Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Transfer of Primer Binding Site-Mutated Simian Immunodeficiency
Virus Vectors by Genetically Engineered Artificial
and Hybrid tRNA-Like Primers
ANETTE CHEMNITZ HANSEN,1THOMAS GRUNWALD,2ANDERS HENRIK LUND,1† ALEXANDER SCHMITZ,1
MOGENS DUCH,1KLAUS U¨ BERLA,2ANDFINN SKOU PEDERSEN1,3*
Department of Molecular and Structural Biology1and Department of Medical Microbiology and Immunology,3Aarhus University,
DK-8000 Aarhus, Denmark, and Institute of Virology, Leipzig University, D-04103 Leipzig, Germany2
Received 19 October 2000/Accepted 14 February 2001
Simian immunodeficiency viruses (SIV) harbor primer binding sites (PBS) matching tRNA3Lysor tRNA5Lys.
To study determinants of primer usage in SIV, a SIVmac239-based vector was impaired by mutating the PBS to a sequence (PBS-X2) with no match to any tRNA. By cotransfection of a synthetic gene encoding a tRNAPro-like RNA with a match to PBS-X2, the activity of this vector could be restored to a transduction
efficiency slightly lower than that of the wild-type vector. A vector with a PBS matching tRNAProwas functional
at a level slightly below that of the wild-type vector, but higher transduction efficiency could be obtained by cotransfection of a gene for an engineered tRNAPro-tRNA
3
Lyshybrid with a match to PBS-Pro. The importance
of tRNA backbone identity was further analyzed by complementing the PBS-X2 vector with a gene for a matching x2 primer with a tRNA3Lysbackbone, which led to three- to fourfold-higher titers than those observed
for the x2 primer with the tRNAPro backbone. In summary, our results demonstrate flexibility in PBS and
primer usage for SIVmac239, with PBS-primer complementarity being the major determinant, in analogy with previous findings for murine leukemia viruses and human immunodeficiency virus type 1.
Retroviruses replicate through reverse transcription of the viral RNA genome into a double-stranded DNA form that is integrated into the genome of the host. The virus-encoded reverse transcriptase (RT) enzyme mediates this process through the use of a host-encoded tRNA. The 3⬘-terminal 18-nucleotide (nt) segment of the tRNA primer molecule an-neals to the RNA genome at a complementary sequence known as the primer binding site (PBS). In order to synthesize a double-stranded DNA copy, two template shifts are involved. In plus-strand synthesis, the 3⬘ 18-nt segment of the tRNA primer is reverse transcribed into a DNA copy, which mediates the second template shift by annealing to the minus-strand DNA copy of the PBS. Accordingly, the PBS sequence of the transduced provirus may originate from a copy of the genomic PBS RNA sequence or from copying of the 3⬘ 18 nt of the tRNA primer.
Different tRNA species are used as primers among retrovi-ruses; however, for each group of viruses, the PBS is stably maintained during replication. Murine leukemia viruses (MLV) and human T-cell leukemia virus use a tRNAPro, the
avian sarcoma-leukosis virus (ASLV) group uses a tRNATrp,
and a tRNA3Lysis used as a primer for reverse transcription in
mouse mammary tumor virus and in human and simian immu-nodeficiency viruses (HIV and SIV, respectively) (reviewed in references 33 and 35).
Mechanisms of PBS maintenance during replication have
been studied in a variety of viruses. MLV mutated in the PBS to match the 3⬘end of other tRNAs are able to use noncognate primers in multiple rounds of replication (6, 30, 36). In con-trast, PBS-mutated HIV-1 and ASLV may initially replicate, but in time revert to their cognate tRNA during serial transfer of PBS-mutated viruses (7, 25, 47, 49). The selective reversion and maintenance of the cognate primer in these viruses may be accomplished by non-PBS determinants.
The lentiviruses of the SIV group have attracted attention due to the development of animal models for the study of AIDS (9) and the development of lentivirus vectors for gene therapeutic purposes (34, 43). Although SIVs and HIV-1 share common features, the SIVmac239 isolate used in this study differs from HIV-1 in lacking an adenosine-rich loop situated in the U5 hairpin (3, 4), which has been implicated in deter-mining primer specificity in HIV-1 (19–24, 27, 46, 52). To study the determinants for primer specificity in SIV, we have estab-lished a system in which the viral RNA genome as well as the tRNA primer may be manipulated. By using synthetic genes encoding tRNA-like molecules complementary to PBS-mu-tated SIV vectors, it is demonstrated that the PBS is the pri-mary determinant for primer specificity in initiation of first-strand synthesis and plus-first-strand transfer resulting in the formation of a provirus. In this experimental setup, it is pos-sible to study the effect on the transduction titers exerted by sequences outside the PBS-complementary 3⬘ 18 nt of the tRNA-like primer, because no endogenous competitor tRNA molecule is able to compete for binding to the mutant PBS.
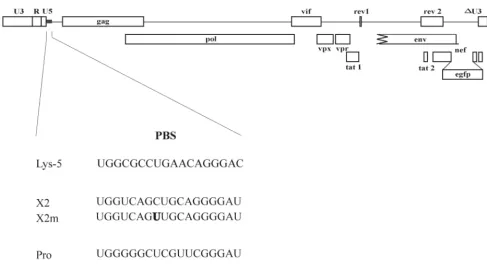
Design of PBS-mutated vectors and engineered tRNA-like primers. The PBS of a SIV-based green fluorescent protein (GFP) vector (SIVGFP) was mutated, and constructs encoding putative tRNA-like primers were designed. SIVGFP-Pro con-tains a PBS-Pro for which the corresponding tRNA proline is * Corresponding author. Mailing address: Department of Molecular
and Structural Biology, Aarhus University, C.F. Moellers Alle´, Bldg. 130, DK-8000 Aarhus, Denmark. Phone: 45 89423188. Fax: 45 86196500. E-mail: [email protected].
†Present address: Division of Molecular Carcinogenesis, The Neth-erlands Cancer Institute, NL-1066CX Amsterdam, The NethNeth-erlands.
4922
on November 9, 2019 by guest
http://jvi.asm.org/
found in the cell. SIVGFP-X2, however, harbors a PBS-X2 for which no matching endogenous tRNA exists (Fig. 1). In the case of SIVGFP-Pro, a PBS for the tRNA proline was intro-duced into the PBS of SIVGFP by PCR-mediated site-directed mutagenesis (29) in which primer set A (primers 1 and 2) (Table 1) and primer set B (primers 3 and 4) generated two PCR products with SIVGFP as a template. The two PCR
products were combined in an overlap extension reaction with primers 2 and 4. The resulting fragment was cloned into SIVGFP. For SIVGFP-X2, the PBS was mutated at 11 nt positions that were originally selected to give a minimal match to any known murine tRNA (28) with primer set A (primers 5 and 2) and primer set B (primers 6 and 4). The SIVGFP vector is anenvdeletion mutant of SIVmac239, in which thenefgene has been replaced by the enhanced GFP (EGFP) reporter gene (Fig. 1) (⌬envgfp in reference 43).
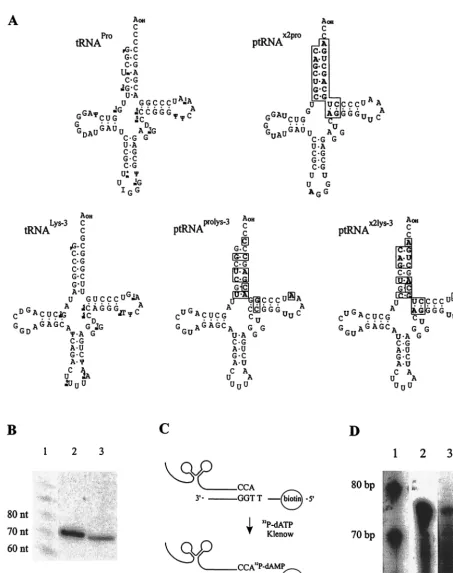
The constructs designed to encode the synthetic tRNA-like primer molecules ptRNAx2pro, ptRNAx2lys-3, and ptRNAprolys-3
were constructed from deoxyoligonucleotides and cloned as described in reference 28. ptRNAx2prois identical to ptRNAx2
in reference 28, whereas ptRNAx2lys-3and ptRNAprolys-3were
constructed by annealing the 22-nt elongation primer 7 to the 127-nt primers 8 and 9, respectively. The sequences of primers 8 and 9 were based on that of tRNA3Lys(GenBank accession
no. K01797) (41). The predicted structures of ptRNAx2proand
ptRNAprolys-3are shown in Fig. 2A. The 3⬘18-nt sequence of
ptRNAx2pro matches that of PBS-X2, but otherwise it
resem-bles the sequence of tRNAPro, whereas ptRNAprolys-3matches
PBS-Pro at the 3⬘end, but resembles tRNA3Lys. The synthetic
tRNA-like primers were processed correctly with regard to trim-ming of the tRNA precursor at the 5⬘and 3⬘termini, because the transcript sizes correspond to the expected 75 and 76 nt for ptRNAx2proand ptRNAx2lys-3, respectively (Fig. 2B), as shown
by Northern hybridization after transfection into human BOSC 23 packaging cells (37), as described in reference 28. The addition of the nonencoded CCA sequence to the trimmed 3⬘
[image:2.612.61.548.70.337.2]end of the synthetic tRNAs was verified by a tRNA tagging assay (31), in which the presence of the CCA tail was a de-FIG. 1. Vector structure. The SIVmac239-based vector SIVGFP is shown at the top. The sequences of the introduced PBS mutations are shown below. The mismatch marker mutation relative to the engineered corresponding tRNA-like primer is shown in boldface.
TABLE 1. Oligonucleotides used in this study
Primer Sequence (5⬘33⬘)
1 ...p-TTGGGGGCTCGTYCGGGATTTGAAGGAGAGTGA GAGACTCCTGA
2 ...CGGGCTTAATGGCAGGTGGACA
3 ...p-GTACTCAGGAGTCTCTCACTCTCCTTCAAATCCCG RACGAGGGGC
4 ...CGCTGAAACAGCAGGGACTT
5 ...p-TTGGTCAGYTGCAGGGGATTTGAAGGAGAGTGA GAGACTTCTGA
6 ...p-TTCAAATCCCCTGCARCTGACCAATCTGCTAGGG ATTTTC
7 ...GGAAAGCTTAAAAAAGACATGC
8 ...CCGGAATTCAAAAAGTAAAGCTCTCGTGAAGACA GCTGCTAGCTCAGTCGGTAGAGCATCAGACTTTT AATCTGAGGGTCAGGGGTTCAAATCCCCTGCAG CTGAGCATGTCTTTTTTAAGCTTTCC
9 ...CCGGAATTCAAAAAGTAAAGCTCTCTCGTGAAGAG GCTTAGCTCAGTCGGTAGAGCATCAGACTTTTAA TCTGAGGGTCCCGGGTTCAAATCCCGGACGAGC CCGCATGTCTTTTTTAAGCTTTCC
10 ...TGGTCAGCTGCAGGGGAT
11 ...Biotin-AAAAATTGGTCAGCTGCAGGGGAT 12 ...GGGGAATTCTGGAAGGGATTTATAAGAG 13 ...CCCCTTCCTGGATAAAAGACA
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.59.294.496.727.2]FIG. 2. (A) Structure of the murine tRNAPro(16) and tRNA 3
Lys(38) and putative structures of synthetic tRNAs. Mutations relative to tRNAPro
and tRNA3Lysare boxed. (B) Northern hybridization. Ten micrograms of total RNA from human BOSC 23 cells transfected with ptRNAx2lys-3(lane
2) or ptRNAx2pro(lane 3) was separated on an 8% polyacrylamide–urea gel, blotted, and hybridized to an X2-specific probe (primer 10). The DNA
markers are shown in lane 1. (C) tRNA tagging assay. Total RNA from BOSC 23 cells was incubated with a biotinylated oligonucleotide complementary to the 3⬘18 nt of tRNAPro(31) or tRNAx2lys-3(primer 11), respectively. Following magnetic separation, the annealed tRNA was
extended in the presence of [32P]dATP and separated on an 8% polyacrylamide–urea gel. (D) tRNA tagging gel showing total RNA from BOSC
23 cells incubated with the tRNAPro-specific oligonucleotide (lane 2). In lane 3, total RNA from BOSC 23 cells transfected with ptRNAx2lys-3was
incubated with primer 11. Lane 1 shows DNA size markers.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.74.527.70.643.2]signed prerequisite (Fig. 2C) for the synthesis of the radioac-tively labeled product seen in Fig. 2D.
A mismatch between vector PBS and primer was introduced by altering 1 nt in the PBS of SIVGFP-X2 and SIVGFP-Pro, yielding SIVGFP-X2m and SIVGFP-Prom, respectively (Fig. 1), to distinguish the PBS sequences of the primer and PBS origin during replication.
Complementation of PBS mutations by designed tRNA-like primers. The effect of PBS mutations on the infectivity of SIVGFP was determined by transient transfections of 293T cells (293ts/A1609) (10) with the different mutants of SIVGFP together with the vesicular stomatitis virus glycoprotein G (VSV-G)-encoding pHIT-G plasmid (14) following calcium phosphate coprecipitation (44). Table 2 shows the SIVGFP vector titers in the supernatant of the transfected cells deter-mined on 293A cells (Quantum Biotechnologies, Laval, Can-ada). 293A cells, a subclone of 293 cells, were used for titration since they show stronger adherence to plastic dishes than 293T cells. The transfection efficiency was monitored by determina-tion of capsid antigen levels in the supernatant of the trans-fected cells by employing an HIV-1 capsid enzyme-linked im-munosorbent assay kit (Innogenetics, Ghent, Belgium). The 293A target cells were seeded in 24-well plates at a density of 5⫻ 104 cells per well. The following day, the medium was
removed and cells were incubated for 2 to 4 h with serial dilutions of 200 l (per well) of the supernatant from the transfected cells. Two days after infection, the number of GFP-positive cells was counted. The infectivity of the vectors was calculated by dividing the vector titer by the amount of capsid antigen and is expressed relative to the infectivity of wild-type SIVGFP.
Mutation of the PBS of SIVGFP to the X2 sequence re-duced the titer by at least 4 orders of magnitude with essen-tially no background (Table 2). By the addition of a synthetic tRNA-like primer, ptRNAx2pro, matching PBS-X2, the
infec-tivity of SIVGFP-X2 was increased by at least 4 orders of magnitude and was only fivefold lower than the infectivity of SIVGFP wild-type particles. The PBS-mutated vector could therefore be complemented by the synthetic ptRNA-like primer matching the 18 nt of the PBS-mutated vector.
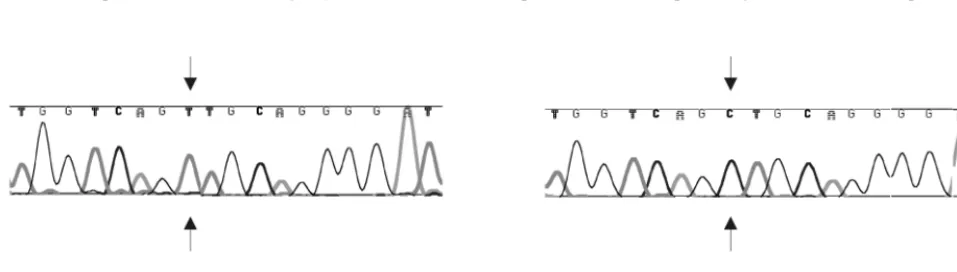
Direct sequence evidence for ptRNAx2pro usage was
[image:4.612.52.555.84.223.2]ob-tained from amplification with primers 12 and 13 and sequenc-ing the PBS region of a mixed pool of cell lysates originatsequenc-ing from transductions with the supernatant of SIVGFP-X2m transfected cells. In the left panel of Fig. 3, the PBS sequence of the PBS-X2m origin is shown, whereas the panel to the right shows the PBS sequence, originating from a copy of ptRNAx2pro, thus providing the evidence for primer usage.
FIG. 3. Genetic evidence for tRNAx2prousage. PCR products spanning the PBS region from lysates prepared from a mixed pool of transduced
[image:4.612.64.543.559.695.2]cells were cloned and sequenced. In the left and right panels are shown the PBS sequences resulting from a copy of the vector SIVGFP-X2m and primer tRNAx2pro, respectively. The mismatch positions are indicated by arrows.
TABLE 2. Vector transduction efficiencies
Plasmid cotransfected with VSV-G Results for:
PBS mutants tRNA/mock
Expt 1 Expt 2
Titer (GFU)/ng
of p27CAa Infectivity% b Titer (GFU)/ngof p27CAa Infectivity% b
SIVGFP pBluescript 5.5⫻105/2.7 100 1.5⫻105/9.2 100
SIVGFP-X2 pBluescript ⬍5/3.1 ⬍0.0008 5/11.4 0.0027
SIVGFP-X2 ptRNAx2pro 8.5⫻104/2.6 16 2.4⫻104/6 25.9
SIVGFP-X2m pBluescript ⬍5/5.7 ⬍0.0004 20/14 0.0088
SIVGFP-X2m ptRNAx2pro 9.8⫻104/2.7 18 8.7⫻104/6.1 87.5
SIVGFP-Pro pBluescript 7.7⫻104/8.7 4.3 2.4⫻104/14.6 10
SIVGFP-Pro ptRNAprolys-3 2.8⫻105/8.7 16 9.2⫻104/15.8 36
SIVGFP-Prom pBluescript 1.9⫻104/6.4 1.5 3.1⫻103/14.9 1.2
SIVGFP-Prom ptRNAprolys-3 1.8⫻105/7.7 11.5 2.1⫻104/11 11.9
aValues are in green fluorescence-forming units (GFU) and nanograms of p27CA antigen per milliliter of supernatant of 293T cells transfected with the indicated
plasmids.
bPercentage of infectivity (titer of p27CA antigen) relative to wild-type SIVGFP. Of each plasmid DNA, 2.5g was used for transfection of a six-well plate.
on November 9, 2019 by guest
http://jvi.asm.org/
According to the mechanism of reverse transcription, the PBS sequence of a transduced provirus originates from copying the genomic PBS RNA or the 3⬘18 nt of the tRNA primer. Con-sequently, it would be predicted that 50% of the clones would contain the primer sequence. It was demonstrated that 5 sub-clones out of 13 analyzed contained the artificial tRNA se-quence.
Effect of using primers with a tRNA3Lys-like backbone. A
comparison of the infectivity of SIVGFP-Pro in the presence or absence, respectively, of a hybrid primer, tRNAprolys-3,
which had an acceptor stem matching PBS-Pro but otherwise resembled the tRNA3Lysmolecule (Fig. 2A), showed that
co-transfection of ptRNAprolys-3increased the infectivity fourfold
(Table 2). This suggests that sequences or structures in the backbone of the cognate tRNA3Lys compared to the
endoge-nous tRNAProare important for optimal initiation of reverse
transcription or packaging. In these experiments, accurate quantification of the expression levels is difficult due to abun-dant expression of endogenous tRNAs closely homologous to the transfected tRNAs. However, primer selectivity by the ret-roviral machinery during primer packaging, placement, and utilization may be the determining factor rather than abun-dance of the tRNA. To compare the effects exerted by the tRNA sequences outside the PBS complementary region, ptRNAx2lys-3 was designed, containing a tRNA
3
Lys backbone,
but matching the PBS-X2 (Fig. 2A), and used in transient transfections. The infectivity of the SIVGFP-X2 obtained by cotransfection with ptRNAx2lys-3resulted in three- to
fourfold-higher transduction titers relative to cotransfection with ptRNAx2pro(Table 3).
While normalization of transfection efficiency by p27CA an-tigen levels within each experiment gave consistent results, normalization of transfection efficiency between different ex-periments was not found to be very useful. Although CA an-tigen levels differed largely between different experiments (compare Tables 2 and 3), vector titers did not seem to in-crease in a linear way with increasing p27CA antigen levels. Therefore, CA antigen levels might not be the limiting factor for vector titer, or increased toxicities at higher transfection rates might lead to stronger inhibition of formation of infec-tious vector particles than protein expression. To confirm that the vector titer depends on the amount and type of tRNA, two independent titration experiments were performed with cotransfection of increasing amounts of ptRNAx2pro and
ptRNAx2lys-3, respectively, in cotransfections with SIVGFP-X2
(Fig. 4A). For amounts of tRNA expression plasmid up to 1
g, the transduction titer increases for a given amount of vector DNA. Increasing the amount of ptRNA above the sat-urating level seems to have an inhibitory effect on transduction efficiency. Throughout the entire range of DNA concentra-tions, the cotransfection of ptRNAx2lys-3results in higher
trans-duction titers than those of ptRNAx2pro, with the exception of
the experiment using 2.5g of ptRNAx2pro. Such a preference
for ptRNAx2lys-3 was not observed in an MLV system using
PBS-mutated Akv-based vectors in which the addition of
tRNAx2proresults in slightly higher titers than those used for
addition of the ptRNAx2lys-3(Fig. 4B). Construction of
MLV-based vectors and the titer assay was described previously in reference 28. Thus, it seems that SIV shows a weak preference for synthetic tRNA-like primers with a tRNA3Lysbackbone.
Our data demonstrate that complementarity between the PBS and a matching synthetic tRNA-like primer is the primary determinant for selection and use of the tRNA species in initiation of reverse transcription in SIV. However, we note that the transduction titers are lower than for the wild-type vector. An MLV Akv-based vector carrying PBS-X2 was greatly impaired in its ability to replicate in single-round trans-fections, but could be rescued by the addition of the designed complementary tRNAx2pro, resulting in a complete provirus
(28). Similarly, an HIV-1 provirus with a PBS complementary to the yeast tRNAPhe relied on the transfection of a yeast
tRNAPhe, using a single-round transfection system (51). Thus,
SIVmac239 seems to be similar to MLV, ASLV, and HIV-1, in which the PBS and tRNA complementarity is the primary determinant for selection of the tRNA primer for initiation of reverse transcription and the ability to proceed through a com-plete replication cycle.
Northern analysis of the expression levels of the synthetic tRNAs relative to a control U2 small nuclear RNA probe (48) has shown that ptRNAx2lys-3is expressed at higher levels than
ptRNAx2proin the producer cells used (data not shown).
How-ever, it is not clear whether the level of tRNA abundance per se has any effect on transduction efficiency, because the amount of synthetic tRNA DNA added in the transfections seems to be saturating (Fig. 4). Secondary determinants such as non-PBS interactions may be responsible for the apparent preference for the tRNA3Lys backbone, as has been
[image:5.612.49.557.84.186.2]demon-strated for HIV-1. Viral genomic RNA sequences other than TABLE 3. Effect of tRNA backbone identity on vector transduction efficiencies
Plasmid cotransfected with VSV-G Results for:
PBS mutants tRNA mock
Expt 1 Expt 2 Expt 3
Titer (GFU)/ng
of p27CAa Infectivityb Titer (GFU)/ngof p27CAa Infectivityb Titer (GFU)/ngof p27CAa Infectivityb
SIVGFP-X2m pBluescript ⬍5/131 10/95 ⬍5/86
SIVGFP-X2m ptRNAx2pro 1.4⫻105/132 1⫻103 5.9⫻104/45 1.3⫻103 4.9⫻104/44 1.1⫻103
SIVGFP-X2m ptRNAx2lys-3 4.1⫻105/106 3.9⫻103 2.4⫻105/54 4.4⫻103 3.1⫻105/109 2.8⫻103
x2lys3/x2pro ratio 2.9 4 3.9 3.4 6.3 2.5
aValues are in green fluorescence-forming units (GFU) and nanograms of p27CA antigen per milliliter of supernatant of 293T cells transfected with the indicated
plasmids on 293A cells.
bInfectivity is given as the titer of p27CA antigen. ptRNA expression plasmid DNA (1.25g) was cotransfected with 2.5g of SIVGFP-X2 and 2.5g of pHITG.
The total amount of DNA transfected was adjusted to 10g for a six-well plate.
on November 9, 2019 by guest
http://jvi.asm.org/
the A-rich loop may interact with the primer in SIVmac239, although interactions between tRNAPro and the Moloney
MLV RNA have been found to be restricted to the PBS (13). In HIV-1, primer placement on the viral genome and primer packaging may depend on destabilization of the secondary structure of the tRNA and the viral RNA template by the p55gagprecursor (1, 5, 8, 12, 17, 18, 26, 32, 39, 40), which also
seems to be the case for avian leukosis virus, but not for MLV (15). Primer and RT interaction may also be necessary for initiation of reverse transcription (1, 2, 11, 42, 50), although RT may recognize the overall L-shaped tRNA rather than specific features of its cognate tRNA primer (45). Thus, we cannot exclude that one or several of the possible non-PBS interactions may contribute to the observed higher titers in SIV for the tRNA3Lys backbone in this study; however, the
primary determinant for primer specificity is the complemen-tarity between the viral PBS and the tRNA primer in SIV-mac239.
The first two authors contributed equally to this work and should both be considered the first author.
The technical assistance of Ane Kjeldsen and Katrin Bra¨utigam is gratefully acknowledged.
A. Schmitz acknowledges support by the German Academic Ex-change Service (DAAD/HSPIII) Program. This work was supported by Bavarian Nordic Research Institute A/S, the Karen Elise Jensen Foun-dation, the Danish Cancer Society, the German Research Foundation
(Ue45–3/1), and the Danish Natural Science and Health Science Re-search Councils.
REFERENCES
1.Barat, C., V. Lullien, O. Schatz, G. Keith, M. T. Nugeyre, F. Gruninger-Leitch, F. Barre-Sinoussi, S. F. LeGrice, and J.-L. Darlix.1989. HIV-1 reverse transcriptase specifically interacts with the anticodon domain of its cognate primer tRNA. EMBO J.8:3279–3285.
2.Barat, C., O. Schatz, S. Le Grice, and J.-L. Darlix.1993. Analysis of the interactions of HIV-1 replication primer tRNALys, 3with nucleocapsid
pro-tein and reverse transcriptase. J. Mol. Biol.23:185–190.
3.Berkhout, B.1997. The primer binding site on the RNA genome of human and simian immunodeficiency viruses is flanked by an upstream hairpin structure. Nucleic Acids Res.25:4013–4017.
4.Berkhout, B., and I. Schoneveld.1993. Secondary structure of the HIV-2 leader RNA comprising the tRNA-primer binding site. Nucleic Acids Res. 21:1171–1178.
5.Cen, S., Y. Huang, A. Khorchid, J.-L. Darlix, M. A. Wainberg, and L. Kleiman.1999. The role of Pr55gagin the annealing of tRNA
3
Lysto human
immunodeficiency virus type 1 genomic RNA. J. Virol.73:4485–4488. 6.Colicelli, J., and S. P. Goff.1987. Isolation of a recombinant murine
leuke-mia virus utilizing a new primer tRNA. J. Virol.57:37–45.
7.Das, A. T., B. Klaver, and B. Berkhout.1995. Reduced replication of human immunodeficiency virus type 1 mutants that use reverse transcription primers other than the natural tRNA3Lys. J. Virol.69:3090–3097.
8.De Rocquigny, H., C. Gabus, A. Vincent, M. C. Fournie-Zaluski, B. Roques, and J.-L. Darlix.1992. Viral RNA annealing activities of human immuno-deficiency virus type 1 nucleocapsid protein require only peptide domains outside the zinc fingers. Proc. Natl. Acad. Sci. USA89:6472–6476. 9.Desrosiers, R. C.1990. The simian immunodeficiency viruses. Annu. Rev.
Immunol.8:557–578.
10. DuBridge, R. B., P. Tang, H. C. Hsia, P.-M. Leong, J. H. Miller, and M. P. Calos.1987. Analysis of mutation in human cells by using an Epstein-Barr virus shuttle system. Mol. Cell. Biol.7:379–387.
FIG. 4. Titration of engineered tRNA-like DNAs differing in their tRNA backbone compositions. Dose-response curves of the effects of increasing amounts of engineered tRNA-like primer DNA ptRNAx2proand ptRNAx2lys-3on the infectivity of an SIV-based vector (A) and the titer
of an MLV-based vector (B) are shown. (A) Engineered tRNA-like primers were cotransfected in the indicated amounts with SIVGFP-X2 and pHITG into 293T cells. The infectivity (green fluorescence-forming units [GFU] per nanogram of p27CA) in the supernatant of the transfected cells was determined on 293A cells. (B) Engineered tRNA-like primers were cotransfected in the indicated amounts with the MLV-based vector pPBS-X2 (28) into the packaging cell line BOSC 23. Vector titers in the supernatant of the transfected cells were determined on NIH 3T3 cells. Black and grey bars represent cotransfections with ptRNAx2proand ptRNAx2lys-3, respectively. Replica experiments are shown for each amount of
ptRNA added.
on November 9, 2019 by guest
http://jvi.asm.org/
11.Dufour, E., J. Reinbolt, M. Castroviejo, B. Ehresmann, S. Litvak, L. Tar-rago-Litvak, and M. L. Andreola.1999. Cross-linking localization of a HIV-1 reverse transcriptase peptide involved in the binding of primer tRNALys3. J.
Mol. Biol.285:1339–1346.
12. Feng, Y. X., S. Campbell, D. Harvin, B. Ehresmann, C. Ehresmann, and A. Rein.1999. The human immunodeficiency virus type 1 Gag polyprotein has nucleic acid chaperone activity: possible role in dimerization of genomic RNA and placement of tRNA on the primer binding site. J. Virol.73:4251– 4256.
13. Fosse, P., M. Mougel, G. Keith, E. Westhof, B. Ehresmann, and C. Ehres-mann.1998. Modified nucleotides of tRNAProrestrict interactions in the
binary primer/template complex of M-MuLV. J. Mol. Biol.275:731–746. 14. Fouchier, R. A., B. E. Meyer, J. H. Simon, U. Fischer, and M. H. Malim.
1997. HIV-1 infection of non-dividing cells: evidence that the amino-termi-nal basic region of the viral matrix protein is important for Gag processing but not for post-entry nuclear import. EMBO J.16:4531–4539.
15. Fu, W., B. A. Ortiz-Conde, R. J. Gorelick, S. H. Hughes, and A. Rein.1997. Placement of tRNA primer on the primer-binding site requirespolgene expression in avian but not murine retroviruses. J. Virol.71:6940–6946. 16. Harada, F., G. G. Peters, and J. E. Dahlberg.1979. The primer tRNA for
Moloney murine leukemia virus DNA synthesis. Nucleotide sequence and aminoacylation of tRNAPro. J. Biol. Chem.254:10979–10985.
17. Huang, Y., A. Khorchid, J. Gabor, J. Wang, X. Li, J.-L. Darlix, M. A. Wainberg, and L. Kleiman.1998. The role of nucleocapsid and U5 stem/A-rich loop sequences in tRNA3Lysgenomic placement and initiation of reverse
transcription in human immunodeficiency virus type 1. J. Virol.72:3907–3915. 18. Huang, Y., A. Khorchid, J. Wang, M. A. Parniak, J.-L. Darlix, M. A. Wain-berg, and L. Kleiman.1997. Effect of mutations in the nucleocapsid protein (NCp7) upon Pr160gag-poland tRNALysincorporation into human
immuno-deficiency virus type 1. J. Virol.71:4378–4384.
19.Huang, Y., A. Shalom, Z. Li, J. Wang, J. Mak, M. A. Wainberg, and L. Kleiman.1996. Effects of modifying the tRNA3Lysanticodon on the initiation
of human immunodeficiency virus type 1 reverse transcription. J. Virol. 70:4700–4706.
20.Isel, C., C. Ehresmann, G. Keith, B. Ehresmann, and R. Marquet.1995. Initiation of reverse transcription of HIV-1: secondary structure of the HIV-1 RNA/tRNA3Lys(template/primer). J. Mol. Biol.247:236–250.
21. Isel, C., R. Marquet, G. Keith, C. Ehresmann, and B. Ehresmann.1993. Modified nucleotides of tRNA3Lysmodulate primer/template loop-loop
in-teraction in the initiation complex of HIV-1 reverse transcription. J. Biol. Chem.268:25269–25272.
22. Isel, C., E. Westhof, C. Massire, S. F. Le Grice, B. Ehresmann, C. Ehres-mann, and R. Marquet.1999. Structural basis for the specificity of the initiation of HIV-1 reverse transcription. EMBO J.18:1038–1048. 23. Kang, S. M., Z. Zhang, and C. D. Morrow.1999. Identification of a human
immunodeficiency virus type 1 that stably uses tRNALys1,2 rather than
tRNALys,3for initiation of reverse transcription. Virology257:95–105.
24. Kang, S.-M., Z. Zhang, and C. D. Morrow.1997. Identification of a sequence within U5 required for human immunodeficiency virus type 1 to stably maintain a primer binding site complementary to tRNAMet. J. Virol.71:207–
217.
25. Li, X., J. Mak, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A. Parniak.1994. Effects of alterations of primer-binding site sequences on human immunodeficiency virus type 1 replication. J. Virol.68:6198–6206. 26. Li, X., Y. Quan, E. J. Arts, Z. Li, B. D. Preston, H. de Rocquigny, B. P.
Roques, J.-L. Darlix, L. Kleiman, M. A. Parniak, and M. A. Wainberg.1996. Human immunodeficiency virus type 1 nucleocapsid protein (NCp7) directs specific initiation of minus-strand DNA synthesis primed by human tRNA3Lys
in vitro: studies of viral RNA molecules mutated in regions that flank the primer binding site. J. Virol.70:4996–5004.
27.Liang, C., X. Li, L. Rong, P. Inouye, Y. Quan, L. Kleiman, and M. A. Wainberg.1997. The importance of the A-rich loop in human immunodefi-ciency virus type 1 reverse transcription and infectivity. J. Virol.71:5750– 5757.
28.Lund, A. H., M. Duch, J. Lovmand, P. Jo¨rgensen, and F. S. Pedersen.1997. Complementation of a primer binding site-impaired murine leukemia virus-derived retroviral vector by a genetically engineered tRNA-like primer. J. Virol.71:1191–1195.
29. Lund, A. H., M. Duch, J. Lovmand, P. Jo¨rgensen, and F. S. Pedersen.1993. Mutated primer binding sites interacting with different tRNAs allow efficient murine leukemia virus replication. J. Virol.67:7125–7130.
30. Lund, A. H., J. Schmidt, A. Luz, A. B. So¨rensen, M. Duch, and F. S. Pedersen.1999. Replication and pathogenicity of primer binding site mu-tants of SL3–3 murine leukemia viruses. J. Virol.73:6117–6122.
31. Lund, A. H., A. Schmitz, F. S. Pedersen, and M. Duch.2000. Identification of a novel human tRNASer(CGA)functional in murine leukemia virus
repli-cation. Biochim. Biophys. Acta1492:264–268.
32.Mak, J., M. Jiang, M. A. Wainberg, M. L. Hammarskjold, D. Rekosh, and L. Kleiman.1994. Role of Pr160gag-polin mediating the selective incorporation
of tRNALysinto human immunodeficiency virus type 1 particles. J. Virol. 68:2065–2072.
33. Mak, J., and L. Kleiman.1997. Primer tRNAs for reverse transcription. J. Virol.71:8087–8095.
34. Mangeot, P. E., D. Negre, B. Dubois, A. J. Winter, P. Leissner, M. Mehtali, D. Kaiserlian, F. L. Cosset, and J.-L. Darlix.2000. Development of minimal lentivirus vectors derived from simian immunodeficiency virus (SIVmac251) and their use for gene transfer into human dendritic cells. J. Virol.74:8307– 8315.
35. Marquet, R., C. Isel, C. Ehresmann, and B. Ehresmann.1995. tRNAs as primer of reverse transcriptases. Biochimie77:113–124.
36. Nikbakht, K. N., C. Y. Ou, L. R. Boone, P. L. Glover, and W. K. Yang.1985. Nucleotide sequence analysis of endogenous murine leukemia virus-related proviral clones reveals primer-binding sites for glutamine tRNA. J. Virol. 54:889–893.
37. Pear, W. S., G. P. Nolan, M. L. Scott, and D. Baltimore.1993. Production of high-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad. Sci. USA90:8392–8396.
38. Raba, M., K. Limburg, M. Burghagen, J. R. Katze, M. Simsek, J. E. Heck-man, U. L. Rajbhandary, and H. J. Gross.1979. Nucleotide sequence of three isoaccepting lysine tRNAs from rabbit liver and SV40-transformed mouse fibroblasts. Eur. J. Biochem.97:305–318.
39. Remy, E., H. de Rocquigny, P. Petitjean, D. Muriaux, V. Theilleux, J. Pa-oletti, and B. P. Roques.1998. The annealing of tRNA3Lysto human
immu-nodeficiency virus type 1 primer binding site is critically dependent on the NCp7 zinc fingers structure. J. Biol. Chem.273:4819–4822.
40. Rong, L., C. Liang, M. Hsu, L. Kleiman, P. Petitjean, H. de Rocquigny, B. P. Roques, and M. A. Wainberg.1998. Roles of the human immunodeficiency virus type 1 nucleocapsid protein in annealing and initiation versus elonga-tion in reverse transcripelonga-tion of viral negative-strand strong-stop DNA. J. Virol.72:9353–9358.
41. Roy, K. L., H. Cooke, and R. Buckland.1982. Nucleotide sequence of a segment of human DNA containing the three tRNA genes. Nucleic Acids Res.10:7313–7322.
42. Sarih-Cottin, L., B. Bordier, K. Musier-Forsyth, M. L. Andreola, P. J. Barr, and S. Litvak.1992. Preferential interaction of human immunodeficiency virus reverse transcriptase with two regions of primer tRNALysas evidenced
by footprinting studies and inhibition with synthetic oligoribonucleotides. J. Mol. Biol.226:1–6.
43.Schnell, T., P. Foley, M. Wirth, J. Mu¨nch, and K. U¨ berla.2000. Develop-ment of a self-inactivating, minimal lentivirus vector based on simian immu-nodeficiency virus. Hum. Gene Ther.11:439–447.
44.Soneoka, Y., P. M. Cannon, E. E. Ramsdale, J. C. Griffiths, G. Romano, S. M. Kingsman, and A. J. Kingsman.1995. A transient three-plasmid expression system for the production of high titer retroviral vectors. Nucleic Acids Res.23:628–633.
45.Thrall, S. H., J. Reinstein, B. M. Wohrl, and R. S. Goody.1996. Evaluation of human immunodeficiency virus type 1 reverse transcriptase primer tRNA binding by fluorescence spectroscopy: specificity and comparison to primer/ template binding. Biochemistry35:4609–4618.
46.Wakefield, J. K., S.-M. Kang, and C. D. Morrow.1996. Construction of a type 1 human immunodeficiency virus that maintains a primer binding site complementary to tRNAHis. J. Virol.70:966–975.
47. Wakefield, J. K., A. G. Wolf, and C. D. Morrow.1995. Human immunode-ficiency virus type 1 can use different tRNAs as primers for reverse tran-scription but selectively maintains a primer binding site complementary to tRNA3Lys. J. Virol.69:6021–6029.
48. Westin, G., J. Zabielski, K. Hammarstrom, H. J. Monstein, C. Bark, and U. Pettersson.1984. Clustered genes for human U2 RNA. Proc. Natl. Acad. Sci. USA81:3811–3815.
49. Whitcomb, J. M., B. A. Ortiz-Conde, and S. H. Hughes.1995. Replication of avian leukosis viruses with mutations at the primer binding site: use of alternative tRNAs as primers. J. Virol.69:6228–6238.
50. Wo¨hrl, B. M., B. Ehresmann, G. Keith, and S. F. Le Grice.1993. Nuclease footprinting of human immunodeficiency virus reverse transcriptase/ tRNALys-3complexes. J. Biol. Chem.268:13617–13624.
51. Yu, Q., and C. D. Morrow.1999. Complementarity between 3⬘ terminal nucleotides of tRNA and primer binding site is a major determinant for selection of the tRNA primer used for initiation of HIV-1 reverse transcrip-tion. Virology254:160–168.
52. Zhang, Z., S. M. Kang, A. LeBlanc, S. L. Hajduk, and C. D. Morrow.1996. Nucleotide sequences within the U5 region of the viral RNA genome are the major determinants for a human immunodeficiency virus type 1 to maintain a primer binding site complementary to tRNAHis. Virology226:306–317.