Vol.40,No.1 JOURNALOF VIROLOGY, Oct.1981,p.45-55
0022-538X/81/100045-11$02.00/0
Fv-1
Host Restriction
of Friend Leukemia Virus: Analysis of
Unintegrated
Proviral DNA
JEFFREY CHINSKY' AND RUY SOEIRO'-*
Departments ofMedicine2andCellBiology',AlbertEinsteinCollege ofMedicine,Bronx, New York 10461 Received 6 March 1981/Accepted 15 June 1981
The murine gene Fv-1 predominantly controls the outcome of infection by
murineecotropic retroviruses. The inhibition of virus replicationby theFv-1gene
product has been determined to be at an early stage in virusreplication.
Mecha-nistically, its effectappears tobe on the accumulation of unintegrated proviral
DNA oritsintegration or both. We investigated the synthesis of unintegrated
proviral DNA, usingseveral clones of B-, N-,orNB-tropic Friend murineleukemia
virus. Our results indicate that the accumulation of B-tropic proviral DNA in
NIH cellsmaybeinhibited ateither the level of linear (form III) orcovalently
closed circular DNA (forn I), dependingupon the degree of restriction of the
clone of virus used. We confirmed that there isaneffect of theFv-1gene on the
accumulation of form I DNA of eitherB- or N-tropic Friend murine leukemia
virus. However, the decrease in infectiouscenterseffected by the Fv-1 genedid
notcorrelatequantitatively with the effectonform Iproviral DNA produced by
N-tropic Friend murine leukemia-virus in nonpermissive cells. Lastly, we
dem-onstrated in nonpermissively infected NIHcells thatarapidly migrating doublet
of viral DNA is formed.
ThemurinegeneFv-1 isanautosomal
domi-nantwhichexerts amajor controloverthe
out-comeof infection of micebymostmurine
retro-viruses(3, 14; forareviewseereferences6 and
12). This gene, whose phenotype canbe
dem-onstrated in tissue culture, confers resistanceto
thereplication of retroviruses. Thegenehastwo
codominant alleles, Fv-ln and Fv-1
b.
Fv-1nhomozygousanimalsarepermissive for N-tropic,
but resistant to B-tropic, murine leukemia
vi-ruses(MuLV's). Fv-lb1b animalsarepermissive
for B-tropic, but nonpermissive for N-tropic,
viruses. Most murine virusesmay beclassified
as either N- or B-tropic, according tothe host
range they exhibit. However, some
laboratory-derived ecotropic viruses exhibit NB-tropism,
whichis definedasthe abilitytoreplicate well
incells ofanyFv-1 type.
Thedegreeof host resistanceconferred by the
Fv-1geneis relative (3);thatis,a50- to
10,000-fold decrease in infectious centers is observed
whenagivenvirus infectsapermissiveversus a restrictive host.The degree of restriction, which
dependsupon the exact virus andhostcellused,
may showeither single- or two-hit kinetics on
thenonpermissivecelltype (1, 7, 15, 19).
The replication of virus in a restrictive as
comparedwith apermissivehost wasstudied for
bothearlyandlateeventsin the virus cycle. It
was shown in our laboratory, as well as
else-where, thatearlysteps,suchasadsorption,
pen-etration, and initiation ofproviralDNA
synthe-sis,are unimpaired in the restrictive host (5, 8,
11,23).However,biological studies indicate that
the Fv-1 restriction effectoccursduring theearly
phase of the viralreplication cycle (2, 24).
Fur-thermore,ourunpublishedstudies indicate that
if an infectious center is established by a
re-stricted virus, there is no restriction of the
expression of either viralproteinsorRNA. Our
conclusion, therefore, is that the Fv-1gene
prod-uct acts at anearlyphase in viralreplicationto
decrease the efficiency ofestablishingan
infec-tiouscenter.
Weand others have shown that the
integra-tion ofproviralDNA afterexogenousinfection
issuppressedduring Fv-1 host restriction(8, 23).
Thestudies ofYangetal. (26)andofJolicoeur
and Rassart (9), suggestedthatthe
Fv-1-medi-ated inhibitionofproviralDNAintegrationmay
be due to an inhibition of the
production
oraccumulationorboth of form I circular
proviral
DNA, thepresumedprecursortotheintegrated provirus.
We studiedthe earlyforms ofnonintegrated
proviral DNAproduced during acute infection
byclonesof either B-tropic orN-tropicFriend
MuLV (F-MuLV). Ourresults indicatethat,
de-pending upon the clone of B-tropic F-MuLV
used, restriction in NIH cells (Fv-1
f/f)
may be45
on November 10, 2019 by guest
http://jvi.asm.org/
observed at either oftwosteps ofproviralDNA
accumulation. That is, highly restricted clones failed to accumulate linear proviral DNA, whereas less restrictedvirusshowedadecrease only in form I circular DNA. Secondly,
N-tro?ic
F-MuLV infection of BALB/c cells (Fv-116)
manifested a decrease in the level of circular DNA accumulated. However, the decrease in the level of circular DNA found in the restrictive infection did not correlate quantitatively with the decrease in the number of infectious centers formed.(The data in this paper are from a thesis to be submitted inpartial fulfillment of the require-ments for the Ph.D. degreein the SueGolding
Division of Medical Sciences, Albert Einstein College ofMedicine, Yeshiva University.)
MATERIALS AND METHODS Cells.NIH, BALB/c,and XC cellswereobtained from Janet Hartley and Wallace Rowe. They were
maintained inmonolayer culture withJoklikmodified Eagle medium containing 10% fetal calf serum and nonessential amino acids. DBA/2cells were the gen-erous gift of Robert Bassin and weremaintained in monolayer culture with Dulbecco modifiedEagle
me-diumcontaining10%ofetal calfserum.
Viruses.N-tropicandB-tropic F-MuLVwere ob-tained from Frank Lilly ashomogenates ofspleens
from infected NIH andBALB/c animals,respectively.
The viruscomplex containingboth thespleen
focus-forming virus, as well as the replicative competent helperF-MuLV, waspassed in tissue cultureathigh
dilutioninFv-1permissivemousefibroblastcell lines. Progeny viruswassubsequentlypassed in uninfected fibroblastcellstoeliminate the spleenfocus-forming
virus component. The N and B typelymphatic leu-kemia virus components (F-MuLV) were then cloned twicebylimitingdilution (using XC assay[18]),and thecloned viruswaspassagedtohightiter andfrozen inaliquots.
Virus infection. Cloned virus passaged to high
titer was usedtoinfect permissivesubconfluent mouse fibroblasts. Virusaliquotswerequicklythawed, passed through Millex HA 0.20-,um filter units to remove virus clumps, and titers were determined by direct XC plaque assay.Similarly,processed aliquots were used forexperiments. Confluent mouse fibroblastcelllines weretrypsinized and seeded at adensity of1 x 106 cellsper 100-mmdish or 3 x 106 to 5x 106cellsper T150 flask (Falcon Plastics, Oxnard,Calif.)inmedium containingPolybrene at5
jig/ml.
At 9to 12 h later, the cellswere infectedwith appropriate amounts of virusin mediumcontaining Polybrene (5iLg/ml).At thistime,asample dishwastrypsinized, andthe cellswere counted to ensure anaccurate multiplicity of infection(MOI) determination.
Isolation of viral DNA.Atappropriatetimes after infection withvirus,cellsweretrypsinized,rinsed with serum containing medium, centrifuged into apellet,
andsuspendedin10mMTris-hydrochloride (pH 7.5)-10mM EDTAat <2.0 x 106cellsper ml.Cellswere
heatedat65°C (personal communication,AnnSkalka)
for 15 min, then lysed in 1.0% sodium dodecyl sulfate at65°C for 20 min. NaCl was added to 1 M (4), and thesampleswerekept at4°C overnight. Samples were
centrifugedat12,000rpm in aSorvallSS 34 rotor, and the supematantswereextracted withphenol and chlo-roform-isoamyl alcohol (24:1) and precipitated with 2 volumes of ethanol at -20°C.
Agarose gelelectrophoresisandblot hybridi-zation. DNA samples wereseparated byhorizontal electrophoresis in 0.8% agarose slabgelssubmerged2
to 3mmunder therunningbuffer(0.089MTris-0.082 Mboric acid-0.002 M EDTA). The gel was stained with 1,ug of ethidium bromide per ml in 0.5 M am-monium acetatefor Polaroid photography under UV illumination, and the DNA wastransferred to nitro-cellulose by the Southern transfer procedure (22). Pretreatment with0.15M HClwas used forpartial depurinationof DNAtoenhance the transfer of the larger fragments from the gel (25). Viral DNA trans-ferred to nitrocellulose paper was detected by molec-ularhybridization to 32P-labeled F-MuLV DNA
pre-paredby nick translation (17) of F-MuLV DNA cloned into the plasmid vector pBR322, a generous gift of A.
Oliff (13). The specific activities routinely obtained
were 0.5x 108to 2.0 x108 cpm/yg of DNA. Hybridi-zationof the nitrocellulose-boundviral DNA to the
[32P]DNAprobewasperformedin 10% dextran sulfate (25), 5xSSC (lx SSC=0.15MNaCl,0.015Msodium
citrate),50%formamide,0.01Mphosphate buffer (pH 7.0), and100jigof salmon sperm DNA per ml at 42 to
44°Cfor 12to 16 h.This wasfollowed by extensive
sequential washingsat650Cin3xSSC,lxSSC, 0.4x
SSC, and 0.2x SSCcontaining0.2%sodiumdodecyl sulfate. Filterswereairdried andexposedat-700Cto
KodakX-Omat Rfilm, usingaPickerMAX-1
inten-sifyingscreen.
Quantitationof viral DNA. The band intensities of theautoradiogramsweremeasured with a double-beamrecording microdensitometer (MKIII C; Joyce, Loebl and Co., Ltd., England). Several
autoradi-ographic exposures ofeach hybridizedblotwere ob-tainedsothatasingle wedge settingonthe microden-sitometer could be usedtocompareadjacentlanes for bands representing both forms I and III DNA. The
backgroundexhibitedonthemicrodensitometry
trac-ing was thencarefullyconsidered, and the peak area representing either form III orform I wascutfrom thetracingandweighedon ananalyticalbalance (Met-tler).Itwasdemonstratedthat,withinawell-defined range,the microdensitometer tracing method gavea
linear responsetodifferentamountsofhybridizedviral DNA, similar to the standardizationprocedures
de-scribed by Keshet et al. (10). Therefore, only those peak weights obtained from asingle autoradiogram whichfellwithinthisdefined rangewerecompared.
Restriction endonucleases. Restriction enzymes were obtained from Bethesda ResearchLaboratories,
Gaithersburg,Md. Twotofive units ofenzyme perjig ofDNA wasused with the reaction conditions pro-vided by the supplier. The extent of reaction was
monitored with either lambda DNA or adenovirus
type2DNA.
RESULTS
Quantitation of nonintegrated proviral
on November 10, 2019 by guest
http://jvi.asm.org/
Fv-1 HOST RESTRICTION OF F-MuLV 47 DNA from acutely infectedcells. Acute
in-fection by retroviruses initially results in the
formation of free nonintegrated proviral DNA
andsubsequently in theintegration of thisDNA
into the host genome. Previous studies have
demonstrated two predominant forms of free
DNA in the infected cells: a double-stranded
linear DNA (forn III), which gives rise to a
covalently closed circular supercoiled DNA
(form I), which in turn is presumed to be the
precursor of the integrated provirus (21).
Be-cause relatively few copies of viral DNA are
synthesized percell,ourinitial studieswere
di-rectedatoptimizingtherecoveryof thesemajor
nonintegrated DNA forms.Avery poorrecovery
of closed circular proviral DNA was obtained
when the Hirt extractionprocedure (4)was
ap-pliedto 5 x 107permissivelyinfectedcells
grow-ing in roller bottles. Our suspicions were
that
this lowyield of circular DNA could be dueto
extraction artifacts. Accordingly, we modified
both the extractionprocedure andtheinfected
cell number. The modification of the Hirt
pro-cedure involveslysingandtreating the infected
cells at 65°C before salt precipitation of
chro-mosomal DNA.Using this method,wewereable
toisolatesignificantamountsof both form I and
form Ill DNA from as few as 1 x 106 cells.
However, scaling up the procedure to process
larger numbers of cells grown inroller bottles
still resulted inapoorrecoveryofform I DNA.
We attribute these results eithertomechanical
entrapment ornuclease degradation of
uninte-gratedproviral DNAdue to agreatercell
con-centrationduring extraction.
With this new meihod, and using only
be-tween1 x106and5 x106cells,weexamined the
proviral DNAaccumulated inseveral cell lines
considered either permissive or restrictive in
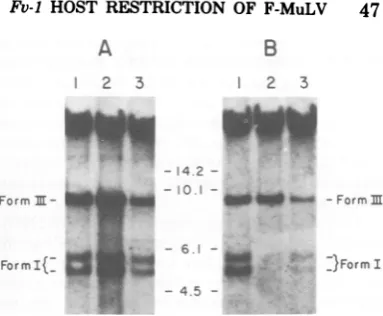
termsof theirFv-1genotype.Ourresults, shown
in Fig. 1A, indicated that distinctly different
amounts of nonintegrated proviral DNA are formedindifferent celltypes evenif the cellsare
consideredequallypermissiveatthe Fv-1 gene.
UsinganNB-tropicF-MuLV,avirus whose host
rangeis notlimited bythe Fv-1 gene, we
com-pared the proviral DNA accumulated in NIH
(Fv-1f'/),
BALB/c (FV-lb/b),
and DBA/2(Fv-IJn/f)
cells.Extrachromosomal DNA obtained bythe modified Hirt extractionwasseparated by
electrophoresis inagarose, transferred to
nitro-cellulose (22), and hybridized to a
[32P]DNA
probeprepared by nicktranslationofcloned
F-MuLVDNA(13).It is evident thatthe recovery
ofboth form I and form III from each of the
three cell lines was different. Recovery was
greatest fromNIH (lane 2), least from DBA/2
(lane 3), and intermediate from BALB/c (lane
1) cells. A study comparing N-tropic virus in
A
2 3
123
w:p1q
Form
II-FormI(- Cs
B
2 3
-14 2
--iO __ m mFo
- 6.1 -- 4.5
[image:3.496.247.439.52.210.2]-U:.
FIG. 1. Autoradiograms offilterblothybridization analysis ofHirt DNAextractsfromvirally infected cells. Subconfluentmousefibroblastcelllines(5.0x
106cells)wereinfected with either B-tropic F-MuLV orNB-tropicF-MuLV(MOI of1.0)inthepresenceof
Polybrene (5pg/ml) for 16 h. After infection, cells
wereextracted bythemodifiedHirtprocedure (see
Materials andMethods), and the Hirt supernatant DNAwasloadedonto0.8%agarosegels. Electropho-resiswasfor 20 hat50 V. Eachlanecontainsthe DNAobtainedfromthe entireextractof5x10'cells.
Afterelectrophoresis, the DNAwastransferredtoa nitrocellulose filterandhybridizedto a nick-trans-lated [32P]DNA (specific activity, 0.5 to 1.0 X 108
cpm/pg)probecontainingF-MuL Vsequences. Auto-radiography wasperformedwith X-Omatfilm and
intensifierscreensat-70°C. (A) NB-tropicF-MuLV
infection ofBALB/ccells(lane 1),NIHcells(lane 2),
and DBA/2cells(lane3). (B)B-tropic
F-MuLVinfec-tionofBALB/ccells(lane1),NIHcells(lane 2),and
DBA/2cells(lane 3). The bandsof hybridizedDNA
areindicatedatthe sidesofeachautoradiogramand
representformIcircular andform III linear viral
DNA. The numbers between theautoradiograms
rep-resentthe locationsand sizes (inkilobasepairs) of BamHI-digested adenovirus type2 DNAfragments migratingin the samegel. The broad bands atthe
topofthegelsrepresentresidual hostcellDNAthat wasnotcompletelyeliminatedbytheHirtprocedure.
NIH versus DBA/2 cells gave similar results (datanotshown). The basisfor thesedifferences
maybeatthe levelofsynthesis,degradation,or extractability of proviralDNA from the individ-ualcell lines. The relative platingefficiency of each cell line also differsand must be controlled to ensure identical cell numbers, both at the time of infection and at the time of harvest.
These possibilities have not all been explored. We concludedthatthe differencesintotal
recov-ery between cell lineswould complicate an at-tempttocompare the effectof the Fv-1 alleles in these different cell lines and that this fact mustbeconsidered inanysuchanalysis.
Inadditiontovisualanalysis of the
autoradi-ogram,wecomparedthe amounts of form I and form III DNA by assaying the labeled probe })FormI VOL. 40,1981
on November 10, 2019 by guest
http://jvi.asm.org/
48 CHINSKY AND SOEIRO
bound tothe nitrocellulose filter. Inourhands,
the totalamountofradioactivity that could be
directly ascribed to the DNA bands was very
low,usually of the order of50 to 200cpmabove
abackgroundof approximately50cpmfoundin
theinterband regions. We consideredthat these
small differences of radioactivity wouldnot
per-mitus tomakeanaccuratecomparison between
different cells.
Toquantitate thepresenceofproviral DNA,
microdensitometrytracing of the autoradiogram
presentedan alternative toscintillation
count-ing. By analyzing the weight of theareaunder
the curveof the microdensitometry tracing,we
measured the total amount of annealed
radio-active DNA and minimized the background.
Furthermore,weelectedtocomparetheratioof
formsIand III DNA inthosecells which differed
inaccumulationor recovery or both ofproviral
DNA. Inthisway, ananalysis ofratios could be
made with amethodthat wouldnormalize
re-sults, regardless of the total proviral DNA
re-covered. In Fig. 1A, for example, the ratios of
form IIItoform I DNA for the threeNBtropic
F-MuLVinfectedcellsareverysimilar; forNIH
it is1.34, for DBA/2 it is1.39,and for BALB/c
it is 2.12. We feel that, within the limits of
resolution of the technique, these numbersare
not significantly different. Comparison of the
ratiosof the proviral forms thereforepermits the
determination of the effect of different Fv-1
al-leles onthesynthesis oraccumulation orboth
of these proviral DNAforms by asingle virus
type.
B-tropic F-MuLV infection of Fv-_jln
cells. Thedegreeofrestriction of growth of
B-tropic F-MuLV in NIH Swiss fibroblastcelllines
can vary up to 100-fold depending upon the
stock of B-tropic virus examined. We cloned
stocks of B-tropic F-MuLV that are restricted
either 10-2or10-4whenanalyzed by the direct
XCplaqueassay onNIHversusBALB/ccells.
The synthesis of unintegrated viralDNA after
infection of various cell lines by the
10-2
re-stricted B-tropic F-MuLV
(B-2)
was examinedby the Southern blot hybridization technique.
The resultsareshown in Fig. 1B. In both
per-missiveBALB/ccells(Fv-1b/b)and
nonpermis-siveNIH cells
(Fv-1'1)
(lanes 1 and2,respec-tively), nearly equivalentamounts of linear
pro-viral DNA (form III) werefound. However,in
the nonpermissive cell, essentially no form I
DNAcould bedetected(lane 2). By comparison
with DNA size standards, weestimatethe size of thelinearproviralDNAinbothcelltypes to be about 8.8 to 9.1 kilobase pairs (kb). Using
CsCl-ethidium bromide density gradients, we
demonstrated for the permissive cell that the
bands migrating between the 6.1- and 4.5-kb
markersrepresent the two closed circular
pro-viralDNAspecies(formI) known toaccumulate
inretrovirus-infected cells (20, 27). By analogy
withpublished data, the slower migrating of the
twospeciesof formIshould containtwocopies
of the long terminal repeat sequence (LTR),
whereas the faster band should contain onlya
single copy (20, 27). This clone ofB-tropic
F-MuLV thusappearednottoberestricted in its
production of form III DNA, but accumulated
markedly diminished amounts of both form I
DNA species. In contrast, in another Fv-1
1'-type cell (DBA/2), the accumulation of both
form III and form Iwasdiminiishedrelative to
the NIH cell (lanes 3 and 1, respectively) as
would bepredictedfromourearlierresults(Fig.
1A). InthecaseofDBA/2cells,however, both
species of formIDNAareevident(Fig. 1B,lane
3).
The ratio of linearDNA tothe combinedtwo
species of closed circular DNAwasobtainedby
means of comparison of the areas under the
curves describedbymicrodensitometry. In the
case ofB virus infection ofBALB/c cells, the
formn
III/form Iratio equaled 0.76, whereas Bvirusinfection of DBA/2 cells resulted inaratio
of 1.38. Noratio could be determined for NIH
cells because no measurable form I was
accu-mulated (Fig. 1B, lane2). By direct XCassay,
this clone ofB-tropic virus wasrestricted
100-to 500-fold on NIH cells, but only 50-fold on
DBA/2 cells. In agreement with published
re-sultsfrom otherlaboratories,Fv-1 host
restric-tion of this B-tropic F-MuLV in NIH cells is
associated withadrastic inhibitionof the
accu-mulationofformI (9, 26). However, in the less
restrictive DBA/2 cells,clear-cut accumulation
ofclosed circular DNAwasfound,andat aratio
tolinear formsonlyslightlydifferent from that
found in thepermissiveinfection. Forreference,
on this autoradiogram, a nonrestricted
NB-tropic virus infection ofDBA/2cells resulted in
aratio of1.67 (Fig. 1A). These resultssuggest
thatthe decreased accumulation of formIDNA
found inB-tropicinfection of restrictiveDBA/2
cells (ratio, 1.38)relativetothepermissive
infec-tionofBALB/c cells(ratio, 0.76)maynot
com-pletelyexplainthe50-folddecrease in infectious
centersobservedin theXCassay.
ThesynthesisofproviralDNA in NIH Swiss
cells was further examined with a clone of
B-tropicF-MuLVthat exhibits10,000-fold
restric-tion (B-4) whenexamined bydirect XC assay.
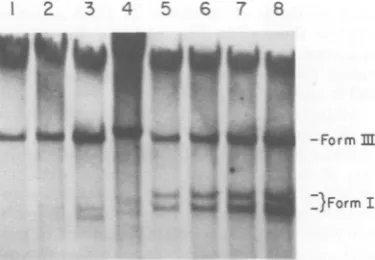
Theresultsareshown inFig.2.Linearproviral
DNAspecies(form III)weredrasticallyreduced
in thenonpermissive NIHcells(Fig. 2A,lane3;
Fig.2B, lane7)comparedto
permissive
BALB/ccells (Fig. 2A, lane 2; Fig. 2B, lane 6) in two
different experiments withtwo different
input
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 40, 1981
A
12 3
B
[image:5.496.43.235.63.197.2]1 2 3 4 5 6 7
FIG. 2. Autoradiograms offilter blot hybridization analysis of cells infected with highly restricted B-tropic F-MuLV. Subconfluent mouse fibroblast cell lines (1.5 x 106 to 2x 106 cells) were infected with either B- orN-tropic F-MuLVfor 24 h. Hirt
super-natantDNA was subjected to electrophoresis in0.8% agarosefor (A) 11.5 h at 50 V or (B) 16 h at 50 V. DNA, transferred to nitrocellulose, was hybridized to
32P-labeled MuLV DNA. (A) Lane 1, N-tropic
F-MuLVinfectionof NIH cells (MOI of 0.4); lane2,
B-tropic F-MuLV infection of BALB/c cells (MOI of 0.4);lane 3, B-tropic F-MuL V infection of NIH cells
(MOI of0.4). (B) Lanes 1 to 5, N-tropic F-MuLV infection of: NIH cells (MOI of 1.0), DBA/2 cells
(MOIof 1.0), BALB/c cells (MOI of 1.0), BALB/c cells(MOIof 2.0), BALB/c cells (MOI of 5.0), respec-tively; lane 6, B-tropic F-MuL V infection ofBALB/c cells(MOIof 1.0); lane 7, B-tropic F-MuL V infection of NIH cells(MOI of 1.0).
multiplicities. Othersetsof permissiveand
non-permissive infectionsarepresentedinthis figure
forcomparisonoftheamountsof formIII DNA
synthesized; thesewill be discussedlater. This
result, demonstrating theabsenceofformIand
reducedamountsofformIII,isclearly different
fromthat whichwasobserved for the
100-fold-restricted B virus(B-2),whichexhibited little to
nodecreaseinlinearDNA(formIII).
Independ-entclonesofB-tropic viruscanthereforeexhibit
different types of restriction of their proviral
DNAinNIHSwiss cells. Thishighly restricted
viral clone (B-4) did not appear to be able to
accumulate linear proviral DNA to the same
extent as the less restricted clone (B-2). These
datamayexplain the different degreesof
restric-tionexhibitedbythese twoB-tropic clones.
The effect of the multiplicity of infection
(MOI)onthe synthesis ofproviral DNA in
Fv-1permissivecellscomparedwith nonpermissive
cells was examined. B-tropic F-MuLV which
exhibits 102restriction in NIH cells (B-2)was
used to infectNIHand BALB/c cellsatMOIs
rangingfrom 0.25 to 1.0. Theresultsareshown
inFig.3. Asexpected,thetotalamountof
pro-viral DNA increased relative to the MOI. At each MOI, the amount of form III DNA
ob-Fv-1 HOST RESTRICTION OF F-MuLV 49
served in thenonpermissive cellswasequivalent
to that observed inthe permissive cells.
How-ever,regardless of the MOI, theamountof form
I DNA in the nonpermissivecells was greatly
reduced.Itisunclear whether the lack of
detec-tion of form I in thenonpermissive hostat an
MOI of0.25(lane 1)is duesimplytoinsufficient
numbers ofinfected cells usedinthisanalysisor
whether this form is completely absent atthis
MOI.
Two rapidly migrating bands of DNA are clearlyobserved at thehigherMOI in the
non-permissivecells(lanes3and4).Thesebandsare
seen to migrate more rapidly than the fonn I
DNAs found in the permissive BALB/c cells
(lanes5 to8). Because these bands in the
non-permissivecellsarepresentatlowconcentration,
wehave thus far beenunabletodemonstrate,as
wehave inpermissive infections, thattheyare,
in fact, form I DNA either by CsCl-ethidium
bromide density gradient orby restriction
en-zyme criteria (datanot shown). In the
nonper-missivecell, these presumed form I bands were
present at aratio of1:10tothe amountofform III present. If theproviralDNAsynthesizedin the nonpermissive cell had a lower molecular
1 2 3 4 5 6 7 8
-Formm
-}Form I
FIG. 3. Autoradiogram of filterblothybridization analysis ofeffect ofMOIbyB-tropicF-MuLV. Sub-confluent NIH Swiss (lanes1 to4) andBALB/c cells
(lanes5 to8)wereinfectedwithB-tropicF-MuL Vat
variousMOIsfor16h andsubjectedtomodifiedHirt extraction asdescribed in thelegendtoFig. 1. Hirt supernatantDNAwassubjectedtoelectrophoresisin 0.8% agaroseat 60Vfor16h.Afterelectrophoresis, DNAwastransferred bythe Southern blotprocedure
to nitrocellulose and hybridized to 32P-labeled F-MuLV DNA.Autoradiographywasperformedasin Fig. 1. Lanes 1 to 4: B-tropic F-MuLVinfection of NIH Swiss cellsatMOIsof0.25(lane 1),0.50(lane 2),
1.0(lane 3), and2.0(lane 4). Lanes5 to 8:B-tropic F-MuLV infection ofBALBIc cells at MOIs of0.25
(lane 5), 0.50 (lane6), 1.0(lane 7), and2.0(lane 8).
The hybridized bands of viral DNA representing
formIIIlinear andformIcircularDNA are
indi-catedonthe sidesof the autoradiogram. knothi[L
and ud
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.248.436.348.478.2]50 CHINSKY AND SOEIRO
weight than thatin thepermissivecell,
circular-ization would result in a form I DNA which
would migratemorerapidly than itspermissive
equivalent. Asmall difference in the size of the
linear proviral DNA (form III) synthesized in
thenonpermissive cell wouldnotbe detected by
directvisual examination ofourgels because of
thelimitedresolvingpowerof 0.8%agarosegels
for DNA species thataregreaterthan8.0kb in
size. We therefore digested the proviral DNA
obtained fromnonpermissiveversuspermissive
cells with the restriction endonuclease EcoRI.
This enzyme cuts the viral genome only once,
producingtwobands of5.7and3.4 kb
(unpub-lished data and reference 13). Ifadifference in
the sizeof the linear DNA from the
nonpermis-sive cellswerepresent, itmight be detectedas
an altered migration ofone ofthe viral bands
resulting from EcoRI digestion. The results of
thisanalysisareshown inFig.4. Thepattern of
digestion obtained from proviralDNAextracts
from nonpermissive cells was similar to that
obtained fromproviralDNAextracted from the
permissive cells. Preliminary studies using
HindIII,anenzymewhichappearsto cuttwice,
confirmed these results. Therefore, it does not
appearthat the basis for the restriction in the
nonpermissive cells isamajordeletionor
incom-plete synthesis of the form III DNA
accumu-lated in these cells. The basis for the altered
migration pattem of the doublet DNA
(pre-sumed circular species) is not yet understood
and isunderinvestigation.
Additionally, we tested whether extensive
methylation of form III in different host cells
could be associated withdifferential
accumula-tion of circular DNA. The results (data not
shown) ofHpall digestionoflinear DNA
accu-mulated in eitherapermissiveor a
nonpermis-sive host reveal noresistant fragments of
pro-viral material. Since resistance tothis enzyme
correlates with methylation of the internal
cy-tosine in the HpaII recognition sequence (16),
these resultssuggestthat extensivemethylation
of proviral DNA doesnot occur in either cell
type,butthey donotexclude limitedsite-specific
methylbasemodification.
Insummary, independent clones ofB-tropic
virus exhibited wide variation in the restriction
byNIHSwisscells whenexaminedbythe direct
XCplaque assay. Thedegreeofrestriction
ap-pears to correlate not only with the ability to
accumulate normal amounts of proviral DNA butalso with thetypes of DNA made.B-tropic F-MuLV thatexhibitsahigh degree of
restric-tion in NIHcells
(B-4)
appearedtoaccumulateseverely reducedlevels oflinear,andessentially
no circular, forms of proviral DNA. Viruses
which are lessrestricted (B-2)were ableto
ac-64
-9.1
-57
[image:6.496.307.423.67.265.2]_Irni * -.34
FIG. 4. EcoRI restriction enzyme analysis offormn IIIDNA.Subconfluent ceU lines (5 x 106 cells) were
infectedwith B-tropic F-MuLV(MOI of1.0) in the presenceofPolybrene (5pg/mi)for 23 h and subjected
tomodifiedHirt extraction. Viral DNA samples were treated withendonuclease EcoRI (5U/pgofDNA) at 38°C for 3 h and then subjected to electrophoresis on 0.8% agarose to 60 V for 17 h. After electrophoresis, DNAwas transferred to nitrocellulose and hybrid-izedto32P-labeledF-MuLV DNA.Autoradiography wasperformed as in Fig. 1. Undigested DNA from B-tropic F-MuLV infection ofBALB/c cells (lane 1) and NIH cells (lane 2) was run in parallel with
digestedDNAfromBALB/c cells (lane 3) and NIH cells(lane 4). The numbers at the side of the autora-diogram represent the molecular size (in kilobase pairs) estimated from restriction fragmentsof ade-novirustype2DNA and lambda DNA that were run in parallel. In lane 4, the band migrating at 9.0 kb represents endogenous cellular sequences found in genomic DNAofuninfectedaswell as infected cells. Inthis gel, these sequences resultfrom the contami-nating chromosomal DNA that is found in all Hirt
extracts. In lane 3, the bands migrating to 9.0 kb represent these same cellular sequences as well as form III (DNA) derivedfrom formIafter a single cut
byendonuclease EcoRI.
cumulate normal amounts oflinearDNA, but
accumulatedmarkedlyreducedlevels of
abnor-mally migrating presumptive circular DNA.
Theseresultssuggestthat,in NIHcells,
restric-tion ofB-tropic virusreplicationmay occur at
either the accumulation of linear or circular
DNA.Whetheronlyone orbothof theseeffects
may be correlated with the Fv-1phenotypeisat
thistime notresolved.
Furthermore, in DBA/2 cells, B-tropic
F-MuLVwasrestrictedto alesserdegreethanthat
observedwith NIHcells. However, the ratio of
the accumulation of circular forms relative to
J.VIROL.
j
::
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 40, 1981
linearappearsequivalenttothatobserved with
apermissive infection (NB-tropic virus) in those
same cells (Fig. 1). Therefore, the degree of
biological restriction observed in tissue culture assays with DBA/2 cells didnotcorrelate well withadecreased accumulation of form I proviral DNA.
N-tropic F-MuLV infection of BALB/c
cells: analysis of nonintegrated proviral
DNA. Several clones ofour stock N-tropic F-MuLVweretested and allconsistently exhibited between 100- and 500-fold restrictionon
nonper-missive BALB/c cells when examined by the direct XCplaqueassay.Tocompare thesite(s) of Fv-1 restriction, we examined the proviral DNAspecies of N-tropic F-MuLV accumulated inNIHversusBALB/c cells. The results repre-sentinfectionatarangeof MOI from 0.25to1.0
at16to 17 hpostinfection (Fig. 5). Visual eval-uation of theautoradiogram of the blot hybrid-ization suggests thatateach MOI, theamount
of formIIIspecies accumulatedwasslightly less in thenonpermissive, when compared with the permissive, infection.Inotherexperiments, the
amounts of form III were identical. However,
theamountofform I DNA in theBALB/c cell
1 2 3 4 5 6 7 8 9
[image:7.496.246.439.323.549.2]Y
FIG. 5. Autoradiogram of filterblothybridization analysis of effect ofMOIby N-tropicF-MuLV.
Sub-confluentBALBIc(lanesIto4)andNIH Swisscells
(lanes5to8)wereinfectedwithN-tropicF-MuLVat
variousMOIsfor16 h andsubjectedtomodifiedHirt extractionasdescribed in thelegend for Fig.1.Hirt
supernatant DNAwassubjectedtoelectrophoresisin 0.8%agaroseat60 Vfor16h.Afterelectrophoresis,
DNAwastransferred bySouthern blotprocedureto
nitrocelluloseandhybridizedto32P-labeled F-MuLV DNA.AutoradiographywasperformedasinFig. 1. Lanes1to4:N-tropicF-MuLVinfection of BALB/c cellsatMOIsof0.25(lane1),0.50(lane2), 0.75(lane 3),and 1.0(ane4). Lanes5to8: N-tropicF-MuLV
infection ofNIHSwisscellsatMOIsof0.25(ane5),
0.50(lane 6), 0.75 (lane 7), and1.0(lane 8). Lane9
representsDNAobtainedfrom B-tropic F-MuLV
in-fection ofNIHSwiss cellsatanMOIof5.0and is
shownfor comparison oftheelectrophoretic
migra-tionof formI(Fig. 3).
Fv-1 HOST RESTRICTION OF F-MuLV 51
(lanes 1 to4) was reduced ateach MOI, when
compared with the permissive cell (lanes 5to8). Microdensitometry tracingsweretaken foreach
lane, and theareasunderthepeakforeach DNA
form were weighed for quantitation (Table 1). Two points are evident: (i) at each MOI the amountof form III inpermissiveversus
nonper-missive cellswasnearlyequivalent, and (ii) the amountsof form Ifound in the nonpermisive
cellwerereduced. The ratiosof form III to form Iindicate that onlyatwo-tothreefolddecrease
in the amount of circularspecies occurredinthe restrictive infection. Aseries of similar experi-ments, in which the proviralDNA formswere examinedatbetween 6and20 h afterinfection, all confirmed these findings. Within 6 h after
infection, we documented the accumulation of
linear DNA inexcessofcircularDNA,thatis,a high ratio of form III toform I. With time this
ratioapproachedthatseenat12 to 16 h(Table 1). In the nonpermissive infection, a parallel time-dependent changeinaccumulation of DNA
species occurred.But thesamerelative decrease
in accumulation of circular DNAseen at 16 h
TABLE 1. EffectofMOI onproviralDNA in
N-tropicF-MuL Vinfection
Hostcell MOI Form 11 Form1
Ratio
(A) NIHSwiss 0.25 0.016 0.015 1.07 0.50 0.031 0.030 1.03 0.75 0.040 0.050 0.80
1.00 0.057 0.0655 0.87 (B)BALB/c 0.25 0.0285 0.011 2.59
0.50 0.045 0.019 2.37 0.75 0.065 0.027 2.41 1.00 0.092 0.030 3.07
NIHSwiss 0.25 0.053 0.055 0.96 0.50 0.076 0.066 1.15 (C) BALB/c 0.50 0.049 0.021 2.30
0.75 0.059 0.022 2.68 1.00 0.061 0.023 2.65 NIHSwiss 0.50 0.034 0.024 1.42
0.75 0.051 0.032 1.59 1.00 0.074 0.045 1.64
(D)NIHSwiss 1.00 0.123 0.062 1.98 BALB/c 1.00 0.092 0.017 5.41 2.0 0.115 0.030 3.83
5.0 0.199 0.048 4.15 aNumbers indicate weightingramsofpeakareas
cutfrom themicrodensitometrytracingsof autoradi-ograms.All the measurements in each letteredgroup
weremadeusingasinglewedge settingonthe micro-densitometer. (A) and(B)representdifferent
autora-diographic exposurestothesamehybridized Southern
blot. (A) and (B):5 x 10'cells wereinfectedin the
presence ofPolybrene (5
,ug/ml)
for16h. Theauto-radiogramcanbeseeninFig.5.(C):2x 106cellswere
infected in the presence ofPolybrene (5 Lg/ml)for16
h.(D):1.5x 106 cellswereinfected in the presence of
Polybrene (5 Lg/ml)for24h. Theautoradiogramcan
beseeninFig.2B.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.496.43.233.349.482.2]was observedateach timepoint in the nonper-missive infection. That is, although both the absoluteamountand thetypeofproviral DNA species accumulated in cells varied with time, therewas aconsistentfinding (indicated in the ratio ofform IIItoformI) ofadecreased
accu-mulation of only two- to fourfold of circular DNA inBALB/c cells infected with N-tropic F-MuLV relative to that seen in the permissive infection. These data are consistent with the decreased accumulation of forn I reported by other investigators (9, 26). However, they are
inconsistent with the degree of reduction of form Inotedby Jolicoeur and Rassart (9). Yangetal. (26) suggested a multiplicity-dependent effect ontheformation of form I DNA in nonpermis-sivecells.Our datasuggestthat thedecrease in form I DNA accumulation is independent of multiplicity, since the ratio of form IIItoform Iwasconstantoverawiderangeof viralinput.
The phenomenon of the Fv-1generestrictive
effecthas, however, been shown inmany labo-ratoriestofollow two-hitkinetics. That is, titra-tion of either infectious centers or ofprogeny
virus production in nonpermissive cells de-creases geometrically rather than linearly with virusdilution. Incontrast tothepermissive cell infections, these findings have been interpreted
to indicate that a nonpermissive cell must be infected by at least two infectious viruses to
establish an infectious center. By contrast, the permissivecellrequiresonlyasingle "hit" (15). However, single-hit restriction curves, appar-ently due to the Fv-1 host gene, have been
reported inmanylaboratories (reviewed by
Jol-icoeur,6).
OurN-tropic F-MuLVappearstoberestricted by single-hit kineticsasmeasuredby the direct XC assay. These kinetics may also be
demon-stratedbyprogenyyield. NIH andBALB/c cells
wereinfectedatanMOI ofeither 2.0or0.2. The
progenyyield from suchpermissivelyor
nonper-missively infected cells wasmeasured on NIH
cells at24 through 96 h postinfection. The
re-sults, shown in Table 2, were (i) at 24 h in
permissive cellstherewasthe expected 10-fold
difference in yield, comparing an MOI of0.2 with oneof2.0; (ii) atboth 24 and 48 h
postin-fection, there was a 100-fold decrease in yield,
comparingpermissivewithnonpermissive
infec-tion, and this result occurred at eitherMOI; (iii)
at72 and96h, therewasonlya10-fold restric-tion inyield,aresultpresumably due to
cell-to-cellspread,aphenomenon previously found by
us (Soeiro, unpublished data) and others (7).
Similar experimentsstudying infectiouscenters
producedby N-tropic F-MuLV infection of NIH versus BALB/c cells have also indicated such single-hit kinetics. Our results, interms of the ratio of form IIItoform I inanFv-1 restrictive infection, musttherefore be interpreted in the light ofapparentsingle-hit kinetics.
Toexamineproviral DNA formed undermore extremeconditions ofmultiplicity,wecompared theN-tropic F-MuLV infection of BALB/c ver-susNIHcellsatanMOI of 5.0. At thisMOI,at
least 96% of the cellswere infected bytwo or moreinfectiousparticles. It would be expected, accordingtothe two-hitmodel, thatatthisMOI restriction would be completely overcome and the patterns ofproviralDNAsynthesismightbe
identical. For comparison, we also infected a muchlarger numberofBALB/ccellsatanMOI of0.5,sothat theamountsoftotallinear DNA produced by N-tropic F-MuLV would be
ap-proximately equaltothat foundby infection of fewercellsatahigherMOI. Theresultsof this experiment depictedasmicrodensitometry
trac-ings from the autoradiogram are shown inFig.
6. Figure 6A represents the twocircular forms (form I) and the linear formIII ofpermissively infectedNIHcells.Figure6Brepresents similar tracings takenfromBALB/ccellsinfected atan MOI of 5.0. Figure 6C is the tracing obtained
fromagreaternumber ofBALB/ccellsinfected atan MOIof0.5. Theratio ofweights of form TABLE 2. Effect ofMOIonprogenyyield from permissive versus restrictive infection'
Progenyyield at hpostinfectionof:
MOI 24 48 72 96
N B N B N B N B
2.0 4 x 104 3x 102 1.2x 107 1.2x 105 0.9 x 106 1.3 x 105 1.4x 106 2 x 105
0.2 3x 103 NDb 8 x106 4 x 104 9 X 105 5 x 104 3 x 106 3 x 105 aSubconfluent mouse fibroblast cell
lines
(1.0 x106
cells) were infected with N-tropic F-MuLV in the presenceof Polybrene (5,ug/ml)at an MOI of 2.0 or 0.2. After a 3.5-h adsorption period,cellswerewashed and given fresh medium. At thetimesindicated,the medium was replaced, and the titers of the progeny virus in the removed mediaweredetermined on permissive (NIH) cells by direct XC plaque assay. The results obtained from media removed from infected NIH Swiss cells (N) or BALB/ccells (B) are given as the number of macroscopic PFU permilliliter.bND,Notdetected.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
Fv-1 HOST RESTRICTION OF F-MuLV 53
A
B
C
DNA Forms: I m I m I m
Area Under: Curve (g)
Ratio m/I:
.147 .133
0.90
.064 .147 .051 .142
2.30 2.78
FIG. 6. Microdensitometrytracings offilterblothybridizationanalysesof infection by N-tropicF-MuLV.
SubconfluentceU lines wereinfectedfor24h in thepresenceof Polybrene (5
lg/ml)
andsubjected tomodifiedHirtextraction(seeMaterialsandMethods).Afterelectrophoresisthrough 0.8%agarosefor16h at 50V,the DNA wastransferred bythe Southern blotproceduretonitrocellulose andhybridized to 32P-labeledF-MuLV DNA.Autoradiographywasperformedasdescribed in thelegendtoFig. 1,and theexposed filmwasanalyzed
lanebylanewithadouble-beamrecordingmicrodensitometer.After carefulconsiderationof background,the peak areas were cut out and weighed; thus, the areas under the curves are given in gram units. (A) N-tropic
F-MuLVinfectionof 1.0 x106NIHSwiss cells,MOI of5.0. (B)N-tropic F-MuLVinfection of 1.0 x106BALBE
c cells, MOIof5.0. (C) N-tropic F-MuLVinfection of4.0 x 106 BALBIc cells, MOI of 0.5. Thepeaks representingformIDNA andformIIIDNA are indicated.
III toform I is included in the figure.We can
conclude from comparing both the totalamount
of linear DNA formed and the ratio of linearto
circular formsthat,even at a veryhighMOI,at
which restriction is expected to be overcome,
there is a two- to threefold reduction in the
accumulation ofform I in nonpermissive cells.
Furthermore,even at anMOI of0.5,by
compar-isonwith5.0,the ratio of lineartocircularforms
innonpermissive cells remains thesame.
Theseresultsconfirm that,in anonpermissive
infection, accumulation offormIis reduced. But
they also show that thedegree of reduction is
much less than that expected by the 100-fold
restriction ofviral progenyyield. Furthermore,
atMOIs of5.0and0.5,the ratiosof form III to
form I are essentiallyidentical, a result
incon-sistent with a two-hit restriction phenomenon.
We conclude that decreased accumulation of
formIdoesoccurin restrictive infectionwith
N-tropic F-MuLV but that thedecrease is
incon-sistent with the degree ofrestriction observed in
tissue cultureassays. In addition,with our virus
and cell combination, the ratio ofform III to
formI inBALB/c cells did not reflect a two-hit
effectonthe decrease ofcircularDNA.
DISCUSSION
Fv-1-mediated host restriction of
B-tropic
MuLV affects the accumulation of
noninte-gratedproviralDNAinFv-1 'i" cells. The results
previouslyreported byotherinvestigators (9, 26)
have shownthat, in NIHcells,there isablock
in theaccumulationof forn III DNA(and
there-fore no
form
I is found) after infection withWN1802B. Other Fv-1'1'cells(DBA/2)infected
with thissamevirusproduceamountsof linear
DNA equivalent to that forned in permissive cells, but levels offorn I are reduced. It has been suggested that the differences observed
between these cell types could be due to the
presence of a second gene in NIH cells which
modifies the Fv-1 effect (9,26).
N-tropic
MuLV(WN1802N) infection ofFv-1b/b cells resulted
also in theaccumulationofnormalamountsof
linearbut decreasedamountsofcircular DNA.
Since previous resultsfromthislab and others
(8, 23) haveindicated that Fv-1 restriction
oc-curs at or before proviral DNA integration, it
wasconcludedthat theblockinaccumulation of
circular proviralDNA is the primary effect of the Fv-1 gene.
VOL. 40, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.496.69.399.56.294.2]54 CHINSKY AND SOEIRO
Wehave demonstrated that individual clones
of B-tropic F-MuLV can exhibit different
de-grees of restriction byNIH cells. Cloned virus
stock exhibitingalow level of restriction (10-2)
byXCassay wereshowntoaccumulatenormal levels of form III, but reduced levels of form I,
DNA. However, a second clone, showing high
(10-4)restriction,wasunabletoaccumulateeven
normallevels of formIII DNAinthesecells.
ThesedatasuggestthatNIHcellsareableto
restrict B-tropic virus atseveral points in the
earlyproviralDNAphase of replication. These
effectscould be dueto asingle pleiotropicFv-l1
allele capable of interacting at different steps
during proviral DNA accumulation.
Alterna-tively,NIHcellsmay carry asecondgenewhich
modifies the Fv-l" allele. Our studies withtwo
clones ofonevirus thatappearblockedat
differ-ent steps leadingto integration do notresolve
thisquestion.They do, however, show that these
two clones are recognized differentially by the
samecell, suggesting thepossibilityofmorethan
asingle recognition feature in host cell
restric-tion. There might be two sites on the virus
recognized by a single Fv-1 gene product or,
altematively, two gene products. This
conclu-sion is underscored by the fact that both
B-tropic clonesarerestricted tothe same degree
(50-fold) in DBA/2 cells
(Fv-1'1).
ThissuggeststhatDBA/2 cellsmayonly be abletorecognize
asingle feature shared by both viruses. These
patternsofproviralDNAaccumulationmaybe duetothepresenceofasecondrestrictinggene
product(s) inNIH cells which isnotpresent in
DBA/2 cells or to an increased efficiency of a
single Fv-l"geneproductinthe NIHcells.
Our studies of the restriction ofN-tropic virus
inBALB/ccells have revealed adecreased
ac-cumulation of form I DNA in agreement with
studies from other laboratories. However, we
observed that the reduction of form Iwas only
of theorder oftwo- tothreefold when compared
with theamountofproviralDNAobtained from
permissively infected cells. Our results are
clearly different from the estimated 25- to
50-fold reduction observed by Jolicoeur and
co-workers (9) and do not show amultiplicity
de-pendence as observed by Yang et al. (26). A
possible reasonfor the differences observed,in
addition to modifications of technical
proce-dures, may reside in the virus stocks used in
thesestudies.The titrationpatternobservedby Yang and co-workers isclearlytwohit in
non-permissive cells, whereas our results
demon-stratedaone-hitrestriction curve, thatis,alack
ofmultiplicity effects on virus yield or on the
proviralDNApattern.
SeveralfeaturesofFv-1restriction have been
described, including "hitness," "refractoriness,"
J. VIROL.
"expression,"and"prozone" effects(15). These
arewell discussed by Jolicoeur (6), whose
con-clusion is that both refractoriness and two-hit
kineticsappear tobepartof the Fv-1geneeffect.
Our findings are that decreases in the
accu-mulationof form I (two-tothreefold)are
incon-sistentwith the degree of restriction (100-fold)
and thatthe decreaseinform I accumulationis
notmultiplicity dependent (Fig. 6).These data
suggestthat forourcells and virus the decrease
informImaybe associated withrefractoriness,
thatis, anon-multiplicity-dependent
Fv-1-me-diatedrestriction effect. (The word
"refractori-ness" is usedtoimplyadecreased efficiency of
formation of infectiouscenters in nonpermissive
hostcells.)
The data of Yangetal., which showatwo-hit
effectonthe accumulation of form IDNA,
sug-gestthatadecrease incircular DNA is the site
ofFv-1 restriction. Our data, which didnotshow
such a multiplicity-dependent effect on form I
accumulation,suggestthat Fv-1 restrictionmay
notbe due simply to effects on circular DNA
accumulation. An alternative model for the
site(s) of Fv-1 gene effect is suggested by our
data.If theefficiency of integration varies geo-metricallywith theconcentration ofform I, then small decreases in accumulation of form I might
result in large differences in the establishment
of infectiouscenters.Therefore, in additiontoa
small decrease in form I accumulation, there
might bealarge effectontheintegrationprocess
itself. This second effect might be considered
secondary, but could result ina geometric
out-come ofalinear process. There is no evidence
against suchamodel thatwe are awareof.Also,
itmustbeconsidered that thebiological
signifi-canceof theaccumulatedproviral DNA species
found after the first few hours ofinfectionis yet
tobecompletely defined andmayrepresent an
easily measurable but dead-end feature of an
acuteinfection.
Lastly, we have attempted to assay
mecha-nisms whichmight resultindecreased
accumu-lation ofform I DNAintherestrictivecell.Our
data, which show no residual HpaII-resistant
proviral fragments,suggest thatextensive
meth-ylationof the DNA atcytosineresidues in either
a permissive or restrictive infection is not
in-volvedinthis effect. Also,no large deletionsin
the length of linear DNA could be found by
preliminary restriction analysis of the forn III
DNAaccumulatedinrestrictivecells.However,
these studies do notexcludethepossibilitythat
verysmall, especiallyterminal, fragments may
bemissing or base modified. The easily
recog-nizedalterationintheelectrophoretic migration
of putative form I in Fig. 3 and 5 is as yet
unexplained. It may represent a more highly
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 40, 1981
supercoiled structurethan isnormally
accumu-lated. Whether this structure is a result ofan interaction of proviral DNA with the Fv-1 gene
productmustbe determined. An understanding of this result may define mechanistically how the Fv-1 gene effect is mediated. Finally, the mechanism(s) for the decreased accumulation of form I hasnotbeen determined andmayresult from eitherchangesinsynthesisordegradation orboth of circlesfonned in therestricting cell.
ACKNOWLEDGMENTS
This workwassupported bygrantsMV-40from the Amer-ican CancerSociety, P3 OCA-13330 from the NationalCancer Institute, andtraininggrant5T32GM7288 fromthe National Institute of General Medical Sciences.
LITERATURE CMD
1. Decleve, A.,0.Niwa, E. Gelmann, and H. S.Kaplan.
1975.Replication kinetics of N- and B-tropic murine leukemia virusesonpermissiveand non-permissivecells
invitro.Virology 65:320-332.
2. Duran-Troise, G., R H. Bassin, A. Rein, and B. L.
Gerwin. 1977. Loss ofFv-1 restriction inBALB/3T3
cellsfollowing infection withasingle N-tropic murine leukemia virus particle. Cell10:479-488.
3. Hartley,J.W.,W. P.Rowe,and R. J.Huebner. 1970. Hostrangerestrictions of murine leukemiaviruses in mouseembryo cell cultures. J. Virol. 5:221-225.
4. Hirt,B. 1967. Selective extraction ofpolyomaDNAfrom infectedmousecellculture. J.Mol. Biol. 26:365-369.
5. Huang, A. S., P. Besmer,LChu,and D.Baltimore. 1973. Growth ofpseudotypes of vesicular stomatitis virus withN-tropic murine leukemia viruscoatsincells
resistanttoN-tropic viruses. J. Virol.12:659-662. 6. Jolicoeur,P.1979.TheFv-i geneof themouseand its
control ofmurine leukemia virusreplication. Curr. Top. Microbiol. Immunol. 86:67-112.
7.Jolicoeur, P.,and D. Baltimore. 1975. Effect ofthe Fv-1locusonthe titration ofmurine leukemiaviruses.J.
Virol.16:1593-1598.
8.Jolicoeur, P., andD.Baltimore. 1976. Effect of Fv-1
geneproductonproviralDNAformationand integra-tion in cells infected withmurine leukemia viruses.
Proc. Natl.Acad. Sci. U.S.A. 73:2236-2240.
9. Jolicoeur, P.,and E. Rassart. 1980.Effect ofFv-i gene
production synthesis oflinear and supercoiled viral DNA incellsinfected with murineleukemia virus. J.
Virol.33:183-195.
10.Keshet, E., J. O'Rear, and H. Temin. 1979.DNA of noninfectiousand infectiousintegrated spleen necrosis virus(SNV) is colinear with unintegrated SNV DNA and notgrossly abnormal. Cell16:51-61.
11. Krontiris, T. G., R. Soeiro, and B. N. Fields. 1973. Host restrictionof Friendleukemia virus: role ofthe viraloutercoat.Proc.Natl. Acad. Sci. U.S.A. 70:2549-2553.
12.Lilly,F., and T. Pincus.1973.Genetic control of murine
Fv-1 HOST RESTRICTION OF F-MuLV 55
viral leukemogenesis.Adv. Cancer Res. 17:231-277. 13. Oliff, A. I., G. L. Hager, E. Chang, E. M. Scolnick, H.
W. Chan, and D. R. Lowy. 1980. Transfection of molecularly cloned Friend murine leukemia virus DNA yields a highlyleukemogenic helper-independent type C virus. J. Virol. 33:475-486.
14. Pincus,T., J. W.Hartley, and W. P. Rowe. 1971. A major genetic locus affecting resistance to infection with murine leukemiaviruses. I. Tissue culture studies of naturally occurring viruses. J. Exp. Med. 133:1219-1233.
15. Pincus,T., J. W. Hartley, and W. P. Rowe. 1975. A major genetic locusaffecting resistance to infection with murineleukemia viruses. IV. Dose-response relation-ships in Fv-1 sensitive and resistantcell cultures. Virol-ogy 65:333-342.
16. Razin, A.,and A.Riggs. 1980. DNAmethylationand gene function.Science210:604-610.
17. Rigby, P. W. J., M. Dieckmann,C.Rhodes, and P. Berg. 1977.Labellingof DNAtohigh specificactivity
by nick translation. J. Mol. Biol. 113:237-258. 18.Rowe, W. P., W. E.Pugh, and J. W.Hartley. 1970.
Plaque assay techniques for murine leukemia viruses. Virology 42:1136-1139.
19. Schuh, V., M.Blackstein, and A. Axelrad. 1976. In-herited resistance to N-andB-tropicmurine leukemia viruses in vitro: titration patterns in strains SIM and SIM.R congenic at the Fv-1 locus. J. Virol. 18:473-480. 20.Shank,P.RK, S.H.Hughes,H. J.Kung,J. E.Majors,
N. Quintrell, R. V.Guntaka, J. M.Bishop,and H. E.Varmus. 1978.Mapping unintegratedavian sarcoma virus DNA: termini of linear DNAbear 300 nucleotides present once or twice intwospecies of circular DNA. Cell 15:1383-1395.
21.Smotkin,D., A. M.Gianni,S.Rozenblatt,and R. A. Weinberg. 1975. Infectious viral DNA of murine leu-kemia virus. Proc. Natl. Acad. Sci. U.S.A. 72:4910-4915. 22. Southern, E. M. 1975.Detection ofspecificsequences among DNAfragmentsseparated bygel
electrophore-sis. J.Mol. Biol. 98:503-517.
23. Sveda, M. M., and R. Soeiro.1976.Hostrestriction of Friend leukemia virus:synthesisandintegrationof the provirus. Proc. Natl. Acad. Sci. U.S.A. 73:235-2360. 24. Tennant,R. W., B. Schluter, W. K. Yang, and A.
Brown.1974.Reciprocalinhibitionofmouseleukemia virusinfection byFv-i allele cell extracts. Proc. Natl. Acad.Sci. U.S.A. 71:4241-4245.
25. Wahl, G. M., M. Stern,andG.Stark. 1979.Efficient transfer oflargeDNAfragments from agarosegelsto
diazobenzyloxymethyl-paperandrapid hybridization by
using dextran sulfate. Proc. Natl. Acad. Sci. U.S.A.76: 3683-3687.
26.Yang, W.K,J.0.Kiggans,D.Yang,C.Ou,R. W.
Tennant,A.Brown,and R. H. Bassin.1980. Synthe-sisand circularization of N- andB-tropic retroviral DNAin Fv-i permissive and restrictivemouse cells. Proc.Natl. Acad.Sci. U.S.A.77:2994-2998.
27. Yoshimura, F. K., andRKA.Weinberg.1979. Restric-tion endonucleasecleavageof linear and closedcircular murine leukemia viral DNAs:discovery ofasmaller circular form.Cell 16:323-332.
on November 10, 2019 by guest
http://jvi.asm.org/