatleastfivediscreteregionsoftheDNAreplicationorigin. Theviralprotein requiredfor
EBV plasmid replication, EBV nuclearantigen 1 (EBNA-1), binds to specific sequences within the origin
region. The HeLa cellproteins competed withEBNA-1for bindingtoEBVoriginDNA invitro,leadingtothe
possibilitythat these cellularproteins regulateEBV DNAreplication bydisplacingEBNA-1attheoriginsites.
The herpesvirus Epstein-Barr virus (EBV) infects and
immortalizes human B lymphocytes in culture(reviewed in
references5 and 13).The EBV genomeis usuallymaintained as a multicopy plasmid in latently infected cells (7, 10).
Extrachromosomal maintenance of EBV plasmids requires onlytwoviralgenetic elements: 1,800bp oftheviralgenome thatfunctionsas anorigin ofreplication (oriP)(6, 11, 17,21) anda trans-actingfactor, EBV nuclearantigen 1 (EBNA-1)
(9, 11, 22). Maintenance ofEBV latent infection is depen-denton EBNA-1both forreplicationof EBV episomes and
fortransactivation oftranscription fromthe EBVBamHI-C latency promoter (18). Viral plasmid replication is
coordi-nately regulated with cellular DNA synthesis in that each
EBV genome duplicates only once pergeneration in the S
phase ofthe cellcycle(1,8).
The EBVorigin of replication,oriP,containstwoessential
components (11, 17, 21). Onecomponent is aregion
com-posed of20tandemly repeatedcopies ofa30-bp sequence. Each of these repeated sequences contains a 12-bp palin-dromiccorewhich constitutesanEBNA-1binding site.The
family of 30-bp repeats can function as a transcriptional
enhancer element in the presence of EBNA-1 (16, 18). The second essential component of oriP is a region of dyad
symmetry which constitutes the actual site of initiation of viral DNAreplication (6). Thisregion contains fourrelated
copies ofthe 30-bp repeats, of which two are embedded withina65-bp sequence of dyad symmetry and the
remain-ing two are located together at the base of the dyad se-quence. Previous studies using deletion mapping have
iden-tifiedacritical140-bp region encompassing the dyad element
(between the EcoRV and HpaI sites within oriP) as neces-saryincombination with the 30-bp repeats for EBV plasmid
maintenance(3, 11, 17).
With the exception of EBNA-1, EBV relies entirely upon the cellular DNA replication machinery. The properties of EBVplasmidreplication suggest that viral DNA synthesis is linkedtothecellularmechanisms thatgovern chromosomal DNA synthesis. Thus, it is possible that the virus exploits cellularfactors which bindto oriPand regulate the replica-tion of EBV. To identify cellular proteins possibly involved in EBV plasmid maintenance, protein-DNA gel retardation
experiments and DNase I footprinting were performed,
*Corresponding author.
using HeLa cell nuclear extracts and the 140-bp oriP dyad symmetry region. Recombinant oriP plasmids encoding EBNA-1 replicate autonomously in HeLa cells (11, 22), demonstrating that these cells express all trans-acting
fac-torsnecessaryfor EBVplasmidmaintenance. In thisstudy, weidentify thebinding sites of HeLa cellproteinswithin the EBVoriPdyadregion. These cellular factorscompetedfor
binding with a bacterially produced carboxy-terminal do-main of EBNA-1 (28,000-molecular-weight EBNA-1 [28K EBNA-1]) (14, 15).
To detect cellularproteins that bind to the 140-bp oriP
dyad region, nuclearextractspreparedfrom HeLa cells (4) were used in a protein-DNA gel shiftassay (Fig. 1). Blue-script(Stratagene) plasmidscontaining subcloned oriP DNA
fragments (3) were used to generate end-labeled probes representing the 140-bp EcoRV-to-HpaI oriPdyad symme-tryregion (nucleotides 8897to9136;numberingasdescribed by Baer et al. [2]). Plasmids were linearized at polylinker
restriction sites adjacent toeither the EcoRV or the HpaI
end of the dyad region, end labeled with a-32P-labeled
deoxynucleoside triphosphates and Klenow fragment, and
digested with a second restriction enzyme to excise the
140-bp-labeledfragments. Labeled oriPfragmentswere
pu-rified after isolation fromagarosegels. Nuclearextracts(5to
10 ,ug) wereincubated for 30 minatroomtemperature with 0.5 to 1 ng of end-labeled 140-bp probe and 1
pug
of poly(dI-dC) in 12.5 ,ul of buffer containing 15 mMN-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES; pH 7.9),50 mMNaCl,0.5 mMEDTA,and1 mM
dithiothre-itol. Samples were electrophoresed at 200 V on 4% poly-acrylamide gels in 10 mM Tris hydrochloride (pH 8.0) containing0.1mM EDTAfor 2.5 h.
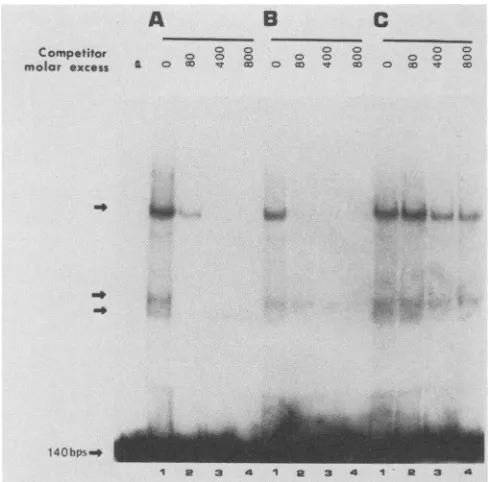
Three DNA bands ofreduced mobility wereobservedin thesenondenaturing gels (Fig. 1A),and thespecificityof the
majorshifted bands wasinvestigated bycompetition exper-iments. Two DNA fragments were employed as specific competitors: the homologous 140-bp EcoRV-to-HindIII dyadfragmentanda36-bp oligonucleotide correspondingto
the consensus 30-bp oriP repeat sequence that binds EBNA-1 (termed the EBNA-1 consensus oligomer; see
legend to Fig. 1B). As described above, four partial
ho-mologs ofthisconsensusrepeat sequencearepresentwithin the 140-bp dyad region. Addition of the homologous unla-beled 140-bp dyad fragment during the binding reactions caused the disappearance ofall bands(Fig. 1A). The36-bp
514
on November 10, 2019 by guest
http://jvi.asm.org/
A B 0a
Competitor 0 0 0 0 0
molar excoss ° e sr a:0o It e
so
-140bps
FIG. 1. Detection of cellular proteins that bindtc dyad region by protein-DNA gel shiftassays.A1-n,
end-labeled probe corresponding to the 140-bp
dyad region and 1,ug of poly(dI-dC) were added HeLa nuclear extract together with several diff double-strandDNAcompetitors,asindicated below binding reaction, the mixturewas electrophoresed
naturing polyacrylamide gel. Competition experin
formed by including in the binding reaction mixturt
gous(A)orheterologous (B and C)competitor DN excess over theprobeDNA indicated above each petitor DNAs were as follows: (A) the 140-bp oi
fragment; (B)a36-bpEBNA-1consensusoligomer(
TTAGGATAGCATATGCTACCCAGATATA-3 ATCTGGGTAGCATATGCTATCCTAATCTG-nonspecific oligonucleotide (5'-GGCACCAGCT'
ACAGTGTAAAAAAGGGCC-3':5'-GGCCCTTI TGACTGATTGAGCTGGTGCC-3'). Arrows in( of bands of altered mobilitywhich result fromspec
of HeLa nuclearproteins and the140-bp oriPdyad
EBNA-1 consensus oligomer also effectively
binding of cellular factors to the 140-bp dya( (Fig. 1B), whereas a 37-bp DNA fragment
sequence did not compete (Fig. 1C), indical complexes resulted from sequence-specific This result indicates that those cellular factoi specifically with the 140-bp dyad region als EBNA-1 consensus repeat oligomer. Furth findings suggest that the cellular factors bin recognition sites, sinceboththe 140-bp dyad EBNA-1 consensus oligomer sharethis sequt
In order to identify the nucleotide sequen
oriPdyadregion (nucleotides 8897to9136)th; factors, DNase I footprinting was performec
finalreaction mixture (50
RI)
contained 4ng(labeled140-bporiPdyadprobe (seeabove), 1 dC), 8 mMHEPES(pH7.9),24 mMKCI, 1m
mM EDTA, 0.2 mM dithiothreitol, 8% glyce 200 pug of HeLa nuclear extract. A 46-pd re
that containedallcomponents exceptthelab incubatedonicefor15min. Theprobe DNA
c
4-pd
volume,
and the reactionmixturewasincubated at 25°Cfor10min. DNase I was
added,
andincubationwascontin-COg 0 ued at room
temperature
for 1 min. The concentration of DNase I required to obtain uniformdigestionateachextractprotein concentration was determined
empirically
andrangedfrom 0.6 to 7.5
,ug/ml.
After theadditionof 300RI
of 10 mM Tris hydrochloride (pH7.4)-300
mM NaCl-20 mMEDTA-1% sodiumdodecyl sulfate-50
p.g
of tRNA perml,
the DNA waspurifiedby extraction withphenoland chlo-roform andanalyzedon8%polyacrylamide
sequencing gels.
_"*s1+iiiii
flok The sizes of thedigestion products
were compared with thoseof productsgeneratedby
specific
chemicalcleavage
of thelabeled DNA(12).DNase Iprotectionanalysisdemonstrated that the
140-bp
oriPdyadregion containsmultiplebinding
sites for cellular factors. At least five DNase I-protected regions were de-tected within the 140-bp oriP dyadregion
on thecoding
strand, designated 1 to5(Fig. 2A), andatleast fourregions
were detected on the noncoding strand,designated
1 to 4 (Fig. 2B). Anadditionalprotectedregion
(broken
line inFig.
2A)was alsoobserved in thecoding
strand. The nucleotide sequenceofeachoftheseprotected
regions
is summarized in Fig. 2C.Mostoftheprotectedregions
areflankedby
DNase 1 a 3 4 I-hypersensitivesites. One of theseregionscorresponds
toaZ
the 140-bp oriP sequence
previously
identifiedby
deletionanalysis
asessen-thamount
of
32p_ tial forplasmid
maintenance(region
1 inFig. 2C) (3).
TwoEcoRV-to-HpaI protected
regions,
2 and5,
are located onthe stem of theto 5 to 10 ,ug of 65-bp dyad. Footprints 1, 3, 4, and the broken-line region
rerent unlabeled overlapwith the EBNA-1recognitionsequences and sharea e. After 30minof core ATATsequenceand similar adjacent sequences. These on a 4%nonde- findings and the results of gel shift assays (Fig. 1) suggest nents were per- that one or more proteins in HeLa cell extracts bind to
eeitherhomolo- sequences within oriP thatcoincide with the EBNA-1 rec-[As inthemolar ognition
sites.
.riP
dyad region Consistent with thispossibility, specific HeLa cell-protein 'S'-GATCCAGA interactions with the EBNA-1 consensus oligomer were':5'-GATCTAT detected by gel shift assays (Fig. 3). A nuclear extract from -3'); (C) a 37-bp HeLa cells was
subjected
tochromatography
on acation-CAATCAGTC exchange matrix
(SP-5PW),
and fractions were testedby
arTTTACACTG band shift assay using a
32P-5'-end-labeled
EBNA-1consen-dicate positions sus oligomer (Fig. 3). This analysis revealed three
protein-cific
interactions DNA complexes of reducedmobility (arrows
a,b,
and c,Iprobe. Fig. 3). The activities giving rise to the a and bcomplexes (fraction 9) were separated from those
producing
the ccomplex (fraction 13,
Fig. 3A).
Competition
experiments
competed for wereperformedwith the EBNA-1 consensus
oligomer
probe
d region probe and either fraction 9
(Fig. 3B)
or fraction 13(Fig.
3C).t of unrelated Complexes a, b, and c were inhibited
by
the addition ofting that these unlabeled
homologous
DNAduring
thebinding
reaction but interactions. were notinhibitedby
anonspecific
oligonucleotide
(Fig.
3B rs thatinteract and C). Nonspecificoligonucleotides
containing
single-io bind to the stranded DNA end structures similar to the EBNA-1
oligo-termore, these mer also did not compete for
binding
to thesecomplexes
id to EBNA-1 (data not shown). These results indicate that allthree
activ-region and the ities resulted from
sequence-specific
interactions.ence. Rawlins et al.
(15)
demonstrated that a 28-kDa fusionIces within the protein
containing
thecarboxyl-terminal
one-third of atbind cellular EBNA-1 (28KEBNA-1)
iscapable
ofbinding
torecognition
I (Fig. 2). The sites within the tandem30-bp
repeatregion
and thedyad
of the
32P-end-
symmetryregion
of oriP. To examine therelationship
be-.
jig
ofpoly(dI- tween EBNA-1binding
and the HeLacellfactorsbinding
toM
MgCl2,
0.08 oriP, the in vitro interactions between the HeLa cellular -rol, and 25 to proteins and 28K EBNA-1 with the EBNA-1 consensusaction volume oligomer were studied. 28K EBNA-1 was
expressed
in eled DNA was Escherichia coliharboring
pNAK28
andpartially purified
was added in a according to the method of Milman et al.
(14).
Theon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.55.299.72.313.2]A.
Coding
B. Non-coding
G
1
2
34
A
_. A
0
a
_ *-W., n_ _
_w
-_
_
_
0
S"
a
a0
a0
e0
j0
II
a, 4
I I
Is
I"
I
0
A
1 2 3
Nw_
-0-_
U-_ a _
wew
4~... M
am
P
_~..0 am .
a
$4..,
C. A
B
88971 p 4-i
"ATATTCC C! GTTCCTTAGGACCCTTTTACTAACCCTAATTCGATAGCATATGCTTCCCGTTGGGTAACATATC
EcoRV
a3-C
D
9136 TTGFAATTAGGGTTAGTCTGGATAGTATATACTACTACCCGCGAAGCATATGCTACCCGTTTAGGGTTAAC
[image:3.612.69.550.69.562.2]|
¶-s--
----
~~~~~~
Hpal
FIG. 2. DNase Ifootprinting of theoriPdyadsymmetryregion (nucleotides8897 to9136)of EBV. The140-bp EcoRV-to-HpaIoriPdyad regionwas32Pendlabeledasdescribedinthetext.(A) Coding strand.Theprobewasincubated withnoprotein (lane 1)orwith HeLa extract (lane 2, 25 ,ug; lane 3, 100 ,ug; lane 4,200 jLg) anddigested with DNaseI. DNAwasfractionated by electrophoresis on8% denaturing polyacrylamide gels, and the cleavage products were visualized by autoradiography. Lanes G+A correspond to apurine-specific DNA
sequencemarkerasdescribed by Maxam and Gilbert (12). Arrows indicate the 65-bp dyadsymmetrystructure.Proteinprotected regionsare
numbered 1to5.Thebroken line indicates weaker levels of protein protection. (B) Noncodingstrand. Lane1,Noprotein;lanes 2and3,100 and200 jigofHeLa nuclearextract,respectively. (C) Summary of protected regions within the140-bporiPdyadsymmetryregion.Thick
horizontallinesrepresentfootprints 1 through 5 shown in panel A. Four partial homologs of the30-bp repeatedmotif(EBNA-1binding sites)
aremarked Athrough D. Arrows indicate the 65-bp dyad symmetry structure. NucleotidenumberingisasdescribedbyBaeretal.(2).
ally synthesized 28K EBNA-1 bound to the consensus
oligomerDNA inagelshiftassay(Fig. 4A).Twospeciesof reduced mobility (Fig. 4A, arrows) were shown to be the
resultsofspecific interactions between 28K EBNA-1 protein
and DNA by competition experiments, while the third
ap-pearedtobenonspecific (lower band, Fig. 4A). In order to determine whether 28K EBNA-1 could compete for the
same sites on the oligomer which are recognized by the HeLa cell factors, increasing concentrations of 28K EBNA-1 were added toa fixed level of HeLa cell extract
G
A
I
I
Ia
13
P 4
on November 10, 2019 by guest
http://jvi.asm.org/
input
In
_~~~~~~~
PT
0~~~ 0
S
_£~~~~~~
I I
_. ~ ~ 2
k~~~~~~~s a0
'a
0
2O
Z
so
U,, a
~~son
I
ow
~~~~~Ipu
t o
r
8~~~0
<I
* 00s 1 80SO
0-C)
4a
on November 10, 2019 by guest
http://jvi.asm.org/
A Competitor
Specific
N.pidfic
0
5
8o
a CM a v- 0*a 0la
0
0
B
1 2 3 4 5 6 7
F -_ F4
FIG. 4. Interaction of HeLa cell proteins and 28K EBNA-1 with the EBNA-1 consensus oligomer. (A) Gel mobility shift assay. Competition experiments were performed with the homologous specificEBNA-1 oligomer andaheterologous nonspecific oligomer (see
legendtoFig. 1B and C). Arrows indicate positions of bands of altered mobility which result from specific interactionsof the bacterial28K EBNA-1protein (0.2 ,ugperreactionmixture) and the EBNA-1consensusoligomer. (B)Gel retardationassaywith variousconcentrations of bacterial28K EBNA-1 protein. Binding reactionswerecarriedoutwith 4,ug ofpoly(dI-dC) ina10-pl reaction volume containingaconstant
amountof HeLa cellextract(12 ,ug) and increasing concentrations ofanextractcontaining the 28K EBNA-1protein. Finally,the32P-labeled EBNA-1consensusoligomerwasadded, and the reaction mixtureswereincubated for 30minatroomtemperature.DNA-protein complexes
wereseparated byelectrophoresison a4%polyacrylamide gel. Lane 1, Noextract;lane 2, HeLaextract(12 ,ug); lanes 3to7, binding reaction mixturescontainingconstantHeLa nuclearextract(12
jig)
and increasingamountsof 28K EBNA-1fusionproteinextract(lane3through 7, 4, 8, 12, 16, and 20 ,ug, respectively). Arrows indicate positions of DNA-protein complexes whichwereinhibitedby competition.Extractscontaining the cellular DNA-binding proteins and EBNA-1 28K proteinwerenotpurifiedorenriched.
togetherwiththeEBNA-1consensusoligonucleotide probe. Competition experiments were carried out with concentra-tions of DNAoligomer probesandproteinatwhichallofthe
oligomer is bound (and is limiting in the reaction). As the amountof 28K EBNA-1wasincreased,thebands represent-ing HeLa cellprotein-DNA complexeswereeliminated (Fig.
4B). Under the conditions used in these experiments, the nonspecific third bandfrom 28K EBNA-1(Fig. 4A)wasnot apparent. The reciprocal experimentwasperformed with a fixedlevel of 28KEBNA-1and increasingconcentrations of HeLaextract. Inthis experiment, atleastoneband derived
from the 28K EBNA-1-oligomer complex was inhibited by the addition of HeLa extract (data not shown). These findingsindicate thatHeLacell factors competed with 28K EBNA-1 in vitro for binding to the EBNA-1 consensus
oligomer.
It ispossiblethatcompetition between cellular factors and
EBNA-1 could regulate viral DNA replication in vivo by excluding EBNA-1fromthe binding sites withinoriP. Con-ceivably, theinteraction of such cellular proteins withoriP could be involved in the control of EBV replication within
the cell cycle or could govern the copy number ofEBV
plasmids. It has been reported that two cellular factors
produced bya12-O-tetradecanoylphorbol-13-acetate
(TPA)-treated, EBV-negative Burkitt's lymphomacellline(BJAB)
are able to compete for and uncouple EBNA-1 binding to oriP(19). It remains tobe determined whether these TPA-induced cellularfactors are the same as, or relatedto, the
HeLacellfactorsdetected in this study. Binding ofEBNA-1
tooriPappearstoberequiredbothfor EBVreplicationand for activation of the30-bprepeatenhancerelement(16, 18, 20). Recent experiments by Sugden and Warren (18) dem-onstrated that EBNA-1 can trans activate the BamHI-C
latency promoter via the oriP 30-bp repeat enhancer ele-ment. Thus, cellular factor(s) that compete for EBNA-1
binding sites could also playan importantrole in the regu-lation of viralgene expression inlatently infected cells.
WethankUlrich Muller forhelpful discussions,Matthew Marton for providing SP-5PW fractions of HeLa cell extract, Gregory Milman forsupplying the plasmidpNAK28, and EduardoMontalvo
forreviewing the manuscript.
S.-J.OhwassupportedinpartbyaKorea Science and Engineer-ing Foundation scholarship. This work was funded by grant CA49271from the National CancerInstitute.
REFERENCES
1. Adams, A. 1987. Replication of latent Epstein-Barr virus
ge-nomesinRaji cells. J. Virol. 61:1743-1746.
2. Baer,R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell,T.J. Gibson, G. Hatful, G. S. Hudson, S. D. Satchwell,
C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA
sequence and expression ofthe B95-8 Epstein-Barrvirus
ge-nome.Nature(London) 310:207-211.
3. Chittenden, T., S.Lupton, and A.J. Levine. 1989. Functional
limitsoforiP, the Epstein-Barr virus plasmid origin of replica-tion.J. Virol. 63:3016-3025.
4. Dignam, J. D., R. M. Lebovitz, and R. Roeder. 1983. Accurate
transcription initiation by RNA polymerase II in a soluble
extract from isolated mammalian nuclei. Nucleic Acids Res.
4-:..
A
--BP
', ;-:;.
-.01.1
VIMP.II
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.105.511.71.308.2]11:1475-1489.
5. Epstein, M. A., andB.G. Achong (ed.).1979.TheEpstein-Barr virus. Springer-Verlag KG, Berlin.
6. Gahn, T. A., and C. L. Schildkraut. 1989. The Epstein-Barr virus origin of plasmid replication, oriP, contains both the initiation andtermination sites of DNA replication. Cell 58:527-535.
7. Gussander, E., and A. Adams. 1984. Electron microscopic evidenceforreplication of circular Epstein-Barr virusgenomes
inlatently infected Raji cells. J. Virol. 52:549-556.
8. Hampar, B., A. Tanaka, M. Nonoyama, and J. Derge. 1974. Replication of the resident repressed Epstein-Barr virusgenome
duringthe early S phase(S-1period) of non-producer Raji cells. Proc.Natl. Acad. Sci. USA 71:631-633.
9. Hearing, J. C., J.-C. Nicolas, and A. J. Levine. 1984. Identifica-tion ofEpstein-Barr virus sequences that encode a nuclear
antigenexpressed in latentlyinfected lymphocytes. Proc. Natl. Acad. Sci. USA 81:4374-4377.
10. Lindahl, T., A. Adams, G. Bjursell, G. W. Bornkamm, C.
Kascha-Dierich, and U. Jehn. 1976. Covalently closed circular duplex DNA of Epstein-Barr virus ina human lymphoid cell
line.J. Mol. Biol. 102:511-530.
11. Lupton, S., and A.J.Levine.1985. Mappinggenetic elements of Epstein-Barr virus that facilitate extrachromosomal persistence ofEpstein-Barr virus-derived plasmids in human cells. Mol. Cell.Biol. 5:2533-2542.
12. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
13. Miller, G. 1985. Epstein-Barr virus,p.563-589. In B. N.Fields
(ed.), Virology. Raven Press Publishers, New York.
14. Milman,G., A. L. Scott, M. S. Cho, S. C. Hartman, D. K.Ades,
G.S.Hayward, P.F.Ki, J.T.August, and S. D. Hayward. 1985.
Carboxyl-terminal domain of the Epstein-Barr virus nuclear antigen ishighly immunogenic inman. Proc. Natl.Acad. Sci. USA 82:6300-6304.
15. Rawlins,D.R., G.Milman,S. D.Hayward, and G. S. Hayward. 1985. Sequence-specificDNAbinding of theEpstein-Barr virus nuclear antigen (EBNA-1) to clustered sites in the plasmid maintenanceregion. Cell42:859-868.
16. Reisman, D., and B. Sugden. 1986. trans activation of an Epstein-Barr viraltranscriptional enhancer bytheEpstein-Barr viral nuclearantigen 1.Mol. Cell. Biol. 6:3838-3846.
17. Reisman, D., J. Yates, and B.Sugden.1985.Aputativeorigin of replication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol. 5:1822-1832.
18. Sugden, B., and N. Warren. 1989. ApromoterofEpstein-Barr virus that canfunctionduring latentinfectioncanbe transacti-vated by EBNA-1, a viral protein required for viral DNA replicationduring latent infection. J. Virol. 63:2644-2649. 19. Wen, L.-T., A. Tanaka, and M.Nonoyama. 1989. Induction of
anti-EBNA-1 protein by 12-O-tetradecanoylphorbol-13-acetate treatment of human lymphoblastoid cells. J. Virol. 63:3315-3322.
20. Yates, J. L., and S. M. Camiolo. 1988. Dissection ofDNA replication and enhancer activation functions ofEpstein-Barr virus nuclear antigen 1. Cancer Cells 6:197-205.
21. Yates, J. L., N. Warren, D. Reisman, and B.Sugden. 1984.A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infectedcells. Proc.Natl. Acad. Sci. USA 81:3806-3810. 22. Yates, J.L., N. Warren, and B.Sugden. 1985. Stablereplication
ofplasmidsderivedfromEpstein-Barr virusin various mamma-lian cells. Nature(London) 313:812-815.