0022-538X/93/084513-08$02.00/0
Copyright X)1993, American Society for Microbiology
Recombinant
Human
Hepatitis B
Virus Reverse Transcriptase
Is
Active in
the Absence
of
the Nucleocapsid
or
the
Viral
Replication
Origin, DR1
MARIASEIFER1 AND DAVIDN.
STANDRINGl12*
HormoneResearch
Institute'*
andDepartmentofBiochemistry andBiophysics,2UniversityofCalifornia atSan
Francisco,
SanFrancisco,
California
94143-0534Received 11February 1993/Accepted 28 April 1993
The double-stranded DNA genome of hepatitis B virus (HBV) is reverse transcribed from the viral pregenomeRNAtemplate bya
virally
encodedreversetranscriptase enzyme (RT) that possesses both priming andelongationactivities. Prior efforts have failedtoexpressanactive form of HBV RT outside thenucleocapsidinanimal cellsortorelease it from viralnucleocapsids, thusrestricting the characterization of this important enzyme.Here,wehaveengineeredepitope-taggedHBV RTproteinsandexpressedtheminXenopusoocytesvia asynthetic RT mRNA which doesnotinclude the viral capsidproteinorthe known initiation site for viral DNA synthesis, DR1. We demonstrate the production ofanimmunoprecipitable 96-kDa HBVRTprotein and show, usingasimpleinvitro RT assay, that oocyte
lysates
containingthisprotein possessanactivity
that(i) catalyzesanRNA-dependentdeoxynucleotidetriphosphate polymerizationreactionby usinganas-yet-unidentifiedRNA
templateand(ii)is sensitivetothe RTinhibitorsactinomycin Dandphosphonoformate. Experimentswith the chain terminator ddATP suggest thatasignificantamountof chainelongationoccursinourin vitro reaction. Electrophoretic
analysis
reveals aheterogeneous array ofRTreactionproducts with sizes ranging from about 100 basestofar larger than that of the input RT mRNA. These products appeartocontaincovalently
bound protein,consistent with the notion that the RTproteinmayhaveprimed their synthesis. We conclude that HBV RTactivity can beuncoupled from both the nucleocapsid and thereplication origin, DR1. Our results raise thepossibilitythat unless HBVemploysnovelmechanisms toregulateits
constitutively
activeRT, cellularRNAs may be reverse transcribed during HBV infection, with potential implications for the development of HBV-related livercancer. The use of the oocyte system should facilitate studies of HBVRT, includingthe development of HBV RT inhibitors for antiviraltherapy.Thereversetranscriptaseenzyme(RT)of humanhepatitis B virus (HBV) has properties that set it apart from its retroviral counterparts. Although retroviral RTs replicate their genomes postinfection within particulate complexes
that may(2) ormaynot(5)contain theviralcapsid protein, the HBV RT isunusual in being able to convert its prege-nomic RNAtemplate into double-stranded DNA inside the
newly assembled viral nucleocapsid (33) in a process that
specifically depends on the HBV p21.5 capsid or core
protein (11, 18, 24,36). Althoughthe HBV pregenome RNA exhibits a retrovirus-like arrangement of open reading
frames(ORFs), in which the RT ORFoverlapsthe3' end of thecore ORF,geneticstudies(3, 26)reveal that HBV RTis translated as a discrete species; there is no evidence fora ribosomal frameshifting event that would yield a core-RT precursoranalogous to the retroviral gag-pol fusion proteins
(14).The final unusualfeature of the HBV RT protein is that it possesses an intrinsic priming activity (see below) that initiates first-strand DNA synthesis and leaves RT (or at least part ofit) covalentlylinkedto the viral DNA(9).
In addition to playing apivotal role in the initiation and
elongation phases ofreverse transcription, RTis essential for the selective uptake of the pregenome RNA into the
nucleocapsid (1, 13). RT, alongwith p21.5 and the prege-nome RNA, isclearlyaparticipantin the undefined
hierar-chyofinteractions that direct theassemblyof thereplicative nucleocapsid. Unfortunately, attempts to study RT at a
*Correspondingauthor.
biochemical level have been stymied by the lack of well-defined antibodies against RT and also by the failure to releaseactive RT fromnucleocapsids(21)or toexpressit in anucleocapsid-freeform inanimal cells. In this study, we set out to remedy this situation by expressing, in Xenopus oocytes, a tagged RTmolecule bearing an in-frame foreign
epitope to render it immunoprecipitable. We describe the successfulaccomplishment of this goal and further show that the resultant RTproteinunexpectedlydisplaysRTactivity.
MATERIALSANDMETHODS
Plasmids. All constructs were generated by standard re-combinant DNAtechniques.TocreateplasmidpSP64T-RT,
the bulk of the RT gene and a small piece of 3' flanking sequenceweremobilizedas arestrictionfragment and the 5' end of the genewas rebuilt via a synthetic oligonucleotide adaptor. The twofragmentswere coligated downstream of theSP6 promoter inplasmid pSP64T(16). Polymerasechain reactionoligonucleotide insertion was used to create epitope fusions bearing in-frame versions of either the FLAG
(Im-munex) epitope (Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys) or the Lepitope
(Asp-His-Gln-Leu-Asp-Pro-Ala-Phe-Gly-Ala-Asn-Ser)corresponding toresidues 27 to38 of theviral LSprotein (HBV subtype adw). These epitopes were intro-ducedimmediately after the initiatingRTMet tocreate the FLAG-RT andL-RT constructs. IntheRT-LandRT-FLAG constructs, the epitopes were inserted between RT amino acids 174 and 175attheuniqueBstEIIsite present inthe RT 4513
on November 9, 2019 by guest
http://jvi.asm.org/
4514 SEIFER AND STANDRING
gene (HBV subtype adw). Mutant fsL-RT is a frameshift mutantwhich is basedonthe L-RTepitopefusionconstruct (see Results) and contains a 1-base deletion located 11
residues downstream from the initiating Met codon (i.e., just beyond the Lepitope), resulting in a termination codon 15
residues later. AllconstructswereverifiedbyDNA
sequenc-ing.
RNA transcription and translation methodology. Capped SP6mRNAs wereprepared and concentrated 2-fold (forin
vitro translations)or15-fold (foroocytestudies)asdescribed elsewhere (28). In vitro translations were performed with reticulocyte lysates supplemented with [35S]methionine (NEN-Dupont) and the appropriate RNAas recommended
bythe manufacturer (Promega). Proteinswere resolvedby
sodium dodecyl sulfate (SDS)-polyacrylamide gel electro-phoresis (PAGE) on either 10 or 8% gels and detected by
exposingthe dried gels toX-rayfilm. Immunoprecipitation ofproteins bearingthe Lepitopewasperformed (30)with 1 ,g of anti-L monoclonal antibody MA 18/7 (12). Xenopus oocytemicroinjectionandlabeling methodologieshave been described earlier (30). Batches of 8 to 12 healthy oocytes
were injected with -40 nl of the appropriate mRNA per
oocyteand labeledmetabolicallywith 0.75 mCi ofamixture of [35SJmethionine and [35S]cysteine (Expresslabel; NEN-Dupont)perml for 12to16 hat17°C. Lysateswereprepared
in 200
RI
of homogenization buffer (30) containing 1 mM phenylmethylsulfonyl fluoride, 1 mM benzamidine, 1 p,M leupeptin-hemisulfate,and 2 ,ugofaprotininperml, clarified by centrifugation,andimmediately subjectedto immunopre-cipitationand SDS-PAGEessentiallyas described above.In vitro reverse transcription assays. Care was taken to inject oocytes with equal amounts of the L-RT or fsL-RT mRNAs, which were generally prepared in parallel and checked for concentration by agarose gel electrophoresis and ethidium bromide staining. LS mRNA was generally
used as a control in amounts that were at least equimolar with theappropriateRT mRNA.Microinjectedoocyteswere
incubated for 12to 14 hand thenhomogenizedinastandard HBVpolymerasebuffer (50mM Tris-HCl [pH 8.0]-40 mM MgCl2-50mMNH4Cl-1% [vol/vol]Nonidet P-40-0.3% [voV vol] 2-mercaptoethanol) containing 1 mM phenylmethylsul-fonyl fluoride,1 p,M leupeptin, and1 p,gofaprotininperml. For RTassays,10-,ul samplesof clarified(Eppendorf centri-fuge; 15,000 xg,10min) lysates (=0.5 oocyte)werediluted in 100 ,ul of polymerase buffer containing 0.2 mM each dCTP, dGTP,anddTT-lP (ultrapure; Pharmacia)and -10,Ci (34to125nM)of[a-32P]dATP (NEN-Dupont). Assays using other labeled nucleotideswereperformed analogously.After 1 h at37°C, the reactionswere stopped by the addition of EDTA (to 25 mM) and the reaction mixtureswere treated with trichloroacetic acid-sodium
PPi
(10 and 1%, wt/vol, respectively) in thepresenceof50 p,gof carrier tRNA. The precipitateswerecollectedbyfiltration throughGF/Cmem-branes(Whatman).The filterswerewashedextensivelyand dried, and radioactivitywas measuredbyCerenkov
count-ing.
For the RT timecourseexperiment,oocyteswereinjected with either L-RT mRNA or an LS mRNA control (see Results). At the appropriate time points, batches of unla-beled oocytes were homogenized and subjected to the RT
assay.
Analysis ofthe nucleic acid productsofreverse
transcrip-tion.Large-scaleRTassayswereperformedwith L-RT- and
fsL-RT-expressing lysates essentially as described above. Aliquots (one-quarter) of each reaction mixture were
ex-tracted twice withphenolwithorwithoutprior digestionfor
45 min at
37°C
with 1 mg ofproteinase
K per ml in the presence of 0.5%(wt/vol)
SDS. Nucleic acids were thenprecipitated,
washed,
dissolvedin TE(10
mMTris-HCl[pH
8.0],
1 mMEDTA), electrophoresed through
1% agarosein0.5xTBE
(44.5
mMTris,
44.5 mMborate,
0.5 mMEDTA),
and visualized
by
autoradiography
of the driedgel.
Theremaining
halves of the RTreaction cocktails weredepro-teinized
(as
describedabove), adjusted
to 10 mM sodiumphosphate
(pH
6.8),
and loaded onto small columns ofhydroxylapatite
(Biogel
HTP;
Bio-Rad).
After extensivewashing
withloading buffer,
single-
and double-stranded nucleic acidswere elutedfrom the columnwith successivesteps of 127 and 187 mM sodium
phosphate
(pH 6.8),
respectively, according
to the manufacturer'sinstructions.Samples
of each eluate werequantitated by
scintillationcounting.
Theremaining
nucleic acidswereprecipitated
(8)
with 5 mM EDTAand 0.1%
(vol/vol)
cetylpyridinium
bro-mide and
analyzed by
agarosegel
electrophoresis
asde-scribed above.
Effects of RNase A and inhibitors ofreverse
transcription.
For RNase A and RT inhibitionexperiments,
we usedlysates prepared
from RT-L- orLS-microinjected
oocytes. To testthe RNAdependence
of RTactivity,
RT-Land LSlysates
fromtwooocyteswereincubatedwithorwithout 20jig
of RNase A(DNase-free)
per ml for 10 min at37°C.
Subsequently,
double-scale,
200-,ul
RT assays wereper-formed
essentially
as describedabove,
exceptthatasimpli-fiedRTbuffer
(50
mMTris-HCl[pH 8.0],
10mMMgCl2,
60 mMNaCl,
10 mMdithiothreitol)
replaced
the HBVpoly-merasebufferand the reactionswere conducted for 30 min rather than 1h.RTinhibitionstudieswere
performed
under thesame conditions in assayssupplemented
with(i)
actino-mycin
D(100 pg/ml;
Sigma), (ii) phosphonoformate
(PFA) (1
mM;
foscarnet;
Sigma),
or(iii)
ddATP(0.83 ,uM;
Boehringer
Mannheim) (see
thelegend
toFig.
5).
The RTnucleicacidproducts
werepurified by
deproteinization,
extraction,
andprecipitation,
resolvedby
agarosegel
electrophoresis,
andvisualized
by
autoradiographyof thedriedgels,
asdescribedabove. The amountsof32P-labeled
products
presentineachlaneorin selected
regions
of eachlanewerequantitated by
scanning
the driedgels
with aphosphorimager
(Molecular
Devices).
RESULTS
Strategyfor
expression
ofanimmunoprecipitable
HBVRT. Thestructureof the HBV pregenomeRNA,
whichserves asthereverse
transcription template
aswellasthe mRNA for boththe 21.5-kDacapsid
(C) protein
and the RTprotein,
is shown inFig.
la. The HBV RTprotein
(with
apredicted
molecular massof -94.7
kDa)
is encodedby
acontiguous
-2.4-kb RT ORF that
overlaps
the 3' end of the CORF. The SORF,
whichspecifies
three relatedenvelope
(S) proteins,
lies
completely
within the RT ORF but utilizes a differentreading
frame.During
HBVinfection,
these Sproteins
aretranslated fromaclass of
subgenomic
mRNAs(32).
Repeats
foundat each end of the pregenome RNA contain
specific
sequenceelementscrucial for HBV
replication,
suchastheencapsidation
signal,
£,whichensurestheselectivepackag-ing
ofthe pregenome RNA into thenucleocapsid
(15),
andthe
replication
origin,
DR1. Intheproposed
(25, 27)
HBVreplication scheme,
the amino-terminaldomain of the mul-tifunctional(20)
RTprotein (Fig.
lb)
initiates first-strand(minus-strand)
DNAsynthesis
at the 3' copy of the short direct repeat DR1. RT remainscovalently
attached to theJ. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
a
DR2 DR1 I
*HAAA
3b
DR1 5§_ -ORF.|
R/
L/RT RT/L FLAGIRT RT/FLAG
5G.=
t
fsL/RT
FIG. 1. Structure of HBV pregenome RNA and synthetic RT mRNA.(a) Pregenome RNA(-3.5kb) with capped 5'(filled circle) andpolyadenylated3'(AAA) ends. Redundantsequences(R)bear onecopyeach of the short directrepeatDR1andanencapsidation signal (£). DR2 isafurther copyofDR1located before the3' R. The 3'DR1elementand DR2provide the initiation sites for first-strand (minus-strand) and second-strand (plus-strand)HBV DNA synthe-sis, respectively (27). Also shown are ORFs for the p21.5 core
protein(C-ORF) and the overlappingRTprotein(RT-ORF); S-ORF (see below) is presentin this RNAbut isnot shown. Theregion betweenthevertical dottedlines represents the sequence that was usedto createplasmid pSP64T-RT (seebelow). (b)Structure of RT mRNA. The RTORFand ashortpiece of 3'flank (line)lie between theXenopus p-globin (, G.) 5' and 3' untranslated sequences. Patternedsegmentswithin theRTORFrepresentfourproposed (20) domains: theterminalprotein(TP) domain, thetetherregion,the RT activity domain, and the RNase H domain. S-ORF, shown below RT-ORF, encodesthe three HBVenvelope proteins; the
approxi-matelocationof the naturalLepitope is indicated byanasterisk. Arrows indicate approximate locations at which the indicated epitopesor aterminationcodonwereintroduced.
elongating minus-strand DNA. Second-strand (plus-strand) DNAsynthesis is primed by an RNA primer and initiates at DR2, which is identical in sequence to DR1 but is located nearthe 3' end of the RT ORF.
Weexpressed the RT protein using a synthetic RT mRNA (Fig. lb). Briefly, this SP6 mRNA contains a minimal RT ORF and ashortpiece of 3' flanking sequence (correspond-ing to the region of the pregenome RNA included between the dotted lines in Fig. la) embedded between the transla-tionally enhancing Xenopus ,-globin flanking sequences (16). According to the HBV replication model (described above), RT mRNA should not be a template for reverse transcription, since it lacks both £ and DR1. To monitor RT production, RT was tagged with either the FLAG (19) epitope or anepitope (L) (29) from the HBV large surface (LS) protein. Epitopeswereinserted in framewith RT either into the amino terminus (FLAG-RT and L-RT fusions) or between RT residues 174 and 175, which lie at the border of anonfunctional tether region (20) of the RT (RT-FLAG and RT-L fusions). We also created a mutant L-RT (fsL-RT)
bearingaframeshiftnearthe 5'end of the L-RT ORF. In this mutantthe L-RTproteintranslation terminatesshortlyafter the frameshift. The epitope and frameshift positions are shown inFig. lb, which also summarizes the approximate
boundaries of the known RTsubdomains (20).
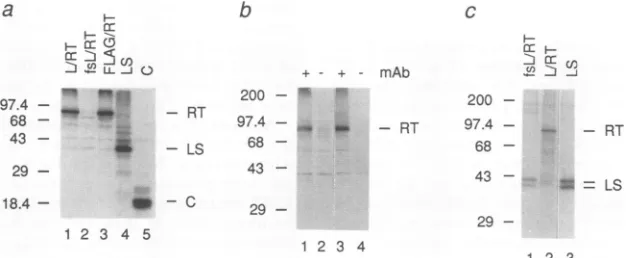
Expression of HBV RT in reticulocyte
lysates
and in frog oocytes. Upon translation in reticulocyte lysates (Fig. 2a),the capped L-RT and FLAG-RT mRNAs, but not fsL-RT mRNA, gavethepredicted-96-kDa35S-labeledproteins,as did RT-FLAG and RT-L mRNAs(datanotshown). The RT RNAs weretranslated lessefficiently than mRNAsencoding
the HBV LS and C proteins. Anti-L monoclonal antibody
MA 18/7 (12) specifically immunoprecipitated both native and denatured forms of L-RT (Fig. 2b) and RT-L but not wild-type RT(data not shown). No 96-kDa bandwas seen when MA 18/7 was omitted or replaced with a control monoclonalantibody of the same isotype(data not shown).
Curiously,we wereunabletospecifically immunoprecipitate
FLAG-RTorRT-FLAG fusions with any of four different anti-FLAG antibodiesunder awide variety of
immunopre-cipitation conditions (datanotshown).
rr
.--
u-0
97.4
-68 - M - RT
43- -LS
29
-18.4 - - C
1 2 3 4 5
b
+ + - mAb
200 - " 97.4 ,,u
68
-43
-29
-1 2 3 4
c
0 ) D J
200
-97.4 -RT
68
-43
-= = LS
29
[image:3.612.67.305.76.248.2]1 2 3
FIG. 2. Expressionof RT invitro andinXenopusoocytes.(a) Translation of mRNAs encodingtheL-RT, fsL-RT, FILAG-RT, LS,and C HBVproteinsinreticulocyte lysatessupplementedwith[35S]methionine. Proteinswerevisualizedbyautoradiographyafter SDS-PAGE.
(b)Monoclonal anti-LSantibodyMA18/7 (mAb) immunoprecipitatesboth native and denatured L-RTprotein.The35S-labeledL-RTproducts
wereimmunoprecipitatedfrom10-pl portionsof invitro translationreactions either in native form(lanes1 and2)orafterdenaturation(lanes
3 and4) (by boilingfor 2minwith1%[wt/vol] SDS). MA18/7wasomitted in lanes 2and 4.ImmunoprecipitationofFLAG-taggedRTswas
attempted unsuccessfully (data notshown) under the same conditionswith commercially available Ml and M2 monoclonal anti-FLAG antibodies(IBI)and withanM5monoclonalantibodyandarabbitanti-FLAGserumkindly provided byImmunex(Seattle, Wash.). (c) Only
oocytesmicroinjectedwith L-RT mRNAsynthesizeanimmunoprecipitable 96-kDa RTprotein.Translationsof the fsL-RT and LS mRNAs
are shown. The positions of prestained (a and b) or "4C-labeled (panel c) marker proteins (sizes in kilodaltons) (Bethesda Research
Laboratories) areindicated.
a
_ .. _
1-R
3--B
G.--]
aseH
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.157.476.498.627.2]4516 SEIFER AND STANDRING
12 dCTP A
* dATP
None LS L/RT RT
(1 5x) (I x)
LS D
URT
6 10 25 34
Time[h]
10.
B
E
0c
U.d
URT fsURT LS
E
0. le
URT fsURT
FIG. 3. Xenopus oocytesexpress anenzymatically active HBV
RT. Crude oocyte lysates stimulate incorporation of radiolabeled
deoxynucleotide triphosphates into acid-insoluble products (Aand
B). (A) Incorporation of [a-32P]dCTPor [a-32P]dATP directed by
lysates of oocytes injected with 15x L-RT, lx RT, LS, or no
mRNA(none). (B) Enhanced incorporation activity dependsonthe
L-RT protein. Shown is a comparison of the activities present in
lysates from oocytes injected with 15x L-RT and 15x fsL-RT
mRNAs. Lysates ofLS-expressing oocytesprovide acontrol. (C)
Immunoprecipitated L-RT protein retains activity. Proteins were
immunoprecipitatedfromfresh L-RTand fsL-RT lysateswith anti-L
monoclonal antibody (MA 18/7) and immediately assayed for RT
activityasdescribedinMaterialsandMethods. (D) RTactivity time
course.Unlabeled L-RTorLSproteinsweresynthesized inoocytes
fortheindicated times. At each timepoint, batches of fiveoocytes
were homogenized and subjected to RT in vitro activity assays.
Furtherdetailsare supplied in Materials and Methods.
We next examined the translation of concentrated L-RT mRNAs injected into Xenopus oocytes (Fig. 2c). L-RT mRNA, but not the fsL-RT and LS mRNAs, directed productionofa -96-kDa35S-labeled L-RTproteinthatwas
immunoprecipitated byMA18/7 andappearedtobeidentical
totheinvitroproduct by SDS-PAGE.Interestingly,all three
of these RNAs generated immunoprecipitable 39- and 42-kDa LS proteins (31) whichwere not detected for control oocytes (data not shown). For the L-RT and fsL-RT mRNAs, these LS species presumably arise by internal initiation of translation, which seemingly increases upon
abrogationoftheRTORF.
Production ofan enzymatically active RT in oocytes. We next tested whether the oocyte-produced RT proteins had RT activity. Crude oocyte lysates were incubated in a
polymerase buffer(20) inthe presence ofonea-32P-labeled
andthreeunlabeleddeoxynucleotide triphosphates.No
tem-plateorprimerwas added. After incubatingthemixturefor
1 h at 37°C, we measured the conversion of 32p into a
trichloroaceticacid-precipitable form. Remarkably, this sim-ple RT assay revealed (Fig. 3A) that lysates from oocytes
injected with concentrated L-RT mRNA gave upto 20-fold-stronger signals than lysates from oocytes that were either
uninjectedorinjectedwith concentrated LS mRNA. Similar
results were obtained whether the
a-32P-labeled
deoxynucle-otidewasdATP,dCTP(Fig. 3A),ordGTP(datanot
shown).
Another representative experiment (Fig. 3B) revealed an -10- to 19-fold-stronger signal for oocytes injected with L-RT mRNAthan for thoseinjectedwiththe LS andfsL-RT
mRNAs.Theseresults have been confirmed in -20
indepen-dent experiments (data not
shown).
Positive signals were seen only for oocytes injected with concentrated mRNAsencoding any of the RTs and typically varied from 4- to
10-fold above the background level, although we have seen
signals as faint as 2-fold and as strong as 20-fold. A wide
variety of concentrated mRNAs encoding other proteins were tested, but all gave negative results (Fig. 3 and datanot
shown).
The likelihoodof a link between the L-RT protein andthe
putative RT activitywas strengthened by the results of an immunoprecipitation experiment (Fig. 3C) using antibody
MA 18/7 and lysates from L-RT- or fsL-RT-expressing
oocytes. Both the washed immunoprecipitates and samples of theoriginal lysateswerethenassayed for RTactivity.The untreated L-RT lysate displayed 13-fold more activityin the RT assay than did the fsL-RT control (data not shown).
After immunoprecipitation, this activity differenceincreased
to-18-fold, despite a low overall recovery of RT activityby this method.
We also tested (Fig. 3D) how the level of RT activity
present in oocytes depends on the time allowed for protein
synthesis postinjection. Batches of oocytes were injected
with either L-RT mRNA or LS mRNA, incubated for 3, 6, 10, or 25 h prior to lysis, and then immediately subjected to the RT assay. In this experiment, RT activity exceedingthe LS control background level was first detected after -10h of incubation and was still seen after 25 h, though at reduced levels; further experiments have verified that 12 to 14 h postinjection is the optimal time for assaying HBV RT activity in oocytes (data not shown). A parallel analysis of L-RT protein synthesis in
oocytes
(by steady-state labeling of the oocyte proteins with[3
S]Met and[35S]Cys
followed by immunoprecipitation and SDS-PAGE) suggested that the level of L-RT protein is maximal between 6 and 10 h and diminishes slightly by 25 h (data not shown). Thus, there is a general correlation between the levels of RT activity and quantities of the 96-kDa L-RT protein, but the large amount of L-RT protein with no detectable RT activity seen at the 6-h time point suggests that the emergence of RT activitymay lag behind protein production.
Analysis of nucleic acid products of the RT reaction. Next, the products from the in vitro RT assay were deproteinized with proteinase K and phenol and then analyzed by agarose gel electrophoresis. In a representative experiment (Fig. 4a), L-RT-expressing lysates clearly generated more
32P-labeled
products (lane 1) than did the fsL-RT control (lane 3). Although small products (including discrete species with apparent sizes of -150 and 600 bases) predominated, the array of RT products had lengths which extended to at least 23,000 bases. When proteinase K treatment was omitted, phenol extraction eliminated the specific L-RT products (Fig. 4a, lane 2) but not most of the fsL-RT background (lane 4). Since HBV DNA genomes bearing covalently bound RT protein are extracted into phenol (9), this result is consistent with the notion that the RT protein had primed DNA synthesis in oocytes and remained covalently bound to the products. The deproteinized L-RT and fsL-RT products
30
20
10
50-
40-30.
20
E 0.
U.e
E
C)
0
3
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.58.297.72.332.2]b
- P. K
c ss ds
E
C.)
1 2 3 4 127 mM 187 mM
[image:5.612.135.485.60.227.2]M 1 2 3 4 sodiumphosphate pH 6.8
FIG. 4. Analysis of nucleic acid products ofreversetranscription reactions. (a)Mostof the in vitro labeled32P-RTproductsarecovalently
linkedtoprotein. Lanes1and2, L-RT reaction products; lanes 3 and 4,fsL-RT reaction products. Addition (+)(lanes 1 and 3) and omission (-) (lanes 2 and 4) ofproteinase K (P.K)areindicated above the panel. Positions ofDNAsize markersareshownonthe left.(b)Separation
ofsingle-stranded (ss) and double-stranded (ds) RT products by hydroxylapatite chromatography. Lane1,ssL-RT products; lane 2,ssfsL-RT
products; lane 3, ds L-RT products; lane 4, ds fsL-RT products. (c)Quantitation ofssand ds products byCerenkov counting.
werefurther characterized byhydroxylapatite
chromatogra-phy, whichpermits resolution of single- and double-stranded nucleic acid species. Thetwospecieswerethen analyzed by agarose gel electrophoresis (Fig. 4b) and quantitated by
scintillationcounting (Fig. 4c). The data revealed that -20% ofthe RT product eluted at 127 mMsodium phosphate, as
expected for single-stranded material, most ofwhich
ap-pearedtobeshort(Fig. 4b, lane 1). The remaining -80% of the product eluted at 187 mM sodium phosphate (Fig. 4b, lane 3), consistent with its being double-stranded, and in-cluded both short and long species. Preliminary analyses
a
kb 23.1
9.4
-6.6
-4.4
-2.2 2.0
-b
F
toM -1
with restrictionenzymesand RNase Hsuggestthatthe latter consistsof both DNA-DNA and RNA-DNAduplexes. The bulk of thefsL-RTbackground specieswere in the form of
double-stranded material (Fig. 4b, lane 4).
Further characterization of the RT reaction. To reassure
ourselves that our assays are measuring bona fide RT
activity,weturned to teststhat have been usedhistorically todefine RTreactions (Fig. 5). For thesetests, weusedan
optimized 30-min RT assay performed in a standard RT
buffer ratherthan themorecomplex HBV polymerase buffer
(see Materials andMethods), and RT-L mRNA rather than
c
_j F- _ _
d
-j... =__ _ _
CC C:
I
0.56
-0.125- F
M 1 2 3 4
RNase A
t 2 3 4
Act
Act. D
1 2 3 4
P+F
PEA
1 2 3 4 5 6
ddATP
FIG. 5. EffectsofRNaseA andinhibitorsofreversetranscription. In vitro RT productionappearstobemostly RNA dependent (a) and
isrepressed by knowninhibitorsof HBV RT(b through d). Lysates ofRT-L- and LS-injectedoocytes(indicated above the lanes)wereused
forthisexperiment.32P-labeledproductsweregenerated byinvitro RTreactionsaftertreatment(+) with 20 ,ug of RNase Aperml (panel
a,lanes 2and4), 100 pLgofactinomycinD(Act.D)perml(panel b, lanes 2 and 4), 1 mM PFA (panelc,lanes 2and 4),or0.83 ,uM ddATP
(panel d,lanes2 and4).Ineachpanel, lanes 1 and 3 show the corresponding products from untreated (-) control reactions. Reaction products
wereanalyzed byagarosegel electrophoresis and visualized byautoradiography. Lanes 5 and 6 in panel d show lighterexposuresof lanes
1 and2. Theend-labeledHindIllfragments of A phageDNAwereusedassize markers (M). Fragment sizesaregiven in kilobase pairs.
a
kb 23.1
4.4
-2.0
-0.56
0.125
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.137.478.431.649.2]4518 SEIFER AND STANDRING
L-RT mRNA provided the source of RT protein, with LS mRNA-injected oocytes as the controls. The 32P-labeled nucleic acid products of the in vitro RT reactions were treated withproteinase K andphenol-chloroformto remove protein, electrophoresed through agarose gels, and then visualized byautoradiography.
The data in Fig. 5 show that the untreated RT-L assay samples(lanes1) always yieldedmore32P-labeled products thandid thecorresponding untreated LS controls (lanes3). Phosphorimaging of the gels revealed that the32P activity in the RT-L lanes ranged from 6.8-fold (Fig. 5a) to 20.8-fold (Fig.Sd) higher than that in the corresponding LS lanes. The RT-L reaction products were more heterogeneous than the LScontrolproductsandincluded largerspecies. The control lane products (a discrete -1.8-kb band and a smear of shorter species) are not dependent on LS mRNA, since essentially the same pattern was seen whether LS mRNA wasomitted or replaced byanother control RNA (data not
shown). Thus, these species presumably represent back-ground products generated in the lysates by an endogenous oocyte enzyme acting on oocyte nucleic acidspecies.
To testwhether the polymerizing activityobserved for the RT-L lysates is dependent on an RNA template, oocyte extracts were treatedfor 10 min at 37°C with 20 Lg of RNase A per ml prior to the RT assay (Fig. Sa). As judged by phosphorimaging, this pretreatment eliminated 83% of the RT-L reaction products (Fig. Sa, lane 2) versus only 8% of the control reactionproducts(lane4).
Actinomycin D(Fig.5b) and PFA (Fig. Sc)weretested by adding them to RT assays to final concentrations of 100 ,ug/ml and 1 mM,respectively. The synthesis of32P-labeled
products directed by theRT-L-expressing lysate was exquis-itelysensitive toinhibition by actinomycin D and PFA(also
known asfoscarnet), which reduced label incorporation by 85 and96%, respectively (Fig. Sb and c, cf. lanes 1 and2).
Thecontrol lane products wereless sensitive to thesedrugs, showing reductions of52% with actinomycin D (Fig. Sb,cf. lanes 3 and 4) and56% with PFA(Fig.Sc,cf. lanes 3 and4). These values are consistent with the known properties of these drugs (4, 17, 33) which preferentially inhibit viral polymerases and RTs but alsoaffect cellularpolymerases at the concentrations used in this study.
The final inhibitor tested, the chain terminator ddATP (Fig. Sd), reduced the overall production of RT-L-derived products by 35% (Fig. Sd, cf. lanes 1 and 2) and caused a 25% reduction in the background laneproducts (cf. lanes 3 and4). These modest inhibition figures reflect the fact that the ddATP concentration used in this experiment(0.83 ,uM)
wasonly about fourfold
higher
than the[32P]dATP
concen-tration. HigherddATP/32
P]dATP ratios elicited a more severe curtailment of [2P]dATP
incorporation (data notshown).An intriguing feature of the data inFig. Sd, lanes 1 and 2, which is more easily seen in a shorter autoradio-graphic exposure (lanes 5 and 6), is that the extent of inhibition due to ddATP varies with the length of the RT-L-derivedproduct, with thelonger products being inhib-ited more severely than the shorter ones. Using the phos-phorimager, we divided each of lanes 1 and 2 of theddATP gelsinto three equal regions, corresponding to three product size classes(<0.1to0.5kb, 0.5to 2.2kb, and 2.2 to >23 kb), and quantitated the32Pactivity present in each region. The levelsofinhibition due to ddATP seen in these size classes were20, 35, and 48%, respectively, whereas the LS controls gave a uniform -25% inhibition for all three size classes. The fact that the longer products of the RT assay are more sensitive to chain termination with ddATP suggests that
there is significant chain elongation occurringin this in vitro reaction(22).
Insummary, these experiments show that RT-L-contain-ing oocyte extracts exhibit an RNA-dependent deoxynucle-otide-polymerizing activity that (i) is distinct from an endo-genouspolymerase detected in control oocyte lysates and (ii) behaves as expected in response to drugsthat have
classi-callybeen usedtodefine RT reactions. Takentogether with thefindings presented in the preceding sections, the results shown inFig. 5makeacompelling case for the fact thatwe aredealingwith thetrueHBVRT. The only puzzling feature of thedata in Fig. 5 is an apparent discrepancy between the RNase A and actinomycin D results. The sensitivity of the RTreaction to RNase Aseemingly implies that the HBV RT is copying an RNA template, and, hence generating a first-strand cDNA product. In contrast, the actinomycin D data imply that the main RT reaction product is double-stranded DNA, since this drug preferentially reduces sec-ond- rather than first-strand DNA synthesis (33). However, these results can perhaps be reconciled if the in vitro RT reaction starts with an initial phase of single-strand synthesis followed by a second phase of double-strand synthesis. Further experiments will berequired to address this issue.
DISCUSSION
We have expressedin frog oocytes animmunoprecipitable form ofhuman HBV RTin the absence of the viral nucleo-capsidprotein. Unexpectedly, oocytelysatescontainingthis soluble RTexhibited adeoxynucleotide polymerizing activ-itythat (i) has the propertiesexpected for a trueRTand(ii) appears toremain covalently linked to the reverse transcrip-tionproducts, acharacteristic and unusual feature of hepad-naviral polymerases. This resultimmediately arguesagainst
thecapsid proteinbeing a prerequisite forRTactivity,even thoughgeneticevidence indicates that thecarboxyl-terminal subdomain ofthe viral capsid protein is required for DNA genomematuration inboth theduck(24, 36) and human(18)
viruses. We speculate that the involvement of the capsid protein in the replication process maybe a helper function enabling HBV RT to overcome the formidable problems of topology and nucleic acid condensation that must arise during the conversion ofsingle-stranded RNA into double-stranded DNAwithin the strict confines of the capsid. We have previously proposed that this help may come in the form of binding energy supplied by specific DNA-binding
motifs presentin the capsid protein carboxyl terminus(11).
Our results are consistent with a recent report (35) that duck HBV RT protein produced by in vitro translation is enzymatically active in the absence of the viral
nucleocap-sid. However, ourstudyprovidesthe first demonstration(i)
that this result holds true for the medicallymoreimportant
human enzyme, (ii) that such activitycanbe detected in an animal cell system, and (iii) that the activity can be unex-pectedly uncoupled from regulatory elements such as the encapsidation signal, s, and, most notably, the replication origin, DR1(25, 27). Thereare somedifferences between the results seen with duck and human HBVRTs. Curiously, the duck RTactivity was limitedlargelytothe covalent addition ofjust fourdeoxynucleotides to the RTprotein, while only
trace amounts of longer productswere seen. Although our results also suggest, albeit indirectly, that the HBV RT protein remains attached to the reverse transcription
prod-ucts and thus has presumably primed DNA synthesis, the oocyte-derived enzyme differs in its polymerizing
capabili-ties in that ityields large amounts of products thatarelonger
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
than 100 nucleotides (some may be as long as 23 kb). We havenotshown that theseproductsarise exclusivelyinour in vitro assay (somemaywell have been started inside the intactoocyte),butthe effect of ddATPonthereaction(Fig.
Sd) argues strongly that our in vitro reaction can support chain elongation. Whether these disparate results reflect differences between the two systems or between the two enzymes and/or their templates orwhethertheyarecaused
byunknown factors remains tobedetermined.
Among the many crucial questions that remain to be answered are those about the nature of the presumptive
RNAreversetranscription template(s) copiedinoursystem and the
identity
oftheprimingsite(s)
used in the absence of DR1. We donotyet knowwhether the reaction templateis the input HBV RT mRNA or an unidentified endogenousoocyte RNA. The fact that some of the polymerized RT
productsare apparently longer thantheinjected RTmRNA could be taken to imply the use of a long oocyte mRNA
template (or templates), butwecertainlydonotruleoutthe
possibility that RT mRNA acts as its own substrate. To
explain overlength products arising byreversetranscription
of RTmRNA,we caninvoke scenarios suchasthe reiterated
copying
of the RNA(via
snapback priming)orthejumpingof thereversetranscription machinery
toadditional templates.These issues arecurrentlyunderinvestigation.
Aswe notedearlier, there is convincing evidence (3, 26)
that HBV RT is translatedas a discrete entityandnot as a core-RTprecursor.Theconstitutive activityofthis discrete RT forces us to reconsider the proposed HBV replication
scheme. Itnowbecomesconceivable thatreverse
transcrip-tion of the HBV pregenome RNA initiates prior to capsid assemblyand thatapartially replicatedgenomecould be the true substrate forselective inclusion into thecapsid. More-over, the notion that HBV
replication
mayinitiate outside the capsid challenges the widely held perception that theproduction of active viral RTs, and hence, the reverse
transcription process, is restricted, or segregated, to the interior of viral
nucleocapsids (6).
Retroviruses anddistantlyrelated viruses such as yeast L-A double-stranded RNA virus
(7) accomplish
suchsegregation
via a common mech-anism.Briefly,
these viruses elaborate inactivegag-pol RT precursors(14)
whichareactivated(by
proteolytic cleavage)only
oncethey
have bound their pregenome RNAs and haveincorporated
into the viralnucleocapsid. Wespeculate that unless HBV uses anovel kind of segregation strategy, the active HBV RT may come into contact with and reverse transcribe other viral andevencellular mRNAsduringHBV infection. That HBV RT is not completely faithful to its pregenome templateis suggested by therecent demonstra-tion thatsubgenomic
viral RNAs are reverse transcribedduring
viral infection(10, 34).
The reverse transcriptionofcytoplasmic
RNAscould haveimplications
for thedevelop-mentofHBV-linkedhepatocellular carcinoma(23).
The demonstration ofafree humanRTthat is bothhighly
active and
immunoprecipitable
opens upnewavenuesin thestudy
of HBVbiology
andpathology.
Our system should proveuseful for characterizingthe RTmolecule anddeter-mining the identity of its active form, for examining its
primaseandRNase Hsubactivities,and forelucidatinghow itinteracts with RNA. In addition, it should be feasible to extend our
prior
oocyte studies on HBV capsid assembly (28, 37, 38)tothekeyissue ofhow theRTprotein,thecapsid protein, andthe pregenomeRNAcommunicateto promote theassembly
ofareplication-competent
nucleocapsid (25).Finally,
the robustactivity
of the human HBV RT seen inthesestudies makes the oocyte systemattractive for testing novel RT inhibitors for antiviral therapy.
ACKNOWLEDGMENTS
We are indebtedtoM. Blanar, I. Caras, J. Glenn, R. Ward, and the referees for suggestions concerning the manuscript; to W. Gerlich (Giessen, Germany) and K. H. Heermann (Gottingen, Germany) for supplyingMA18/7; andto0.Veneki for oligonucle-otidesynthesis.We thankJ. Reardon,L. Condreay,and M. Davis for valuable discussions.
This work wassupported by NIH grantA125056and in partby a Pilot/Feasibility Projectgrantfrom the LiverCenterof the Univer-sityof CaliforniaatSan Francisco(DK-26743) andbyagrantfrom theMSC/REAC Hampton Fund (UCSF).M.S.wassupported bya
fellowship fromtheDeutscheForschungsgemeinschaft. REFERENCES
1. Bartenschlager,R., M. Junker-Niepmann, and H. Schaller. 1990. The P gene product of hepatitis B virus is required as a
structural componentforgenomicRNAencapsidation.J.Virol. 64:5324-5332.
2. Bowerman, B., P.0.Brown, J. M.Bishop, and H. E. Varmus. 1989. A nucleoprotein complex mediates the integration of retroviral DNA. Genes Dev. 3:469-478.
3. Chang,L.J., P. Pryciak,D.Ganem,and H. E. Varmus.1989. Biosynthesisof thereversetranscriptaseofhepatitisB viruses involves de novotranslational initiation notribosomal frame-shifting.Nature(London) 337:364-368.
4. Crumpacker, C. S. 1992. Mechanism of action of foscarnet againstviralpolymerases. Am. J. Med.92(Suppl. 2A):3S-7S. 5. Farnet, C. M., and W. A. Haseltine. 1991. Determination of viral
proteins present in the human immunodeficiencyvirus type 1 preintegration complex.J. Virol. 65:1910-1915.
6. Fuetterer, J., and T. Hohn. 1987. Involvement ofnucleocapsids in reverse transcription: a general phenomenon? Trends Bio-chem. Sci. 12:92-95.
7. Fujimura, T., J. C. Ribas, A. M. Makhov, and R. B. Wickner. 1992.Polofgag-polfusionprotein required forencapsidation of viral RNA of yeast L-A virus. Nature(London) 359:746-749. 8. Geck,P., andI.Nasz. 1983.Concentrated,digestibleDNAafter
hydroxylapatite chromatographywithcetylpyridinium bromide precipitation.Anal. Biochem. 135:264-268.
9. Gerlich, W., and W. S. Robinson. 1980. Hepatitis B virus contains aprotein attached tothe 5' terminus ofitscomplete DNAstrand.Cell21:801-809.
10. Hantz,O., I.Baginski, I. Fourel, I.Chemin,andC.Trepo.1992. ViralsplicedRNAareproduced, encapsidatedandtranscribed duringin vivo woodchuckhepatitisinfection.Virology 190:193-200.
11. Hatton, T., S. Zhou, and D. N. Standring. 1992. RNA- and DNA-binding activities in hepatitis B virus capsid protein: a
model for their roles in viralreplication.J. Virol. 66:5232-5241. 12. Heermann, K.H.,U.Goldmann,W.Schwartz,T.Seyffarth,H. Baumgarten,and W. H.Gerlich.1984.Largesurfaceproteinsof hepatitisBviruscontainingthe pre-s sequence. J.Virol. 52:396-402.
13. Hirsch,R.C., J.E.Lavine,L.J.Chang,H. E.Varmus,and D. Ganem.1990. Polymerasegeneproducts of hepatitisBviruses
arerequiredforgenomicRNApackagingaswellasforreverse
transcription. Nature(London)344:552-555.
14. Jacks, T., H. D.Madhani, F. R. Masiarz, and H. E. Varmus. 1988. Signalsfor ribosomalframeshiftinginthe Roussarcoma
virusgag-polregion.Cell55:447-458.
15. Junker-Niepmann, M.,R.Bartenschlager,andH. Schaller.1990.
Ashort cis-acting sequence is required for hepatitis B virus pregenomeencapsidationandsufficient forpackagingofforeign RNA.EMBO J. 9:3389-3396.
16. Krieg, P. A., and D. A. Melton. 1984. Functional messenger RNAs are produced by SP6 in vitro transcription of cloned cDNAs. Nucleic Acids Res. 12:7057-7070.
17. Mason,W.S., J. Lien,D.G.Petcu,L.Coates,W. T.London,A.
O'Connell,C.Aldrich, andR. P.Custer. 1987. In vivo and in
on November 9, 2019 by guest
http://jvi.asm.org/
4520 SEIFER AND STANDRING
vitro studies on duck hepatitis B virus replication. UCLA Symp. Mol.Biol. 70:3-16.
18. Nassal, M. 1992. The arginine-rich domain of the hepatitis B viruscoreprotein is requiredfor pregenomeencapsidation and productive viralpositive-strand DNA synthesisbutnotfor virus assembly.J.Virol. 66:4107-4116.
19. Prickett, K. S., D. C. Amberg, and T. P. Hopp. 1989. A calcium-dependent antibody for identification and purification of recombinantproteins. BioTechniques 7:580-589.
20. Radziwill, G., W. Tucker, and H. Schaller. 1990. Mutational analysis of the hepatitis B virus P gene product: domain structureand RNase Hactivity. J.Virol.64:613-620.
21. Radziwill, G., H. Zentgraf, H. Schaller, and V. Bosch. 1988. The duckhepatitis B virus DNApolymerase is tightly associated with the viral core structure andunabletoswitchto an exoge-noustemplate.Virology 163:123-132.
22. Reardon, J. E. 1992. Human immunodeficiencyvirus reverse transcriptase: steady-state and pre-steady-state kinetics of nu-cleotide incorporation. Biochemistry31:4473-4479.
23. Rogler, C. E. 1991. Cellular and molecular mechanisms of hepatocarcinogenesis associated with hepadnavirus infection. Curr.Top.Microbiol. Immunol. 168:103-140.
24. Schlicht, H.-J., R. Bartenschlager, and H. Schaller. 1989. The duckhepatitis Bviruscoreprotein containsahighly phospho-rylatedC terminus that is essentialforreplication but not for RNApackaging. J.Virol. 63:2995-3000.
25. Schlicht, H. J., R. Bartenschlager,and H. Schaller. 1991. Bio-synthesisandenzymatic functions of thehepadnaviral reverse transcriptase, p. 171-180. In A. McLachlan (ed.), Molecular biologyofthe hepatitisB virus.CRCPress, Boca Raton, Fla. 26. Schlicht, H. J., G. Radziwill, and H. Schaller. 1989. Synthesis
and encapsidation of duck hepatitis B virus reverse tran-scriptase donot require formation of core-polymerase fusion proteins. Cell56:85-92.
27. Seeger, C., J. Summers, and W. S. Mason. 1991. Viral DNA synthesis. Curr. Top. Microbiol. Immunol. 168:41-59. 28. Seifer, M., S. Zhou, and D. N. Standring. 1993. Amicromolar
poolofantigenically distinctprecursors isrequired toinitiate
cooperative assembly of hepatitis Bvirus capsids in Xenopus oocytes. J.Virol.67:249-257.
29. Sominskaya, I., P. Pushko, D. Dreilina, T. Kozlovskaya, and P. Pumpen. 1992. Determination of the minimal length ofpreSl epitope recognized by a monoclonal antibody which inhibits attachmentofhepatitisBvirustohepatocytes. Med. Microbiol. Immunol. 181:215-226.
30. Standring, D. N., J. H.Ou,F. R.Masiarz, and W. J. Rutter. 1988. A signal peptide encoded within the precore region of hepatitis B virus directs the secretion of a heterogeneous population of eantigensin Xenopus oocytes. Proc.Natl.Acad. Sci.USA 85:8405-8409.
31. Standring, D. N., J. H.Ou, and W. J. Rutter. 1986.Assembly of viral particlesinXenopusoocytes:pre-surface antigens regulate secretion of thehepatitis B viral surface envelope particle. Proc. Natl. Acad. Sci.USA 83:9338-9342.
32. Standring, D. N., W. J. Rutter, H. E. Varmus, and D. Ganem. 1984.Transcription of the hepatitis Bsurface antigengenein cultured murinecells initiates within thepresurface region. J. Virol. 50:563-571.
33. Summers, J.,and W.S.Mason.1982.Replication of thegenome ofhepatitis B-like virus by reverse transcription of an RNA intermediate. Cell 29:403-415.
34. Terre, S.,M.-A.Petit,andC.Brechot.1991. Defectivehepatitis Bvirusparticlesaregenerated bypackaging andreverse tran-scription of spliced viralRNAsinvivo.J.Virol. 65:5539-5543. 35. Wang,G.H., and C.Seeger.1992. Thereversetranscriptase of hepatitis B virus acts as a protein primer for viral DNA synthesis. Cell71:663-670.
36. Yu, M., and J. Summers. 1991. Adomain of thehepadnavirus capsid proteinisspecificallyrequired forDNAmaturation and virusassembly. J.Virol. 65:2511-2517.
37. Zhou,S., and D. N.Standring. 1992. HepatitisBvirus capsids
areassembled fromcoreprotein dimers. Proc. Natl. Acad. Sci. USA 89:10046-10050.
38. Zhou,S.,S.Q. Yang,and D. N.Standring. 1992. Characteriza-tion ofhepatitis B viruscapsid particle assembly in Xenopus oocytes.J. Virol.66:3086-3092.
J.VIROL.