JOURNALOFVIROLOGY, Mar.1978,p.710-718 0022-538X/78/0025-0710$02.00/0
Copyright0 1978 AmericanSocietyforMicrobiology
Vol.25, No.3 Printed inU.S.A.
Integrated
Viral
DNA
Sequences
in
Epstein-Barr
Virus-Converted
Human
Lymphoma
Lines
MARIAANDERSSON-ANVRET* ANDTOMASLINDAHLt
Departments ofChemistyandTumorBiology,KarolinskaInstitute,10401Stockholm,Sweden
Receivedfor publication 19September 1977
Most humanlymphoid celllines containmultiplecopies ofcircular, noninte-grated Epstein-Barr virus (EBV) DNA molecules as wellasviral DNA sequences
withproperties ofintegrated DNA. The
physical
state ofthe EBV DNA ina humanlymphoma line that only containsonevirusgenomeequivalentper cell has nowbeen studiedbythree differentmethods,neutralCsCldensity gradient centrifugation, actinomycinD-CsClgradientcentrifugation,
and Hirt fractiona-tion. This cellline,AW-Ramos,has beenobtainedbyEBVinfection invitro of theapparently EBV-negative Ramoslymphoma line. Theresults indicate that the EBV DNA in AW-Ramos is presentexclusivelyinalinearly integratedform. Similar data were obtained with two otherEBV-converted sublines of Ramos cells.Human lymphoid cells cannot be grown in long-term tissue culture, as a rule, unless the cells have been transformed by Epstein-Barr
virus (EBV;forreviews, see27, 38). Lymphoid celllines derived from blood ofhealthy donors
alwaysareof the B (bone marrow-derived)-cell
type,invariablycontainmultiplegenomecopies
of EBV DNA, and express an EBV-associated nuclear antigen, EBNA. Similar EBV-trans-formed B-cell lines can be obtained at a high frequency fromblood of infectious mononucle-osispatientsorfromBurkittlymphoma biopsies. Although the establishment of EBV-negative
humanlymphoid cell lines from nonmalignant
sourceshasneverbeenreported, biopsiesfrom
afew
undifferentiated,
highly malignanthumanB-cell lymphomas have yielded cell lines that appeartobe free fromEBV,asjudged byEBNA
negativity (22) and by absence of detectable
amountsof EBV DNA (3). Two such
EBV-neg-ative lymphoma lines, Ramos and BJAB, are
of particular interest because they can be in-fected with EBV in vitro and converted to
EBNA-positivesublines(7, 13,21). These
EBV-converted lines contain small but detectable
amountsof viralDNA,andonelineofthistype,
AW-Ramos, only has about one EBV genome
equivalentper cell (3).AW-Ramoscellsare
sta-bly convertedby EBV,andEBNA-negativecells
do not appear in cultures at a detectable
fre-quency.
The latent EBV DNA in typical human lymphoid celllinesismainly presentascircular
tPresentaddress:DepartmentofMedicalChemistry, Uni-versity ofG6teborg,400 33Gi6teborg,Sweden.
DNAmolecules of viral genome length (17, 23), butintegrated viral DNA sequences also seem
tobepresent (1, 2, 17). In this work, we have extended the studies on the physical state of intracellular EBVDNAtoEBV-converted sub-linesof Ramos that carrysmall amounts of viral DNA. We have used three different methods suitable for the analysis ofintegration of
high-molecular-weightviralDNA and show that such lines contain EBV DNA sequences with the expected properties ofintegrated DNA, whereas circular,nonintegrated viral DNA molecules ap-peartobeabsent.
MATERIALS AND METHODS
Cell lines.The humanlymphoma-derived B-cell
lines Raji (10), Ramos, AW-Ramos clone 2,
EHRA-Ramos (21), Ramos/B95-8 (13), and U-698 M (22) were obtained from G.Klein,Department of Tumor
Biology, Karolinska Institute, Stockholm. The cells
were grown at 370C in suspension culture in RPMI 1640medium (Grand Island Biological Co.) supple-mented with 15% fetalbovineserum,100U of penicil-lin perml,and100ug ofstreptomycin perml.Actively growing cell cultures, containing5 x 106 to 1 x 106
cells/mlandmorethan 85%livingcellsaccording to
the trypan blueexclusion test,wereused in all exper-iments. The cells wereharvested by low-speed
cen-trifugation and washed twice in0.01 Msodium
phos-phate, pH 7.4,immediately before use.
DNA preparations and hybridization proce-dures.High-molecular-weight cellular DNA for gra-dientcentrifugation experimentswasobtainedby lysis
ofacellsuspensioncontaining107cells/mlin0.01M
sodiumphosphate (pH 7.4) byaddition of0.5volume of 0.075 M Tris-hydrochloride (pH 8.5), 0.025 M
EDTA,and 1.5%sodiumdodecylsulfate. After20min
at 20°C, 0.2 volume of 0.5% Pronase (Calbiochem;
710
on November 10, 2019 by guest
http://jvi.asm.org/
self-digestedin 0.05 MTrs-hydrochloride[pH7.5]at
46°C for45minbeforeuse)wasadded,and thelysate
wasincubatedat370Cfor 8 h(25).The viscous
solu-tionwasthenbroughtto200C, diluted withanequal
volume of 0.1 M NaCl-0.075 M Tris-hydrochloride
(pH 8.0), and extracted with 1 volume offreshly redis-tilledphenol equilibratedwith the samebuffer. The phenol extraction was performed by slowly pouring
the mixtureback and forthbetween twoglassbeakers.
The mixture was then chilled to 8°C, and the two phases wereseparatedbycentrifugation.The aqueous phase wasextracted once more with phenol in the same fashion and then dialyzed against 0.1 M
NaCl-0.01 M Tris-hydrochloride (pH 8.0)-0.001 M
EDTA for 48 h at20C,followedby dialysisfor 4 h
againstthesamebuffer withoutNaCl.
Nonradioactive and 3H-labeled (105cpm/pg)EBV
DNA preparations from virus particles released by
the P3HR-1celllineweregiftsfrom A. Adams. The viral DNApreparations had more than 8.5% of the DNA in intact, 59S form. KlebsielIa pneumoniae
[3H]DNA (10' cpm/,ug, p = 1.717 g/cm3, molecular
weight=3x 107) wasprepared by standard methods.
The preparation of3P-labeled EBV
complemen-taryRNA(cRNA)and the conditions for DNA-cRNA
hybridizationwereasdescribed (23).
COsCldensity gradient centrifugation.ForCsCl
gradient centrifugation, DNA solutions were diluted
tolowconcentrations,2to6
jg/ml,
andsupplementedwith solidCsCl (MerckSuprapur) to afinaldensity
of 1.710 to 1.714 g/cm3 and a trace amount of K.
pneumoniae[3H]DNA.The DNAsolutionswere
cen-trifugedin18.5-mlsamplesinaSpinco6OTirotor at
33,000 rpmand210Cfor65h.Fractions(0.4ml)were
collected through alargehole in the bottom of the
tube with the aid ofaclosed-systemcollection device
and analyzedfor refractive index, radioactivity,
ab-sorbancy at 260 nm, and hybridizability with EBV
[32P]cRNA
asdescribed(23).FurtheranalysesofCsClgradientfractionsbyneutralglycerol gradient
centrif-ugationwerealsoperformedasdescribed(23).
Actinomycin D-CsCl gradient centrifugation.
Theexperimentswereperformedessentially according
toBinstieletal.(5).DNAsolutionswereextensively
dialyzed against0.05M sodium tetraborate (pH 9.0)
at30Cand dilutedto aDNA concentration of2
i&g/ml.
Fourteenmiflilitersof each DNAsolutionwassupple-mented with a trace amount of K. pneumoniae
[3H]DNA, 0.2 ml ofa 0.2% actinomycin D (Sigma)
solution,andsolidCsClto arefractive index of 1.3885
at30C.Thesolutionswerethenoverlaidwithparaffin
oil andcentrifuged ina Spinco 6WTi rotor at33,000
rpm and 30C for 96 h. Collection and analysis of fractionswere asforCsCl gradientswithout
actino-mycin D,except that eachfractionwasdilutedwith
an equalvolume of0.05 M sodium tetraborate (pH
9.0) before denaturationforhybridizationanalysis.
Hirtfractionationprocedure.The fractionation
wasperformedafter Pronasetreatment(16)and at a
relatively low cell concentration (4). Cells (2 x 107
living cells) were suspended in 8 ml of 0.13 M
NaCl-0.01 M sodium phosphate (pH 7.4). An equal
volume of 1.2% sodium dodecylsulfate-0.01 M
Tris-hydrochloride (pH 8.0)-0.01 MEDTA wasadded at
200C, and thesolutions were gently mixed. After 60
min, 4 mlofpreincubated 0.5% Pronase was slowly
added, followedby incubation at370C for 4 h. The
solution wasthen chilled to 0°C and gently mixed
with 6.7 ml of4 M NaCl-0.4 M Tris-hydrochloride
(pH 7.6) (at00C).After 12 h at00C,themixturewas
centrifugedat24,000xg for 45 min. Thesupernatant
wasrecovered anddialyzedagainst 0.1 M
Tris-hydro-chloride (pH8.0)-0.001 MEDTA, and the precipitate
wasdissolvedinthe same buffer at200C.The solutions
wereseparatelyextracted with 1 volume of phenol at
200C and freed from phenol by centrifugation and
dialysisof the aqueousphaseagainst 1 MNaCl-0.05
MTris-hydrochloride(pH 8.0)-0.001 M EDTA for 48
h,followed bydialysis againstthe same buffer without
NaCl. Severalsamples (10to 12,ugeach)of the DNA
solutions were thendenatured byincubation with an
equal volumeof0.5MNaOH at800Cfor 10min,and
thisprocedure also served to cleave large, covalently
closed circular DNA molecules and to degrade
con-taminatingRNA. The DNA was subsequentlyfixed
to membrane filters and analyzed by hybridization with EBV [uP]cRNA. The DNA-containing filters wererecovered after theradioactivitymeasurements, and the DNA on eachfilterwasacidhydrolyzedand
quantitated by the diphenylamine reaction as
de-scribed (23). The latter values were used to correct
thehybridizationdata to 10.0
itg
of DNA per filter. RESULTSDistribution of integratedviral DNA se-quences after CsCl
density
gradientcen-trifugation. DNA molecules of different base
compositioncanbefractionated byCsCldensity
gradient centrifugation, and this method has often been usedtoseparatefree viral DNA and host DNA andtostudytheintegrationofviral DNA (for reviews, see 9, 24). In its simplest
form, the method can be used to measure the
integration ofasmall viral genome, or a
frag-ment of a virus DNA molecule, into a large
piece of host DNA. In this case, theintegrated viral DNA sequenceswillexhibit a density very close to that of the cellular DNA, and it has been shown in this fashion that fragments of adenovirus 12DNAareintegratedinto
cellular
DNA after abortive infection ofbaby hamster
kidney cellsgrown inthe presence of bromode-oxyuridine (8), and that the DNA provirus of
spleen necrosis virus is integrated into the chicken cell genome (14). When the virus
ge-nome is large, as in the case of a herpesvirus DNA molecule, the analysis becomes slightly more complicated, because standard methods
for the preparation of high-molecular-weight
DNA from
mammalian
cellsusing Pronase andphenol treatments (33) yield shear-produced
large fragments of the host chromosome that
are of amolecularweightcloseto
10',
i.e.,similarin sizetothe viralgenome under
study.
Inthis case,integratedviralDNAsequences should be found atdensities intermediate between thoseon November 10, 2019 by guest
http://jvi.asm.org/
712 ANDERSSON-ANVRET AND LINDAHL
offree viral DNA and host DNA afterdensity
gradient centrifugation ifentire virus genomes are integrated, because random fragmentation
of hostchromosomescontainingintegratedvirus
genomes will result in the formation of "joint
molecules"comprisedofvarying proportionsof
viral and cellular DNA sequences With the assumption that the joints between viral and
cellular DNA sequences are not anomalously
shear sensitive, a typical jointmolecule might
then be comprisedofapproximately equalparts
ofviral and cellular DNA.
There isnomethod availabletoequally mea-sureallsuch joint molecules.Instead, viralDNA sequencesinjoint moleculescanbe determined
by nucleic acid hybridization, usingaradioactive
virusnucleic acid probe. Thismeansthatajoint
molecule comprised ofalongsequence of viral
DNAandashortsequenceofcellular DNA will
provide more sequencescomplementaryto the
radioactive probe than a joint molecule
com-prised ofashortsequence ofviral DNA and a
long sequence ofcellular DNA. Consequently, eveniffree virusDNAmolecules,cellular DNA
"molecules," and viral-cellular DNA joint mol-ecules are all of the same size, the profile
ob-tainedbyhybridizationwithaviral nucleic acid
probe over aCsClgradient containing cellular
DNA with integrated virus genomes will not
yield a symmetric radioactivity peak at equal
distance between viral and cellular DNA. In-stead, a skewed peak localized closer to the
positionof free viral DNAthantocellular DNA
would beexpected.
When DNA molecules isolated from
trans-formedcellsand free viralgenomes areof similar
size, the determination ofthe expected distri-bution of integrated viral DNA after density gradient centrifugation becomes formally equiv-alenttothe theoretical analysis of the density
distribution of bacteriophage T4 recombinant
DNA molecules performed by Tomizawa and Anraku(34). These authors infected cells simul-taneously with bromouracil-containing,
nonra-dioactiveT4particlesofhigh density andasmall
amountof32P-labeledT4phageofnormal
den-sity,andregistered joiningofparentalT4DNA
molecules bymeasuring the amounts of
radio-active T4 DNA foundatanomalousdensities in
CsClgradientsafter extractionandfractionation
of high-molecular-weight DNA from infected
cells. In thisconnection,anequationwasderived
toshow theexpected distribution of radioactive
residuesinjoint molecules (equation2in
refer-ence34). Here,wehaveapplied the
Tomizawa-Anrakuequationtoestimatethe expected
den-sity distribution in neutral CsCl gradients of
EBV DNA integrated into human DNA. EBV
DNA sequences can be separated from host
cellularDNA inCsClgradientsbecausealarge,
natural density difference exists between the virus DNA (p = 1.718 g/cm3) and mammalian
DNA (p = 1.700 g/cm3), whereas EBV DNA
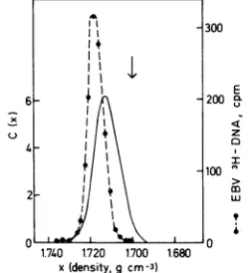
has little internal density heterogeneity (30). Figure 1 shows the experimentally determined density values for free EBV DNA from virus particles and for host DNA, aswell as the
ex-pectedtheoretical distribution ofintegrated
vi-usDNA sequencesfor thecase ofintegration
of entire EBV genomes as single entities.
Whereasthe freevirus DNA is foundas a narrow peakatadensityof 1.718g/cm3,theintegrated viral DNA would be detected as a broader,
slightly skewed profile with a peak density of
1.713g/cm3 by hybridizationwithaviral nucleic
acid probe. There are no detectable free viral
DNAsequencesatthedensityof cellularDNA, whereas a signiicant tail of integrated viral
DNAsequenceswould be found at thisdensity.
Thelatter material wouldrepresent
viral-cellu-lar DNA joint molecules mainly consistng of
cellular DNA. Several ssumptions have been
made inthisanalysis.
(i)All cellularDNAmoleculeshave thesame
density.This is notentirelycorrect,but
moder-ate density heterogeneity within the peak of
cellular DNAwould not markedly change the
theoretical distributionprofileofintegratedviral
DNA (34). It should be noted, however, that
E
z
I
ro m m
UJ
wp
V 1.740 1720 1700 1.680 V
x(density,gcm-3)
FIG. 1. Estimateofthedensitydistribution of in-tegratedEBVDNA inaneutralCsClgradient. The dashed line shows experimental data for EBV
PHIDNA,isolatedfromvirusparticles, after centrif-ugationat33,00()rpminaSpinco6i rotorfor65 hat20°Cinan185-mlneutralCsClgradient. The datacanbeapproximated byaGaussiandistribution with apeak density of1.718 g/cm3 anda = 0.004
g/cm3.Thearrowshows thepeak position ofhuman ceUDNA (1.700g/cm3)asdetermined inaseparate experiment. The solid line shows the theoretically expected distribution ofEBV DNA in theform of viral-cellularjointmolecules, c(x), as afunction of
density,calculatedaccordingtoTomizawa and
An-raku(34).Forfurther details,seetext. ' 4
I
4 I I
I
1~~~~~~1
2 /
J. VIROL.
DO
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.299.424.392.529.2]the presentestimate concerns viral DNA inte-grated into cellular DNA of typical base
com-position (p = 1.700 g/cm3). It is obvious that differentdistributionswould be expected for the unlikely cases of integration of EBV DNA into guanine-cytosine-rich DNA such as nucleolar DNA, or into satellite DNA of unusual base composition.
(ii) All DNA molecules are of the same length. Thisrequirement isessentially followed in the presentwork, as the isolated cellular DNA was ofsimilar orslightlylargersizethan intact EBV DNA (seebelow).
(iii) Fragmentation of cellular chromosomes containingintegrated viral DNA occurs by ran-dom shear-induced breakage, so that ratios of viral to cellular DNA in joint molecules range from zero to infinity, andall values occur with
equal probability.
(iv) Viral DNA is integrated in the form of entire butsingle genomes. Clearly, ifthe virus DNA isintegratedasmany smallfragments of viral DNA at multiple positions, the integrated virus DNA sequences should be found at the
density positionofcellularDNA. On the other hand, if several EBV genomes are tandemly
integrated, theywouldbe,difficulttodistinguish fromnonintegratedvirus DNAby CsCl density
gradient centrifugation.
(v) Differences in DNAmethylationbetween EBV DNA from virusparticlesand intracellular EBVDNA, ifthey exist, would notbe of such amagnitude as to cause marked differencesin
buoyantdensity(20). In support of this
assump-tion,it is noted that intracellularnonintegrated
EBV DNAmolecules,isolatedmainly as nicked circular DNA molecules, have densities very closetothat of EBV DNA from virusparticles
(17).
(vi) The radioactive viral nucleic acid probe
usedtodetect viral DNA sequencesby
hybridi-zation experiments isequally representative of
allsequences inthe EBV genome. In the present work we have used 32P-labeled EBV cRNA, made with Escherichia coli RNA polymerase (containingsigma factor),and such cRNA
prep-arationshave been showntorepresentmost or
all of the EBV genome (3). However, it is
pos-siblethatabundancedifferences between
differ-entsequencesoccurinthe cRNApreparations.
As the aboveassumptions maynotbestrictly
fulfilledinthepresent case, the theoretical
dis-tribution shown inFig.1shouldonlyberegarded
as an approximation. Nevertheless, it seems
likelythatthisestimateis aconsiderablybetter representation of the expected location oflong integrated sequences of EBV DNA than the
simple assumption that all integrated
herpes-INTEGRATED
virus DNA sequences might be found at the density of cellular DNA.
Fractionation of lymphoma cell DNA by neutral CsCl density gradient centrifuga-tion. Three different EBNA-positive sublines of the EBV-negative Ramos cell lines were used. Two of these lines, AW-Ramos and EHRA-Ra-mos, were obtainedby infection of Ramos cells with the P3HR-1 strain of EBV (21), whereas the third line, Ramos/B95-8, was obtained by infection with the B95-8 strain of EBV (13). Inagreement with previous results (3, 12), the AW-Ramos cells were found to contain ap-proximately one EBV genome equivalent per cell, as determined by DNA-cRNA hybridiza-tion, whereas EHRA-Ramos had four and Ra-mos/B95-8 had two EBV genome equivalents per cell. High-molecular-weight DNAwas pre-pared from these linesby lysisof thecells with
sodiumdodecylsulfateand
EDTA,
followedby prolongedtreatmentwithahigh concentration ofPronase, and finallytwogentle phenolextrac-tions followedby dialysis.The DNAwas
subse-quentlymixed witha traceamountofadensity marker,K.pneumoniae[3H]DNA,fractionated
byneutral CsCldensitygradient centrifugation,
and collected under conditions minimizing
shearingforces(23). Such DNA hada sedimen-tation coefficient of62S,correspondingtoa
mo-lecular weight of 1.1 x 108 (reference 11), as
determined by cosedimentation of [3H]DNA
from AW-Ramos, recovered from a CsCl
gra-dient, with phage T4 [14C]DNA in a neutral glycerol gradient.
As a control, DNA was isolated from the
extensively studied Raji cell line by the same
procedures,and EBV DNA sequences were lo-calizedby hybridization ofindividualCsCl gra-dient fractions. This Burkittlymphoma-derived
linecontains 50 to 60EBVgenomeequivalents per cell (28), and most of this virus DNA is presentascircular, nonintegratedDNA,but vi-rusDNAsequences withproperties of integrated DNA arealso present (1). The data are shown in Fig. 2. The peak of EBV DNA sequences from Raji cells is found at a density of 1.716 g/cm3, i.e., at adensity lowerthan thatof free viralDNA(1.718
g/cm3)
butat adensity higher than that expected for singly integrated viral genomes (1.713 g/cm3). Furthermore, the peakisslightlyskewed inshapeandextendsinto the
densityregionofcellular DNA. These observa-tions areconsistent with the presence ofboth
nonintegrated and integrated EBV DNA se-quencesinthesecells. Thehybridizationprofile obtained isrepresentativeof theintrinsic density of the intracellular EBV DNA, because it has
been shown (2) that the radioactiveviralDNA
on November 10, 2019 by guest
http://jvi.asm.org/
714 ANDERSSON-ANVRET AND LINDAHL
o
0L' " &
10 20 30
Fraction number
T
x
E
0X
N
15.
10z lx
uJ
0I
FIG. 2. Fractionation of high-molecular-weight
DNAfromRajicellsbyneutralCsCIdensitygradient
centrifugation.DNAwasextractedfromthe cellsby
lysis with sodium dodecyl sulfate and EDTA,
fol-lowedbyextensivetreatmentwith Pronase. The DNA wasthen extracted twice withphenol, dialyzed,mixed
witha trace amountofK.pneumoniae13HJDNAas densitymarker, diluted,andsupplementedwith solid
CsCIto afinaldensityof1.710to1.714g/cm3.Each 18.5-mlgradientcontained the DNAfrom4 x106to
6x 106cellsin 0.01 MTris-hydrochloride-0.001 M
EDTA
LpH
8.0)andwascentrifugedinaSpinco6OTi rotor at 33,000 rpm and 200Cfor 65 h. Fractionswere slowly collected through a large hole in the
bottom ofthetube, and thecellular DNA and the
densitymarker were localizedby absorbancyat260
nm
(A200
andradioactivitymeasurements.TheDNAin theindividualfractionswasthenalkali denatured and immobilized on membrane filters. Each filter
washybridizedwith6ngofEBV[2PJcRNA(8.8x
I&.cpmafter correctionfor32Pdecay),and the
back-groundof cRNAnonspecificallyboundtofilters
con-tainingheterologousDNA(80() cpm)wassubtracted.
Thegradients werelinear within the entiredensity
rangeofinterest(1.68to1.75g/cm3) asdetermined
byrefractiveindexmeasurements.Symbols:Celular
DNA of buoyant density 1.700g/cm3 (0); K.
pneu-moniae pHlDNA of buoyant density 1.717 g/cm3
(a); EBV DNA sequences(A).
component inanartifical mixture of3H-labeled EBVDNAfromvirusparticles and
high-molec-ular-weight cellularDNAbands at the density of free viralDNA inneutral CsClgradients, that theintracellular EBV DNA sequencesfromRaji
cells found at an anomalously low density re-main atthe same density inrebanding
experi-ments at low DNA concentrations, and that
suchlow-densityEBVDNAisalsofound after CsClgradientcentrifugation ofPronase-treated
celllysatesnotextractedwithphenol.
Figure3showsaneutral CsCl density gradient
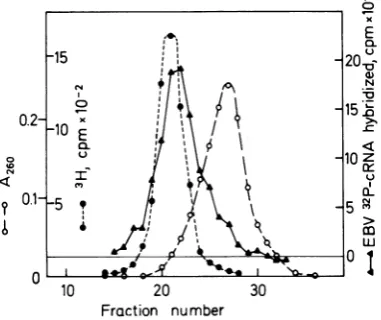
fractionation experiment with high-molecular-weightDNAfromAW-Ramoscells,which
con-tam 50timesless EBV DNA than Raji cells. In
[image:5.501.65.256.78.238.2]this case, the EBV DNA sequences are found at apeak density of 1.713g/cm3,and the hybrid-ization profileis similar to the theoretical esti-mate for an integrated EBV DNA molecule in Fig. 1. These data indicate that the viral DNA sequences in AW-Ramos are present in inte-grated form. The results do not seem compatible with a situation in which AW-Ramoscells con-tain onenonintegratedEBV DNAcircle percell
of density of 1.718
g/cm3.
In agreement withthisnotion, no viralDNAwith the sedimenta-tion properties of covalently closed circular DNA of viral genome size was detected when the AW-Ramos DNA banding at a density of 1.710 to 1.718
g/cm3
inCsClgradients was fur-ther fractionated by neutral glycerol gradientcentrifugation and analyzed by hybridization
(datanot shown). Moreover, the viral DNA in AW-Ramos cells is notpresent in the form of many smallfragments integrated at several lo-cations,as thereare few EBV DNA sequences
foundatthedensityofcellularDNA.The data donotdistinguishbetween thepossibilitiesthat AW-Ramos contains one integrated complete EBVgenome or a couple oflargefragments of
viralDNA.AW-Ramos cells, incontrast to Raji
cells,cannotbeinduced to express EBV antigens associated with an abortivelyticvirus cycle by
iododeoxyuridine treatment (21), and it is not knownifallsequences of the EBV genome are present in AW-Ramos.
When DNA from AW-Ramos cells was re-duced in sizeby shear treatment to a molecular weight of 6 x 106 before CsCl density gradient
centrifugation, the EBV DNA sequences were instead foundasapeakat1.718
g/cm3,
i.e., thedensityof freeviral DNA (3). This showsthat
0
0.6-6 0
N
0
0--0.~~~~~~~~~1
--10 20 30
Fraction number
FIG. 3. FractionationofA W-Ramos DNA by neu-tral CsCl density gradient centrifugation. Experi-mental conditions andsymbolsasinFig.2.
J. VIROL.
0
XD
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.272.461.470.624.2]715
the low density observed for the intracellular
EBVDNA after fractionation of high-molecular-weightDNA from AW-Ramos cells is notdue toanunusualbase
composition
of the viral DNA sequences presentinthecells.Results similar to those obtained with AW-Ramos were also observed on fractionation of
high-molecular-weightDNAfrom the two other EBV-convertedRamossublines,EHRA-Ramos andRamos/B95-8,by CsCl density gradient
cen-trifugation. In both cases, the EBV DNA se-quences were detected as broad hybridization profiles at densities markedly lower (1.710 to 1.714g/cm3) than that offreeEBVDNA (data notshown).
Actinomycin D-CsCl gradient centrifu-gation. InCsClgradientscontaining actinomy-cin D, guanine-cytosine-rich DNA bands at a lower density thanadenine-thymine-rich DNA (5, 18). Consequently, integrated EBV DNA
would be expected to band at a higher density than free EBV DNA insuchgradients, and the
distribution ofintegratedviral DNAsequences would be approximated by a mirror image of Fig. 1.
When high-molecular-weight DNA from an EBV-negative humanlymphoma linewasmixed
with small amounts of EBV DNA from virus
particles andK.pneumoniae[3H]DNA,the
lat-terDNAserving asdensity marker,theresults shown in Fig.4were obtained by actinomycin
D-CsCl gradient centrifugation. Although K.
pneumoniae DNA and EBV DNA have very similar base compositions, the EBV DNA banded at a slightly higher density than the
bacterialDNA,presumably becauseof sequence effects. As expected, the human cellDNA was foundat amarkedly higherdensity than either of the two more guanine-cytosine-rich DNAs. The properties of high-molecular-weight AW-RamosDNA,centrifuged togetherwith K.
pneu-moniaeDNA in thesamefashion,areshownin
Fig. 5. The separation between the bacterial DNA and the human cell DNA is similar to that observed in the controlexperiment. How-ever, theintrinsicEBVDNA sequences of AW-Ramos arefound at asignificantly higher
den-sitythan that of free viral DNA in this type of gradient. These data are in agreement with those
showninFig.3andprovide additional evidence
for the presence ofintegrated EBV DNA
se-quencesinAW-Ramos cells.
Hirt fractionation procedure. DNA from
smalltumorvirusescanbeefficiently separated fromhigh-molecular-weight cellular DNA in cell lysates by coprecipitation of the latter DNA
withsodiumdodecyl sulfate in cold 1 M NaCl. The relative sizes of the DNA molecules are clearlyimportant forthefractionation, and the
0I
4
n
Q
E
a-C-)
0)
N
.n
10
->
z
5DL
N
m
uJ
0
1
10 20
[image:6.501.253.444.58.260.2]Fraction number
FIG. 4. Actinomycin D-CsClgradient centrifuga-tion ofhuman DNA from an EBV-negative
lym-phoma line, U-69a, mixed with trace amountsof
EBV DNAfromvirusparticksandK. pneumoniae
pHJDNA. A 30-pg amount ofthe
high-molecular-weight DNA was dialyzed against 0.05M sodium
tetraborate(pH9.0),suppkmentedwithactinomycin
D andCsCl,andcentrifugedin afinal volumeof19 ml in aSpinco6OTi rotorfor4days at33,000rpm and3°C. Fractions (0.4ml) were coUectedfromthe bottom of the tube, followed by measurements on
individualfractionsof refractive index, absorbancy
at 260nm(A2,3Hradioactivity,andhybridizability
withEBVf2PJcRNA.SymbolsasinFig.2.
method is also applicable to Pronase-treated
lysates (16). It has recently been found that
molecules as large as intact herpesvirus DNA
can be separated from cellular DNA in this fashion (4, 29).Here, we have used this proce-dure to obtain partial separation of
noninte-grated EBV DNA from human cellular DNA. In control experiments with phage T4
[14C]DNA,which issimilar in sizetoherpesvirus DNA, added in trace amounts to Pronase-treatedlysates ofhumanlymphomacells, con-siderabletrappingofthe T4 DNAwasobserved.
Using a
relatively
low cell concentration(106
cells/ml),which has beenreported tofacilitatethe separation of herpesvirus DNA from cell DNA (4),weobtained 13 to15% of the cellular
DNAand50 to60% of the T4 DNA in the "Hirt
supernatant." It is clear that the presence of
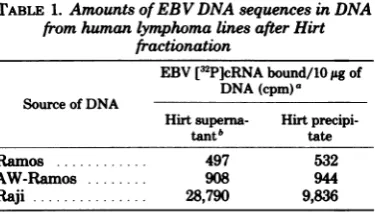
viral DNAin the "Hirt precipitate" cannot be takenasevidence forintegrationwhenthevirus genome hasa molecularweightof108, but it is alsoclear that theT4DNAwasenriched in the Hirtsupernatant. Table1shows that whenRaji
cellDNA wasfractionatedinthesamefashion,
theHirtsupernatant wasthreefold enrichedin
EBV DNAsequences. These data indicate that
on November 10, 2019 by guest
http://jvi.asm.org/
716 ANDERSSON-ANVRET AND LINDAHL
o 0. N
q 0.
0
10 20
[image:7.501.65.254.54.260.2]Fraction number
FIG. 5. Analysis of intrinsic EBV DNAsequences
in AW-Ramos cells byactinomycin D-CsCl gradient centrifugation. ExperimentalconditionsasinFig.4
andsymbolsasinFig.2.
some nonintegrated EBV DNA is present in
Raji cells. There is much better evidence for
this notion obtained by othertechniques (23),
and the EBV DNA present in the Hirt
super-natantfromRajicellshas beenfurtherpurified
and shown to consist largely of circular viral
DNA molecules (data not shown). Incontrast,
therewas nodifference between theproportion
ofEBV DNAtocellular DNA betweenthe Hirt
supernatant andthe corresponding precipitate
fromAW-Ramoscells(Table 1). These dataare
again consistent with the absence of
noninte-gratedEBV DNA molecules inAW-Ramoscells.
Furthermore, the results indicatethat the EBV
DNAsequencesinAW-Ramoscellsarenot
pres-ent in the forn of circular DNA molecules of
viralgenomesizecomprisedof bothcellularand
viral DNAsequences.
DISCUSSION
Several differentmethodshave been usedto
study the integration oftumor virus DNA
se-quences in virus-transformed host cells, and
thereare nowmanyexcellenttechniques
avail-abletoconvincinglydemonstrate theintegration
ofsmall tumor virus genomes or fragments of
virus DNA (6, 8, 15, 31,36,37). Unfortunately,
most ofthese methods depend on the smaller
size of the tumor virus genomes under study
than ofgentlyisolatedcellular DNA, and they
arenotreadilyapplicabletothe characterization
of intracellular forms ofhigh-molecular-weight
viral DNA. In that case, nonintegrated viral
genomesmaybe foundas amixture of covalently
closedcircular DNA and nicked circular DNA, and one orthe other of these forms islikelyto havefractionation propertiessimilartothose of
thehost DNA in different types ofcentrifugation
experiments. Inaddition, methodsthat involve
precipitationstepstoseparate viral and cellular
DNA are marred by trapping artifactsthat do
notoccurtoasignificantextentwith small DNA
molecules. When alargenaturaldensity
differ-ence between the viral DNA and host DNA
exists, asinthepresentcase,neutral CsCl
den-sity gradient centrifugation is a useful method
forsearchingforlarge viral DNA molecules in
integratedform. Alkaline CsClgradientsareless
satisfactoryfor suchstudieson
high-molecular-weightDNA because of theslow,salt-promoted hydrolysisof DNA thattakesplace in that sol-vent (19, 35). Inthe presentwork,the analysis of EBV-converted lymphoma lines that carry
relativelysmall amountsof EBVDNAby
neu-tral CsClgradient centrifugation yieldedresults
that are strongly indicative of the presence of
integrated viral DNA sequences in the cells,
whereasthe data donotseemcompatiblewith
the occurrence of intracellular nonintegrated viral DNA. Similar results were obtained by
centrifugation of AW-Ramos DNA in
actino-mycin D-CsCl gradients. Although the latter
methoddoesnotseemtoofferanygainin
sen-sitivityoverregularCsClgradient centrifugation
inthepresentcase(Fig.3and5), the dataserve
to confirm that the anomalousdensity
proper-tiesof theintracellular viral DNAsequencesare
due to an association with DNA of relatively
lowerguanine-cytosinecontent.
The EBV-converted sublines of the Ramos
lymphomaline havedifferentgrowth
character-istics andsurvive much better in crowded
sus-pensioncultures than the parent line (32), and
theyalsoexpressEBNAincontrast tothe
par-TABLE 1. AmountsofEBV DNA sequences in DNA
fromhumanlymphomalinesafterHirt fractionation
EBV[32P]cRNAbound/10jigof
DNA(cpm)a
Source of DNA
Hirtsuperna- Hirt
precipi-tantb tate
Ramos ... 497 532
AW-Ramos ... 908 944
Raji... 28,790 9,&36
aAverage values of
triplicate
determinations. Each0.3-mlreaction mixture contained2.5ng(370,000 cpm) of EBV[3P]cRNA.
bThe Hirtsupernatantfractions contained14± 2% of the total cellular DNA, using a low initial cell concentration andPronase treatmentbefore fraction-ation.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.272.460.486.593.2]25,1978
entline,sofrom thispointof viewlines suchas
AW-Ramos may be regarded as
"EBV-trans-forned." On the otherhand, the parental,
ap-parently EBV-negative Ramoslymphoma cells havealready beentransformed in an unknown event, since they grow indefinitely in culture and show malignant potentialoninjectioninto nude mice (21).Toavoidanomenclature prob-lem,the term "EBV conversion" has been used inthe present work todescribe the stable alter-ation ofpropertiesobserved after EBVinfection ofanEBV-negative lymphoma line.
Theintegration of EBV DNAinto the DNA of human lymphocytes does not appear to be
sufficient by itself for transformation to
malig-nancy, since small amountsofintegratedEBV DNA sequenceshave been found innewly estab-lished human lymphoblastoid cell lines of dip-loidkaryotypethatdonot cause tumorsinadult nudemice(17, 26).It may be necessarytostudy
the sites of viral DNA integration to evaluate the transforming potential of EBV, and such
measurements will have to await the
develop-ment of suitable
technology
fordetailedchar-acterizationof
integrated
sequences oflarge
viral DNAmolecules.ACKNOWLEDGMENTS
We thankBirgittaMoller andUlfBrodin oftheKarolinaka InstituteComputerCenterforwritingaFortran program of theTomizawa-Anrakuequation.
This work was supported bygrants from the Swedish
CancerSocietyand theSwedish Natural ScienceResearch
CouncilandbyPublicHealthServicecontractsNO1CP33316 and NOI CP81020within the Virus CancerProgramof the
U.S.National CancerInsttute.
LITERATURE CiTED
1. Adams,A., andT.Lindahi.1975.Intracellularforms of
EBVDNAinRajicelis,p. 125-132. In G. deThM,M.
A.Epstein,and H. zurHausen(ed.), Oncogenesisand
herpeaviruse II,part1.InternationalAgency for
Re-searchonCancer,Lyon.
2. Adams, A., T.Lindahi, and G. Klein. 1973. Linear association betweencellularDNAand EBV DNA in a humanlymphoblastoidcell line. Proc.Natl.Acad.Sci. U.S.A.70:2888-2892.
3. Andersson,M., and T. Lindali. 1976.EBV DNA in
human lymphoidcell lines:invitro conversion.Virology
73:96-105.
4. Ben-Porat,T., A. S. Kaplan, B. Stehn, and A. S. Rubinstein.1976.Concatemeric forms ofintracellular herpesvirusDNA.Virology 69:547-660.
5. Birnstiel, M,J.Telford,E.Weinberg,andD.
Staf-ford. 1974.Isolation andsomepropertiesofthegenes
codingfor histoneproteins.Proc. Natl.Acad. Sci. U.S.A. 71:2900-2904.
6. Botchan,M.,W.Topp,and J.Sambrook.1976. The arrangement ofSV40 sequences in the DNA of trans-formedcells.Cell9:269-287.
7. Clements,G.B.,G.Klein,and S.Povey.1975. Produc-tion by EBV infection of an EBNA-positivesubline from an EBNA-negativehuman lymphoma cell line
withoutdetectableEBVDNA.Int.J.Cancerl6:125-133.
8. Doerfler,W.1970.Integrationofthe DNA ofadenovirus
INTEGRATED EBV DNA
type 12into the DNAofbaby hamsterkidneycells.J. Virol. 6:652-666.
9. Doerfler, W. 1975. Integration of viral DNA into the host genome.Cun.Top.Microbiol.Immunol.71:1-78. 10. Epstein,M.A.,Y. M.Achong,Y.Barr,B.Zajac,G.
Henle,and W. Henle. 1966.Morphologicaland
viro-logical investigations oncultured Burkitt tumor lym-phoblasts (strain Raji).J.Nati.Cancer Inst. 37:547-559. 11. Freffelder, D. 1970. Molecular weights of DNA from
bacteriophagesT4, T5 andT7 and thegeneral problem
of determination of M. J.Mol. Biol. 54:567-577. 12. Fresen,K. O.,B.Merkt, G.W.Bornkamm,andH.
zurHausen.1977.Heterogeneityof EBVoriginating
fromP3HR-1cells. I.StudiesonEBNA induction.Int. J.Cancer19:317-323.
13.Fresen, K. O., and H.zurHausen.1976.Establishment
ofEBNA-expressing cell lines byinfection of
EBV-genome-negative human lymphomacellswith different
EBV strains. Int. J.Cancer17:161-166.
14. Fritsch, E., and H. KL Temin. 1977. Formation and
structureof infectious DNA ofspleennecrosisvirus. J. Virol. 21:119-130.
15.Groneberg, J.,Y.Chardonnet,and W.Doerfler.1977. Integratedviral sequences inadenovirus type 12-trans-formedhamster cells.Cell10:101-111.
16.Hirt,B.1967.Selective extraction ofpolyomaDNAfrom
infectedmousecellcultures. J. Mol. Biol. 26:365-369. 17.Kaschka-Dierich, C.,LFalk, G. Bjursell,A.Adams, andT. Lindahl. 1977. Humanlymphoblastoidcell lines derived from individuals withoutlymphoproliferative
disease contain thesamelatent formsof EBV DNAas those found intumorcells. Int.J. Cancer 20:173-180. 18.Kersten, W.,H. Kersten, and W. Szybalski. 1966.
Physicochemical propertiesofcomplexesbetween DNA and antibiotics which affect RNA synthesis (actino-mycin, daunomycin, cinerubin, nogalamycin, chromo-mycin, mithrachromo-mycin, and olivomycin). Biochemistry
5:236-242.
19. Kiger,J.A.,E. T.Young,and R. LSinsheimer.1968. Purification and properties of intracellular lambda DNArings.J.Mol. Biol. 33:395-413.
20. Kirk,J. T.0. 1967. Effect ofmethylationofcytosine
residueson thebuoyant densityof DNA incaesium
chloride solution. J.Mol.Biol. 28:171-172.
21. Klein, G.,B.Giovanella,A.Westman,J.S.Stehlin, andD.Mumford.1975.AnEBV-genome-negativecell
line established fromanAmerican Burkittlymphoma;
receptorcharacteristics,EBVinfectibility,and perma-nentconversion intoEBV-positivesublinesbyin vitro infection.Intervirology5:319-334.
22. Klein, G.,T.Lindahl,M.Jondal,W.Leibold,J. Me-nezes, K.Nilsson,andC.Sundstrom.1974. Contin-uouslymphoidcell lines withcharacteristics of B cells
(bonemarrow-derived), lackingthe EBV genome and derived from three human lymphomas. Proc. Natl. Acad.Sci. U.S.A.71:3283-3286.
23.Lindahl, T.,A.Adams,G.Bjursell,G.W.Bornkamm, C.Kaschka-Dierich,andU.Jehn.1976.Covalently
closed circularduplexDNA of EBV inahuman lymph-oidcell line. J. Mol.Biol.102:511-530.
24. Martin,M. A., andG. Khoury. 1976. Integration of
DNAtumorvirus genomes. Curr.Top.Microbiol. Im-munol.73:35-65.
25. Miller,R.C.,Jr.1975.Replicationand molecular recom-bination ofT-phage.Annu.Rev. Microbiol.29:355-376. 26. Nilsson, K.,B.Giovanelia,J. S.Stehlin,andG. Klein. 1977.Tumorigenicityof humanhematopoieticcell lines inathymicnude mice.Int. J. Cancer19:337-344. 27. Nilsson, K., and J. Pont6n. 1975. Classification and
biologicalnatureofestablished humanhematopoietic
cell lines.Int.J. Cancer15:321-341.
28. Nonoyama, M., and J. S. Pagano. 1973. Homology
betweenEB virus DNA and viral DNA from Burkitt's 717
on November 10, 2019 by guest
http://jvi.asm.org/
718 ANDERSSON-ANVRET AND LINDAHL
lymphomaand nasopharyngeal carcinoma determined byDNA-DNA reassociation kinetics. Nature (London) 242:44-47.
29. Pater,M.M., R. W. Hyman, and F. Rapp. 1976.
Isola-tionof herpessimplex virus DNA from the "Hirt
su-pernatant." Virology 75:481-483.
30.Pritchett, R. F., S. D. Hayward, and E. Kieff. 1975. Comparative studies of the DNA of Epstein-Barr virus fromHR-1 and B95-8cells.J.Virol. 15:556-569.
31. Sambrook,J., H. Westphal, P. R.Srinivasan, and R. Dulbecco. 1968. The integratedstateof viral DNA inSV40-transformed cells. Proc. Natl. Acad. Sci. U.S.A. 60:1288-1295.
32. Steinitz, IL,and G. Klein. 1975. Comparison between
growth characteristics ofanEBV-genome negative
lym-phoma line and its EBV-converted subline in vitro. Proc.Natl. Acad. Sci. U.S.A. 72:3518-3520.
33. Thomas,C. A., K. L Berns, and T. J. Kelly. 1966.
Isolation ofhigh molecular weight DNA from bacteria andcellnuclei.Procedures NucleicAcid Res. 1:536-0.
34. Tomizawa, J. I., and N.Anraku.1964.Molecular mech-anisms ofgenetic recombination in bacteriophage. II.
Joining ofparental DNA molecules of phage T4. J. Mol. Biol. 8:516-540.
35.Tomizawa, J.I.,and N.Anraku.1965.Molecular mech-anisms ofgenetic recombination in bacteriophage. IV. Absence ofpolynucleotide interruption in DNA of T4 and A phageparticles, with special referenceto heter-ozygosis. J. Mol. Biol. 11:509-527.
36. Varmus, H.E., P. K. Vogt, and J. M. Bishop. 1973. Integration of DNA specific for Roussarcoma virus after infection ofpennissive and non-permissive hosts. Proc.Natl. Acad. Sci. U.S.A. 70:3067-3071.
37.Wall, R.,J.Weber,Z.Gage,andJ. E.DarnelL 1973. Production of viralmRNAinadenovir-transformed
cellsbythepost-transcriptionalprocesingof
hetero-geneousnuclearRNAcontaining viral and cellular se-quences.J.Virol.11:953-960.
38.zurHausen,H.1975.Oncogenic herpesviruses. Biochim.
Biophys. Acta 417:25-53.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/