JOURNALOFVIROLOGY, 7Nov.1994, p.6947-6958 0022-538X/94/$04.00+0
Copyright C 1994, AmericanSociety for Microbiology
Identification and
Characterization of
an
Epstein-Barr Virus
Nuclear
Antigen
2-Responsive
cis
Element
in the Bidirectional
Promoter Region of Latent Membrane Protein
and
Terminal Protein
2
Genes
GERHARD LAUX,* FRANKDUGRILLON,t CHRISTINE ECKERT, BRIGITTE ADAM, URSULAZIMBER-STROBL, ANDGEORG W. BORNKAMM
Institutfiir KlinischeMolekularbiologie und
Tumorgenetik,
im GSF-Forschungszentrumfiir
Umweltund GesundheitGmbH,
D-81377Munchen,
Germany
Received5May1994/Accepted 20July 1994
Epstein-Barr virus (EBV) transforms resting B cells in vitroveryefficiently. The nuclear viral protein EBV
nuclearantigen 2 (EBNA2) isabsolutelyrequiredfor thisprocessandalsoactsas atranscriptional activator
of cellular and viral genes. As shown previously, EBNA2 transactivates the promoters of the viral latent membrane proteins. It interacts indirectlywith anEBNA2-responsive cis element of the terminal protein 1
(TP1) promoter. To identify the sequences mediating EBNA2 transactivation of the bidirectional promoter
region driving expression of thelatent membrane proteins LMP and TP2 in opposite directions,weassayed the
effects of EBNA2ontheactivities ofpromoterdeletion and site-directedmutantsof TP2 and LMPpromoter
luciferasereportergeneconstructsbycotransfections into EBNA2-negative Burkitt's lymphoma cells. Wewere
abletodelineatean80-bpEBNA2-responsive region (EBNA2RE) between -232 and -152 relativetothe LMP RNAstartsitewhich could also mediate EBNA2-dependent activationonaheterologouspromoter. Sequences of20 and 32bplocatedat the 5' and 3' ends, respectively, of theEBNA2REwereboth essential for EBNA2
responsiveness. Full transactivation of the LMP and TP2 promotersseemedtorequire 20 bp of 5' adjacent
sequencesinadditiontothe80-bp element. Electrophoreticmobilityshiftassaysrevealed specific protein-DNA complexes formed attheEBNA2RE. Oligonucleotides from -181to -152 and -166to -132 relativetothe LMP RNAstartsitevisualized oneB-cell- andoneB-cell-plus-HL60-specific retarded protein-DNA complex, respectively.Additionally,anoligonucleotide from-253to-210 revealedtwospecific protein-DNA complexes with nuclearextractsfrom different B and non-Bcells, suggesting also the binding of ubiquitously expressed proteins on the EBNA2RE. Thus, these experiments defined a 80-bp cis element sufficient for conferring EBNA2 inducibilityand demonstrated specific interactions of cellular proteinsatDNAsequenceswithin the EBNA2RE, whichare critical fortransactivation by EBNA2.
Epstein-Barrvirus (EBV), a ubiquitous herpesvirus, is the causative agent of infectious mononucleosis, a self-limiting
lymphoproliferative disease. Additionally, the virus has been
implicatedinthedevelopmentof several humanmalignancies, including endemic Burkitt's lymphoma, nasopharyngeal
carci-noma, B-lymphoproliferative disorders in
immunocompro-mised humans, andHodgkin's disease(see reference 46 fora
review).
Invitro,EBV infectsprimary restingBcellsveryefficiently, resulting inunlimitedproliferation andoutgrowth oflatently
infected lymphoblastoid cell lines. In these immortalized cell
lines,theexpressionof EBVgenesis restrictedtothe so-called latent genes,whichencode six nuclearantigens (EBNA-1, -2, -3A,-3B, -3C,and-LP),threemembraneproteins(LMP, TP1,
andTP2; also denotedLMP1, LMP2A, and LMP2B,
respec-tively), and two small nuclear nonpolyadenylated RNAs, EBER1 and EBER2 (for a review, see reference 36). As
recently shown, the latent genes for EBNA2, EBNA-3A, EBNA-3C, and LMParerequiredforB-cell immortalization,
*Corresponding author. Mailing address: Institut fiir Klinische
Molekularbiologie und Tumorgenetik, GSF-Hamatologikum, Mar-chioninistr. 25,D-81377 Munchen, Germany. Phone: 49-89-7099510. Fax:49-89-7099500. Electronic mail address: [email protected].
tPresent address:Ciba-GeigyAG,CH-4002Basel, Switzerland.
whereas the others are not (14, 29, 35, 59). However, the
mechanism ofgrowth transformation by EBV is still poorly
understood.
The EBNA2 gene, which is absolutely essential for B-cell immortalization in vitro (14, 29), codes fora 487-amino-acid
phosphorylated protein (27, 28).Its unusualprimarystructure is characterized byacontinuous stretch of40proline residues nearthe N terminus,a negatively chargedC terminus, and a
glycine/arginine-rich region. EBNA2 is one of the firstgenes
expressed afterprimary infection ofB lymphocytes (2, 3, 49, 53, 67)andinduces theexpressionof cellulargeneslikeCD21, CD23,andc-fgr(9, 16, 17, 39,62, 63, 64).Inaddition,the viral
promotersforLMP, TP1, TP2,and the EBNAgenesarealso transactivated by EBNA2 (1, 58, 63, 65, 70). Since specific
mutations of the EBNA2geneabolished itsabilityto transac-tivate LMP andatthesametime itscapacitytoimmortalize B
lymphocytes in vitro (13, 65), it is assumed that EBNA2 contributestoB-cell transformationbytransactivatingcellular and viralgenes. Although a transactivation domainhas been mappedattheC terminus of EBNA2by usingGAL4-EBNA2 fusion proteins (12, 31), it is not yet known how EBNA2
transactivates its target genes. Recently, we showed that
EBNA2 interacts with an EBNA2-responsive cis element (EBNA2RE) of theTP1 promoter, presumablyviaa cellular DNA-binding proteinwithoutbindingthe DNAdirectly (69). 6947
Vol.68,No. 11
on November 9, 2019 by guest
http://jvi.asm.org/
6948 LAUX ET AL.
Additionally, it was demonstrated that a cellular protein could target EBNA2 to an EBNA2RE of the EBNA gene's Cp promoter (44).
To further elucidate the mechanism of EBNA2-mediated transactivation, we studied the bidirectional LMP/TP2 pro-moter region in more detail. The LMP promoter initiates transcription at position 169515 (numbering of Baer et al. [4]) leftward of the linear EBV genome (24, 33), whereas the adjacentTP2promoter directs transcription to the right of the standard EBV genome (41, 55). The exact transcription initi-ation site for the TP2 mRNA has not been mapped; however, the 5' end of the longest published cDNA clone was mapped to position 169752 (55). The bidirectional nature of this promoter region suggests that cis elements are shared by the two promoters. Previous reports indicated that DNA sequences downstream of -234 relative to the LMP mRNA start site are necessary fortransactivation of LMP by EBNA2 (22, 26, 60), and an inducible enhancer element which mediated transacti-vation by EBNA2 was localized between positions -92 and -234 of the LMP promoter (60). However, effects on the TP2 promoter have not been considered.
Wetherefore began to furtherdefine the EBNA2RE in the bidirectional LMP/TP2 promoter region in more detail by using progressive 5' deletions and internal mutations of the promoter region in front of reporter genes. This kind of analysis showed in transient transfection assays that common cis-regulatory elements are necessary for transactivation of the LMP and TP2 genes by EBNA2, affecting both genes differ-entially. Additionally, we could demonstrate specific protein-DNAinteractions within the described EBNA2RE by electro-phoretic mobility shift assays (EMSAs). The expression of two of these cellular DNA-binding activities is restricted to the hematopoietic and B-cell lineage, whereas two other
protein-DNAcomplexes demonstrated by EMSA within sequences of the EBNA2RE seem to derive from ubiquitously expressed cellular proteins which might be related to a protein also binding within the EBNA2RE of theTP1 promoter.
MATERIALS AND METHODS
Cell linesandculture conditions.Cell lines DG75 (5), Jijoye (30),IB4(37),BL41 (42), BL41.P3HR-1 and BL41.B95-8 (9), Raji(52), P3HR-1 (47), M-ABA (19), HL60(15), HeLa (25), HaCaT(6), and Jurkat (56) were used in transfection experi-mentsand for nuclear extract preparations. All cell lines except HaCaT weregrown inRPMI1640 mediumsupplemented with 10% fetal calf serum, 100 U of penicillin per ml, and 100 ,ug of streptomycin per ml. HaCaT was grown in Dulbecco modified Eagle medium with the same supplements.
Plasmids. The eukaryotic EBNA2 expression vectors were kindlyprovided by W. Hammerschmidt (29). p554 encodes a fully functional EBNA2, whereas p615 harbors a linker inser-tion with stop codons in all reading frames expressing only about one-third of theN-terminal region of EBNA2. Expres-sion of the EBNA2 gene is driven by the Cp and/or Wp promoters of EBV.
All standard manipulations involved in plasmid construc-tions were carried out by established procedures described by Maniatis et al. (45). PCRs were performed on plasmid TP2CAT (41), and the PCR products were cloned upstream of the luciferase (LUC) reporter gene. The primers RO (gccgca gatctcgcctctttgtgcagattacact) and LO (gcaagagatctcagggcagtgt gtcaggag) containing a BglII restriction site were used to amplify the entire bidirectional promoter regions of LMP (-327 to +40) and TP2 (-276 to +91), including their mRNA initiation sites. The amplified DNA product was cut withBglII
and inserted into the BglII site upstream of the promoterless LUC gene of the pBLLUC5 vector (39a), giving rise to reporter gene constructs LLO (-327 to +40) and, in reverse orientation, TLO (-276 to +91), respectively. The 5'-deletion plasmids of the TP2 promoter region were generated by directional cloning of the T4 polymerase-treated andBglII-cut PCR products obtained after amplification with primers RO and L6 (positions 169648 to 169669 in the numbering of Baer et al.[4]),RO and L7 (positions 169668 to 169687), and RO and L8 (positions 169688 to 169707) between PstI (blunt ended) and BglII sites ofpBLLUC5. The resulting DNA clones were designated TL6 (-104 to +91), TL7 (-84 to +91), and TL8 (-64 to +91), respectively. The 5'-deletion constructs of the LMP promoter region were prepared in the same way, using primers LO and R3 (positions 169714 to 169690), LO and R4 (positions 169669 to 169650), LO and R5 (positions 169647 to 169628), LO and R7 (positions 169594 to 169575), and LO and R9 (positions 169549 to 169530), generating plasmids LL3 (-199 to +40), LL4 (-154 to +40),LL5 (-132 to +40), LL7 (-79 to +40), and LL9 (-34 to +40), respectively. Plasmid LLla (-234 to +40) was constructed by digestion ofLLO with MscI andHindlll,blunt ending with T4 DNA polymerase, and self-ligation.
Reporter gene constructs Ga5O/7 and Ga68/8 were con-structed by inserting an oligonucleotide containing the 3-glo-bin minimal promoter without and with an Spl-binding site (66), respectively, in front of the promoterless LUC gene of pBLLUC5. Plasmids E/3g,
80/4g,
80'/3g, E/s3g, 80/s3g, and 80'/sIg were cloned on the basis of Ga5O/7 (,Bg) and Ga68/8 (s,Bg) and harboradditionally in front of the 3-globin minimal promoter either a 100-bp fragment (E, positions 169648 to 169747), an 82-bp fragment (80, 169647 to 169729), or an 80-bp fragment (80', 169667 to 169747) of EBV DNA of the LMP/TP2promoter region.Plasmids BA16 and BA17 were generated by inserting the XhoI (blunt ended)-HindIII fragments of TL7 and TL8 into the StuI-HindIII-cut plasmids LL5 and LL4, respectively. Constructs BA18 and BA19 were made by ligating theXhoI (blunt ended)-HindIII fragments of LL5 and LL4 into the Stul-HindIII-cut plasmids TL7 and TL8, respectively. Plasmids BA20 and BA21 were constructed by ligation of the blunt-ended XhoI-MscI fragment of TL8 into the blunt-ended HindIII site of plasmid LL3 and by ligation of theXhoI (blunt ended)-HindIII fragment of LL3 into the HindIII-MscI-cut plasmid TL8, respectively.
CMVLTRLUC was generated by replacing the chloram-phenicol acetyltransferase (CAT) gene of CMVLTRCAT (10) with the LUC gene (39a). CMV,3gal (Clontech Laboratories) expresses
3-galactosidase
(P-Gal)
under the control of the cytomegalovirus immediate-early promoter and was used as an internal control fortransfection efficiency (2 ,ug per transfec-tion).Allreporter gene constructs areschematically shown in Fig. 2 to 4 and have been verified by DNA sequencing. Plasmid DNA was prepared by the alkali lysis method (45) and purification on CsCl gradients (p554 and p615) or by column chromatography, using resins of Quiagen and Promega as instructed by themanufacturers.
Transfection of cells and LUC andn-Gal assays. Cells were transfected by electroporation as described by Cann et al. (10), withminormodifications (41). LUC assays were performed as previously described (69). ,B-Gal assays were done with 5,ul of the same extracts used for the LUC assays in order to normalize for transfection efficiency. ,B-Gal activities were measured in 0.1 mMMgCl2-100mMsodium phosphate buffer (pH 8.0); 10 ,ug of3-(4-methoxyspirol[1,2-dioxetane-3,2'-tricyclo
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
EBNA2-RESPONSIVE ELEMENT OF LMP/TP2 PROMOTER REGION 6949
,GCGTGTGTTTGTGTI CGCACACAAACACAJ
ICGCACACAAACACA&TCTTTTCGCCC
UAC
;CGTGTGTTTGTGTTAG GTGC CGCACACAAACACAATCTTTTC
CMG F
CAAGTGCAACAGG&TGGA GTTCACG1TGTCCTTTACCT
F2
1 2 3
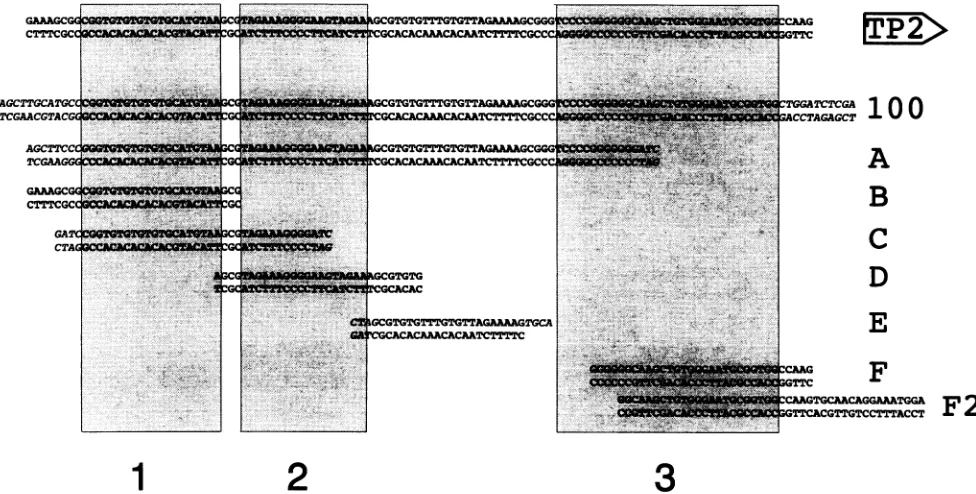
[image:3.612.64.552.80.327.2]FIG. 1. Oligonucleotides used for EMSA. Thetoplinerepresentssequencesof theLMP/TP2bidirectionalpromoterregionfrom -125to-237 relativetothe LMP RNAstartsite. Thedirection of the TP2geneisindicatedontheright byanarrow.Thesequencesofthe123-bpfragment (100) and ofoligonucleotides A, B, C, D, E, F, and F2 used for EMSAarelisted below. Bases initalic shownon-EBVsequencesusedtofacilitate cloning. Boxes 1, 2, and 3 indicate mutatedordeletedsequencesinplasmids BA16 plus BA17, BA18 plus BA19, and BA20 plus BA21, respectively.
[3.3.1.13.7]decan]-4-yl)phenyl f3-D-galactopyranoside (AMPGD; Tropix) permlwas usedas thesubstrate, and the reactionwas started with0.2 MNaOHand10% Emerald Enhancer(Tropix). Bioluminescence was measured (in relative luciferase units) withaLumat LB9501 (Berthold, Wildbad, Germany).
Nuclear extract preparation and EMSA. Nuclear extracts wereprepared byamodification ofthemethod of Dignamet al. (20) as described by Zimber-Strobl et al. (70). Binding
reactions for EMSAwere carried outaspreviously described
(69). The reaction products were separated on 3.25 or 5%
polyacrylamide (30:1) gelsrunin1xTris-borate-EDTA buffer. Gelswere dried and autoradiographed.
Oligonucleotides.The positionsofoligonucleotides used in relationtotheEBVgenomicsequenceasdeterminedbyBaer et al. (4)were as follows: A, 169649 to 169726; B, 169640 to 169670; C, 169648to169681; D, 169667to 169696; E, 169691 to 169711; F, 169721 to 169752; and F2, 169725 to 169768. Some oligonucleotidescontained additionallyunrelated bases at their ends to facilitate cloning (Fig. 1). Oligonucleotides
were synthesized by a Pharmacia Gene Assembler or pur-chased from MWG-Biotech. The123-bp fragment (Fig. 1)was
isolatedafterdigestionofplasmidTL6dwithHindlllandXhoI
harboringEBVsequences ofpositions 169648to169747. The
fragment and oligonucleotides were used in EMSA after
labelling byT4 DNApolymerasein thepresenceof[32P]dCTP.
RESULTS
Identification of an EBNA2RE within the bidirectional LMP/TP2 promoter region. As shown previously, LMP and TP2promoterCATconstructswere transactivatedbyEBNA2 in B cells (70). The clones contained either 327 bp of EBV DNA sequence upstream of the LMP transcription initiation siteor276bpupstreamof the 5' end of thelongest published
cDNA of TP2 (55),which isherein referred toasposition +1
of the TP2promoter. To delineate the EBNA2REmediating
transactivationbyEBNA2,weconstructedreportergene plas-mids which contain progressively truncated 5' upstream se-quencesofthe overlappingLMPandTP2 promotersin front of the LUC gene (Fig. 2). The plasmids were tested for
expression after transient cotransfections together with the EBNA2 expression plasmid p554 into EBNA2-negative BL41.P3HR-1 cells. CotransfectionswiththeEBNA2 expres-sionplasmid p615, carryingalinkerinsertion withstopcodons after about one-third of the EBNA2openreading frame,were carriedoutas acontrol forplasmid p554 expressingwild-type
EBNA2. Expression of the EBNA2 gene was driven by the viral EBNA promoters Cp and Wp (67). Plasmid
CMV-LTRLUCwasused as apositive control fortransfection, and thepromoterlessreportergeneplasmid pBLLUC5 servedas a
negative control for LUC expression. LUC expression was normalized for transfection efficiency by using a-Gal activity expressedbythe cotransfected CMV,Bgal plasmid.
The results of transfection experiments to analyze transac-tivation by EBNA2 with the individual promoter deletion
constructs are summarized in Fig. 2. Cotransfections of the LMPpromoter-LUCplasmidswithp615inBL41.P3HR-1 cells resulted in low or modest expression of the LUC gene. The basalactivitywasincreased about 10-foldbydeletingupstream
sequencesfrom -232 to -199 (data notshown). Cotransfec-tions withp554inducedLUCexpression51.9 + 16.0- and 26.8
+ 8.5-fold whenupstreamsequencesfrom +40to-327(LLO)
and -232(LLla), respectively,werepresent.The other dele-tionconstructs(LL3, LL4,LL5, LL7, andLL9) showednoor
onlyalow increase in reportergeneactivity (3.8 + 2.0-,1.7 ±
1.1-, 2.6 + 1.6-, 2.8 + 1.0-, and 1.3 + 0.7-fold, respectively)
after cotransfectionwith p554.
~~~~~~~~~ACCTAGAGCT
VA
B
C
D
E
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
6950 LAUX ET AL.
T Tr _
'%'+40 A AA A -0 +91
F LMPI A TP2
169515 169752
IUD
1 2 3
mm
-ILLmm
=
LL3_-199
L/cifoinsIE Z ZI I -1s4
LLS -132
LL9 4
B
+91 J-327
>-
+91-276L
LLO lL1a L13
[image:4.612.66.564.71.306.2]LL4 LL5 LL7 LL9 CMV pBL TLO TL6 TL7 TL8
I-I
FIG. 2. Deletion analysis of LMP and TP2 promoter reporter gene constructs for EBNA2inducibility. (A)Sequences coveredbythe promoter
deletion constructs used in transient transfections. The bidirectional promoter region indicated at the top shows the location of the LMP
transcription startsite (bentarrow)and the 5' end of thelongestpublishedTP2 cDNAclone(dottedbentarrow)with thepositionsrelativeto the B95-8 standard EBV genome(4).TATA-box-like sequences(TACAandTAGA),thesites used forcloning(verticalbarsat+40and+91),
andthreeregionsof the EBNA2RE(boxes 1to3)important for EBNA2responsivenessare also shown. The LMP(LLO,LLIa,LL3, LL4, LL5, LL7,andLL9)and TP2(TLO, TL6, TL7,andTL8)promoterdeletionconstructs arerepresentedasrectangularboxesshowingthedirection of the LUC reporter gene and also theendpointsof the promoter upstream sequences. (B)Activation of the individual clones after cotransfection with the EBNA2 gene(expression plasmidp554)isdemonstratedbyhorizontal bars with standard deviations. CMV andpBL representplasmids
CMVLTRLUC andpBLLUC5, whichwereusedascontrols.
The same cotransfection experiments wereperformed with consecutive deletion constructs of the TP2 promoter region. Luciferase activities were also low or modest with
cotrans-fected p615. The basal activity was increased fourfold and decreased fivefoldby deletingpromoter sequencesfrom -104
to -84 and -84 to -64, respectively (data not shown). Expression of the reporter gene increasedbyfactors of29 ±
14.6 and 41.1 ± 15.4when p554was cotransfected with TL0 (+91 to -276) and TL6 (+91 to -104), respectively. Only
a very low induction (2.4 + 1.1- and 2.8 ± 1.2-fold) of LUC expression by EBNA2 was detected after cotransfec-tion with plasmids TL7 (+91 to -84) and TL8 (+91 to
-64), respectively.
Since p554 could also expressEBNA-LP, EBNA2 expression plasmids which express EBNA2
but not EBNA-LP were additionally used to exclude the involvement ofEBNA-LPintransactivating theLMPandTP2 promoter region. Results for transactivation by EBNA2 were similar to those obtained with plasmids p554 and
p615(datanotshown),which is consistent with datapublished by Fahraeus et al. (22). The results presented above indi-cated that sequences between -232 and -199 of the LMP promoter and sequencesbetween -104 and -84 of the TP2 promoter contain ciselements essential fortransactivation of
the LMP and TP2 promoters, respectively. The DNA se-quences of LLla and TL6overlap by 100bp, suggesting that this common cis element constitutes essential parts of the EBNA2RE. However, results of the set of experiments
de-scribed above leave open the possibility that additional se-quenceelements adjacenttothe 100-bpcommon sequence are necessary toconfertransactivationbyEBNA2 ontheLMP or
TP2promoter.
Identification of important sequences within the 100-bp
EBNA2RE.Further plasmidswere constructedto create
spe-cific mutations in regions 1 to 3 of the LMP/TP2 promoter
region (Fig. 3). The constructs were tested for activation in
cotransfection experiments. Mutations in region 1 (plasmids
BA16 and BA17) reduced the inducibility byEBNA2 of the LMP promoterfrom 51.9 ± 16.0-to20.0+ 9.1-fold andofthe TP2 promoterfrom29.0 ± 14.6- to7.0 + 1.3-fold inducibility. Clones BA18 and BA19 harboring mutations in region 2
reduced transactivation by EBNA2 more than 10-fold
com-pared withthe unmutatedplasmids, leaving only5.1 ±2.0- and
1.8 ± 0.9-fold activation of the LMP and TP2 promoters,
respectively. Mutations in region3(plasmidsBA20 andBA21)
also diminished activation by EBNA2 for both promoters
drastically,to 2.0 ± 1.3- and 1.6 + 0.9-foldinduction,
respec-tively. Comparison of the effects on EBNA2 responsiveness due to the mutations in different regions of the 100-bp
EBNA2REshowed thatthe level of transactivationbyEBNA2
is reduced to a much lesser extent by mutations in region 1
thanbythose inregion2or3. Inaddition, region 1 mutations
affected the EBNA2-mediated induction ofthe TP2 promoter more strongly than induction of the LMP promoter. Also, region 2mutations impaired EBNA2 inducibility of the TP2 gene moreseverelythaninducibilityof theLMPgene.Region 3mutations affected theEBNA2responsiveness of bothgenes to the same degree.
Transfectionexperiments withreporter gene constructs
con-taining the EBNA2-responsive and nonresponsive promoters
were also carried out in EBV-negative Burkitt's lymphoma DG75 cells. Comparable resultswere obtained, although the
A _ T
fold activation
0
,10,.
20 30 40 50 6070
.,. .,
-104
IN
cfra9s-"imzzzM2
84
ImzzMsI
J. VIROL.
+40
--q
s
on November 9, 2019 by guest
http://jvi.asm.org/
EBNA2-RESPONSIVE ELEMENT OF LMP/TP2 PROMOTER REGION 6951
A +91
LMPI
A1
TP21615 169752
+4o--i
ILLO
1 2
3>
-1II--+91
1-327
IBA16
lR
1IBA18
1l
-I
+40
-276
F
;']--B
LLO
BAl
6
BAl
8
BA20
+91
TOLdI
,.
mmi BA171
1'-1
so
mBAI9
71NON ..,I Luc
1'F-1
n
EWII'TLO
BAl
7
BAl
9
BA21
fold activation
25C
::G:CATGCAAGC:TGCATGCC::./::::::::::::::17
IGGCGGTGTGTGTGTGCATGTAAGCGTAGAAAGGGGAAGTAGAA
AG/ /GGGTCCCCGGGGGGCAAGCTGTGGGAATGCGGTGGCCA:::::::::::::::::::::::::GCATGCAA:CTT:C:TGCC:://::::::::::::::::::::::::::::::::::::::
I wt BA18/19
.:::::::::::::::::::::::::::::::::::::::: :://:::G:ATGCAA:CT---::: BA20/21
1
2 3FIG. 3. Analysisofinternalmutations in theLMP andTP2promoters for EBNA2responsiveness. Aschematic representation of plasmids mutated(cross-hatched boxes)inregions 1to3 oftheEBNA2RE(A) and their activation by EBNA2 (B)areshown.Otherindicatedfeaturesare
thesameasdescribedforFig.2.(C)ThemutatedsequencesofplasmidsBA16toBA21.Thewild-type(wt)EBVsequenceisboxed. Identical and deleted bases of the EBNA2REarerepresented bycolonsanddashes,respectively.Twodiagonalstrokesindicate unmutatedsequencesbetween regions2 and3omitted forclarity.
transactivationefficiencybyEBNA2wassomewhat lower(data notshown).
A DNA sequence of 80 bp of the bidirectional LMP/TP2 promoter is suficient to confer EBNA2 inducibility on a
heterologous minimal promoter. Since the LMP and TP2
promotersappearedtorequire region2 and 3sequencesof the EBNA2REandregion 1sequencesonlytoalesserextentfor
mediatingtransactivationby EBNA2,itwasof interesttostudy
the cis sequence in the context of a heterologous minimal promoter. The 100-bp overlappingsequences (containing
re-gions 1 to 3) and the 80-bp sequences (containing either
regions1 and 2or 2and3)were cloned in front of the LUC reportergeneandthe ,B-globin minimal promotercontaining onlytheTATA boxortheTATA boxandanSpl-bindingsite
(66). Theresults, summarizedin Fig.4, clearlyshow that the
80-bp sequence containing regions 2 and 3 is sufficient to transmit about a20-fold EBNA2inducibilityto the
heterolo-gous 3-globin minimal promoter regardless of whether an
Spl-binding site is present. Region 1 was dispensable for transactivation of the heterologous promoter by EBNA2,
whereasregion3wasrequired. Thenecessityofregion2was nottestedinthissystem.
Specific protein-DNAinteractionswithin theEBNA2RE of
the LMP/TP2 promoter region. EMSAs were performed to identify specific protein-DNA interactions within the
delin-eated EBNA2RE of the bidirectional LMP/TP2promoter. A
123-bp DNA fragment containing the 100-bp EBNA2RE
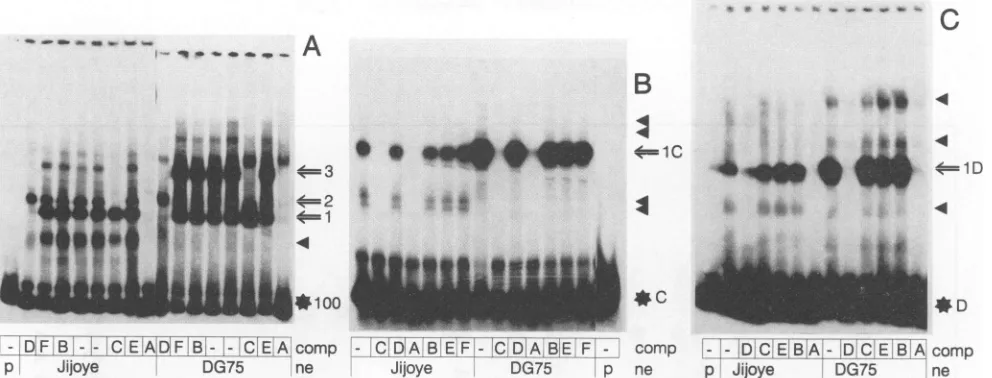
(-132 to -232 relative to the LMP transcription initiation site)wasexcised fromplasmid TL6d, labelled, andincubated with nuclear extracts of DG75 (EBV-negative) and Jijoye (EBV-positive) cells (Fig. 5A). The EMSA revealed three
prominent protein-DNA complexes ofJijoye and DG75 nu-clear extracts binding to the EBNA2RE. A fourth
protein-DNAcomplex migrating faster than complex 1 wasobserved withextractsofJijoye cells. Thiscomplexwas notspecificfor
EBNA2-positive cells(seealsoFig. 6A)butratherrepresents adegradation productpresent insomeextractpreparationsas a result ofprotease activities. Competition experimentswith unlabelled oligonucleotidesAtoF(see Fig. 1 forsequences)
were carried out in 100-fold molar excess over the labelled fragment to define shorter sequences within the 100-bp
EBNA2REinvolved inprotein-DNAinteractions.
Oligonucle-otide A(Fig. 5A), encompassing80bpof the100bp,competed
for all protein-DNA complexes generatedwith both extracts exceptforaweakcomplexinDG75 extractsmigrating slightly
more slowly than complex 3. This complex was also not competed for by a 50-fold molar excess of the unlabelled 123-bp fragment (data not shown) and therefore mostlikely represents an unspecific protein-DNA interaction. The re-tarded complexes 1 and 3were efficiently competed forby a
50 75
--I
I
-~~~~~~~~~~-'I'
VOL.68, 1994
0
IFsm
BA16/17
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.66.557.78.287.2]6952 LAUX ET AL.
+40 A
I c +91
1 2
3
-132 AA AG
gobTATA|
IN
ABlob
TATA Luciferase_MIJI[L]
l3glob
TATA-152 -232
-132 A A T
Spl
I
Bglob
TATA|*-
AiSpl BgIobTATA
IEZII
ISpl IBglobTATA
pI
IBglob
TATAl
-152 -232
activation
0.9
17.0
0.9
22.5
0.9
17.5
0.8
24.5
FIG. 4. A80-bpsubsequence of the 100-bp EBNA2RE (E)canconfer EBNA2responsivenesson aheterologous 3-globin minimalpromoter
(,BglobTATA)oronthesamepromoterwithanadditionalSpi transcriptionfactor-bindingsite(Spi ,Pglob TATA).The -232to-132 sequences of the LMP promoterwereinsertedimmediatelyupstreamofthe minimal promoters ofplasmids Ga5O/7andGa68/8; 20-bp deletionsatthe 3' (80/3g and80s,gb) and 5' (80'/,g and80'/sIg) ends ofthe 100-bpsequence were also testedforEBNA2inducibility. The average levelsof activationbyEBNA2 in threeindependent experimentsareshownattheright. Otherfeaturesare asdescribed inlegendtoFig. 2.
100-foldmolar excessofoligonucleotide D,whereas oligonu-cleotide C competed for complexes 2 and 3. None of the complexeswerecompeted for byoligonucleotides B, E,and F.
Complexes 1 and 2mightrepresent thesingle bindingofonly
one of two different factors to the 123-bp fragment
distin-guished bythedifferentpositionsofmigrationinEMSA andby competitionwitholigonucleotides C andD, respectively. The binding of both factorstothe123-bp labelled fragment might
result inan even strongerretardationpossiblyrepresented by complex3. This view issupported bythe factthat complex3 was specifically competed for by oligonucleotide C or D,
resulting in thevisualization ofonlyoneretardedcomplexin thecompetition experiments (Fig. 5A).
Further studies of the protein-DNA interactions on the EBNA2REwere performed with labelled oligonucleotides C and D(Fig.SBandC),since theseoligonucleotides competed specifically for individual protein-DNA complexes detected
withthe123-bpsequence as aprobe. One major complexwas visualized with each of labelled oligonucleotides C (complex 1C) and D(complex 1D). Complex 1C migratedmore slowly than complex 1D, as was expected from the competition
experiments shown inFig. 5A.Additionally,minorcomplexes
could be detected with both probes. Faster-migrating
com-plexeswere seenwithvariable strength depending on extract preparations and storage times, which suggested that they
might represent degradation products of the major protein-DNA complexes. Minor complexes with slowermobility than
complexes 1C and 1D were also seen and appeared to be specific, since they could be competed for by unlabelled oligonucleotides C and D. The nature of these complexes
remainsunclear. Oligonucleotides B,E, and F,derived from
adjacentsequences, didnot compete foranyof the retarded
complexes.
No major differencesbetween nuclear extractsof
EBNA2-positive (Jijoye) andEBNA2-negative (DG75) cells could be detectedbyEMSA. Control shiftexperimentsruninthesame
gels with oligonucleotide 054bp of the TP1 promoter (69) easily demonstrated the EBNA2-containing complexes de-scribedbyZimber-Strobletal. (70).Inaddition,EMSAswere
performed with the 123-bp labelled DNA fragment of the
LMP/TP2 promoter region andEBNA2-negative nuclear ex-tracts of DG75 cells supplemented with in vitro-translated
EBNA2. Under theseconditionsalso, no additional
EBNA2-specific retardedband couldbedetected evenafter overexpo-sure of theautoradiograms (datanotshown).
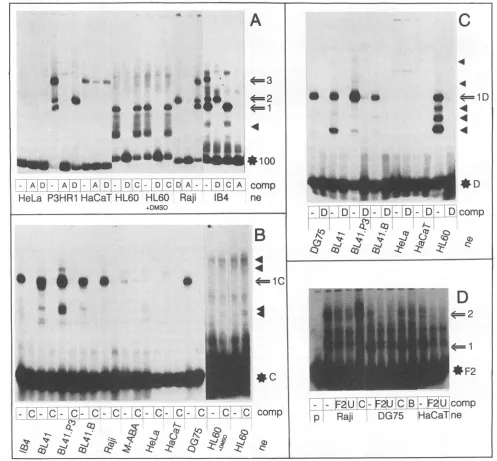
Tissue specificity of DNA-binding proteins detected by EMSA using sequences of the EBNA2RE of the LMP/TP2 promoter region. EMSAs were performed with nuclear ex-tractspreparedfromavarietyof differentcelllines,including EBV-negative (DG75 andBL41), EBV-positive but EBNA2-negative (BL41.P3HR-1 and P3HR-1), and EBV- and EBNA2-positive (BL41.B95-8, Raji, Jijoye, and IB4) B lym-phocytesaswellasnon-B-cell lines(HeLa, HaCaT,andHL60) inorderto studythe tissuespecificity oftheproteins involved
in complex formation. The results with the labelled 123-bp
DNAfragment andoligonucleotidesC,D, andF2areshown in
Fig. 6. All specific complexes shown in Fig. 5 were detected with nuclear extracts made from other B-cell lines, both EBNA2 positive and negative. Nuclear extracts made from
HeLa or HaCaT cells did not form specific protein-DNA complexes inEMSA performed witha labelled 123-bp frag-ment(Fig. 6A).Oligonucleotide F2, overlapping partiallywith the123-bpfragmentatits 3'endby23bp,revealedtwospecific protein-DNA complexes seen with nuclear extracts of Raji, DG75, and HaCaT cells (Fig. 6D). However, the complexes
were observedonlywhen theelectrophoreticconditionswere
LMP
A
TP2
169515 169752
Ga5O/7
E/Bg
80/Bg
80'/Bg
Ga68I8
E/sBg
80/sBg
80'/sBg
J. VIROL.
T T T
A A A 1.
G G G
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.75.557.78.338.2]EBNA2-RESPONSIVE ELEMENT OF LMP/TP2 PROMOTER REGION 6953
I- V
___
A
B
v*
e
*
I_
ALar-
a_*
=4xc=1D
IE
I
4
#C
-DF B--C EADF B - -CE A comp
p Jijoye DG75 ne
comp - - D C EB.A - D C E B A comp
[image:7.612.63.559.77.266.2]ne p Jijoye DG75 ne
FIG. 5. Autoradiograms of EMSA using nuclear extracts (ne) of EBV-positive (Jijoye) and EBV-negative (DG75) cells. Free labelled
oligonucleotidesandmajor and minorretardedcomplexes are shown on therightby a starplus the relevant name, by an arrow plus numbers, and
by filledtriangles, respectively. Oligonucleotides Ato Fwere added in 100-foldmolar excess for competition (comp) to binding reactions as
indicatedbeloweachlane.Dashes representreactionsperformed without competingoligonucleotides,and pindicates that nonuclear extract was
added.(A)The labelled123-bpfragment, containing100bpofLMP/TP2promotersequences,revealed threeretardedcomplexes, shown byarrows and numbers. A minorcomplex (filled triangle)possibly represents adegradationproduct of themajor complex 1,sinceit isalso competed for
by oligonucleotide D. Complexes 1 and 3 were competed for by oligonucleotide D, whereas complexes 2 and 3 were competed for by
oligonucleotide C, indicatingthatcomplex3 may represent thebindingofbothcomplexes1and 2 to the labelled123-bp fragment. Oligonucleotide Acompetedforallretarded complexes. (B) Labelled oligonucleotide Cvisualizedone major complex (1C)and four minor complexes (filled triangles). (C) Labelled oligonucleotideDrevealedalso onemajorcomplex(1D)and three minorcomplexes.
changedto arunningtime of 2 hat 130Vand4°C. Addition-ally,theF2-specificcomplexeswerealsospecificallycompeted forbya100-fold molar excess of oligonucleotide 054bp of the
TP1 EBNA2RE(Fig. 6D, lane U)butnotby oligonucleotides C and B. EMSAs performed under low-ionic-strength and low-temperature conditions with anoligonucleotide (position 169721 to 169746) verysimilar to oligonucleotide F revealed thebindingofcellularprotein(s)which could becompeted for by theEBNA2REoftheTP1promoter. EBNA2 could alsonot
bevisualized in these complexes.Additionally,the oligonucle-otide competed for the binding of a cellular factor to the EBNA2RE of theTP1 promoter, though about 25 times less efficiently than the unlabelled TP1 promoter oligonucleotide (datanotshown). Complexes1(withthe123-bpfragment)and 1D (with oligonucleotide D), which were not detected with
HeLa and HaCaT nuclear extracts, were easilyvisualized in EMSAperformed with nuclearextractsmadefromHL60cells, whether dimethylsulfoxide induced or not. Inaddition,three faster-migrating complexes, mostlikely representing degrada-tionproducts stillcontaining theDNA-bindingdomain,were
detected. Specific complexes2and1C(Fig.6AandB),which
were not seenwithHeLaandHaCaT cell nuclear extracts, also couldnotbe detected with HL60 cell nuclearextracts orwith
extracts made from the T-cell line Jurkat (data not shown). These resultsclearlyidentify theDNA-binding activitiesseen
witholigonucleotides C(complex 1C) andD(complex 1D)as B cell andB cellplus HL60cell specific, respectively.
DISCUSSION
It has been suggested that EBNA2 contributes to B-cell immortalizationby itsabilitytotransactivate viral and cellular genes(13). Therefore, it is conceivabletoelucidate the
mech-anism of EBNA2-mediatedtransactivation in orderto under-stand the processofimmortalization at amolecular level.We
chose the bidirectionalLMP/TP2promoterregion for studying
transactivationbyEBNA2. Previousreportshad indicatedthat
LMP, TP1, and TP2expression wastranscriptionally upregu-latedby EBNA2inBurkitt'slymphoma cells (65,70) andthat
promoterCAT reporterplasmids of these geneswere transac-tivated by EBNA2 (22, 70). Additionally, EBNA2REs have
beenroughly delineated for theLMP(22, 60) and CD23 (64) promoters.Onlythe EBNA2REsof theCp (34)and TP1(69) promoterswere analyzed in more detail, providing clear
evi-dence forprotein-DNA interactionsonfunctionally definedcis
sequences.
Theexperiments described here identified theEBNA2RE in
the bidirectional promoter region and characterized protein-DNAinteractions on the delineatedcis-acting elements medi-ating EBNA2 induction. Inafirst step,we wereabletolocate
cis elements between positions -232 to +40 of the LMP
promoter and -104 to +91 of the TP2promoter, conferring EBNA2 responsiveness to the LMP and TP2 promoters, respectively. The data for the LMP promoter are consistent with similar results ofTsangetal. (60).However, the authors did notconsider the transactivation ofthe TP2 promoter by EBNA2. The derived EBNA2REs of the LMP and TP2
promoters overlap by 100 bp as a result of the bidirectional
natureof this promoterregion, which ishighlysuggestive ofa commonEBNA2REforbothpromoters, probablyfunctioning
in coordinate expression of thesetwo latent membrane
pro-teins.
In a second set of experiments, we tested three specific mutations within the 100-bp common cis element in the contextof the whole bidirectional promoter
region.
Theresultspinpointed tworegions(region 2and3)whicharecrucial for
transactivationbyEBNA2of boththe LMPandTP2
promot-ers. Additionally, athird region (region
1)
within the 100-bp sequence isrequiredfor full transactivation ofthe TP2 andto VOL. 68, 1994on November 9, 2019 by guest
http://jvi.asm.org/
6954 LAUX ET AL.
A
*
*
**
&*
,A*.1
z | ~~~~~~~~~~~~~~~~~:;...~ ~
~~~~~~~wq
_W
O-
*
100
I-p
4=1
-
AID
-AOAD!
Al
DV-
IDL
1D|
Ci
A111i
DlCEj
Comp
HeLa
P3HR1 HaCaT
HL60
HL60
Raji
1B4
ne
+DMSO
B
4=1
C
l..
C~
C-
C!I
C
C
C-
CI- C
cC
C-C
COmP
q
x
UK
Z 2 Z 89 :rS
W
CZ) ci] N4)0
!Ce0
Co0 Co~~~~~~~~~(ZCi,4
q~~~~z
ci 0-44C
4
a
*
*
.4
*
4=1D
.
4
40
.4
4
jD-
D!-'Di-
D-
D,-1D-
Dcomp
1
C
as)
zU
27t SPA..
'V. V6t"4'
CT
q-.-.
V,4
"4 C) t .~
zzcAV
r
-
t!F-U
C-IF2UjCj-
jFgIU
Comp
p
Raji
DG75
HaCaT
neFIG. 6. Autoradiogram of EMSA using nuclear extracts (ne) of cervical carcinoma cells (HeLa), promyelocyte cells(HL60),keratinocyte cells
(HaCaT),EBV-negative B cells (BL41 and DG75), andEBV-positive B cells (P3HR-1, Raji, IB4,BL41.P3HR-1, BL41.B95-8, and M-ABA).
Oligonucleotidesaddedfor competition (comp) are shown below each lane by the respective letters. (A) All extracts of B cells revealed the retarded complexes 1 to 3 with the labelled 123-bp fragment and were competed for as in Fig. 5. Nuclear extracts of HeLa and HaCaT cells visualized no specific binding to the 123-bp fragment, whereasHL60extracts contained one binding activitycomigrating with complex 1 as well asseveral faster-migrating complexes (filled triangles). No difference could be observed between extracts ofHL60cells and ofHL60cellstreated
with dimethylsulfoxide (DMSO). (B) One retarded major complex(1C)wasdetected with nuclear extracts of B cells andoligonucleotide C. No
specific complexesweredetected with extracts of non-B cells. Even a very long exposure of the lanes withHL60nuclear extracts did not visualize anyspecificcomplexes. (C) Labelled oligonucleotide D revealed one major retarded complex(1D)withnuclear extracts of B cells. Fourcomplexes wereseen withnuclear extracts ofHL60cells.HeLa and HaCaT nuclear extracts did not show any specific binding activity tooligonucleotide D.
(D) OligonucleotideF2visualized two specific complexes (1 and 2) with nuclear extracts of Raji, DG75, or HaCaT cells which could be competed
for witholigonucleotidesF2and U but not witholigonucleotides C and B.
-J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.68.565.136.597.2]EBNA2-RESPONSIVE ELEMENT OF LMPITP2 PROMOTER REGION 6955
homology
II
cagtgyg
homology
I
yygtgggaa
homology
II
cagtgyg
Cp
AA!&ATC
AxTG A
TI
--
----CGTGC,
ViGMIAL
TP1
ic
M
AG4CCGT
CGAATT4ACD23
=
GAQWIA
+
CTCA
CD48
AL
LMP/
TCCCGG;Gd
GGG
TP2
[image:9.612.56.549.73.353.2](SV40)
Cc
TT
FIG. 7. Similarities between EBNA2-inducibleviral and cellular promoter upstream sequences. The known EBNA2-responsive regions of viral
Cp(-403to-332),TP1(-258to-178), andLMP/TP2 (-232to-131 and -106 to -5) promoters were compared with sequences of the cellular promoters of the CD23(-231to-159) and CD48(-639 to -590) genes. Identical nucleotides in the different sequences are boxed. The dark andlightgreyboxes representhomology regionsIandII, respectively,presentin all five promoters. The bidirectionalLMP/TP2promoter region harbors additional sequences which show strong similarity to simian virus 40 (SV40) sequences containing the binding site for the Spi-1/Pu.1 proto-oncogene product belonging to the Ets family of transcription factors. Region 1 to 3 sequences of the bidirectional LMP/TP2 promoter region mutated in plasmids BA16 to BA21 are represented by numbered black bars.
alesser extentof the LMPpromoterby EBNA2. This
differ-encemight simply reflect the overall weaker induction of the TP2 promoterby EBNA2 compared with the induction of the
LMPpromoter. Bytesting the effect of the EBNA2RE on a heterologous promoter, we could show that regions 2 and 3 were essential for transactivating the heterologous promoter by EBNA2. Theoligonucleotide C (region 1)-binding protein could thus be a promoter factor acting mainly on the TP2 promoter, which does not harbor a classical TATA box. Similarity searches identified a potential Oct-binding site at positions169656to 169665(-87to-96)inthe TP2 promoter (26)next tosequences(homologyregionII) present in
differ-ent EBNA2-responsive promoters (Fig. 7). However,we did
notdetect any competition of the protein-DNA complex 1C either withaclassicalOct-binding siteorwitholigonucleotide
B containing the putative Oct recognition site (data not
shown). This observation suggests that the target DNA
se-quence for this B-cell-specific factormightbe located
down-stream of the putative Oct-binding site relative to the TP2 promoter,which issupportedby DNaseIfootprinting analysis (40, 57).
Ithas been shown (60) that sequences between -234 and -205 are essential for EBNA2 responsiveness on the LMP
promoter butnotsufficient forconferringEBNA2inducibility
to a heterologous promoter. Ourexperiments confirmed and extended these studies by showingthat sequences from -232
to -199 (region 3) are essential for both LMP and TP2
activationby EBNA2. Fahraeus etal. (22), however, claimed
that sequences between -232 and -214are notnecessaryfor induction of theLMPpromoterby EBNA2. Thenatureof this discrepancy isnotclear.Itmight be due tothe use of slightly different promoter deletion constructs with different expres-sionvectorsandtransient transfections into different cells.
Two specific protein-DNA interactions, which are not re-strictedtoB-cell nuclear extracts, were detected with oligonu-cleotide F2, representingDNA-binding activities in the LMP promoter upstreamregion from -253 to -211. This sequence containshomologyregion I, which is mostobviously detected in similarity searches of known viral and cellular promoters respondingtoEBNA2(Fig. 7).A consensussequence,YYGT GGGAA, which is identical to the recently discovered core
recognition sequence of the highly conserved DNA-binding proteinRBP-JK(61),canbe derived. ThemouseRBP-JKbinds
next to the heptamer sequence of JK and appears to be ubiquitously expressed. Itsfunction isnotclear. The available data indicate that RBP-JK may also bind the homology I
sequence presentin the differentEBNA2-responsive
promot-ers(Fig. 7).TheTP1promoterbinds aubiquitously expressed cellular protein(s) interacting with EBNA2 (69). In EMSA withanoligonucleotide containingEBVsequences from posi-tions 169721 to 169746 (numberingofBaer etal. [4]) orwith
oligonucleotide F2 of the LMP/TP2 promoter, we could not detect interactions with EBNA2, but theprotein-DNA com-plexeswerecompeted for byoligonucleotide054bpof theTP1
promoter (Fig.6D, laneU). Additionally, theoligonucleotide (positions 169721 to 169746) competed for the protein-DNA
VOL. 68,1994
on November 9, 2019 by guest
http://jvi.asm.org/
6956 LAUX ET AL.
complex on homology region I (CGTGGGAA) of the TP1 promoter, supporting the possible binding of the same or a related cellular factor(s) on the EBNA2RE of the TP1 and
LMP promoters. The competition was about 25 times less efficient than the competition with the unlabelled TP1 pro-moter oligonucleotide. This finding indicates that homology regions I of the LMP and TP1 promoters probably represent low- and high-affinity binding sites, respectively, of the same cellular protein. The inability to demonstrate the presence of EBNA2 in complexes at the EBNA2RE of the LMPiTP2 promoter region might be due to the less stable interaction of
a protein(s) with sequences containing homology region I of the LMP promoter compared with sequences containing ho-mology region I of the TP1 promoter (oligonucleotide 054bp), since these binding sequences are not completely conserved betweenTP1andLMP/TP2promoters.The factthat there are
two repetitive copies of this sequence present in the TP1 EBNA2RE might also contribute to a more stable complex formation with EBNA2 compared with the single-copy degen-erated sequence of theLMP/TP2 promoter region.
In addition, we identified upstream sequences between -173 and -156of the LMP promoter and -81 to -64 of the TP2 promoter (region 2) which were also essential for the EBNA2 responsiveness of both genes. Oligonucleotide D was used to demonstrate protein-DNA interactions within this region2.Since sequence similarity to simian virus 40 sequences harboring abindingsite for the transcription factor Pu.1/Spi-1 of the Etsfamily(38)wasobserved, itseemsverylikely that an Ets-related transcription factor interacts with oligonucleotide D. We are investigating this possibility by competition and supershift experiments with oligonucleotides containing bind-ing sites for factors of the Ets family and antibodies against thesetranscription factors, respectively.
The importance of cellular factors has been noted for EBNA2-mediated upregulation of cellular genes (17), the Cp promoterdrivingEBNAexpression (34,51,54), and the LMP and TP1 genes (18,69). But it is also possible, depending on the cellularbackground, thatLMPis expressed in the absence ofEBNA2,asrecentlyshown(18, 23) and as seenin nasopha-ryngealcarcinoma cells(7,8, 21,68)and EBV-positive periph-eral angiocentric T-cell lymphomas (48). In these cases, it appears that the LMP expression in the absence of EBNA2 may be regulated by the methylation status of upstream sequences(32, 48).
Our findings suggest that EBNA2 induces LMP and TP2 expression in B cells through interaction with cellular factors binding to the EBNA2RE as has been shown for the TP1 promoter(69).Asimilarmechanism is known to be used by the adenovirus Elaprotein transactivating the viral E4 promoter viainteraction with promoter-bound ATF-2 (43). However, we
were unable to demonstrate the presence of EBNA2 in protein-DNA complexes at the EBNA2RE of the LMP/TP2 promoter region. This could be attributed to conditions not sensitive enough for the detection ofEBNA2 oralternatively
toEBNA2actingat adistance. EBNA2 could affect complexes formedbetweenacellularfactor(s) involved in binding to the
EBNA2REand some control protein(s), thereby activating the transcription factor(s). Such a mechanism is reminiscent of adenovirus Ela protein transactivating the E2 promoter in
some cellsby liberating the cellular transcription factor E2F from complexes with either the retinoblastoma gene product
orcyclinA(11, 50).
In conclusion, the data presented are consistent with a model for transactivation by EBNA2 through an EBNA2RE composed of at least two different specific DNA sequences which bind different cellular proteins. Identification of the
proteins binding to the EBNA2RE is nowrequired as thenext
step in elucidating the mechanism of action of EBNA2 during B-cell immortalization by EBV.
ACKNOWLEDGMENTS
We thank W. Hammerschmidt for providing plasmids p554 and p615 andC. Meitinger forprovidingoligonucleotides B and F.
This study was supported by grants Fa 138/3-4and Fa138/3-7 fromthe
Deutsche Forschungsgemeinschaft (Forschergruppe Virus-Zellwechsel-wirkung) to G.L. and FondsderChemischen IndustrietoG.W.B.
REFERENCES
1. Abbot,S. D.,M. Rowe,K. Cadwallader, A. Ricksten, J. Gordon, F.
Wang, L. Rymo, and A. B. Rickinson. 1990. Epstein-Barrvirus
nuclear antigen 2 induces expression of thevirus-encoded latent
membraneprotein. J.Virol. 64:2126-2134.
2. Alfieri, C., M. Birkenbach, and E. Kieff. 1991. Early events in
Epstein-Barr virus infection of human B lymphocytes. Virology 181:595-608.
3. AIlday, M. J., D. H. Crawford, andB. E.Griffin. 1989.
Epstein-Barr virus latent gene expression during the initiation of Bcell
immortalization. J. Gen.Virol. 70:1755-1764.
4. Baer, R., A. T. Bankier,M. D. Biggin,P. L.Deininger, P. J.Farrell,
T. J. Gibson, G. Hatfull, G. S.Hudson, S. C.Satchwell, C.Seguin,
P.Tuffnell, and B. Barrell. 1984. DNAsequence and expression of
the B95-8Epstein-Barr virus genome. Nature (London)310:207-211. 5. Ben-Bassat, H., N. Goldblum, S. Mitrani, T. Goldblum, J. M. Yoffey, M. M. Cohen, Z. Bentwich, B. Ramot, E. Klein, and G. Klein. 1977. Establishment in continuous cultureof a newtype of
lymphocyte from a "Burkitt like" malignant lymphoma (line D.G.-75). Int.J. Cancer 19:27-33.
6. Boukamp, P., R. T. Petrussevska, D. Breitkreutz, J. Hornung,A. Markham, and N. E.Fusenig. 1988. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 106:761-771.
7. Brooks, L., Q.Y. Yao, A. B. Rickinson, and L. S. Young. 1992. Epstein-Barr virus latent gene transcription in nasopharyngeal carcinoma cells: coexpression of EBNA1, LMP1, and LMP2 transcripts. J. Virol. 66:2689-2697.
8. Busson, P., R. McCoy, R. Sadler, K. Gilligan, T. Tursz, and N. Raab-Traub. 1992.Consistent transcription of theEpstein-Barrvirus
LMP2 gene innasopharyngeal carcinoma. J. Virol. 66:3257-3262. 9. Calender, A., M. Billaud, J. P.Aubry, J.Banchereau, M.
Vuil-laume, and G. M.Lenoir. 1987. Epstein-Barr virus (EBV) induces expression of B-cell activation markers on in vitro infection of EBV-negative B-lymphoma cells. Proc. Natl. Acad. Sci. USA 84: 8060-8064.
10. Cann, A. J., Y.Koyanagi, andI.S. Y. Chen. 1988. High efficiency transfection of primary human lymphocytes and studies ofgene expression. Oncogene 3:123-128.
11. Chellapan, S. P.,S.Hiebert, M.Mudryi,J. M.Horowitz, andJ.R. Nevins.1991. The E2F transcription factor is a cellular targetfor the RBprotein. Cell 65:1053-1061.
12. Cohen, J. I., and E. Kieff. 1991. An Epstein-Barr virus nuclear protein 2domain essential for transformation is a direct transcrip-tionalactivator. J. Virol. 65:5880-5885.
13. Cohen, J. L., F. Wang, and E. Kieff. 1991. Epstein-Barr virus nuclearprotein 2 mutations define essential domains for transfor-mation andtransactivation. J. Virol. 65:2545-2554.
14. Cohen, J. I., F. Wang, J. Mannick,and E. Kieff. 1989. Epstein-Barr virus nuclear protein 2 is a key determinant of lymphocyte transformation. Proc. Natl. Acad. Sci. USA 86:9558-9562. 15. Collins, S.J., R. C. Gallo, and R. E. Gallagher. 1977. Continuous
growth anddifferentiation of human myeloid leukaemic cells in suspension culture.Nature (London) 270:347-349.
16. Cordier, M., A.Calender, M. Billaud, U. Zimber, G. Rousselet,0.
Pavlish, J. Banchereau, T. Tursz, G. Bornkamm, and G. M. Lenoir. 1990. Stable transfection of Epstein-Barr virus (EBV) nuclearantigen 2 in lymphoma cells containing the EBV P3HR1 J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
EBNA2-RESPONSIVE ELEMENT OF LMP/TP2 PROMOTER REGION 6957
genome induces expression of B-cell activation molecules CD21
and CD23. J. Virol. 64:1002-1013.
17. Cordier-Bussat, M., M. Billaud, A. Calender, and G. M. Lenoir.
1993. Epstein-Barr virus (EBV) nuclear-antigen-2-induced up-regulation of CD21 and CD23 molecules is dependent on a permissivecellular context. Int. J. Cancer 53:153-160.
18. Cordier-Bussat, M., A. Calender, M. Vuillaume, G. W.
Born-kamm, and G. M.Lenoir. 1993. Expression of the Epstein-Barr virus (EBV) latent membrane protein is tightly regulated, inde-pendentlyofEBnuclear antigen 2 and of EBV integration or copy
number. Virus Res.27:55-69.
19. Crawford, D.H., M. A. Epstein, G. W. Bornkamm, B. G. Achong,
S.Finerty, and J. L.Thompson. 1979. Biological and biochemical
observations onisolates of EB virus from the malignant epithelial
cells of twonasopharyngeal carcinomas. Int. J. Cancer 24:294-302.
20. Dignam, J. D., R. M.Lebovitz, and R. G. Roeder. 1983. Accurate transcription initiation by RNA polymeraseII in a soluble extract
fromisolatedmammalian nuclei. Nucleic Acids Res. 11:1475-1489.
21. Fahraeus, R., H. L. Fu,I. Ernberg, J. Finke, M. Rowe, G. Klein,K.
Falk, E. Nilsson, M. Yadav, P. Busson, T. Turz, and B. Kallin. 1988.Expression ofEpstein-Barr virus-encoded proteins in naso-pharyngeal carcinoma. Int. J. Cancer 42:329-338.
22. Fahraeus,R, A.Jansson, A. Ricksten, A.Sjoblom, and L. Rymo. 1990. Epstein-Barr virus-encoded nuclear antigen 2 activates the virallatentmembraneproteinpromoter by modulating the activity
of anegativeregulatory element. Proc. Natl. Acad. Sci. USA 87: 7390-7394.
23. Fahraeus, R, A. Jansson, A. Sjoblom, T. Nilsson, G. Klein, and L. Rymo. 1993. Cell phenotype-dependent control of Epstein-Barr
virus latent membrane protein 1 gene regulatory sequences. Virology195:71-80.
24. Fennewald, S., V. Van-Santen, and E. Kieff. 1984. Nucleotide sequence of an mRNAtranscribed in latent growth-transforming
virusinfection indicatesthat itmayencode a membrane protein. J.
Virol.51:411-419.
25. Gey, G., W. Coffman, and M. Kubiceck. 1952. Tissue culture studies of the proliferative capacity of cervical carcinoma and
normalepithelium. CancerRes. 12:264-271.
26. Ghosh, D., and E. Kieff. 1990.cis-acting regulatory elements near the Epstein-Barr virus latent-infection membrane protein tran-scriptional start site. J.Virol. 64:1855-1858.
27. Grasser, F. A., S. Gottel, P. Haiss, B. Boldyreff,0.G. Issinger, and N.Mueller-Lantzsch. 1992. Phosphorylation of the Epstein-Barr virus nuclear antigen 2. Biochem. Biophys. Res. Commun. 186:
1694-1701.
28. Grasser, F. A., P.Haiss, S. Gottel, and N. Mueller-Lantzsch. 1991.
Biochemical characterization ofEpstein-Barr virus nuclear
anti-gen 2A.J.Virol. 65:3779-3788.
29. Hammerschmidt, W., and B. Sugden. 1989. Genetic analysis of immortalizingfunctionsofEpstein-Barr virus in human B
lympho-cytes. Nature (London)340:393-397.
30. Hinuma, Y., M.Konn, J. Yamaguchi, D. Wudarski, J. Blakeselee, and J. Grace. 1967. Immunofluorescence and herpes-type virus particles in the P3HR-1 Burkitt lymphoma cell line. J. Virol. 1:1045-1051.
31. Horvath, G. C., and W. H.Schubach. 1993. Identification of the Epstein-Barrvirusnuclear antigen 2transactivation domain.
Bio-chem. Biophys. Res. Commun. 191:196-200.
32. Hu, L. F., J.Minarovits,S. L. Cao, B. Contreras-Salazar, L. Rymo, K.Falk, G. Klein, and L. Ernberg. 1991. Variable expression of latent membrane protein in nasopharyngeal carcinoma can be related to methylation status of the Epstein-Barr virus BNLF-1
5'-flanking region. J. Virol.65:1558-1567.
33. Hudson, G. S., P. J.Farrell, and B. G.Barrell.1985. Two related but differentiallyexpressed potential membrane proteins encoded bytheEcoRI Dhetregion ofEpstein-Barr virus B95-8. J. Virol.
53:528-535.
34. Jin, X. W., and S. H. Speck. 1992. Identification of critical cis
elementsinvolved inmediatingEpstein-Barr virus nuclear antigen
2-dependent activity of anenhancer located upstream of the viral BamHICpromoter.J.Virol.66:2846-2852.
35. Kaye, K. M., K. M. Izumi, and E. Kieff. 1993. Epstein-Barr virus
latent membrane protein 1 isessential for B- lymphocyte growth
transformation. Proc. Natl. Acad. Sci. USA 90:9150-9154.
36. Kieff, E., and D. Liebowitz. 1990.
Epstein-Barr
virus and its replication, p. 1889-1920. In B. N. Fields, D. M. Knipe, R. M. Chanock, M. S. Hirsch, J. L. Melnick, T. P. Monath, and B. Roizman(ed.), Virology. Raven PressLtd., NewYork.37. King, W., A. L. Thomas-Powell, N. Raab-Traub, M. Hawke, and E.
Kief. 1980. Epstein-Barr virus RNA. V. Viral RNA in a restrin-gently infected, growth-transformed cell line.J.Virol.36:506-518. 38. Klemsz, M. J., S. R McKercher, A. Celada, C. Van Beveren, and R A. Maki. 1990. The macrophage and B cell-specific
transcrip-tion factor
PU.1
is related to the ets oncogene. Cell 61:113-124. 39. Knutson, J. C. 1990. The level of c-fgr RNA is increased byEBNA-2, an Epstein-Barr virus gene required for B-cell
immor-talization. J. Virol. 64:2530-2536.
39a.Laux, G., et al. Unpublisheddata.
40. Laux, G., F. Dugrillon, C. Eckert, U. Zimber-Strobl, and G. W.
Bornkamm. 1993. Delineation of the cis-acting sequence mediat-ing transactivation of a bi-directional latent promoter region by
Epstein-Barr virus nuclear antigen 2 (EBNA2), p. 211-217. In T.
Tursz, J. S. Pagano, D. V. Ablashi, G. de The, G. Lenoir, and G. R.
Pearson (ed.), The Epstein-Barr virus and associated diseases. Colloque INSERM/John Libbey Eurotext Ltd., Paris/London.
41. Laux, G., A. Economou, and P. J. Farrell. 1989. The terminal protein gene 2of Epstein-Barr virus is transcribed from a
bidirec-tional latent promoter region. J. Gen. Virol. 70:3079-3084.
42. Lenoir, G. M., M. Vuillaume, and C. Bonnardel. 1985. The use of
lymphomatous and lymphoblastoid cell lines in the study of Burkitt's lymphoma. IARC Sci. Publ. 60:309-318.
43. Lin, F., and M. Green. 1990. A specific member of the ATF transcription factor family can mediate transcription activation by the adenovirus Ela protein. Cell 61:1217-1224.
44. Ling, P. D., D. R Rawlins, and S. D. Hayward. 1993. The
Epstein-Barr virus immortalizing protein EBNA-2 is targeted to DNA by a cellular enhancer-binding protein. Proc. Natl. Acad. Sci. USA 90:9237-9241.
45. Maniatis, T., E. F. Fritsch, and J.
Sambrook.
1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Press,Cold Spring Harbor, N.Y.
46. Miller, G. 1990. Epstein-Barr virus, p. 1921-1958. In B. N. Fields, D. M. Knipe, R. M. Chanock, M. S. Hirsch, J. L. Melnick, T. P. Monath, and B. Roizman (ed.), Virology. Raven Press, New York. 47. Miller, G., J. Robinson, L. Heston, and M. Lipman. 1974. Differ-ences between laboratory strains of Epstein-Barr virus based on immortalization, abortive infection, and interference. Proc.
Natl.
Acad. Sci. USA 71:4006-4010.48. Minarovits, J., L.-F. Hu, S. Imai,Y. Harabuchi, A. Kataura, S. Minarovits-Kormuta, T. Osato, and G. Klein. 1994. Clonality, expression and methylation
patterns
of the Epstein-Barr virus genomes in lethal midline granulomas classified as peripheral angiocentric T cell lymphomas. J. Gen. Virol. 75:77-84. 49. Moss, D. J., T. B. Sculley, and J. H. Pope. 1986. Induction ofEpstein-Barr virus nuclear antigens. J. Virol. 58:988-990. 50. Mudryi,M., S. H. Devoto, S. W.
Hiebert,
T. Hunter, J. Pines, andJ. R Nevins. 1991. Cell cycle regulation of the E2F transcription factor involves an interaction with cyclin A. Cell 65:1243-1253. 51. Nilsson, T.,A. Sjoblom, M. G. Masucci, and L. Rymo. 1993. Viral
and cellular factors influence the activity of the Epstein-Barr virus BCR2 and BWR1 promoters in cells of different phenotype. Virology 193:774-785.
52. Pulvertaft,RJ. V. 1965. A study of malignant tumours in Nigeria by shorttermtissue culture. J. Clin. Pathol. 18:261-273. 53. Rooney, C., J. G. Howe, S. H. Speck, and G. Miller. 1989.Influence
of Burkitt's lymphoma and primary B cells on latent gene expres-sion by the nonimmortalizing P3J-HR-1 strain of Epstein-Barr virus. J. Virol. 63:1531-1539.
54. Rooney, C. M., M. Brimmell, M. Buschle, G. Allan, P. J. Farrell, and J. L. Kolman. 1992. Host cell and EBNA-2 regulation of Epstein-Barr virus latent-cycle promoter activity in B lymphocytes. J. Virol. 66:496-504.
55. Sample, J., D. Liebowitz, and E.Kieff.1989. Two related Epstein-Barr virus membrane proteins are encoded by separate genes. J. Virol. 63:933-937.
56. Schwenk, H. U., and U. Schneider. 1975. Cell cycle dependency of
VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
6958 LAUX ET AL.
aT-cell marker on lymphoblasts. Blut 31:299-306.
57. Sjoblom,A., A. Jansson, T.Nilsson, S. Lain, and L. Rymo.1993. On the mechanism of transcriptional activation of the LMP1 promoterby EBNA2, p.153-158.In T.Tursz, J.S. Pagano, D. V.
Ablashi, G. de The, G. Lenoir, and G. R. Pearson (ed.), The
Epstein-Barr virus and associated diseases. Colloque INSERM/ JohnLibbeyEurotextLtd., Paris/London.
58. Sung, N. S., S. Kenney, D. Gutsch, and J. S. Pagano. 1991. EBNA-2 transactivates a lymphoid-specific enhancer in the
BamHI C promoterofEpstein-Barrvirus. J.Virol.65:2164-2169.
59. Tomkinson, B., E. Robertson, and E. Kieff. 1993. Epstein-Barr virus nuclear proteins EBNA-3Aand EBNA-3C are essential for
B-lymphocytegrowth transformation. J.Virol.67:2014-2025. 60. Tsang, S. F., F. Wang, K M. Izumi, and E. Kief. 1991. Delineation of
thecis-acting element mediatingEBNA-2 transactivation of latent
infectionmembraneprotein expression.J. Virol. 65:6765-6771. 61. Tun, T., Y. Hamaguchi, N. Matsunami, T.Furukawa,T.Hono, and
M. Kawaichi. 1994. Recognition sequence ofa highly conserved
DNAbindingprotein RBP-Jk.Nucleic AcidsRes.22:965-971.
62. Wang, F., C. D.Gregory,M.Rowe, A. B.Rickinson,D.Wang, M. Birkenbach, H. Kikutani, T. Kishimoto, and E. Kief. 1987.
Epstein-Barr virus nuclear antigen2specifically induces
expres-sion of the B-cell activationantigen CD23.Proc.Natl. Acad. Sci. USA 84:3452-3456.
63. Wang, F.,C. D. Gregory, C. Sample, M. Rowe, D.Liebowitz, R. Murray, A. B. Rickinson, and E. Kieff. 1990. Epstein-Barr virus
latent membrane protein(LMP1) andnuclearproteins 2and3C are effectors ofphenotypic changes in B lymphocytes: EBNA-2
andLMP1cooperatively induce CD23. J. Virol.64:2309-2318.
64. Wang, F., H.Kikutani, S. F. Tsang, T. Kishimoto, and E. Kief. 1991. Epstein-Barrvirus nuclear protein 2 transactivates a
cis-acting CD23DNAelement. J. Virol. 65:4101-4106.
65. Wang, F., S. F. Tsang, M. G. Kurilla, J. I. Cohen, and E. Kieff. 1990. Epstein-Barr virus nuclear antigen 2 transactivates latent
membrane proteinLMP1. J.Virol. 64:3407-3416.
66. Westin, G., T. Gerster, M. M. Muller, G. Schaffner, and W. Schaffner. 1987.OVEC,aversatilesystem tostudy transcriptionin mammalian cells and cell-free extracts. Nucleic Acids Res. 15: 6787-6789.
67. Woisetschlaeger, M., X. W. Jin, C. N. Yandava, L. A. Furmanski, J. L.Strominger, and S. H.Speck.1991. Role for theEpstein-Barr
virus nuclear antigen 2 in viral promoter switching duringinitial
stagesofinfection. Proc. Natl. Acad.Sci. USA88:3942-3946. 68. Young, L.S., C. W. Dawson, D.Clark,H.Rupani,P.Busson, T.
Tursz, A.Johnson, and A. B. Rickinson. 1988.Epstein-Barr virus
gene expression in nasopharyngeal carcinoma. J. Gen. Virol.
69:1051-1065.
69. Zimber-Strobl, U., E. Kremmer, F. Grasser, G. Marschall, G. Laux, andG. W. Bornkamm.1993.The Epstein-Barrvirus nuclear antigen2interacts withanEBNA2responsive cis-element of the terminalprotein 1gene promoter. EMBOJ. 12:167-175. 70. Zimber-Strobi, U., K 0. Suentzenich, G. Laux, D. Eick, M.
Cordier, A. Calender, M. Billaud, G. M. Lenoir, and G. W. Bornkamm. 1991. Epstein-Barrvirusnuclear antigen 2activates transcription oftheterminalproteingene.J. Virol. 65:415-423.
J. VIROL.