0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.14.9197–9205.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
In Vitro Suppression of Human Immunodeficiency Virus
Type 1 Replication by Measles Virus
Mayra Garcı´a,
1,2Xiao-Fang Yu,
1Diane E. Griffin,
1and William J. Moss
1,3*
W. Harry Feinstone Department of Molecular Microbiology and Immunology, Bloomberg School of Public Health,1
Cellular and Molecular Medicine, School of Medicine,2and Department of Epidemiology, Bloomberg
School of Public Health,3Johns Hopkins University, Baltimore, Maryland
Received 18 January 2005/Accepted 27 March 2005
During the acute phase of measles, human immunodeficiency virus type 1 (HIV-1)-infected children have a transient, but dramatic, decrease in plasma HIV-1 RNA levels (W. J. Moss, J. J. Ryon, M. Monze, F. Cutts, T. C. Quinn, and D. E. Griffin, J. Infect. Dis. 185:1035–1042, 2002). To determine the mechanism(s) by which coinfection with measles virus (MV) decreases HIV-1 replication, we established an in vitro culture system that reproduces this effect. The addition of MV to CCR5- or CXCR4-tropic HIV-1-infected human peripheral blood mononuclear cells (PBMCs) decreased HIV-1 p24 antigen production in a dose-dependent manner. This decrease occurred with the addition of MV before or after HIV-1. The inhibition of HIV-1 p24 antigen production was decreased when UV-inactivated MV or virus-free supernatant fluid from MV-infected PBMCs was used. Inhibition was not due to increased production of chemokines known to block coreceptor usage by HIV-1, a decrease in the percentage of CD4ⴙT cells, or a decrease in chemokine receptor expression by CD4ⴙ T cells. Viability of PBMCs was decreased only 10 to 20% by MV coinfection; however, lymphocyte proliferation was decreased by 60 to 90% and correlated with decreased production of p24 antigen. These studies showed that an in vitro system of coinfected PBMCs could be used to dissect the mechanism(s) by which MV suppresses HIV-1 replication in coinfected children and suggest that inhibition of lymphocyte proliferation by MV may play a role in the suppression of HIV-1 p24 antigen production.
Measles virus (MV) and human immunodeficiency virus type 1 (HIV-1) infection are important causes of morbidity and mortality in many parts of the world. Both viruses induce immunosuppression, and disease is associated with an in-creased susceptibility to secondary infections. In individuals infected with HIV-1, coinfections and vaccinations are often associated with transient increases in plasma HIV-1 RNA lev-els (26), presumably due to an increase in the number of activated CD4⫹T-lymphocyte target cells and the induction of proinflammatory cytokines that favor HIV-1 replication.
Surprisingly, MV infection, which results in both immune activation and induction of proinflammatory cytokines (12), is associated with a transient but profound reduction in plasma levels of HIV-1 in coinfected children (19). Decreased plasma HIV-1 RNA levels also have been reported following coinfec-tion with Orientia tsutsugamushi (29), the etiologic agent of scrub typhus, and dengue virus (30) and with chronic infection with GB virus C (GBV-C) (32). The mechanisms for in vivo reduction in HIV-1 replication by these various pathogens have not been identified.
Several viruses inhibit HIV-1 replication in vitro. Superna-tant fluids from peripheral blood mononuclear cells (PBMCs) stimulated with live or inactivated influenza A virus inhibit replication of CCR5- and CXCR4-tropic HIV-1 isolates, an effect partially mediated by alpha interferon (22). Human her-pesvirus 6 inhibits replication of CCR5-tropic but not
CXCR-4-tropic HIV-1 in human lymphoid tissue, an effect mediated by RANTES (13). Similarly, GBV-C was shown to inhibit HIV-1 replication and induce chemokine production by PB-MCs in vitro (35). Studies of MV-coinfected children suggest that the decrease in plasma HIV RNA levels is not due to a reduction in the number of CD4⫹T lymphocytes but that an increase in soluble suppressive factors, such as-chemokines, may contribute to the inhibition of HIV-1 replication (19). To better understand the reduction in plasma HIV-1 levels in children with measles, we developed an in vitro system to investigate the mechanisms by which MV suppresses HIV-1 replication.
MATERIALS AND METHODS
Cells.Peripheral blood samples were obtained from healthy, anonymous do-nors through the blood bank of the Johns Hopkins Hospital. PBMCs were separated by Ficoll-Hypaque (Pharmacia, Uppsala, Sweden) density gradient centrifugation and cultured in RPMI 1640 supplemented with 20% fetal bovine serum (FBS) and 10 U/ml interleukin 2 (IL-2) (ICN, Aurora, Ohio) (RPMI–20% FBS–IL-2). Vero cells were obtained from the American Type Culture Collec-tion (Manassas, VA).
Viruses and controls.The Chicago-1 and Edmonston strains of MV were obtained from lysates of infected Vero cells and assayed by plaque formation on Vero cells. MV stocks and Vero cell lysates were prepared by freezing
MV-infected or unMV-infected Vero cells at⫺80°C for at least 2 hours and thawing at
37°C. The lysates were pelleted at 12,000 rpm for 5 min, passed through a
0.22-m filter, and frozen in aliquots at⫺80°C. The Chicago-1 strain was isolated
in 1989 during a measles outbreak in Chicago, Illinois, and has been classified as genotype group D3 (33). The wild-type Edmonston strain (American Type Cul-ture Collection) is a minimally passaged derivative of the original genotype A Edmonston strain of MV isolated by Enders and Peebles (11). MV stocks and
Vero cell lysate controls were tested for lipopolysaccharide (LPS) by theLimulus
amebocyte lysate assay QCL-1000 Chromogenic LAL (BioWhittaker, Walkers-ville, MD). The concentration of LPS was 0.01 to 0.04 ng/ml for Chicago-1 MV, 0.005 to 0.0005 ng/ml for Edmonston MV, and 0.008 ng/ml for Vero cell lysates.
* Corresponding author. Mailing address: W. Harry Feinstone De-partment of Molecular Microbiology and Immunology, Johns Hopkins Bloomberg School of Public Health, 615 N. Wolfe St., Baltimore, MD 21205. Phone: (410) 955-3859. Fax: (410) 955-0105. E-mail: wmoss @jhsph.edu.
9197
on November 8, 2019 by guest
http://jvi.asm.org/
For some experiments, MV was inactivated with 3,000 W/cm2
UV radiation for 2 min. Free radicals were allowed to decay for at least 1 week prior to use. Inactivation was confirmed by culture on Vero cells.
Stocks of HIV-1BaL, a CCR5-tropic laboratory strain, were grown on PBMCs
obtained from healthy donors that were cultured for 10 days in RPMI–20%
FBS–IL-2. Stocks of HIV-1IIIB, a CXCR4-tropic laboratory strain, were obtained
from chronically infected MOLT-3 cells. HIV-1-infected cells were pelleted at
3,000 rpm for 20 min, and supernatant fluid was passed through a 0.2-m filter
and frozen in aliquots at⫺80°C. The titers of stocks were determined using TZM
cells (9).
HIV-1 and MV infection of PBMCs. HIV-1Ba-Lwas incubated with 10 7
PBMCs/ml in RPMI–20% FBS–IL-2 at a multiplicity of infection (MOI) of
0.00005 for 48 h. For HIV-1IIIB, PBMCs were stimulated with 2g/ml of
phy-tohemagglutinin (PHA-P; Sigma, St. Louis, MO) for 24 h in RPMI–20% FBS–
IL-2 with 1⫻glutamine, 100 U/ml penicillin, and 100g/ml streptomycin before
the addition of HIV-1IIIBat a MOI of 0.0004 for 24 h in RPMI–20% FBS–IL-2.
Serial 10-fold dilutions of MV were added to 2⫻106
PBMCs infected with
HIV-1BaLor HIV-1IIIB, starting with 3⫻106PFU/ml of Chicago-1 MV (MOI of
1.5) or 3⫻107
PFU/ml of Edmonston MV (MOI of 15) (Fig. 1). Lysates of uninfected Vero cells and media alone served as controls. For later experiments, only a single MOI (0.15) of MV was used. Vero cell lysates were diluted to correspond with the dilution of the MV stock. To assess the impact of the relative timing of MV and HIV-1 infection, MV was added 3 h before, concurrently, or
15 h after HIV-1 infection. For these experiments, HIV-1BaLwas used at a MOI
of 0.00007.
Supernatant fluid was harvested at 4, 7, and 12 days after the addition of MV
(corresponding to 6, 9, and 14 total days in culture) and assayed for HIV-1 p24 antigen by enzyme immunoassay (EIA) (NEN Life Science Products, Boston, MA) (Fig. 1). Control samples consisted of supernatant fluids from uninfected PBMCs (negative control) and from PBMCs infected with HIV-1 alone (positive
control). Percent inhibition was calculated as follows: 100⫺[(concentration of
p24 antigen in the test sample⫻100)/(concentration of p24 antigen in the
positive-control sample)].
Supernatant fluid from PBMCs infected with MV.PBMCs (2⫻106
) were
incubated with 3⫻105PFU of Chicago-1 MV or Vero cell lysate (1:10) in
RPMI–10% FBS (Fig. 1). Supernatant fluid was collected at 24, 48, and 72 h after
infection and added to PBMCs infected with HIV-1BaLor HIVIIIB. Supernatant
fluid obtained at 24 and 72 h was passed through a 0.1-m filter, and the absence
of MV was confirmed on Vero cells. After 96 h, supernatant fluid from the coculture was collected and assayed for HIV-1 p24 antigen by EIA.
Cell viability.Cell viability was determined by propidium iodide (PI) exclusion or staining with 7-amino actinomycin D (7AAD) (21) 4 days after the addition of MV to HIV-infected PBMCs. Cells were incubated on ice with a 1:5,000 dilution of PI for 10 min and analyzed by flow cytometry. For experiments involving costaining for CD4, CD8, and CXCR4, 7AAD (1:100) was used. Cell death attributed specifically to MV was calculated as the percentage of coinfected,
PI-positive cells⫺the percentage of HIV-1-infected, PI-positive cells.
Cell surface expression of CD4, CD8, CCR5, and CXCR4.PBMCs harvested 4 days after the addition of MV to HIV-infected cells were studied. Cells were
incubated with a 1:5 dilution of human serum, washed with 1⫻
phosphate-buffered saline supplemented with 2% FBS, stained with fluorescein isothiocya-nate-conjugated mouse monoclonal antibodies against human CD4 and CD8, phycoerythrin-conjugated mouse monoclonal antibodies against human CXCR4 and CCR5, or isotype control antibodies (Pharmingen, BD Biosciences, San Diego, CA), washed, and fixed with BD Cytofix/Cytoperm (Pharmingen, BD Biosciences, San Diego, CA). Cells were analyzed on a FACS Calibur using Cell Quest software (Becton Dickinson).
Cytokines and chemokines.The levels of RANTES, stromal-cell-derived fac-tor 1 (SDF-1) (R&D, Minneapolis, MN), and macrophage inflammafac-tory protein
1␣(MIP-1␣) and MIP-1(Biosource International, Camarillo, CA) in
superna-tant fluids from HIV-1-infected PBMCs cocultured with MV or Vero cell lysates were measured by EIA according to the manufacturer’s instructions. To assess
the effects of-chemokines in the culture on HIV-1 p24 antigen production,
neutralizing antibodies to RANTES, MIP-1␣, and MIP-1(R&D, Minneapolis,
MN) were added at 0.5, 1, or 5g/ml to coinfected PBMCs. Isotype control
antibodies or media alone were used as controls. After 96 h, p24 antigen was measured by EIA.
Cell depletion experiments.PBMCs were stimulated with Chicago-1 and
Ed-monston MV strains after removal of CD14⫹cells. PBMCs were incubated with
microbead-labeled antibody to CD14 (MACS; Miltenyi Biotec, Auburn, CA),
resuspended, and passed over a magnetic column. CD14⫹cells remained bound
to the column, and the CD14⫺cells were collected. Cell purity was confirmed by
flow cytometry. The depleted cell population was infected with MV and HIV-1, and p24 antigen was measured after 96 h.
Lymphoproliferation assays.HIV-1-infected PBMCs were plated in triplicate
in 96-well round-bottom plates at a concentration of 105
cells/well in 100l of
RPMI–20% FBS–IL-2. Four test samples were added to the wells: (i) Chicago-1 MV at a MOI of 1.5, 0.15, and 0.015; (ii) Edmonston MV at a MOI of 15, 1.5, and 0.15; (iii) Vero cell lysate appropriately diluted; and (iv) media. The cells
were incubated at 37°C for 72 h, and 1Ci/well (10l) of [3
H]thymidine was added overnight. DNA was harvested onto filter paper and read in a scintillation counter. The reduction in lymphoproliferation attributed to MV was calculated
as follows: 100⫺(cpm of the test samples⫻100)/(cpm of HIV-infected cells),
where cpm is counts per minute.
Statistical analysis.Paired comparisons were made with the Wilcoxon signed-rank test. The relationship between numerical variables was measured by esti-mating the correlation coefficient, with confidence intervals calculated on the basis of Fisher’s transformation. Statistical analyses were conducted using Stata statistical software release 7.0 (Stata Corporation, College Station, TX).
RESULTS
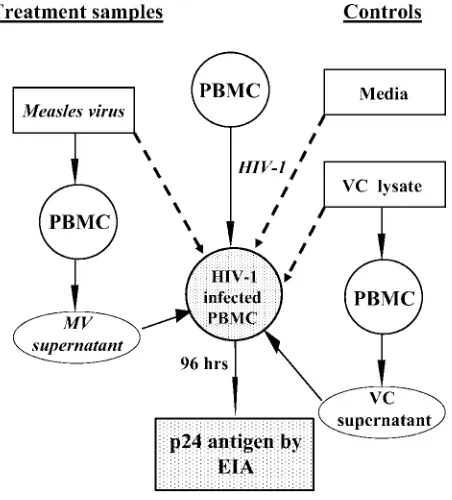
[image:2.585.54.280.68.318.2]Reduction in HIV-1 p24 antigen production following the addition of MV.The addition of MV to CCR5- and CXCR4-tropic HIV-1-infected PBMCs resulted in a dose-dependent decrease in p24 antigen production after 96 h of coculture (Fig. 2A and B). A 10-fold-higher concentration of the Edmonston FIG. 1. Scheme for the in vitro evaluation of interactions between
measles virus and HIV-1. MV, Vero cell (VC) lysate, or media were added to HIV-1-infected cells and incubated for 96 h. Alternatively, PBMCs were incubated with MV or VC lysate for 24, 48, or 72 h, and supernatant fluids were collected and incubated with HIV-1-infected PBMCs for 96 h. The levels of p24 antigen were determined by enzyme immunoassay. For HIV-1 infection with CCR5-tropic HIV-1BaL,
PBMCs were infected for 48 h in the presence of 10 U IL-2/ml. For CXCR4-tropic HIV-1IIIB, PBMCs were stimulated with 10 U IL-2/ml
and 2g phytohemagglutinin/ml for 24 h before the addition of HIV-1. The medium was removed, and the cells were infected for 24 h in the presence of 10 U/ml of IL-2.
9198 GARCI´A ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
strain of MV was required to achieve a decrease in p24 antigen production comparable to that of the Chicago-1 strain. When MV was added at a MOI lower than 0.015, the decrease in p24 antigen production was not significant and was similar to that observed with lysates of uninfected Vero cells. The reduction in p24 antigen production persisted for at least 12 days of coculture (Fig. 2C and D). Inhibition of HIV-1 replication was independent of the relative timing of the addition of MV. Reduction in p24 antigen production by PBMCs infected with
HIV-1BaL(Fig. 3A) or HIV-1IIIB(Fig. 3B) was observed when
MV was added 3 h before HIV-1, concurrently with HIV-1, or 15 h after HIV-1. Similar reductions in p24 antigen production were observed using cell populations depleted of CD14⫹cells. The reductions in p24 antigen production by PMBCs infected with HIV-1BaLwere 83% for Chicago-1 and 59% for
Edmon-ston MV strains. For HIV-1IIIB, the reductions in p24 antigen
production by Chicago-1 and Edmonston MV strains were 78% and 44%, respectively.
FIG. 2. Effect of MV on HIV-1 p24 antigen production. Effects of different concentrations of MV on p24 antigen (Ag) production by CCR5-tropic HIV-1BaL-infected PBMCs (A) or CXCR4-tropic HIV-1IIIB-infected PBMCs (B) using Chicago-1 or Edmonston MV at a MOI of
15 or 1.5 to 0.00015 and Vero cell lysates at dilutions comparable to those used for MV. Data are presented as means⫾standard errors of the means (SEM) (error bars) of duplicate wells with PBMCs from 4 to 13 donors. Values that are significantly different (P⬍0.05) between MV and Vero cell lysates are indicated by an asterisk. ND, not done. (C and D) Time course of p24 antigen production by HIV-1-infected PBMCs after the addition of MV. PBMCs were infected as described above with CCR5-tropic HIV-1BaL(C) or CXCR4-tropic HIV-1IIIB(D) and Chicago-1 and
Edmonston MV at a MOI of 0.15. Vero cell lysates (1:10) served as control. Data are presented as means⫾SEM (error bars) of duplicate wells with PBMCs from 7 to 11 donors. Values that are significantly different (P⬍0.05) between MV and media are indicated by an asterisk. Ag conc., antigen concentration.
on November 8, 2019 by guest
http://jvi.asm.org/
MV replication and inactivation.To determine whether MV replicated in the presence of HIV-1, MV in supernatant fluids was assayed (Fig. 4). MV replicated in HIV-infected PBMCs, and virus production 4 days after infection was highest at an inoculating MOI of 0.015 or 0.0015 (Fig. 4A and B). Replica-tion of Edmonston MV was maintained over 12 days in cell culture, but the amounts of Chicago-1 MV declined 10- to
100-fold (Fig. 4C and D). To assess whether replicating MV was necessary for the reduction in p24 antigen production, the experiments were repeated using UV-inactivated MV. Inacti-vated Chicago-1 and Edmonston MV reduced p24 antigen production by PBMCs infected with HIV-1BaLand HIV-1IIIB
but to a lesser extent than live virus did (Fig. 4E and F).
Effect of MV on cell viability.To assess whether the reduc-tion in p24 antigen producreduc-tion following the addireduc-tion of MV was due to a decrease in the number of cells capable of sup-porting HIV-1 replication, the viability of coinfected PBMCs was measured (Fig. 5). Death due to HIV infection (range, 3 to 9%) was subtracted to determine excess cell death due to MV. The percentage of cells that failed to exclude PI due to MV increased with the MV MOI and was higher for the Chicago-1 strain than for the Edmonston strain (Fig. 5A and B), although these differences were not statistically significant. The percent-age of PI-staining cells ranged from 0 to 17% for the MV MOI used in most experiments (0.15).
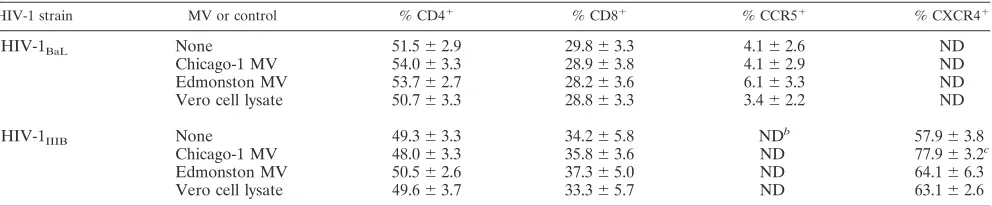
Effects of MV on cell surface expression of CD4, CCR5, and CXCR4.To determine whether MV coinfection decreased the number of target cells for HIV-1 replication by reducing avail-ability of the HIV-1 cellular receptors, surface expression of CD4, CCR5, and CXCR4 was assessed by flow cytometry (Ta-ble 1). The proportion of CD4⫹ and CD8⫹ cells was not altered by coinfection with MV, and cell surface expression of the HIV coreceptors CCR5 and CXC4 did not decrease. In fact, surface expression of CXCR4 increased after the addition of Chicago-1 MV to PBMCs infected with HIV-1IIIB(Table 1).
The number of nonviable cells expressing the surface markers CD4, CD8, and CXCR4 was assessed by costaining for 7AAD and did not demonstrate preferential death of a specific lym-phocyte subset (data not shown).
Effect of MV on lymphoproliferation.Although the addition of MV did not result in the loss of potential target cells, the ability of cells to proliferate could be impaired in the presence of MV (14, 28). MV inhibited the proliferation of PBMCs infected with HIV-1 as measured by incorporation of [3
H]thy-midine (Fig. 6). Inhibition increased with the MV MOI and was greater with the Chicago-1 strain than with the Edmonston strain. The reduction in p24 antigen production was strongly correlated with the extent to which lymphoproliferation was inhibited when the mean values for the two MV strains (at four different MOIs) and the two HIV-1 strains were compared (r
⫽0.72; 95% confidence interval [CI], 0.35 to 0.90). The cor-relation between p24 antigen production and lymphoprolifera-tion was strongest for Chicago-1 MV (r⫽0.91; 95% CI, 0.55 to 0.98) and weakest for CXCR4-tropic HIV-1 (r⫽0.57; 95% CI, 0.23 to 0.91).
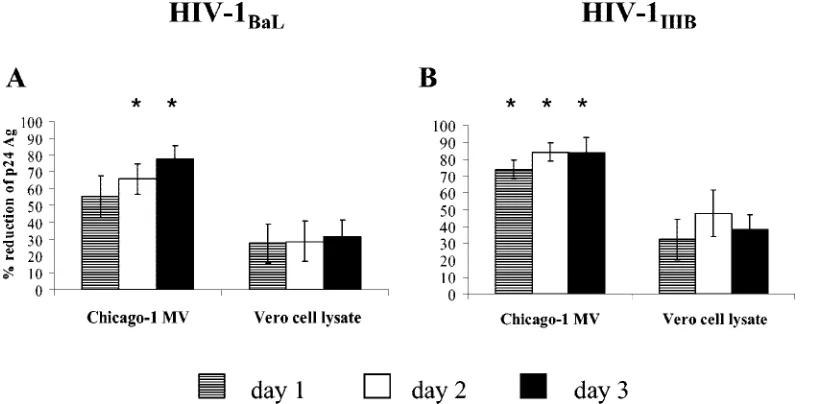
Effects of supernatant fluids from MV-infected PBMCs on HIV-1 p24 antigen production.To assess whether soluble fac-tors produced by MV-infected PMBCs inhibit HIV-1 replica-tion, supernatant fluids were harvested from PBMCs infected with Chicago-1 MV (MOI of 0.15) at 1, 2, and 3 days following infection and added to PBMCs infected with HIV-1BaL or
HIVIIIB(Fig. 7). Supernatant fluids from MV-infected PBMCs
reduced p24 antigen production by PBMCs infected with HIV-1BaL(Fig. 7A) and HIVIIIB(Fig. 7B). Similar reductions in p24
[image:4.585.43.285.67.444.2]antigen production were observed when the supernatant fluids were filtered to remove possible contaminating viral particles and when PBMCs were infected with the wild-type Bilthoven FIG. 3. Inhibition of p24 antigen production in HIV-1-infected
PBMCs after the addition of measles virus (MV) at different time points. Chicago-1 MV or Edmonston MV (MOI of 0.15) was added to PBMCs infected with CCR5-tropic HIV-1BaL (A) or CXCR4-tropic
HIV-1IIIB (B) 3 h before 1 infection, concurrently with
HIV-infection (0 h), or 15 h after HIV-1 HIV-infection. The levels of p24 antigen (Ag) were measured after 96 h. Experiments were performed in du-plicate with PBMCs from three to five donors, and data are presented as means⫾standard errors of the means (error bars). The value that is significantly different (P⬍0.05) between MV and the corresponding dilution of Vero cell lysate is indicated by an asterisk.
9200 GARCI´A ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 4. Replication of MV in HIV-1-infected PBMCs and requirement for live MV. Chicago-1 or Edmonston MV was added to CCR5-tropic HIV-1BaL-infected PBMCs (A) or CXCR4-tropic HIV-1IIIB-infected PBMCs (B) at a MOI of 15 or 1.5 to 0.00015. Production of MV was
measured after 96 h of coculture by plaque formation on Vero cells. Data are presented as means⫾standard errors of the means (SEM) (error bars) of duplicate wells with PBMCs from two to six donors. Values that are significantly different (P⬍0.05) between Chicago-1 and Edmonston strains of MV are indicated by an asterisk. ND, not done. (C and D) Time course of MV production by HIV-1-infected PBMCs. PBMCs were infected with HIV-1 CCR5-tropic HIV-1BaL(C) or CXCR4-tropic HIV-1IIIB(D), and Chicago-1 or Edmonston MV was added on day 2 of cells
in culture at a MOI of 0.15. Data are presented as means⫾SEM (error bars) of duplicate wells with PBMCs from three to seven donors. Values that are significantly different (P⬍0.05) between Chicago-1 and Edmonston strains of MV are indicated by an asterisk. MV and Vero cell lysates were UV irradiated for 2 min and added to HIV-1BaL-infected PBMCs (E) or HIV-1IIIB-infected PBMCs (F) and tested at 96 h. p24 antigen (Ag)
production was determined by EIA. Values that are significantly different (P⬍0.05) between Chicago-1 and Edmonston MV strains are indicated by an asterisk and bracket. Data are presented as means⫾SEM (error bars) of duplicate wells with PBMCs from 10 donors. Values that are significantly different (P⬍0.05) between live or inactivated MV and Vero cell lysate are indicated by an asterisk.
on November 8, 2019 by guest
http://jvi.asm.org/
strain of MV (data not shown). The reduction in p24 antigen production following the addition of supernatant fluid from MV-infected PBMCs was not as marked (maximum, 70 to 80%) as following the direct addition of MV to PBMCs (max-imum, 85 to 95% [Fig. 2A and B]).
Measurement of chemokines in supernatant fluids.To de-termine whether MV coinfection increased production of HIV-inhibitory chemokines, the levels of chemokines were measured in the supernatant fluids from PBMCs coinfected with MV and HIV-1 (Table 2). The levels of the-chemokines RANTES, MIP-1␣, and MIP-1were measured for cultures infected with CCR5-tropic HIVBaL and of the ␣-chemokine
SDF-1 for CXCR4-tropic HIVIIIB. The levels of MIP-1␣,
MIP-1, and SDF-1 were increased, but these increases were similar for MV-infected PBMCs and Vero cell lysate controls and did not correlate with decreases in p24 antigen production. Fur-thermore, the addition of high concentrations of neutralizing antibodies to the-chemokines RANTES, MIP-1␣, and MIP-1had no effect on the inhibition of p24 antigen production by MV (data not shown).
DISCUSSION
During measles there is a transient, but profound, reduction in plasma HIV-1 RNA levels in coinfected children (19). In these studies, we have shown that two different strains of MV inhibited replication of both CCR5- and CXCR4-tropic strains of HIV-1 in PBMCs cultured in vitro. This inhibitory effect persisted for at least 12 days, was dose dependent, and was independent of the relative timing of the addition of MV. Inactivated MV and supernatant fluid from PBMCs stimulated with MV also inhibited HIV-1 replication, although not to the same degree as for replication-competent MV. This suppres-sion was not associated with MV-induced death of HIV-sus-ceptible cells or decreased expression of HIV-1 coreceptors and could not be correlated with production of HIV-inhibitory chemokines. Rather, MV-induced suppression of lymphocyte proliferation correlated with decreased p24 antigen production and may contribute to inhibition of HIV-1 replication.
[image:6.585.81.504.71.253.2]We explored several potential mechanisms by which MV could suppress HIV-1 replication. Measles is accompanied by FIG. 5. Effect of MV on cell death. Proportion of propidium iodide-staining HIV-1BaL-infected PBMCs (A) or HIV-1IIIB-infected PBMCs (B) 4
days after the addition of Chicago-1 MV (MOI of 1.5 to 0.015) or Edmonston MV (MOI of 15 to 0.15). Controls were equivalent 10-fold dilutions of Vero cell lysate. Background cell death due to HIV-1 alone (3 to 9%) was subtracted. Data are presented as means⫾standard errors of the means (error bars) of triplicate wells of PBMCs from two or three donors. ND, not done.
TABLE 1. Surface expression of CD4, CD8, CCR5, and CXCR4 4 days after the addition of MV (MOI of 0.15) to HIV-1-infected PBMCsa
HIV-1 strain MV or control % CD4⫹ % CD8⫹ % CCR5⫹ % CXCR4⫹
HIV-1BaL None 51.5⫾2.9 29.8⫾3.3 4.1⫾2.6 ND
Chicago-1 MV 54.0⫾3.3 28.9⫾3.8 4.1⫾2.9 ND
Edmonston MV 53.7⫾2.7 28.2⫾3.6 6.1⫾3.3 ND
Vero cell lysate 50.7⫾3.3 28.8⫾3.3 3.4⫾2.2 ND
HIV-1IIIB None 49.3⫾3.3 34.2⫾5.8 NDb 57.9⫾3.8
Chicago-1 MV 48.0⫾3.3 35.8⫾3.6 ND 77.9⫾3.2c
Edmonston MV 50.5⫾2.6 37.3⫾5.0 ND 64.1⫾6.3
Vero cell lysate 49.6⫾3.7 33.3⫾5.7 ND 63.1⫾2.6
aVero cell lysate (1:10) was used as control. Data represent the means⫾standard errors of the means of the percentage of positive cells from five to nine
experiments.
bND, not determined.
cP⬍0.05 compared to Vero cell lysate.
9202 GARCI´A ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.46.542.588.691.2]a transient lymphopenia, with a reduction in the number of circulating CD4⫹and CD8⫹T lymphocytes. Increased surface expression of Fas (CD95) and annexin V staining suggests that apoptosis of uninfected lymphocytes may contribute to the lymphopenia (23). However, the number of viable cells was not markedly reduced in our in vitro system. Only 10 to 20% of cells were nonviable after 4 days of culture with MV. This degree of cell death is insufficient to explain the marked re-duction in p24 antigen prore-duction. In addition, cell surface
expression of CD4, CD8, CCR5, or CXCR4 was not down-regulated following MV infection, further evidence that loss of target cells did not account for the reduction in p24 antigen production.
[image:7.585.96.492.72.291.2]The inhibitory effect was reduced but not abrogated by in-activation of MV, suggesting that MV replication is necessary for maximal suppression of HIV-1 replication but that suppres-sion may be mediated in part by MV proteins or factors in-duced by MV. The MV immune status of our anonymous FIG. 6. Effect of MV on proliferation of HIV-infected PBMCs. Incorporation of [3H]thymidine by HIV-1
BaL-infected PBMCs (A) or
HIV-1IIIB-infected PBMCs (B) after the addition of Chicago-1 (MOI of 1.5 to 0.015) or Edmonston (MOI of 15 to 0.15) MV or Vero cell lysate at an
equivalent dilution. Data are presented as means⫾standard errors of the means (error bars) of triplicate wells with PBMCs from 6 to 10 donors. Values that are significantly different (P⬍0.05) between MV and Vero cell lysate are indicated by an asterisk. ND, not done.
FIG. 7. Effect of supernatant fluid on p24 antigen production. Supernatant fluid from PBMCs incubated with Chicago-1 MV (MOI of 0.15) or Vero cell lysate at 10-fold dilutions were added to HIV-1BaL-infected PBMCs (A) or HIV-1IIIB-infected PBMCs (B). Supernatant fluids were
harvested on day 1, day 2, and day 3 of stimulation, and p24 antigen (Ag) was assayed after 96 h. Data are presented as means⫾standard errors of the means (error bars) of duplicate wells with PBMCs from seven or eight donors. Values that are significantly different (P⬍0.05) between MV and the corresponding dilution of Vero cell lysate are indicated by an asterisk.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:7.585.92.501.475.677.2]donors of PBMCs was not known, but it is assumed that most, if not all, healthy adults in the United States are MV immune. MV antigens can induce synthesis of IL-4 and gamma inter-feron in the absence of replicating virus (10, 15). Supernatant fluid obtained from PBMCs infected with MV also reduced p24 antigen production, suggesting the presence of soluble factors induced by MV capable of suppressing HIV-1 replica-tion. Several classes of factors with this ability have been identified, including the -chemokines (MIP-1␣, MIP-1, RANTES, and monocyte chemoattractant protein-2) (8, 36), CD8⫹T-cell antiviral factor (17, 27), type 1/2 interferons (24), and the cytokines IL-10, IL-13, IL-16, and IL-18 (1, 2, 5–7). Furthermore, suppression of HIV-1 replication by human her-pesvirus 6 has been linked to production of RANTES (13) and by GBV-C to production of both␣- and-chemokines (35). However, we were not able to identify elevated levels of che-mokines (Table 2) or cytokines (data not shown) in the super-natant fluids of PBMCs stimulated with MV that could account for the suppression, and the addition of neutralizing antibodies to-chemokines did not block the inhibitory effect of MV on p24 antigen production. These results, however, do not exclude the possibility that MV induces as yet unidentified soluble factor(s) that suppresses HIV-1 replication.
Productive HIV-1 replication requires target cell activation and proliferation (25, 37). Specifically, progression to the G1b
phase of the cell cycle is necessary for nuclear importation, integration of the viral genome, and production of viral prog-eny (16). MV inhibits lymphoproliferation (14, 28), an effect that can be mediated by the hemagglutinin (H) and fusion (F) surface glycoproteins (31) and the nucleocapsid protein (18). Coexpression of MV H and F glycoproteins on nonlymphoid cells inhibits proliferation of peripheral blood lymphocytes through a mechanism that requires cleavage of the F protein (31) and impaired protein kinase B (Akt kinase) activation (4), disrupting the downstream pathway critical to IL-2-dependent T-lymphocyte survival and proliferation. In our system, MV inhibited the proliferation of HIV-1-infected PBMCs as mea-sured by incorporation of tritiated thymidine. This inhibitory effect was correlated with the reduction in p24 antigen produc-tion and was more marked with the less attenuated MV strain and with a higher inoculating dose of MV. Thus, the inhibitory
effect of MV on the proliferation of CD4⫹lymphocytes may contribute to the reduction in p24 antigen production.
We observed some differences between the two MV strains in their ability to replicate in this system and in their effects on p24 antigen production, cell viability, and lymphoproliferation. Differences in both structural and nonstructural proteins have been described in attenuated vaccine strains of MV compared to wild-type MV strains (20). Attenuating mutations have not been definitively identified, and differences of unknown impor-tance exist between the D and A clades of MV (34). The fact that the more virulent Chicago-1 strain was more effective in suppressing HIV-1 replication could be related to the induc-tion of a more robust immune response or altered target cell interactions. The Chicago-1 strain replicates better in the hu-man thymus than the Edmonston strain (3), but the Edmon-ston strain replicated better in cultured PBMCs. We cannot exclude the fact that differences in the concentrations of LPS of the two MV strains play a role in some of these differences. However, when LPS was added to HIV-1-infected PMBCs in concentrations similar to those detected in the two MV strains, the effect on p24 antigen production was markedly less, dem-onstrating that LPS was not responsible for the inhibition of p24 antigen production (data not shown).
Multiple mechanisms are likely responsible for the inhibi-tion of HIV-1 replicainhibi-tion by MV. Although this in vitro system provides insights into the potential mechanisms, such a system clearly does not capture the immunologic and virologic com-plexities of a coinfected child. The immune responses to both viruses are dynamic, changing substantially over the course of infection. In addition, cell types necessary for the induction of chemokines or cytokines may not be present in this system. Nevertheless, our in vitro system allows for the investigation of several of these potential mechanisms that is not feasible in coinfected children and suggests that suppression of HIV-1 replication may be mediated by a block in lymphoproliferation induced by MV.
ACKNOWLEDGMENTS
[image:8.585.44.544.81.216.2]This work was funded by the Elizabeth Glaser Pediatric AIDS Foun-dation (PG-51331), the Gustave J. Martin Innovative Research Fund of the Johns Hopkins Bloomberg School of Public Health, and re-search grant AI23047 from the National Institutes of Health. TABLE 2. Chemokine levels in supernatant fluid from PBMCs infected with HIV-1 and MV (MOI of 0.15) for 4 days
HIV-1 strain MV or controla Chemokine level
b(pg/ml)
MIP-1␣ MIP-1 RANTES SDF-1␣
HIV-1BaL None 6,576⫾4,729 7,741⫾4,997 5,218⫾3,510 ND
Chicago-1 MV 12,845⫾9,306 10,227⫾7,389 3,064⫾1,608 ND
Vero cell lysate (1:10) 14,142⫾10,131 11,015⫾9,019 2,567⫾1,326 ND
Edmonston MV 17,216⫾13,277 12,834⫾9,172 3,679⫾1,549 ND
Vero cell lysate (1:100) 18,388⫾18,050 13,314⫾11,699 4,080⫾2,111 ND
HIV-1IIIB None ND ND ND 0⫾0
Chicago-1 MV ND ND ND 83⫾83
Vero cell lysate (1:10) ND ND ND 552⫾249
Edmonston MV ND ND ND 260⫾232
Vero cell lysate (1:100) ND ND ND 290⫾230
aVero cell lysate served as control.
bData represent the means⫾standard errors of the means of duplicate wells from three to eight experiments. ND, not determined.
9204 GARCI´A ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
We are grateful to Robert Siliciano for his many helpful suggestions and provision of reagents.
REFERENCES
1.Akridge, R. E., L. K. M. Oyafuso, and S. G. Reed.1994. IL-10 is induced during HIV-1 infection and is capable of decreasing viral replication in
human macrophages. J. Immunol.153:5782–5789.
2.Amiel, C., E. Darcissac, M. J. Truong, J. Dewulf, M. Loyens, Y. Mouton, A. Capron, and G. M. Bahr.1999. Interleukin-16 (IL-16) inhibits human im-munodeficiency virus replication in cells from infected subjects, and serum
IL-16 levels drop with disease progression. J. Infect. Dis.179:83–91.
3.Auwaerter, P. G., H. Kaneshima, J. M. McCune, G. Wiegand, and D. E. Griffin.1996. Measles virus infection of thymic epithelium in the SCID-hu
mouse leads to thymocyte apoptosis. J. Virol.70:3734–3740.
4.Avota, E., A. Avolts, S. Niewiesk, L. P. Kane, U. Bommhardt, V. ter Meulen, and S. Schneider-Schaulies.2001. Disruption of Akt kinase activation is important for immunosuppression induced by measles virus. Nat. Med.
7:725–731.
5.Bailer, R. T., B. Lee, and L. J. Montaner.2000. IL-13 and TNF-alpha inhibit dual-tropic HIV-1 in primary macrophages by reduction of surface expres-sion of CD4, chemokine receptors CCR5, CXCR4 and post-entry viral gene
expression. Eur. J. Immunol.30:1340–1349.
6.Baler, M., A. Wermer, N. Bannert, K. Metzner, and R. Kurth.1995. HIV
suppression by interleukin-16. Nature378:563.
7.Choi, H. J., C. A. Dinarello, and L. Shapiro.2001. Interleukin-18 inhibits human immunodeficiency virus type 1 production in peripheral blood
mono-nuclear cells. J. Infect. Dis.184:560–568.
8.Cocchi, F., A. L. De Vico, A. Garzino-Demo, S. K. Arya, R. C. Gallo, and P. Lusso.1995. Identification of RANTES, MIP-1alpha, and MIP-beta as the
major HIV-suppressive factors produced by CD8⫹T cells. Science270:
1811–1815.
9.Derdeyn, C. A., J. M. Decker, J. N. Sfakianos, X. Wu, W. A. O’Brien, L. Ratner, J. C. Kappes, G. M. Shaw, and E. Hunter.2000. Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 is modulated by
coreceptor specificity defined by the V3 loop of gp120. J. Virol.74:8358–
8367.
10.Dhiman, N., I. G. Ovsyannikova, R. C. Howe, J. E. Ryan, R. M. Jacobson, and G. A. Poland.2004. Interleukin-4 induced by measles virus and measles-derived peptides as measured by IL-4 receptor-blocking ELISA. J. Immunol.
Methods287:217–225.
11.Enders, J. F., and T. C. Peebles.1954. Propagation in tissue cultures of cytopathic agents from patients with measles. Proc. Soc. Exp. Biol. Med.
86:277–286.
12.Griffin, D. E., B. J. Ward, E. Jauregui, R. T. Johnson, and A. Vaisberg.1989.
Immune activation in measles. N. Engl. J. Med.320:1667–1672.
13.Grivel, J. C., Y. Ito, G. Faga, F. Santoro, F. Shaheen, M. S. Malnati, W. Fitzgerald, P. Lusso, and L. Margolis.2001. Suppression of CCR5- but not CXCR4-tropic HIV-1 in lymphoid tissue by human herpesvirus 6. Nat. Med.
7:1232–1235.
14.Hirsch, R. L., D. E. Griffin, R. T. Johnson, S. J. Cooper, I. Lindo de Soriano, S. Roedenbeck, and A. Vaisberg.1984. Cellular immune responses during complicated and uncomplicated measles virus infections of man. Clin.
Im-munol. Immunopathol.31:1–12.
15.Jaye, A., C. A. Herberts, S. Jallow, S. F. Atabani, M. R. Klein, P. Hoogerhout, M. Kidd, C. A. C. M. van Els, and H. C. Whittle.2003. Vigorous but short-term gamma interferon T-cell responses against a dominant HLA-A*02-restricted measles virus epitope in patients with measles. J. Virol.
77:5014–5016.
16.Korin, Y. D., and J. A. Zack.1998. Progression to the G1b phase of the cell
cycle is required for completion of human immunodeficiency virus type 1
reverse transcription in T cells. J. Virol.72:3161–3168.
17.Levy, J. A., C. E. Mackewicz, and E. Barker.1996. Controlling HIV
patho-genesis: the role of the noncytotoxic anti-HIV response of CD8⫹T cells.
Immunol. Today17:217–224.
18.Marie, J. C., J. Kehren, M. C. Trescol-Biemont, A. Evlashev, H. Valentin, T. Walzer, R. Tedone, B. Loveland, J. F. Nicolas, C. Rabourdin-Combe, and B.
Horvat.2001. Mechanism of measles virus-induced suppression of
inflam-matory immune responses. Immunity14:69–79.
19.Moss, W. J., J. J. Ryon, M. Monze, F. Cutts, T. C. Quinn, and D. E. Griffin.
2002. Suppression of human immunodeficiency virus replication during
acute measles. J. Infect. Dis.185:1035–1042.
20.Parks, C. L., R. A. Lerch, P. Walpita, H. P. Wang, M. S. Sidhu, and S. A. Udem.2001. Comparison of predicted amino acid sequences of measles virus
strains in the Edmonston vaccine lineage. J. Virol.75:910–920.
21.Philpott, N. J., A. J. Turner, J. Scopes, M. Westby, J. C. Marsh, E. C. Gordon-Smith, A. G. Dalgleish, and F. M. Gibson.1996. The use of 7-amino actinomycin D in identifying apoptosis: simplicity of use and broad spectrum
of application compared with other techniques. Blood87:2244–2251.
22.Pinto, L. A.,V. Blazevic, S. A. Anderson, D. J. Venzon, C. M. Trubey, T. Rowe, J. M. Katz, D. Liewehr, M. J. Dolan, and G. M. Shearer.2001. Influenza virus-stimulated generation of anti-human immunodeficiency virus (HIV) activity after influenza vaccination in HIV-infected individuals and healthy
control subjects. J. Infect. Dis.183:1000–1008.
23.Ryon, J. J., W. J. Moss, M. Monze, and D. E. Griffin.2002. Functional and phenotypic changes in circulating lymphocytes from hospitalized Zambian
children with measles. Clin. Diagn. Lab. Immunol.9:994–1003.
24.Shirazi, Y., and P. M. Pitha.1992. Alpha interferon inhibits early stages of
the human immunodeficiency virus type 1 replication cycle. J. Virol. 66:
1321–1328.
25.Stevenson, M., T. L. Stanwick, M. P. Dempsey, and C. A. Lamonica.1990. HIV-1 replication is controlled at the level of T cell activation and proviral
integration. EMBO. J.9:1551–1560.
26.Sulkowski, M. S., R. E. Chaisson, C. L. Karp, R. D. Moore, J. B. Margolick, and T. C. Quinn.1998. The effect of acute infectious illnesses on plasma human immunodeficiency virus (HIV) type 1 load and the expression of serologic markers of immune activation among HIV-infected adults. J.
In-fect. Dis.178:1642–1648.
27.Walker, C. M., D. J. Moody, D. P. Stites, and J. A. Levy.1986. CD8⫹
lymphocytes can control HIV infection in vitro by suppressing virus
replica-tion. Science234:1563–1566.
28.Ward, B. J., R. T. Johnson, A. Vaisberg, E. Jauregui, and D. E. Griffin.1991. Cytokine production in vitro and the lymphoproliferative defect of natural
measles virus infection. Clin. Immunol. Immunopathol.61:236–248.
29.Watt, G., P. Kantipong, M. de Souza, P. Chanbancherd, K. Jongsakul, R. Ruangweerayud, L. D. Loomis-Price, V. Polonis, K. S. Myint, D. L. Birx, A. E. Brown, and S. Krishna.2000. HIV-1 suppression during acute
scrub-typhus infection. Lancet356:475–479.
30.Watt, G., P. Kantipong, and K. Jongsakul.2003. Decrease in human immu-nodeficiency virus type 1 load during acute dengue fever. Clin. Infect. Dis.
36:1067–1069.
31.Weidmann, A., A. Maisner, W. Garten, M. Seufert, V. ter Meulen, and S. Schneider-Schaulies.2000. Proteolytic cleavage of the fusion protein but not membrane fusion is required for measles virus-induced immunosuppression
in vitro. J. Virol.74:1985–1993.
32.Williams, C. F., D. Klinzman, T. E. Yamashita, J. Xiang, P. M. Polgreen, C. Rinaldo, C. Liu, J. Phair, J. B. Margolick, D. Zdunek, G. Hess, and J. T. Stapleton.2004. Persistent GB virus C infection and survival in HIV-infected
men. N. Engl. J. Med.350:981–990.
33.World Health Organization.2001. Nomenclature for describing the genetic characteristics of wild-type measles viruses (update)—part I. Wkly.
Epide-miol. Rec.76:242–247.
34.World Health Organization.2003. Update of the nomenclature for describ-ing the genetic characteristics of wild-type measles viruses: new genotypes
and reference strains. Wkly. Epidemiol. Rec.78:229–232.
35.Xiang, J., S. L. George, S. Wunschmann, Q. Chang, D. Klinzman, and J. T. Stapleton.2004. Inhibition of HIV-1 replication by GB virus C infection
through increases in RANTES, MIP-1␣, MIP-1, and SDF-1. Lancet363:
2040–2046.
36.Yang, O. O., E. A. Garcia-Zepeda, B. D. Walker, and A. D. Luster.2002. Monocyte chemoattractant protein-2 (CC chemokine ligand 8) inhibits rep-lication of human immunodeficiency virus type 1 via CC chemokine receptor
5. J. Infect. Dis.185:1174–1178.
37.Zack, J. A.1995. The role of the cell cycle in HIV-1 infection. Adv. Exp.
Med. Biol.374:27–31.