Adenovirus Type 3 (HAdV-3) and HAdV-7 Reside in Multiple Hexon
Hypervariable Regions
Hongling Qiu,a,bXiao Li,aXingui Tian,aZhichao Zhou,aKe Xing,aHaitao Li,aNi Tang,aWenkuan Liu,aPeisheng Bai,aand Rong Zhoua
State Key Laboratory of Respiratory Diseases (Guangzhou Medical University), The First Affiliated Hospital of Guangzhou Medical College, Guangzhou, China,a and Institute for Nutritional Science, Shanghai Institute for Biological Science, Chinese Academy of Science, Shanghai, Chinab
Human adenovirus types 3 and 7 (HAdV-3 and HAdV-7) occur epidemically and contribute greatly to respiratory diseases, but
there is no currently available licensed recombinant HAdV-3/HAdV-7 bivalent vaccine. Identification of serotype-specific
neu-tralizing antibody (NAb) epitopes for HAdV-3 and HAdV-7 will be beneficial for development of recombinant HAdV-3/HAdV-7
bivalent vaccines. In this study, four NAb epitopes within hexon hypervariable regions (HVRs) were predicted for HAdV-3 and
HAdV-7, respectively, by using bioinformatics. Eight hexon chimeric adenovirus vectors with the alternation of only one
pre-dicted neutralizing epitope were constructed. Further
in vitro
and
in vivo
neutralization assays indicated that E2 (residing in
HVR2) and E3 (residing in HVR5) are NAb epitopes for HAdV-7, and E3 plays a more important role in generating NAb
re-sponses. Cross-neutralization assays indicated that all four predicted epitopes, R1 to R4, are NAb epitopes for HAdV-3, and R1
(residing in HVR1) plays the most important role in generating NAb responses. Humoral immune responses elicited by the
re-combinant rAdH7R1 (containing the R1 epitope) were significantly and durably suppressed by HAdV-3-specific NAbs.
Surpris-ingly, the rAd
⌬
E3GFP-specific neutralizing epitope responses induced by rAdMHE3 (R3 replaced by E3) and rAdMHE4 (R4
re-placed by E4) were weaker than those of rAdMHE1 (R1 rere-placed by E1) or rAdMHE2 (R2 relaced by E2)
in vitro
and
in vivo
.
Furthermore, rAdMHE4 replicated more slowly in HEp-2 cells, and the final yield was about 10-fold lower than that of
rAd
⌬
E3GFP. The current findings contribute not only to the development of new adenovirus vaccine candidates, but also to the
construction of new gene delivery vectors.
T
o date, 51 human adenovirus (HAdV) serotypes have been
identified and divided into six groups (A to F) based on their
morphological, immunogenic, and genomic information (
7
). A
novel human adenovirus, HAdV-52, was identified recently, but
whether it should be considered a novel “serotype” remains
dis-puted (
6
,
12
). Most HAdV can cause respiratory disease,
gastro-enteritis, pneumonia, and keratoconjunctivitis, among other
dis-eases (
3
,
10
,
13
). Of these, the acute respiratory diseases induced
by the subspecies B1 adenoviruses HAdV-3 and HAdV-7 have
occurred epidemically and have caused outbreaks in Asia, Europe,
and North America (
2
,
11
,
15
,
21
,
26
,
31
). However, there is
cur-rently no efficient antiviral therapy against adenoviruses and,
therefore, it is necessary to develop licensed HAdV-3 and HAdV-7
vaccines to help prevent adenovirus infection outbreaks. A simple
combined formulation of HAdV-3 and HAdV-7 monovalent
vac-cines combined to produce a bivalent vaccine might introduce
considerable constraints, such as serotype competition and/or
dominance. Therefore, a recombinant vaccine containing
HAdV-3 and HAdV-7 neutralizing epitopes that elicits protective
immunity simultaneously to HAdV-3 and HAdV-7 is required.
However, no neutralizing epitopes have been identified for
HAdV-3 and HAdV-7 until now.
All adenovirus particles contain a common structure
consist-ing of the nonenveloped capsid, which is 60 to 90 nm in diameter
with icosahedral symmetry. Twelve copies of a trimeric hexon
form each of the 20 triangular facets of the icosahedron, and
tri-meric fibers are inserted into the 12 pentatri-meric penton bases at
the 12 vertices. All three major structural proteins, the hexon,
fiber, and penton, can generate neutralizing antibodies (NAbs)
against adenovirus, but the major protein component of the
ico-sahedral capsid, the hexon, is the major target of serotype-specific
NAbs (
17
,
19
,
24
). Alignment of available HAdV hexon protein
sequences reveals high homology except within the seven
hyper-variable regions (HVRs). The highly conserved domains of the
hexon protein form the core tertiary structure, a pseudohexagonal
shape that allows formation of tightly packed hexon arrays within
the facets of the icosahedral capsid, and the seven HVRs are
local-ized on the exterior surface of the virion (
20
). It is therefore
pos-sible to change amino acids in the HVRs without destroying the
core tertiary structure of the hexon protein.
A hexon chimeric adenovirus serotype 5 vector was reported to
circumvent preexisting antivector immunity by replacing the
seven short HVRs on the surface of the HAdV-5 hexon protein
with the corresponding HVRs from HAdV-48 without perturbing
its core structure (
19
). These data indicated that the seven HVRs
contained epitopes recognized by serotype-specific NAbs.
There-fore, if serotype-specific NAb epitopes for different adenoviruses
can be identified, it would be possible to generate polyvalent
vac-cines by coexpressing different serotype-specific NAb epitopes in
the HVRs of hexon proteins. In this study, we sought to identify
the serotype-specific NAb epitopes for HAdV-3 and HAdV-7 by
Received15 December 2011Accepted8 May 2012
Published ahead of print23 May 2012
Address correspondence to Rong Zhou, [email protected]. Q.H., L.X., and T.X. contributed equally to this work.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.07076-11
on November 7, 2019 by guest
http://jvi.asm.org/
construction of a gene replacement strategy. Our study will
con-tribute to the development of new HAdV-3/HAdV-7
recombi-nant vaccines and gene delivery vectors.
MATERIALS AND METHODS
Defining the potential NAb epitopes of HAdV-3 and HAdV-7 hexons. The nonconserved domains of HAdV-3 and HAdV-7 hexon proteins were determined by aligning them with the available human hexon protein sequences using the protein BLAST program (Basic Local Alignment Tool; http://blast.ncbi.nlm.nih.gov/Blast.cgi). The EMBOSS antigenic tool (http://binfo.ym.edu.tw/emboss/Apps/antigenic.html) was used to predict antigenic epitopes of the HAdV-3 and HAdV-7 hexon proteins. The potential template structures for modeling HAdV-3 and HAdV-7 hexon proteins were identified by searching them against information in the Protein Data Bank (PDB) by BLAST-P analysis. The Modeler 9v8 tool was used to model the three-dimensional (3D) structure of HAdV-3 and HAdV-7 hexon proteins (8). The antigenic epitopes that were predicted to be exposed on the capsid surface and located in nonconserved regions were selected as potential sites for recognition by NAbs.
Construction of the mutagenesis vectors MHE1, pBRAd-MHE2, pBRAd-MHE3, pBRAd-MHE4, pBRAd3H7R1, pBRAd3H7R2, pBRAd3H7R3, and pBRAd3H7R4.The plasmid pBRAd⌬E3GFP encod-ing a HAdV-3 GZ-01 vector (GenBank accession no.DQ099432) and enhanced green fluorescent protein (GFP) with an E3 region deletion was constructed as previously described (32). The hexon chimeric adenovirus vector pAd3egf/H7, in which the HAdV-3 hexon gene in the pBRAd⌬E3GFP vector was replaced with the hexon gene from HAdV-7, was constructed as previously described (27). The mutated hexon genes were cloned into small pBRLR shuttle vectors that facilitated transfer of the mutated hexon genes into full-length viral plasmids pBRAd⌬E3GFP or pAd3egf/H7.
(i) Construction of the shuttle vector pBRLR.The L fragment (from nucleotide [nt] 16748 to nt 18435) was amplified from plasmid pBRAd⌬E3GFP using the primers HexonLU and HexonLD (Table 1) and subsequently digested with EcoRI and ClaI and inserted into the EcoRI/ ClaI-digested pBR322 vector to generate the pBRL vector. Fragment R (from nt 21119 to nt 24526) was amplified from plasmid pBRAd⌬E3GFP using the primers HexonRU and HexonRD (Table 1) before being
di-TABLE 1Primers used in this study
Primer Sequencea
HexonLU 5=-AAAGAATTCTATCCAGCCTGAGGTCAAAGTAAGA-3=
HexonLD 5=-GTCATCGATGGGGTGGCCATCTTGAAAG-3=
HexonRU 5=-AAAGGATCCCATGGATGAGCCCACCCTG-3=
HexonRD 5=-AAGGTCGACTGCATGTTCTGGTTTTCTGAAAG-3=
HexU 5=-CCCATCGATGATGCCCCAATGGGCATAC-3=
HexD 5=-CCGGGATCCACCTCAAAAGTCATGT-3=
MHE1D 5=-GTATGTGGTGGCATTGTCTTCTCCCGTTGTAACTATCCA-3=
MHE1U 5=-GGAGAAGACAATGCCACCACATACACATTTGGCATTGCT-3=
MHE2D 5=GTTGTCTGCGGTAATGTCTTTCCCAATTTCCAAACCTTCTTTAG T3=
MHE2U 5=-GAAATTGGGAAAGACATTACCGCAGACAACAAGCCCATTTATGCCGAT-3=
MHE3D 5=-CGAAAAAGCGTCAGCAGCTTC CCTACCATCGAAAAA-3=
MHE3U 5=-GAAGCTGCTGACGCTTTTTCG CCTGAAATTGTGCTT-3=
MHE4U 5=-CCTAGAGACACTGCATGGGAAAAAGATACTAAAGTTGATACAGCTAAT-3=
MHE4D 5=-TTTAGTATCTTTTTCCCATGCAGTGTCTCTAGGTTTAATGCCTTGATA-3=
IMHE1a 5=-GTATGTGGTGGCATTGTCTTCTCCCG-3=
IMHE2a 5=-GGCATAAATGGGCTTGTTGTCTGCGG-3=
IMHE3a 5=-CGAAAAAGCGTCAGCAGCTTCCCT-3=
IMHE4a 5=-TTTAGTATCTTTTTCCCATGCAGTGTCTCTAGGT-3=
A3HVR1u 5=-ACATCTCAGTGGATAGTTACAACGAATCGAGACAATGCAGTAACTACCACCACAAACAC-3=
A3HVR1r 5=-AGCAATGCCAAATGTGTTTGTGGTGGTAGTTACTGCATTGTC TCGATTCGTTGTAACTATCCACTGAGATGTATTGG-3=
A3HVR2u 5=-ATTGGGAAAGACATTACTACTACTGAAGGAGAAGAAAAGCCCATTTATGCCGATAAAAC-3=
A3HVR2r 5=-CTGATATGTTTTATCGGCATAAATGGGCTTTTCTTCTCCTTCAGTAGTAGTAATGTCTTTCCCAATTTCTAAACCTTC-3=
A3HVR5u 5=-TTTGATGGTAGAGATGCTGTTGCAGGAGCTTTAGCGCCTGAAATTGTGCTTTACACGGA-3=
A3HVR5r 5=-TTCCGTGTAAAGCACAATTTCAGGCGCTAAAGCTCCTGCAAC AGCATCTCTACCATCAAAAAATTCCATATCAATATC-3=
A3HVR7u 5=-AAGTTAAAACCGATGACGCTAATGGATGGGAAAAAGATGCTAATGTTGATACAGCTAATGAAATAGCCATAGGCAAC-3=
A3HVR7r 5=-AGCTGTATCAACATTAGCATCTTTTTCCCATCCATTAGCGTCATCGGTTTTAACTTTAATGCCTTGATATGTTTTCGC-3=
HR1r 5=-TACTGCATTGTCTCGATTCGT-3=
HR2r 5=-GCTTTTCTTCTCCTTCAGT-3=
HR5r 5=-CGCTAAAGCTCCTGCAACAGCATC-3=
HR7r 5=-CCATTAGCGTCATCGGTTTTAAC-3=
HU 5=-GGGTATTCCATATGGCCACCCCATCGATGATGCCCC-3=
HD 5=-CCGCTCGAGTTATTGAGGCTCTGGCTGATATG-3=
AD7SPR1 5=-GGCAGTTGTGCTGGGCATG-3=
AD7SPR2 5=-TTCTCTGGAATTAACATTGAAAGGATAG-3=
AD7SPS 5=-CCACTTAATCATAAATCAGGGCAAAA-3=
AD7SPBH 5=-FAM-TGGCTACTGGTGCCCTTACTAATGCTAAAGG-BHQ1-3=b
AD3GZ01F 5=-CGAGACCTCCTACCCATGAACTAA-3=
AD3GZ01R 5=-GACTGTATTGCTGATTTCAAGTAAGTGTCT-3=
AD3GZ01FB 5=-FAM-TCATTGCCCCTACCTTACCCAATCCAA-BHQ1 aUnderlined nucleotides indicate restriction sites used for cloning.
b
FAM, 6-carboxyfluorescein; BHQ1, Black Hole Quencher 1.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.42.548.78.486.2]gested with BamHI and SalI and insertion into the BamHI/SalI-digested pBRL vector to generate the pBRLR vector.
(ii) Construction of HAdV-3 hexon mutagenesis vectors pBRAd-MHE1, pBRAd-MHE2, pBRAd-MHE3, and pBRAd-MHE4.HAdV-3 hexon mutagenesis vectors MHE1, MHE2, pBRLR-MHE3, and pBRLR-MHE4 were created to facilitate the transfer of mutated hexon fragments from the mutagenesis vectors into the full-length plasmid pBRAd⌬E3GFP. The mutated HAdV-3 hexon gene fragment MHE1 (or MHE2, MHE3, or MHE4) was mutated by replac-ing the region encodreplac-ing the potential neutralizreplac-ing epitope R1 (or R2, R3, or R4) of the HAdV-3 hexon gene with the region encoding the potential neutralizing epitope E1 (or E2, E3, or E4) of the HAdV-7 hexon gene. These mutated fragments were produced by overlapping PCR extension mutagenesis. For example, the shuttle vector pBRLR-MHE1 was constructed as follows. The primer pair pBRLR-MHE1U and MHE1D contained the complementary mutated region that encoded the E1 epitope from the HAdV-7 hexon gene. A 2,279-bp segment was amplified from plasmid pBRAd⌬E3GFP by using the primers MHE1U and HexD (Table 1). In a separate PCR, a 433-bp segment was ampli-fied from plasmid pBRAd⌬E3GFP using the primers MHE1D and HexU (Table 1). Subsequently, the two PCR products were purified and annealed as a template in a second round of PCR with the primer pair HexU and HexD, to generate a 2,697-bp PCR product, MHE1. This fragment was digested with ClaI/BamHI and inserted into ClaI/ BamHI-digested pBRLR to construct the pBRLR-MHE1 vector. The vectors pBRLR-MHE2, pBRLR-MHE3, and pBRLR-MHE4 were con-structed using the same strategy.
The vectors MHE1, MHE2, MER3, pBRLR-MHE4, were digested with EcoRI and SalI, and the digested fragments (LRMHE1, LRMHE2, LRMHE3, and LRMHE4) containing mutated HAdV-3 hexon genes were purified using a gel extract kit (Qiagen). The plasmid pBRAd⌬E3GFP was digested with PacI and AvrII, and the 35-kb fragment without the wild-type hexon gene was purified and served as the backbone, which was used to transformed BJ5183 cells with the linear fragment MHE1, MHE2, MHE3, or MHE4 to reconstruct the mutant viral genome. The presence of mutations in the viral vectors pBRAd-MHE1, pBRAd-MHE2, pBRAd-MHE3, and pBRAd-MHE4 was verified by se-quencing cDNA clones for hexon transcripts.
(iii) Construction of HAdV-7 hexon mutagenesis vectors pBRAd-H7R1, pBRAd-H7R2, pBRAd-H7R3,and pBRAd-H7R4.We used the same strategy to construct the HAdV-7 hexon mutagenesis vectors pBRAd-H7R1, pBRAd-H7R2, pBRAd-H7R3, and pBRAd-H7R4. For ex-ample, the mutated fragment H7R1 was produced by overlapping PCR extension mutagenesis with primer pair A3HVR1u/HexD, A3HVR1r/ HexU, and HexU/HexD, using pAd3egf/H7 as DNA templates. Then, the fragment H7R1 was cloned into pBRLR to generate shuttle vector pBRLR-H7R1. Finally, the LR-H7R1 fragment was cloned into the pAd3egf/H7 vector to generate HAdV-7 hexon mutagenesis vector pBRAd-MH7R1 by using recombinant DNA technology. The presence of mutations in the viral vectors H7R1, H7R2, H7R3, and pBRAd-H7R4 was verified by sequencing cDNA clones for hexon transcripts.
Before transfection, 4⫻105HEK-293 cells were added to each well of a 24-well plate (Corning) and grown overnight at 37°C in a 5% CO2 environment. The chimeric viral genomes were released by using AsiSI (New England BioLabs) from the vector backbones and transfected into HEK-293 using the Sofast transfection kit (Xiamen Sunma Biotechnol-ogy, China). After 5 days, the transfected cells were harvested, freeze-thawed for three cycles, and used to infect HEp-2 cells. The recombinant adenoviruses rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3, and rAdH7R4 were harvested when a 95 to 100% cytopathic effect (CPE) was observed after several passages.
Animals, virus strains, and cells.Female BALB/c mice were pur-chased from the Animal Center at Sun Yat-sen University and housed under pathogen-free conditions. HAdV-7 strain GZ08 (GenBank acces-sion no.GQ478341), HAdV-3 strain GZ01, rAd⌬E3GFP, and eight
re-combinant viruses (rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3, and rAdH7R4) were cultured in HEp-2 cells or 293 cells. Virus particles were purified by using CsCl gradient centrifugation. After purification, the viruses were quantified by real-time PCR and stored at⫺80°C. HEp-2 cells and 293 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 100 IU/ml penicillin, 100 mg/ml streptomycin, and 10% newborn calf serum. RT-PCR. To confirm the generation of chimeric viruses, we per-formed reverse transcription-PCR (RT-PCR) assays with specific PCR primers for the eight chimeric viruses, 1, 2, 3, and 4 and rAdH7R1, rAdH7R2, rAdH7R3, and rAdH7R4. The common sense primer HU and antisense primer HD were located in the conserved regions of HAdV-3 and HAdV-7 hexon protein sequences, and the antisense primers (IMHE1a, IMHE2a, IMHE3a, IMHE4a, HR1r, HR2r, HR3r, and HR4r) were located in the eight mutant regions (E1, E2, E3, and E4 from HAdV-7 strain GZ08 hexon and R1, R2, R3, and R4 from HAdV-3 strain GZ01) of chimeric hexons. Total RNA was extracted from HEp-2 cells infected with rAd⌬E3GFP, HAdV-7, rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3, or rAdH7R4 and reverse transcribed to cDNA templates. PCR controls included rAd⌬E3GFP cDNA, HAdV-7 cDNA, and total RNA. A PCR using the primer pair HU and HD was performed to test cDNA quality. A PCR using the primer pair HU and IMHE1a (or IMHE2a, IMHE3a, IMHE4a, HR1r, HR2r, HR3r, or HR4r) was performed to examine the stability of chimeric viruses.
Western blotting.HEp-2 cells were infected with the recombinant viruses rAd⌬E3GFP, rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4. At 96 h postinfection, cells were harvested and freeze-thawed three times; then the virus suspensions were mixed with 2⫻loading buffer (1% so-dium dodecyl sulfate [SDS], 10% 2-mercaptoethanol, 0.0025% phenol red, and 10% glycerol in 50 mM Tris-HCl [pH 6.8]) and boiled for 5 min at 100°C. Samples were then separated on 10% SDS-polyacrylamide gels and electrophoretically transferred onto nitrocellulose membranes. The membranes were blocked in blocking buffer (1% [wt/vol] bovine serum albumin [BSA], 5% [wt/vol] skim milk in 10 mM phosphate-buffered saline [PBS]) for 2 h and probed with the monoclonal antibody 5D4, which specifically recognizes hexons from HAdV-3 and HAdV-7, as pre-viously described (18). The membranes were further incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies. The color reactions were performed with 0.1 M Tris-HCl (pH 6.8) containing 0.5 mg/ml diaminobenzidine (Sigma) and 0.01% H2O2.
Quantitative PCR.To evaluate the replication characteristics of the recombinant viruses rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, and rAd⌬E3GFP, real-time quantitative PCR (Q-PCR) was performed using an adenovirus Q-PCR kit (Huayin Corporation, Guangzhou, China). HEp-2 cells were infected with 1⫻108to 1⫻109genome copies of rAd⌬E3GFP, rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4. The in-fected cells were harvested after 12, 24, 36, 48, 60, and 72 h postinfection, and viral genome copies were determined using adenovirus Q-PCR kits. Forin vivoneutralization assays, the rAd⌬E3GFP, rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3, rAdH7R4, and pAd3egf/H7 genome copy numbers were determined using primers AD3GZ01F, AD3GZ01R, and AD3GZ01FB, and the HAdV-7 genome copy numbers were determined using primers AD7SPR1, AD7SPR2, AD7SPS, and AD7SPBH.
As a standard for the determination of adenovirus genome copies, the rAd⌬E3GFP genome was purified and quantified. Amplification was per-formed using an Applied Biosystems 7500 real-time PCR system. To as-sess reproducibility, each assay was performed independently three times in duplicate.
Fluorescence-based microneutralization assay.Except for HAdV-7, all recombinant viruses (rAd⌬E3GFP, pAd3egf/H7, rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3, and rAdH7R4) used in this study encoded enhanced GFP. Fluorescence read-ings were made using a microplate spectrophotometer (Thermo
on November 7, 2019 by guest
http://jvi.asm.org/
skanFlash) to measure the ability of different sera to neutralize these vi-ruses (28).
Six- to 8-week-old BALB/c mice received 1010genome copies of CsCl gradient-purified rAd⌬E3GFP, pAd3egf/H7, HAdV-7, rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2, rAdH7R3,or rAdH7R4 intraperitoneally five times at 2-week intervals (seven animals per virus group). Mice were sacrificed, and antisera were used forin vitro neutralization assays. The sera from naïve BALB/c mice were used as neg-ative controls. Nonspecific protection was observed when the dilution of the serum from naïve mice was lower than 1:64. Therefore, all sera used in the neutralization assays were used at an initial dilution of 1:64.
Each polyclonal serum was inactivated at 56°C for 30 min and then serially diluted in PBS, and 150l of each dilution was mixed with 150l rAd⌬E3GFP, Ad3/H7, HAdV-7, rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAdH7R1, rAdH7R2,rAdH7R3, or rAdH7R4 recombinant vi-ruses (⬃1⫻107virus particles). The antibody-virus mixtures were incu-bated at 37°C and 5% CO2and then transferred into the HEp-2 cells in triplicate (1⫻104cells per well) in 96-well plates and incubated for 72 h. Enhanced GFP expression in HEp-2 cells was measured using a micro-plate spectrophotometer (Thermo VarioskanFlash [488-nm excitation, 507-nm emission]) (28). Background fluorescence was equalized to wells containing cells only, and maximum fluorescence was determined based on wells with cells infected only with virus. Due to edge effects on fluores-cence readings, the outer 36 wells of 96-well plates were not used in any assay. Each assay included control wells of HEp-2 cells only, HEp-2 cells infected with virus, and HEp-2 cells infected with virus mixed with sera from naïve mice. The neutralizing ability of the serum was calculated as follows: (fluorescence in cells incubated with the antibody-virus mixture
⫺background fluorescence)/(maximum fluorescence). The NAbs titers were defined as the serum dilutions that resulted in a 70% reduction of maximum fluorescence (cells infected only with viruses).
To confirm the results of fluorescence-based microneutralization as-says, neutralization assays were performed under the same conditions, and genome copies of adenovirus were quantified to measure the inhibi-tion of virus infecinhibi-tion by different sera using adenovirus Q-PCR kits. In addition, assays were performed independently, three times in duplicate, to assess reproducibility. Furthermore, for further standardization, neu-tralization assays were performed with the same virus batch and serum batch by the same operator. The NAb titers were defined as the serum dilution that resulted in a 90% reduction of maximum genome copy number (cells infected only with viruses).
CMN assay.The HAdV-7 virus vector does not encode GFP; there-fore, colorimetric microneutralization (CMN) assays were used to exam-ine the neutralization abilities of different sera against HAdV-7, as previ-ously described (5). In brief, sera were inactivated at 56°C for 30 min, then serially diluted in PBS, and 150l of each dilution was mixed with 150l of HAdV-7 virus (100 TCID50[50% tissue culture infectious doses]) at 37°C for 1 h in 5% CO2. Then, 100l was transferred into 96-well plates containing HEp-2 cells in triplicate. The assays included control wells of HEp-2 cells only, HEp-2 cells infected with virus, and HEp-2 cells infected with virus that was mixed with sera from naive mice. After incubation for 72 h, the culture medium was removed from the wells. One hundred microliters of a 1:5,000 dilution of Finter’s neutral red in PBS was added, and the plates were incubated at 37°C in 5% CO2for 1 h. All medium was removed, and the wells were gently washed twice with 150l PBS. A final solution of acid alcohol (50% ethanol, 1% acetic acid in distilled water) was added (100l per well). TheA550of the solubilized dye was read on a microplate spectrophotometer (Thermo Varyskan Flash). The assays were performed three times in duplicate to assess reproducibility. The NAb titers were defined as the serum dilution that protected 90% of cells from HAdV-7 infection. The results of the CMN assay were confirmed using Q-PCR assays as described above.
In vivoneutralization assay.Female BALB/c mice with no previous exposure to rAd⌬E3GFP and HAdV-7 were tested by Q-PCR and en-zyme-linked immunosorbent assay (ELISA) with blood from tail veins. A
total of 168 mice were divided into two groups: 96 mice in group A and 72 mice in group B (seeFig. 5A, below). In group A, 72 mice were intraperito-neally vaccinated with 1010genome copies of different viruses (HAdV-7, rAd⌬E3GFP, rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4) four times at 2-week intervals, with 12 mice per virus group. Forty-eight mice in group B received 1010genome copies of rAd⌬E3GFP, rAdMHE3, rAd-MHE4, or HAdV-7, with the same vaccination schedule and with 12 mice per virus group. Twenty-four nonvaccinated mice in each group were used as negative controls. Two weeks after the last vaccination, vaccinated mice in group A were anesthetized with diethyl ether and challenged in-tranasally with 50l (about 2⫻109genome copies) of live HAdV-7 virus, and vaccinated mice in group B were anesthetized and challenged intra-nasally with 50l (about 2⫻109genome copies) of live rAd⌬E3GFP virus. Twelve nonvaccinated mice in each group were challenged with live HAdV-7 or rAd⌬E3GFP virus and used as negative controls. Twelve naïve mice without virus challenge in each group were used as additional neg-ative controls.
Prior to receiving the virus challenge (day 0), three mice in each sub-group were sacrificed, and the lungs were used to examine the genome copies of challenge virus, HAdV-7 or rAd⌬E3GFP. The remaining mice were sacrificed on days 3, 5, and 7 (three mice per time point) after virus challenge. The lungs from the sacrificed mice were harvested and homog-enized in a Tissuemizer (Tekmar) in 600l of homogenization buffer (10 mM Tris-HCl, 2% SDS, and 1 mM EDTA; pH 8.0). The numbers of HAdV-7 or rAd⌬E3GFP copies in the lung tissues were determined by Q-PCR, as described above.
Adoptive transfer assays.To study the inhibitory effects of HAdV-3-specific NAbs to four rAd3/H7-based hexon chimeric viruses, adoptive transfer studies were performed as previously described (25). BALB/c mice were preimmunized with 1010virus particles of wild-type HAdV-3 GZ01 three times, and then IgG was purified from 2 ml of sera pooled from five immunized mice. The purified antibody was then injected into naive mice via the tail vein. For passive antibody transfer, mice were in-jected via the tail vein with 40l HAdV-3-specific antibody (with a neu-tralizing titer of 1:16,000). On the day following adoptive transfer, the neutralizing antibody titers of the sera from recipient mice were measured in a fluorescence-based microneutralization assay as described above, and then the recipient mice were immunized intramuscularly with 5⫻109 virus particles of rAd⌬E3GFP, rAd3/H7, rAdH7R1, rAdH7R2, rAdH7R3, and rAdH7R4 vectors, which encoded EGFP. Control mice received adoptive transfer of IgG purified from PBS-injected mice.
At days 12 and 18 postimmunization, the EGFP-specific antibody ti-ters from immunized mice were measured by ELISA. Ninety-six-well plates were coated overnight with 100l/well of 1g/ml recombinant GFP (Millipore) and then blocked for 2 h with blocking buffer (PBS, 2% BSA,0.05% Tween 20). Sera dilutions (100-fold dilution of sera obtained at day 12 postimmunization, 40-fold dilution of sera obtained at day 18 postimmunization) were then added to the wells and incubated for 1 h. The plates were washed three times with PBST (PBS, 0.05% Tween 20) and incubated for 1 h with a 1/10,000 dilution of rabbit anti-mouse IgG plus IgM (H⫹L)–peroxidase– conjugated affinity-purified secondary an-tibodies (Jackson ImmunoResearch). The plates were washed four times, and reactions were developed with tetramethylbenzidine, stopped with 2 M H2SO4, and analyzed at 450 nm with an ELISA plate reader (Thermo Scientific Multiskan MK3).
RESULTS
Analysis of potential serotype-specific NAb epitopes for
HAdV-3 and HAdV-7.
Previous studies indicated that epitopes
recognized by serotype-specific NAbs may be exposed on the
vi-rion surface and reside within the seven HVRs of hexon proteins
(
1
,
17
). Seven HVRs that reside within HAdV-3 and HAdV-7
hexon proteins were identified by alignment of the available
hu-man adenovirus hexon protein sequences (
4
). Using sequence
alignment data and the known 3D structure of hexon proteins
on November 7, 2019 by guest
http://jvi.asm.org/
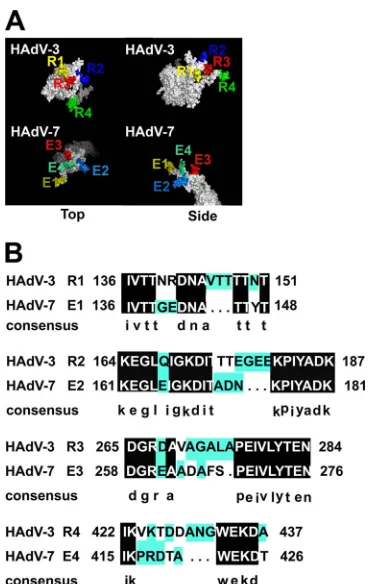
from HAdV-2, HAdV-5, and AdC68 as models, four regions
within the HAdV-3 hexon (R1, residues 136 to 151; R2, residues
164 to 187; R3, residues 265 to 284; R4, residues 422 to 437) and
four regions within the HAdV-7 hexon (E1, residues 136 to 148;
E2, residues 161 to 181; E3, residues 258 to 276; E4, residues 415 to
426) were predicted as potential NAb targets (
Fig. 1A
). These
po-tential neutralization sites for HAdV-3 and HAdV-7 were located
within the HVR1 (R1 and E1), HVR2 (R2 and E2), HVR5 (R3 and
E3), and HVR7 of hexon proteins (R4 and E4) (
Fig. 1A
).
Con-served amino acids were found between the predicted epitopes of
HAdV-3 and the corresponding epitopes of HAdV-7 by sequence
alignment (
Fig. 1B
).
Four hexon chimeric adenovirus vectors were constructed
by replacing the predicted epitopes within the HAdV-3 hexon
protein with the corresponding epitopes from the HAdV-7
hexon.
To identify the neutralizing epitopes for HAdV-7, four
hexon chimeric adenoviruses containing predicted NAb epitopes
from HAdV-7 were constructed. A recombinant HAdV-3-based
vector, rAd
⌬
E3GFP expressing GFP, was constructed in a
previ-ous study (
32
). Four recombinant virus vectors, rAdMHE1 (with
the predicted R1 epitope of the HAdV-3 hexon protein replaced
with the E1 epitope from the HAdV-7 hexon protein), rAdMHE2
(R2 replaced with E2), rAdMHE3 (R3 replaced with E3), and
rAdMHE4 (R4 replaced with E4), were constructed by switching
one of the predicted epitopes within the hexon protein of
rAd
⌬
E3GFP with the corresponding epitope from the
corre-sponding HAdV-7 hexon (see
Fig. 4A
, below). Transmission
elec-tron microscope observation indicated that all four recombinant
viruses were rescued and packaged successfully in HEp-2 cells
(data not shown).
The stabilities of four chimeric hexon sequences were
exam-ined in RT-PCR assays. Total RNA was isolated from HEp-2 cells
infected with rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4 for
10 serial passages, and no virus genomic DNA contamination was
detected in PCR assays (
Fig. 2A
, lanes 20 to 25). Then, cDNA
templates were prepared, and all of them were suitable for the
RT-PCR assays (
Fig. 2A
, lanes 14 to 19). The cDNA template
gen-erated from HAdV-7-infected HEp-2 cells was used as a positive
control, and the cDNA template generated from rAd
⌬
E3GFP-infected HEp-2 cells was used as a negative control. RT-PCR
as-says indicated that all four hexon chimeric vectors remained stable
for at least 10 serial passages
in vitro
without detectable loss of
chimeric hexon sequences (
Fig. 2A
, lanes 9 to 12). No changes to
the chimeric hexon sequence were detected by sequencing the
cloned PCR fragments. The expression of chimeric hexons was
confirmed by Western blotting with the monoclonal antibody
5D4, which recognizes a common epitope of the HAdV-3 and
HAdV-7 hexon proteins (
Fig. 2B
) (
18
).
Replication efficiencies of four chimeric viruses and
rAd
⌬
E3GFP.
During the rescue and purification processes for
recombinant viruses, we observed that under the same conditions,
a shorter time was needed for rAdMHE4-infected HEp-2 cells to
reach a complete CPE than cells infected with other chimeric
FIG 1Analysis of the potential serotype-specific neutralizing epitopes for HAdV-3 and HAdV-7. (A) Modeling of 3D structures of the HAdV-3 hexon and HAdV-7 hexon and map of the potential epitopes on the 3D structures. Potential epitopes of HAdV-3 and HAdV-7 hexons are indicated at the tops of the molecules. (B) Sequence alignment of potential epitopes from HAdV-3 and HAdV-7 hexon proteins.
FIG 2Confirmation of expression of the four chimeric hexons based on RT-PCR and Western blot assays. (A) RT-RT-PCR assays. RT-RT-PCRs were performed to amplify the hexon chimeric regions by using chimeric viral cDNA, and products were analyzed by 1% agarose gel electrophoresis. Lanes 9 and 16: rAdMHE1 cDNA; lanes 10 and 17, rAdMHE2 cDNA; lanes 11 and 18, rAdMHE3 cDNA; lanes 12 and 19, rAdMHE4 cDNA. RT-PCR controls in-cluded rAd⌬E3GFP viral cDNA (lanes 1 to 4 and 14) and total RNA extracted from cells infected with rAd⌬E3GFP (lane 20), HAdV-7 viral cDNA and total RNA extracted from cells infected with HAdV-7 (lane 21), four chimeric virus cDNAs (lanes 16 to 19), and total RNA extracted from cells infected with rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4 (lanes 22 to 25). (B) Western blot assay results. Total protein extracts of HEp-2 cells infected with rAdMHE1 (lane 1), rAdMHE2 (lane 2), rAdMHE3 (lane 3), rAdMHE4 (lane 4), or rAd⌬E3GFP (lane 5) were separated by SDS-PAGE and then analyzed by Western blotting using monoclonal antibody 5D4. MK, protein molecular size
marker.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.71.256.63.355.2] [image:5.585.300.542.69.249.2]viruses or rAd
⌬
E3GFP virus, but it required 2-fold more
rAdMHE4-infected HEp-2 cells to acquire 1
⫻
10
12rAdMHE4
virus particles compared to cells infected with other viruses. We
concluded that the R4 epitope replaced by E4 from the HAdV-7
hexon protein had an impact on the efficiency of rAdMHE4
rep-lication. Q-PCR assays were performed to gain a quantitative
un-derstanding of the replication efficiencies of rAd
⌬
E3GFP and the
four recombinant viruses (
Fig. 3
). Approximately 10
8to 10
9virus
particles were used to infect HEp-2 cells. Two days postinfection,
the CPE ratios for rAdMHE4-infected cells and
rAdMHE2-in-fected cells was 100% and 80%, respectively, while the CPE ratios
for rAdMHE1-, rAdMHE5-, or rAd
⌬
E3GFP-infected cells were
very low (less than 10%). All infected cells were found to reach
CPE 3 days postinfection. In addition, the final yield of rAdMHE4
was 10 times lower than that of rAd
⌬
E3GFP, while rAdMHE1 and
rAdMHE3 replicated as well as the parental virus, rAd
⌬
E3GFP, in
HEp-2 cells. The replication efficiency of rAdMHE2 was the
high-est before 36 h postinfection, but the final yield was similar to that
for rAd
⌬
E3GFP, rAdMHE1, and rAdMHE3. These results
dem-onstrated that all of the chimeric viruses could encapsidate the
recombinant genomes, while the replication efficiency of
rAdMHE4 was lower than that of the other three chimeric viruses.
Of interest, a lower virus titer was required for rAdMHE4 than
other chimeric viruses or rAd
⌬
E3GFP to cause CPE. This may
have been be due to the replaced R4 epitope being very important
for HAdV-3 virus replication and infection.
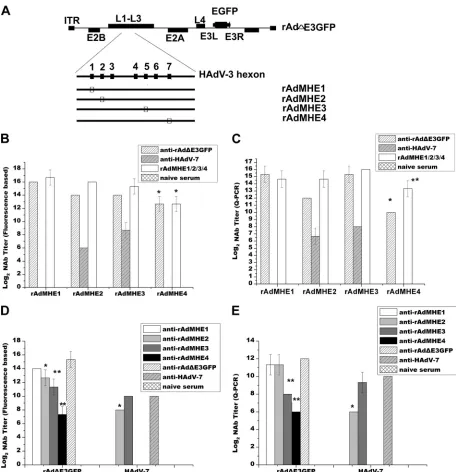
Identification of serotype-specific NAb epitopes for HAdV-7
in vitro
.
In vitro
neutralization assays were performed to detect
whether anti-HAdV-7 sera could cross-neutralize four chimeric
viruses, and the cross-neutralization responses of
anti-rAd
⌬
E3GFP sera to four chimeric viruses were analyzed too (
Fig.
4B
to
E
). The NAb titer was defined as the serum dilution that
resulted in a 70% reduction of the maximum fluorescence signal
(fluorescence signal occurs only in cells infected with viruses that
encode GFP), as assessed in a fluorescence-based
microneutraliza-tion assay as previously described (
23
,
28
). Sera from naive mice
were used as negative controls, and sera from mice preimmunized
with the four chimeric viruses were used as positive controls. With
sera from HAdV-7-preimmunized mice in neutralization assays,
high rAdMHE2-specific NAb titers (log
2titer, 6) and
rAdMHE3-specific NAb titers (log
2titer, 8) were detected, but
rAdMHE1-and rAdMHE4-specific NAb titers were not observed (
Fig. 4B
).
These data indicate that E2 and E3, but not E1 and E4, are HAdV-7
NAbs targets. With sera from rAd
⌬
E3GFP-preimmunized mice
in neutralization assays, high NAb titers were observed against all
four chimeric viruses, and the rAdMHE4 NAb titer was 2 log
2to 4
log
2lower than the NAb titers of the other three chimeric viruses
(
P
⬍
0.05, Duncan’s multiple range test) (
Fig. 4B
).
To confirm the results of the inhibition of virus infection by
measuring GFP expression, the microneutralization assays were
repeated and the chimeric virus genome copy number was
deter-mined by Q-PCR (
Fig. 4C
). The NAb titer was defined as the
serum dilution that resulted in a 90% reduction of maximum of
genome copy number (genome copies in cell infected only with
viruses). rAdMHE2-specific (log
2titer, 6) and rAdMHE3-specific
(log
2titer, 8) NAb titers, but not rAdMHE1- and
rAdMHE4-spe-cific NAb titers, induced by HAdV-7 were observed. As expected,
high NAb titers against all four chimeric viruses were observed.
The rAdMHE4 NAb titer was the lowest (
P
⬍
0.005, Duncan’s
multiple range test). These results supported those of the
fluores-cence-based microneutralization assays (
Fig. 4B
).
Further microneutralization assays were performed to determine
whether rAd
⌬
E3GFP and HAdV-7 could escape the host
neutraliza-tion response generated by four chimeric viruses (
Fig. 4D
and
E
). We
found that sera from all four groups of mice preimmunized with
chimeric virus inhibited rAd
⌬
E3GFP infection; however, there were
meaningful differences in the NAb titers. rAd
⌬
E3GFP-specific NAb
titers induced by rAdMHE4 were 8 log
2lower than those induced by
rAd
⌬
E3GFP (
P
⬍
0.005, Duncan’s multiple range test).
rAd
⌬
E3GFP-specific NAb titers induced by rAdMHE3 were 4 log
2lower than those induced by rAd
⌬
E3GFP (
P
⬍
0.005, Duncan’s
mul-tiple range test), and rAd
⌬
E3GFP-specific NAb titers induced by
rAdMHE2 were 2 log
2lower than those induced by rAd
⌬
E3GFP (
P
⬍
0.05, Duncan’s multiple range test). No statistical differences in
rAd
⌬
E3GFP NAbs titers were found between anti-rAdMHE1 sera
and anti-rAd
⌬
E3GFP sera in fluorescence-based
microneutraliza-tion assays (
P
⬍
0.1, Duncan’s multiple range test). In the
Q-PCR-based microneutralization assay, no statistical differences in
rAd
⌬
E3GFP Nabs titers were found between rAdMHE1,
anti-rAdMHE2, and anti-rAd
⌬
E3GFP sera (
Fig. 4C
). These data indicate
that rAd
⌬
E3GFP NAbs may be directed partially, but not exclusively,
against only one of the four predicted epitopes.
HAdV-7 virus does not encode GFP; therefore, CMN assays
were performed to determine whether HAdV-7 virus could
escape the host neutralizing response generated by the four
chimeric viruses (
Fig. 4D
and
E
). Anti-rAdMHE2 and
anti-rAdMHE3 sera, but not anti-rAdMHE1 or rAdMHE4 sera,
protected HEp-2 cells from HAdV-7 infection.
HAdV-7-spe-cific NAb titers generated by anti-rAdMHE2 sera were 2 log
2lower than those of anti-HAdV-7. HAdV-7-specific NAb titers
generated by anti-rAdMHE3 sera and anti-HAdV-7 sera
showed no significant differences (
P
⬍
0.05, Duncan’s multiple
range test). The Q-PCR results were in accordance with the
results of the CMN assays. These results further confirmed that
FIG 3DNA replication curves of rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, and rAd⌬E3GFP. Cells infected with rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, or rAd⌬E3GFP were harvested at 12, 24, 36, 48, 60, and 72 h, and the genome copy numbers were determined by Q-PCR with Q-PCR kits. Each experiment was repeated independently three times, and the mean values and standard deviations are shown.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.44.286.65.243.2]the predicted epitopes E2 and E3 from the HAdV-7 hexon
pro-tein neutralized epitopes for HAdV-7, and E3 played a more
important role in generating NAb responses.
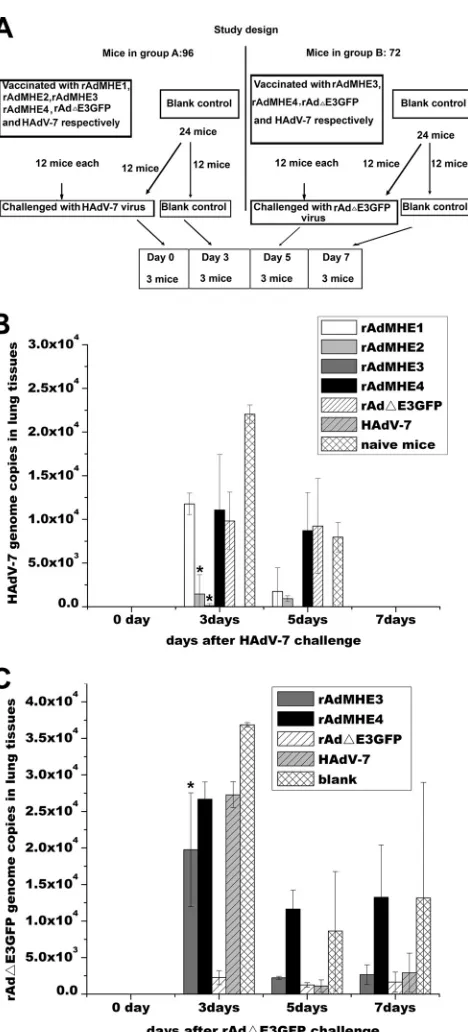
Confirmation of neutralizing epitopes for HAdV-7
in vivo
.
In vivo
neutralization assays were performed to investigate
whether the chimeric viruses could generate a host neutralizing
response against HAdV-7 and HAdV-3 infection (
Fig. 5
). The
study design is shown in
Fig. 5A
. The target organ of HAdV-3 and
HAdV-7 is the respiratory system; therefore, lung tissue was
cho-sen for the detection of adenovirus infection. Groups of mice were
immunized four times with rAdMHE1, rAdMHE2, rAdMHE3,
rAdMHE4, rAd
⌬
E3GFP, or HAdV-7. Two weeks after the last
vaccination, the mice in group A were challenged intranasally with
live HAdV-7 virus, and the mice in group B were challenged
in-tranasally with live rAd
⌬
E3GFP virus. Mice were then sacrificed
after virus infection on days 0 (before HAdV-7 or rAd
⌬
E3GFP
FIG 4Cross-neutralization responses between hexon chimeric vectors and rAd⌬E3GFP or HAdV-7. (A) Schematic depiction of four rAd⌬E3GFP-based hexon chimeric vectors. Epitopes derived from HAdV-7 (E1, E2, E3, and E4) are shown in the empty boxes. (B and C) Assessment of rAdMHE1-, rAdMHE2-, rAdMHE3-, and rAdMHE4-specific NAb titers induced by anti-rAd⌬E3GFP or anti-HAdV-7 sera. Sera from mice preimmunized by the hexon chimeric virus itself were used as positive controls, and sera from naïve mice were used as negative controls. (D and E) Assessment of rAd⌬E3GFP and HAdV-7-specific NAb titers induced by anti-rAdMHE1, anti-rAdMHE2, anti-rAdMHE3, or anti-rAdMHE4 sera. Sera from rAd⌬E3GFP- or HAdV-7-preimmunized mice were used as positive/negative controls. The neutralization assays depicted in panels B to E were performed under the same conditions. (B and D) Hexon chimeric virus NAb titers and rAd⌬E3GFP NAb titers were measured in fluorescence-based neutralization assays. HAdV-7 Nab titers in panel D were measured by CMN assay. (C and E) NAb titers were measured in Q-PCR-based neutralization assays. *,P⬍0.05; **,P⬍0.005.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.63.519.63.535.2]challenge), 3, 5, and 7, and the lung tissues were examined for
HAdV-7 or rAd
⌬
E3GFP infection.
In group A, HAdV-7 genome copies could only be detected in
the lung tissues from naïve mice or mice preimmunized with the
four chimeric viruses or with rAd
⌬
E3GFP on days 3 and 5, with
higher HAdV-7 genome copies present on day 3 than on day 5. No
HAdV-7 genome copies could be detected before HAdV-7
chal-lenge (day 0) or after HAdV-7 chalchal-lenge on day 7 (
Fig. 5B
). No
HAdV-7 genome copies were detected in the lungs of mice
preim-munized with HAdV-7 on day 3. Very low HAdV-7 genome copy
numbers (near the detection limit of the Q-PCR) were detected in
mice preimmunized with rAdMHE3 on day 3, and no genome
copies were detected on day 5; thus, preimmunization with
rAdMHE3 contributed to rapid clearance of HAdV-7
in vivo
.
HAdV-7 genome copies in mice preimmunized with rAdMHE2
were lower than those in mice preimmunized with rAdH1,
rAdMHE4, or rAd
⌬
E3GFP or in naïve mice on day 3 (
P
⬍
0.05,
Duncan’s multiple range test). Thus, preimmunization with
rAdMHE2 also contributed to the clearance of HAdV-7 in BALB/c
mice
in vivo
. In conclusion, rAdMHE2 and rAdMHE3 generate a
host neutralizing response against HAdV-7. The E2 and E3
epitopes were NAb epitopes for HAdV-7, and E3 played a more
important role in generating NAb responses; these results were in
accordance with the results of the neutralization assays
in vitro
.
Further neutralization assays
in vivo
were also performed to
investigate whether the chimeric viruses could generate a host
neutralizing response against rAd
⌬
E3GFP. We found that
preim-munization of mice with rAdMHE1 or rAdMHE2 contributed
equally to the clearance of rAd
⌬
E3GFP in BALB/c mice, while
preimmunization of rAdMHE3 or rAdMHE4 contributed little to
the clearance of rAd
⌬
E3GFP in BALB/c mice (data not shown).
To confirm these results, we repeated the neutralization assays
in
vivo
again and focused on the neutralizing responses of rAdMHE3
and rAdMHE4. As shown in
Fig. 5C
, no significant differences in
the numbers of rAd
⌬
E3GFP genome copies were detected
be-tween the mice preimmunized with rAdMHE4 versus HAdV-7 on
day 3. The number of rAd
⌬
E3GFP genome copies in mice
preim-munized with rAdMHE3 was lower than that of mice
preimmu-nized with rAdMHE4 but higher than that of mice preimmunizied
with rAd
⌬
E3GFP on day 3 (
P
⬍
0.05, Duncan’s multiple range
test). The rAd
⌬
E3GFP genome copy numbers in the lungs of mice
preimmunized with rAdMHE3 or HAdV-7 decreased to almost
the same level as that of mice vaccinated with rAd
⌬
E3GFP on days
5 and 7, while the rAd
⌬
E3GFP genome copy numbers remained
relatively high in lung tissues of mice vaccinated with rAdMHE4
and in naive mice. These data indicated that replacement of either
the R3 or R4 epitope in the rAd
⌬
E3GFP vector has a great effect on
generating the host neutralization response against rAd
⌬
E3GFP
in vivo
, and these results were in accordance with the results of
neutralization assays
in vitro
.
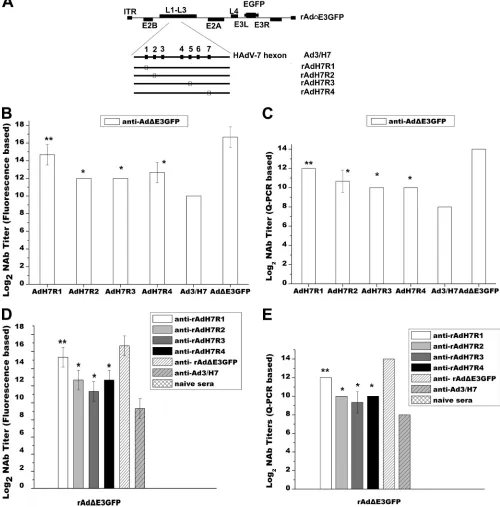
Construction of hexon chimeric adenovirus vectors by
re-placing the predicted epitopes within the HAdV-7 hexon
pro-tein with the corresponding epitopes from the HAdV-3 hexon.
The rAd
⌬
E3GFP-specific NAb responses induced by rAdMHE3 and
rAdMHE4 were weaker than those against rAdMHE1 and
rAd-MHE2. We doubted that these would play a dominant role in
gener-ating the host neutralization response against rAd
⌬
E3GFP.
There-fore, four recombinant adenovirus vectors containing HAdV-7
hexon chimeras were constructed (
Fig. 6A
). A recombinant
FIG 5Analysis of NAb responses of BALB/c mice to rAd⌬E3GFP or HAdV-7 infection generated by preimmunization with rAdMHE1, rAdMHE2, rAdMHE3, or rAdMHE4. (A) Study design. (B) Mice were preimmunized with rAdMHE1, rAdMHE2, rAdMHE3, rAdMHE4, rAd⌬E3GFP, or HAdV-7 and then challenged with HAdV-7. Q-PCR assays were performed to detect genome copy numbers of HAdV-7 in lung tissues. (C) Mice were preimmunized with rAdMHE3, rAdMHE4, rAd⌬E3GFP, or HAdV-7 and then challenged with rAd⌬E3GFP. Q-PCR assays were per-formed to detect genome copy numbers of rAd⌬E3GFP in lung tissues. Controls included mice preimmunized with rAd⌬E3GFP or HAdV-7 or naïve mice *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.47.281.60.576.2]rAd
⌬
E3GFP-based hexon chimeric vector, rAd3/H7, in which the
HAdV-3 hexon was replaced with that of HAdV-7, was constructed
in a previous study (
27
). Previous work showed that the majority of
HAdV-3- and HAdV-7-specific NAbs target their hexon proteins
(
27
). Four rAd3/H7-based, hexon chimeric virus vectors, rAdH7R1
(with the E1 epitope from the HAdV-7 hexon protein replaced by the
R1 epitope from the HAdV-3 hexon protein), rAdH7R2 (E2 replaced
by R2), rAdH7R3 (E3 replaced by R3), and rAdH7R4 (E4 replaced by
R4), were constructed as described above. All four recombinant
vi-ruses were rescued and packaged successfully in HEp-2 cells. cDNA
FIG 6Cross-neutralization NAb responses between rAd3/H7-based hexon chimeric vectors and rAd⌬E3GFP. (A) Schematic depiction of four rAd3/H7-based hexon chimeric vectors. Epitopes derived from the HAdV-3 hexon (R1, R2, R3, and R4) are shown as empty boxes. (B and C) Sera from rAd⌬ E3GFP-preimmunized mice were assessed for NAb titers specific to rAdH7R1, rAdH7R2, rAdH7R3, rAdH7R4, or rAd3/H7. rAd⌬E3GFP was used as a positive control. (D and E) Sera from mice preimmunized with rAdH7R1, rAdH7R2, rAdH7R3, rAdH7R4, or rAd3/H7 were assessed for rAd⌬E3GFP-specific NAb titers. Control sera included anti-rAd⌬E3GFP and naïve serum. The neutralization assays depicted in panels B to E were performed under the identifical conditions. (B and D) NAb titers as measured in fluorescence-based neutralization assays. (C and E) NAb titers measured in Q-PCR based neutralization assays.*,P⬍0.05; **,P⬍ 0.005.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.41.541.73.580.2]sequences in the hexon chimeric domains of the recombinant viruses
were confirmed by sequencing.
Further identification of serotype-specific NAb epitopes
for HAdV-3.
Sera from BALB/c mice preimmunized with
rAd
⌬
E3GFP, rAd3/H7, rAdH7R1, rAdH7R2, rAdH7R3, or
rAdH7R4 were prepared, and NAb titers of these serum samples
ranged from 14 log
2to 16 log
2titer in fluorescence-based
neutral-ization assays.
With sera from mice preimmunized with rAd
⌬
E3GFP,
in vitro
neutralization assays were performed to evaluate
HAdV-3-spe-cific NAb responses against four rAd3/H7-based, hexon chimeric
viruses. Sera from naïve mice were used as negative controls. As
shown in
Fig. 6B
, high rAd
⌬
E3GFP NAb titers (16 log
2) and low
rAd/H7 NAb titers (9 log
2) were detected, as described previously
(
27
). rAdH7R1 NAb titers were 4 log
2higher than rAd/H7 NAb
titers (
P
⬍
0.005, Duncan’s multiple range test). The NAb titers
for rAdH7R2, rAdH7R3, and rAdH7R4 showed no significant
dif-ference (
P
⬍
0.05, Duncan’s multiple range test), and they were
about 2 log
2higher than the rAd/H7 NAb titers (
P
⬍
0.05,
Dun-can’s multiple range test). Further neutralization assays were
car-ried out to evaluate the cross-neutralization responses of sera
from mice preimmunized with the four chimeric viruses to
rAd
⌬
E3GFP. As shown in
Fig. 6D
, the titer of anti-rAdMH1 sera
was 6 log
2higher than that of anti-rAd3/H7 sera against
rAd
⌬
E3GFP (
P
⬍
0.005, Duncan’s multiple range test). The
rAd
⌬
E3GFP NAb titers generated by rAdMH2,
anti-rAdMH3, and anti-rAdMH4 sera showed no significant
differ-ence, and they were at least 2 log
2higher than that of anti-rAd3/H7
sera (
P
⬍
0.05, Duncan’s multiple range test). As a positive
con-trol, rAd
⌬
E3GFP NAb titers generated by anti-rAd
⌬
E3GFP sera
were the highest. The Q-PCR results correlated with the results of
fluorescence-based neutralization assays (
Fig. 6C
and
E
). These
data indicate that all four epitopes, R1 to R4, are NAb epitopes of
HAdV-3, and R1 played the most important role in generating
NAb responses.
Confirmation of serotype-specific NAb epitopes for HAdV-3
in vivo
.
The functional significance of four NAb epitopes, R1 to
R4, was assessed by determining the suppression capacity of
HAdV-3-specific NAbs to the immunogenicity of the four
chime-ric viruses. Purified IgG from mice with wild-type HAdV-3 GZ-01
immunity were confirmed for HAdV-3-specific NAb activity
be-fore adoptive transfer to recipient mice. Groups of naive recipient
mice (
n
⫽
3/group) received 40
l of purified IgG by tail vein
injection before immunization with 5
⫻
10
9virus particles of
rAd
⌬
E3GFP, rAd3/H7, rAdH7R1, rAdH7R2, rAdH7R3, or
rAdMH7R4 vectors (all these viruses encode GFP). To verify
cir-culating NAb titers in these animals, serum was obtained after
adoptive transfer and immediately before vaccination. As
ex-pected, serum NAbs in the recipient mice following adoptive
transfer exhibited similar HAdV-3-specific NAb titers that ranged
from 64 to 128. Vaccine-elicited immune responses were assessed
by enhanced GFP-specific ELISAs on day 12 and day 18. As shown
in
Fig. 7B
and
C
, rAdH7R1- and rAd
⌬
E3GFP-elicited humoral
immune responses were significantly and durably suppressed by
HAdV-3-specific NAbs (
P
⬍
0.05,
t
test), while no significant
sup-pression of humoral immune responses elicited by rAd3/H7,
rAdH7R2, rAdH7R3, or rAdH7R4 were detected by
HAdV-3-spe-cific NAbs (
P
⬍
0.05,
t
test). Together, the results of the
neutral-ization assays and adoptive transfer assays show that R1 appears to
be a major NAb epitope, while R2 to R4 are relative minor NAb
epitopes for rAd
⌬
E3GFP.
DISCUSSION
The subspecies B1 adenoviruses HAdV-3 and HAdV-7 have
occurred epidemically and contribute greatly to pediatric acute
respiratory diseases (
9
,
11
,
14
,
26
). Identification of the NAb
epitopes of HAdV-3 and HAdV-7 will be beneficial for
devel-oping recombinant bivalent HAdV-3/HAdV-7 vaccines and
preventing outbreak of HAdV-3 and HAdV-7 infection. In this
study, four NAb epitopes within hexon HVRs were predicted
for HAdV-3 and HAdV-7 by bioinformatics. Eight hexon
chi-meric adenovirus vectors with the alteration of only one
pre-dicted neutralizing epitope were constructed. All of them were
rescued and packaged successfully in HEp-2 cells. The
replica-tion efficiency of four rAd
⌬
E3GFP-based hexon chimeras
demonstrated that a lower virus titer was required for
rAdMHE4 than the other three chimeric viruses (rAdMHE1,
rAdMHE2, and rAdMHE3) or the parental virus rAd
⌬
E3GFP
FIG 7Adoptive transfer of purified IgG from mice with high levels of anti-HAdV-3 immunity. Purified IgG antibody levels are shown for mice preimmunized with wild-type HAdV-3 adoptively transferred into naive recipient BALB/c mice (n⫽3/group) by tail vein injection before immunization with 5⫻109virus
particles of rAdE3GFP, rAd3/H7, rAdH7R1, rAdH7R2, rAdH7R3, rAdH7R4, or PBS (none). Vaccine-elicited immune responses following immunization were determined by EGFP-specific ELISAs at 12 days (A) or 18 days (B). Controls included mice that received adoptive transfer of naive IgG purified from saline-injected mice (naive). *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.41.540.68.217.2]to cause CPE, but the final virion yield was the lowest. We
specu-late that exchanging the R4 epitope on the surface of the HAdV-3
hexon disturbs its interaction with other cell components and
boosts cell death, and cell death in turn decreases the replication
efficiency of rAdMHE4 in later stages. A recent study indicated
that adenovirus transport occurs through a direct cytoplasmic
dy-nein-hexon interaction (
2
). Exchanging the R4 epitope on the
surface of the HAdV-3 hexon may have an adverse effect on the
dynein-hexon interaction, resulting in the retention of viruses in
the cytoplasm and cell death. However, the role of the hexon in the
CPE needs to be further investigated.
We further identified four serotype-specific NAb epitopes (R1
to R4) for HAdV-3 and two serotype-specific NAb epitopes (E2
and E3) for HAdV-7. Yuan et al. previously reported five NAb
epitopes which reside in HVR1, HVR2, HVR4, HVR5, and HVR7
of the HAdV-3 hexon based on molecular modeling (
30
). Four
NAb epitopes (R1 to R4) identified in this study overlap with four
of the five NAb epitopes the other authors reported. Considering
that HAdV-3 NAbs target partially, but not exclusively, to all four
identified NAb epitopes, further studies will be necessary to
exam-ine whether the remaining three HVRs contribute to HAdV-3 or
HAdV-7 NAb responses.
Cross-neutralization NAb assays between four rAd3/H7-based
hexon chimeric vectors (rAdH7R1, rAdH7R2, rAdH7R3, and
rAdH7R4) and rAd
⌬
E3GFP indicated that all four predicted
epitopes are NAb epitopes of HAdV-3, while R1 play the most
important role in generating NAb responses (
Fig. 6
). Further
adoptive transfer studies confirmed that R1 was a dominant NAb
epitope (
Fig. 7
). We were surprised to find that the rAd
⌬
E3GFP-specific neutralizing epitope responses induced by rAdMHE3 (R3
replaced by E3) and rAdMHE4 (R4 replaced by E4) were weaker
than those of rAdMHE1 (R1 replaced by E1) and rAdMHE2 (R2
relaced by E2)
in vitro
and
in vivo
(
Fig. 4
and
5
). In accordance, E2
and E3 were identified for HAdV-7 serotype-specific NAb
epitopes, but HAdV-7-specific NAb neutralizing epitope
re-sponses induced by rAdH7R2 (E2 replaced by R2) and rAdH7R3
were weaker than those for rAdH7R1 (E1 replaced by R1) and
rAdH7R4 (E4 replaced by R4) (data not shown). Thus, a
domi-nant NAb epitope candidate may be a good candidate for
gener-ating NAb responses, but it may not be a good switched candidate
to evade preexisting host neutralization responses generated by
parental vectors.
In vitro
neutralization assays indicated that E2 and E3 are NAb
epitopes for HAdV-7, and E3 plays a more important role in
gen-erating NAb responses. Results of neutralization assays
in vivo
were in accordance with these results. HAdV-7 virus loads
de-tected in different groups of mice preimmunized with different
viruses were significantly different on day 3. HAdV-7 virus loads
in mice preimmunized with rAdMHE2 and rAdMHE3 were
ob-viously lower than those of mice preimmunized with rAdMHE1,
rAdMHE2, or rAd
⌬
E3GFP (
P
⬍
0.05, Duncan’s multiple range
test). Thus, both E2 and E3 are good candidates for generation of
recombinant vaccines against HAdV-7 infection.
In summary, our studies have identified two serotype-specific
NAb epitopes for HAdV-7 and four serotype-specific NAb
epitopes for HAdV-3. All of them are located in hexon HVRs.
These findings will contribute not only to the development of new
vaccines, but also to the development of new gene delivery vectors.
ACKNOWLEDGMENTS
This study was supported by a foundation of bioMérieux, the National Nature Science Foundation of China (31070150), the Nature Science Foundation of Guangdong (10151009504000006), and School of Guang-zhou research projects (10A006D).
We are grateful for the kind assistance from all of our colleagues.
REFERENCES
1.Bradley RR, et al.2012. Adenovirus serotype 5-specific neutralizing an-tibodies target multiple hexon hypervariable regions. J. Virol.86:1267– 1272.
2.Choi EH, et al.2005. Adenovirus type 7 peptide diversity during out-break, Korea, 1995–2000. Emerg. Infect. Dis.11:649 – 654.
3.Chuang YY, et al.2003. Severe adenovirus infection in children. J. Mi-crobiol. Immunol. Infect.36:37– 40.
4.Crawford-Miksza L, Schnurr DP.1996. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. J. Virol.70:1836 –1844.
5.Crawford-Miksza LK, Schnurr DP.1994. Quantitative colorimetric mi-croneutralization assay for characterization of adenoviruses. J. Clin. Mi-crobiol.32:2331–2334.
6.de Jong JC, Osterhaus AD, Jones MS, Harrach B.2008. Human adeno-virus type 52: a type 41 in disguise? J. Virol.82:3809. (Letter.)
7.Ebner K, Pinsker W, Lion T.2005. Comparative sequence analysis of the hexon gene in the entire spectrum of human adenovirus serotypes: phy-logenetic, taxonomic, and clinical implications. J. Virol.79:12635–12642. 8.Eswar N, Eramian D, Webb B, Shen MY, Sali A.2008. Protein structure
modeling with MODELLER. Methods Mol. Biol.426:145–159. 9.Guo L, et al.2012. Detection of three human adenovirus species in adults
with acute respiratory infection in China. Eur. J. Clin. Microbiol. Infect. Dis.31:1051–1058.
10. Hayashi S, Hogg JC.2007. Adenovirus infections and lung disease. Curr. Opin. Pharmacol.7:237–243.
11. James L, et al.2007. Outbreak of human adenovirus type 3 infection in a pediatric long-term care facility—Illinois, 2005. Clin. Infect. Dis.45:416 – 420.
12. Jones MS, II, et al.2007. New adenovirus species found in a patient presenting with gastroenteritis. J. Virol.81:5978 –5984.
13. Knopf HL, Hierholzer JC.1975. Clinical and immunologic responses in patients with viral keratoconjunctivitis. Am. J. Ophthalmol.80:661– 672.
14. Lee J, Choi EH, Lee HJ.2010. Clinical severity of respiratory adenoviral infection by serotypes in Korean children over 17 consecutive years (1991–2007). J. Clin. Virol.49:115–120.
15. Mitchell LS, Taylor B, Reimels W, Barrett FF, Devincenzo JP.2000. Adenovirus 7a: a community-acquired outbreak in a children’s hospital. Pediatr. Infect. Dis. J.19:996 –1000.
16. Reference deleted.
17. Pichla-Gollon SL, et al.2007. Structure-based identification of a major neutralizing site in an adenovirus hexon. J. Virol.81:1680 –1689. 18. Qiu H, et al.2009. Epitope mapping and cross-reactivity analysis of the
monoclonal antibodies against hexon protein of human adenovirus type 3. Virus Res.146:58 – 65.
19. Roberts DM, et al.2006. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature441:239 –243. 20. Rux JJ, Burnett RM.2000. Type-specific epitope locations revealed by
X-ray crystallographic study of adenovirus type 5 hexon. Mol. Ther.
1:18 –30.
21. Selvaraju SB, Kovac M, Dickson LM, Kajon AE, Selvarangan R.2011. Molecular epidemiology and clinical presentation of human adenovirus infections in Kansas City children. J. Clin. Virol.51:126 –131.
22. Reference deleted.
23. Sprangers MC, et al.2003. Quantifying adenovirus-neutralizing antibod-ies by luciferase transgene detection: addressing preexisting immunity to vaccine and gene therapy vectors. J. Clin. Microbiol.41:5046 –5052. 24. Sumida SM, et al.2005. Neutralizing antibodies to adenovirus serotype 5
vaccine vectors are directed primarily against the adenovirus hexon pro-tein. J. Immunol.174:7179 –7185.
25. Sumida SM, et al.2004. Neutralizing antibodies and CD8⫹T lympho-cytes both contribute to immunity to adenovirus serotype 5 vaccine vec-tors. J. Virol.78:2666 –2673.
26. Tang L, Wang L, Tan X, Xu W.2011. Adenovirus serotype 7 associated
on November 7, 2019 by guest
http://jvi.asm.org/
with a severe lower respiratory tract disease outbreak in infants in Shaanxi Province, China. Virol. J.8:23.
27. Tian X, et al. 2011. Construction and characterization of human adenovirus serotype 3 packaged by serotype 7 hexon. Virus Res.160: 214 –220.
28. Wang Z, Mo C, Kemble G, Duke G.2004. Development of an efficient fluorescence-based microneutralization assay using recombinant human cytomegalovirus strains expressing green fluorescent protein. J. Virol. Methods120:207–215.
29. Reference deleted.
30. Yuan X, et al.2009. Molecular modeling and epitopes mapping of human adenovirus type 3 hexon protein. Vaccine27:5103–5110.
31. Zhang Q, et al.2006. Comparative genomic analysis of two strains of human adenovirus type 3 isolated from children with acute respiratory infection in southern China. J. Gen. Virol.87:1531–1541.
32. Zhang Q, et al.2009. Construction and characterization of a replication-competent human adenovirus type 3-based vector as a live-vaccine can-didate and a viral delivery vector. Vaccine27:1145–1153.
on November 7, 2019 by guest
http://jvi.asm.org/