0022-538X/94/$04.00+0
Copyright © 1994, American Society for Microbiology
Phenotypic Mixing between Different Hepadnavirus Nucleocapsid
Proteins
Reveals C
Protein
Dimerization
To Be cis
Preferential
CHRISTOPHER CHANG,1'3SILIANG ZHOU,2 DON GANEM,3* ANDDAVID N. STANDRINGl.2* Departmentof Biochemistry and Biophysics, 1Hormone Research Institute,2 and Departments of Microbiology and
Medicineand Howard HughesMedical Institute,3 UniversityofCalifomia MedicalCenter, San Francisco, Califomia 94143
Received 24March 1994/Accepted 10 May 1994
Hepadnaviruses encode a single core (C) protein which assembles into a nucleocapsid containing the polymerase (P) proteinandpregenomic RNA during viralreplicationinhepatocytes.Weexamined theability ofheterologous hepadnavirus Cproteins tocross-oligomerize. Usinga two-hybrid assay inHepG2 cells,we observedcross-oligomerization amongthe core proteins from hepatitis B virus (HBV),woodchuckhepatitis virus, and ground squirrel hepatitis virus. When expressed in Xenopus oocytes, in which hepadnavirus C proteins form capsids, the C polypeptides from woodchuck hepatitis virus and ground squirrel hepatitis virus, but not duckhepatitisBvirus, canefficiently coassemblewithanepitope-taggedHBV corepolypeptidetoform mixedcapsids. However, whentwodifferent coremRNAsarecoexpressedinoocytesthe core monomers show astrongpreference forforming homodimers ratherthan heterodimers.Thisholds true even forcoexpression oftwoHBVC proteinsdiffering only by an epitope tag, suggesting that core monomersare notfreetodiffuse and associate with other monomers. Thus, mixed capsids result from aggregation of different species of homodimers.
The core (C) protein of hepatitis B virus (HBV) plays a major role in viral morphogenesis. Core polypeptides, the polymerase (P), and thepregenomic RNAassemble to form the nucleocapsid during viral replication in hepatocytes. The nucleocapsid, in turn, interacts with internal cellular mem-branes that contain HBV envelope glycoproteins to produce enveloped virions. Several laboratories have demonstrated that core protein expressed in the absence of all other viral products can spontaneously assemble into capsid particles in hosts as divergent asbacteria (3, 6), the yeast Saccharomyces cerevisiae (13), insect cells (2), or Xenopus oocytes (21). Nevertheless, the pathway forcapsid assembly remains incom-pletely understood. Zhou and Standring (20, 21) have shown that in the Xenopus oocyte system, unassembled coreprotein exists in a dimer pool that serves as a precursor to capsids. Electron microscopy and velocity sedimentation studies have failed to detect in vivo higher-order multimer intermediates oftenseenin the assembly of other animal viruscapsids(20). Efforts to identify regions of the 185-amino-acid (aa) C protein that are critical for capsid formation have focused primarily on thearginine-richC-terminal tail (2, 3). In various heterologous systems expressing core protein, deletions con-tainingthe N-terminal 144 aaproduce stablecapsids, demon-strating that the Arg-rich tail is not necessary for capsid assembly (2, 3). Moreover, assembly is tolerant of heterologous sequences placed at the N terminus, at the C terminus, and within an immunodominant loop of core protein (4, 5, 16). However, C-terminal truncations resulting in C proteins
shorter than 144residuesabrogatecore particle assembly. To furthercharacterize C-Cinteractions involvedin capsid assembly, we examined the ability of mammalian and avian hepadnavirus C proteins tointeract with each other.Weused both expression in Xenopus oocytes and a mammalian cell-based two-hybrid system. The two-hybrid strategy originally developed by Fields and Song (9) to detect protein-protein
*Correspondingauthors.
interactions relies on themodular nature of the yeast GAL4 DNA-binding and transactivation domains. Fearon and col-leagues modified this systemto study interactions in mamma-lian cellsby using the herpes simplexvirusVP16 transactiva-tion domain (8). Luban et al. have applied this strategy to detect multimerization of Gagpolyprotein from both human immunodeficiencyvirus(HIV)andMoloney murine leukemia virus (MuLV) (11).
In this communication, we report that HBV core proteins (HBV-C) fused to GAL4 and VP16 domains interact in the two-hybridassayandthat thisinteraction behavesaspredicted frominvivostudies ofcapsid formation. Using this two-hybrid assay, we demonstrate cross-oligomerization among the core
proteins from HBV, woodchuck hepatitis virus (WHV), and ground squirrel hepatitis virus (GSHV). Consistent with this finding, when expressed in Xenopus oocytes, the C proteins from WHV and GSHV, but not duck hepatitis B virus (DHBV), efficientlycoassemble with HBV Cproteintoform mixed capsids. Surprisingly, however, when two different C protein mRNAs are translated in the oocyte, the C polypep-tides show a marked preference for forming homodimers rather than heterodimers; thus, mixed capsidsare aggregates of differentspecies of homodimers.
MATERLALSANDMETHODS
Materials. Restriction enzymes werepurchased from New England Biolabs and used as specified by the manufacturer. Radionucleotides were purchased from Amersham Corp. or NewEngland Nuclear. Monoclonal antibody(MAb) 18/7, the MAb directed against the L epitope, was kindlyprovided by Volker Bruss and Wolfram Gerlich. The rabbit polyclonal anti-HBV-C-antibody was purchased from Dako Corp. Anti-DHBV-Cwaskindly provided by Jesse Summers.
Plasmids and RNAs. All HBV nucleotide
positions
arenumbered fromtheuniqueEcoRI site ofHBVsubtypeadw2. Core fusion proteins with GAL4 or VP16 were made
by
subcloning appropriate restriction orPCR-amplified
frag-5225
on November 9, 2019 by guest
http://jvi.asm.org/
ments into the
expression
vectorspGALO
and pNLVP16,which, along
with the reporterplasmid
pGSElbCAT, weredescribed earlier
(8)
andkindly
provided by Eric Fearon. Inbrief,
pGALO
encodes the DNA-binding domain of GAL4,extending
from aa 1 to147,
followedby polylinker
sequence.pNLVP16containsaa411 to455 of the HSV VP16 transacti-vation domain followed
by polylinker
sequence. Full-length HBV,WHV,andGSHVcorefusionsweremadeby subcloninga
581-bp StyI fragment
into thepolylinker
ofpGALO
andpNLVP16
tocreate anin-framefusionwith the C terminus oftheGAL4orVP16domain. C-terminal truncations HBc1-144 and HBc1-117 were
engineered by deleting
sequence fromBspEI
andSspI,
respectively,
tothepolylinker;
HBc29-168wasconstructed
by inserting
a445-bp
BglII fragment
into thepGAL4
polylinker;
HBc38-183wasconstructedby inserting
a449-bp TaqI-StyI fragment
into thepNLVP16polylinker.
To create SP6
expression
vectors, the fragments spanningthe core open
reading
frames ofWHV, GSHV,
and DHBVwere
generated by
PCRamplification
soastoincludeanNcoIsite at the 5' end and a BstEII site at the 3' end. These
fragments
were cloned into the NcoI and BstEII sites of aderivative of
pSP64T.
This strategy puts the core sequences between thetranslation-enhancing
5' and 3' untranslatedregion
sequences of the,-globin
gene. PlasmidspSP64T-C
and A157 are described in detail elsewhere
(19). Briefly,
pSP64T-C
contains the581-bp StyI fragment
fromHBVadw2(nucleotides
1880to2461)
cloned into theBglII
site ofpSP64T.A157 encodesa coredeletioncontainingthe first 157aafused inframetothe lasttworesidues ofcore
protein.
TheLepitope
tag has beenpreviously
described(14b).
We modified theoriginal
Lepitope
sequence with a silent basechange
thatwould introduce aBamHI site: 5'GATCATCAGTTGGATC CTGCATTCGGAGCCAACTCA3'.TheL
epitope
inA157L and HBV-CL is borderedby
coreresidues 78 and80,
deleting
residue79,
in the immunodominantloop
of core. It wasintroduced into this site
by ligation
at the BamHI site of thetwo
PCR-amplified
halves of the core openreading
frameflanking
the Lepitope.
SP6expression
constructs wereverifiedby
sequenceanalysis.
In vitro
transcription.
Synthetic transcripts
were preparedby using
SP6polymerase
(Promega).
Transcription
reactionmixturesweretreated with
DNase,
phenol
extracted,
ethanolprecipitated, resuspended
intoH20,
and storedat-70°C
untilrequired
formicroinjection.
Transfections.
HepG2
cellsweregrownon100-mm-diame-terdishes in DMEH-16 medium
supplemented
with10%fetal bovine serum(Gibco),
0.14% sodiumbicarbonate,
and 2 mMglutamine. HepG2
cellsweretransfected with 5 ,ugof each ofthe GAL4 andVP16 fusionconstructsand the
chloramphen-icol
acetyltransferase
(CAT)
reporterplasmid by
thecalciumphosphate coprecipitation procedure
as describedpreviously
(10).
CAT
activity
assays. Cell extracts for CATactivity
wereprepared by
freeze-thawlysis
of cells 48 h posttransfection.CAT
activity
wasdetermined inextractsby
aphase
separationprocedure.
Inbrief,
5,ul
of cell extract was incubated in a100-pd
volumecontaining
250mMTris-HCl(pH 8.0),
0.5 mMn-butyryl
coenzyme A(Sigma),
and 0.2 ,uCiof["4C]chloram-phenicol (58.1 mCi/mmol)
for 15 to 60 min at 37°C. Phaseextractionwas
performed
withanequal
volume of 2:1hexane-xylene mix,
and levelsofn-butyrylated chloramphenicol
weredetermined
by liquid
scintillationcounting.
CATactivity
isreported
as fold increase relative to CAT activity resultingfromtransfection of the reporter CATplasmid alone.
Oocyte
handlingandmicroinjection. Oocyteswereculturedat
17°C
in modified Barth's mediumcontaining
antibiotics(MBSH) by standard procedures (21). Batches of 10 to 20 oocytes per experiment were injected as describedpreviously (18) by using a programmable microinjection device (Sutter Instruments, Novato, Calif.) to control delivery of RNA (40 nl peroocyte). For experiments involving analysis of assembled capsids, metabolic labeling was initiated within 1 h ofinjection byplacing the oocytes in MBSH containing 0.5 to 1.0 mCi of [5S]methionineand[35S]cysteine(Expresslabel; NEN) per ml and incubating them for a further 35 to 40 h at 17°C. For analysis of the intermediates in capsid assembly, the oocytes were labeled 24 h after injection for approximately 20 h. Lysates were prepared by homogenizing the oocytes in buffer ('20 ,u per oocyte) containing 50 mM Tris-HCl (pH 7.5),1% Nonidet P-40, 100mMNaCl, and 10 mM EDTA. Lysates were then clarified (Eppendorf centrifuge, 15,000 x g, 10 min) before fractionation.
Sucrosegradient fractionation.Oocytelysateswerelayered ontoeither 10 to60% (wt/vol)or3 to 25% sucrose gradients (20, 21).For 10to60%gradients, 200 ,u of lysate was layered ontostepgradients (1.4ml, final volume) with six 200-,ul steps of10, 20, 30, 40, 50, and 60%sucrosein 50 mMTris-HCl (pH 7.5)-100 mMNaCl-1 mM EDTA(TNE) and centrifuged for 40 min at 4°C and55,000 rpm in a TLS-55 swinging-bucket rotor on a TL-100 ultracentrifuge. Gradients of 3 to 25% sucrosewereformedby layering 200-,u steps of 3, 5, 10, 15, 20, and25%sucrose inTNE, immediatelyloadedwith 200 pI of lysate,and spunat55,000 rpm for 4 hasdescribed above.For bothsucrosegradients,14100-p1fractionswerecollected from the top of the gradientand analyzed byimmunoprecipitation and sodiumdodecylsulfate (SDS)-polyacrylamidegel electro-phoresis (PAGE).
Immunoprecipitations and SDS-PAGE. 35S-labeled core proteinswere immunoprecipitated from "30 pA of each gra-dient fraction under native conditions, i.e., without SDS or boiling, with eitherapolyclonal anticore antibody (Dako) or the anti-L MAb 18/7 (15).Immunecomplexeswerecollected with Zysorbin (Zymed); afterwashing, the pellet was resus-pended in sample buffer, and core protein was released by boiling for 10min and analyzedon13.5%polyacrylamidegels.
RESULTS
HBV core protein oligomerization. In order to use the
two-hybrid system to characterize heterologous C-C protein interactions in replication-permissive HepG2 cells, we first demonstrated that we could detect homo-oligomerization of full-length HBV C polypeptides in this system and that the system faithfully reproduces the known aspects of C-C oli-gomerization.
HepG2 cells were cotransfected with plasmids expressing HBV C protein fused to the GAL4 DNA-binding domain
(GAL-core)
andto the VP16 transactivation domain(VP16-core), in the presence of a CAT reporter gene bearing five GAL4DNA-bindingsites. If C-Coligomerizationoccurs,then the VP16 activation domain will be targeted to the DNA region 5' ofCAT and will activate CATexpression. Table 1 summarizes the results of CAT assays performed following these cotransfections. When the GAL4-core fusionwas coex-pressed with the VP16 activation domain alone, no increase in CATactivityoccurred.Similarly,theVP16-corefusion didnot associate with theGAL4DNAbindingdomaintoboost CAT expression. These controls indicate that HBV core doesnot associate with either the GAL4orVP16domain; furthermore,
corebyitself hasnotransactivating potential,nor canit bindto andactivate thetestpromoter whenit is fusedtoVP16. Asa
positive control,we showed that the a and , subunits of the
on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 1. Full-lengthHBV coreproteinoligomerizesa
GAL4 fusion VP16 fusion Relative CATactivity
HBc None 1
None HBc 1
CapZ 1B25 CapZ as49 40
CapZ 1325 HBc 1.5
HBc HBc 40
a HepG2 cells were transfected with three plasmids: a fusion construct
encodingtheGAL4DNA-binding domainand a downstreamproteindomain of interest; a second fusionconstructencoding the VP16transactivation domain and a downstreamproteindomain ofinterest;and a CATreportergene withfive
upstream GAL4 DNA-binding sites. The test protein domains fused to the GAL4 or VP16 domains are indicated. "None" indicates that the fusion
constructencodes either the GAL4 or VP16 domainalone,without a down-stream domain. CATexpressionlevels arereportedasfold increase relative to CATactivity resultingfrom transfection of the CAT reporter plasmid alone.
HBc,HBV coreantigen; CapZ, actin-capping protein.
actin-capping protein CapZ, which are known to form anot-1 dimer, associated to increase CAT activity 40-fold above background levels, as shownpreviously (8). When GAL4-core and VP16-coreclones werecoexpressed, CAT geneexpression likewise increased 40-fold. This result demonstrates that HBV core polypeptides can associate efficiently in the two-hybrid assay,thus enabling VP16 to transactivate the CAT reporter. The C-C interaction was specific, asshownbya lackof CAT activation whenthe core fusion was coexpressed with the CapZ fusionprotein.
Withthis assay,we then evaluated the effect of truncations atthecarboxy terminus of C on itsability to associate with the full-lengthcore. Assummarized in Table2, GAL4 fused to the first 144 aaofcore and without its 41-aaarginine-rich tailwas nonetheless able to complex with the full-length VP16-core fusion as efficiently as the full-length GAL4-core. However, whenweengineereda68-aadeletion from the C terminus, this truncatedcore fusion failedto interact withfull-length VP16-core fusionto activate CATexpression. This behavior closely parallels the known behavior of similar deletions in C when assayed by capsid formation in bacteria or cultured hepato-cytes (3, 14).
Unlike the case for C-terminal deletions, the effect of N-terminal deletions on C-C association has not been re-ported. We engineered a pair of deletions and tested their interactions withfull-lengthcore.AGAL4-corefusionlacking thefirst 28 residues(andtheC-terminal 15residues) failedto
oligomerize with core,as dida VP16-corefusion missing the first 37 aafromcore. However, these data donotprove that these N-terminalresiduesparticipateincore-coreinteractions (either directly or indirectly); we cannot exclude that their absencemight have merelydisrupted the foldingorstability of thefusionproteinsusedin this assay.
TABLE 2. C-terminal and N-terminal deletionsin HBV
coreproteina
GALAfusion VP16 fusion RelativeCAT
activity
HBc HBc 22
HBc1-144 HBc 23
HBc1-117 HBc 1
HBc29-168 HBc 1
HBc HBc38-183 1
[image:3.612.316.559.87.196.2]aSee thefootnote to Table1for details.
TABLE 3. Interactionsamongmammalian HBVcoreproteinsa
GALA fusion VP16 fusion
Relative
CATHBc HBc 23
WHc WHc 18
GSHc GSHc 1
WHc HBc 72
HBc WHc 5
GSHc HBc 20
HBc GSHc 1
GSHc WHc 5
WHc GSHc 1
aSee the footnotetoTable1for details.HBc,WHc,and GSHcdesignatecore
antigensfromHBV,WHC,andGSHV,respectively.
Interactions among mammalian HBV core proteins. We nextusedthis systemtodetermine whethercoreproteinsfrom the different mammalian
hepadnaviruses
could hetero-oli-gomerize.Themammalian Cproteins
displaysignificant
amino acid sequence conservation. HBV-C and WHV-C differ in 46 residues; WHV-C and GSHV-C have diverged only slightly and differ in just 11 residues(17).
Failure of the different mammalian C proteins to associate would imply that amino acids critical forcore-coreinteraction havenotbeenconserved amongtheviruses,whereas hetero-oligomerization would in-dicate that the nonconserved residuesare notessential for this process.Weconstructed both GAL4 and VP16 fusionproteinswith WHV or GSHV C
proteins.
The results of thetwo-hybrid
assaysareshown inTable 3. Like theHBVfusions,
the WHVconstructshomo-oligomerizedandactivated CAT gene expres-sion.However,the assayfailedtodetect
homo-oligomerization
between the GSHV-C fusions. Interestingly, WHV-C fusions could hetero-oligomerize with their HBV counterparts. For the GSHV-C constructs,onlytheGAL4-GSHV-C fusionwasable to
hetero-oligomerize
with both its HBV-C andWHV-Cpartners and activate CAT
expression.
This observation strongly argues that the reason for the failure to detect GSHV-Chomo-oligomerization
inthis assayis that the VP16-GSHVcorefusion is either unstableorimproperly
folded. All attempts to detect DHBV-Chomo-oligomerization
failed,
again mostlikely becauseofimproper
folding
orinstability
of the fusionproteins.
Assembly
of anepitope-taggedHBV coreproteininXenopusoocytes. Our results with the two-hybrid assay indicate that hetero-oligomers can form between C proteins of different
species
but do not establish that such mixedoligomers
canproceed to capsid formation. To do
this,
wecoexpressed
different coreproteins
in Xenopus oocytes andassayed
for mixedcapsids
by sucrosegradient
purification
and immuno-precipitation. For this approach to showmixed capsids,anti-serathat
selectively recognize
atleastoneof the individual Cproteins
must be available. Inpreliminary
experiments,
how-ever, we found that WHV-C and GSHV-C are
efficiently
recognized by thepolyclonalanti-HBVcoreantibody
(Fig.
1, lanes4 to6). Therefore,weconstructeda vectorexpressing
anepitope-tagged,
truncated HBV-CtoserveastheHBVpartnerin
coexpression
studies; thisprotein
can bedistinguished by
both its size and its
reactivity
with antibodiestothe tag.A157 isatruncated HBV-Ccomprisingthefirst 157 residues fusedtothe last 2 residues of HBV-C and assembles into
capsids
asefficiently
as thefull-length
core. The 12-aa Lepitope
tag isfrom the pre-Sl
region
of theHBVlarge
surfaceantigen
andwas inserted into
full-length
HBV-C(to
generateHBV-CL)
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.56.298.638.712.2]aL aHBV-C
/' . / \ /
43-
29-18.4
-A 1 2 3 4 5 6 7 8 9 1011 121314
43
-
29-184
-1463
q-III
6.2
-...
U. 0
14.3-1 2 3 4 5 6
FIG. 1. Analysis ofcross-reactivitywith the anti-L MAb(oaL) and the anti-HBV-C polyclonal antibody (otHBV-C). Lysates from
35S-labeled Xenopus oocytesinjected with the appropriate mRNA were resolvedon stepgradientsof 10 to60%sucrose.Fractionscontaining assembled capsids were combined and immunoprecipitated under nondenaturing conditions with the appropriate antibodyandanalyzed by SDS-PAGE on a13.5%gel.Thepositionsofprestainedmolecular massmarkers areindicated in kilodaltonsatthe left.
and into A157(generating A157L)between residues 78 and80, in a predicted surface loop of the core protein that has previously been shown to be tolerant of insertions (4). To confirm that L epitope-tagged HBV C proteins could be specifically recognized by an anti-L MAb, we expressed HBV-CL(and,ascontrols, the wild-type C proteins of GSHV andWHV)in oocytesand examined theirreactivities with the anti-L MAb 18/7 by immunoprecipitation. As shown inFig. 1 (lanes 1 to 3), only HBV-CL was precipitable with this antibody.
To confirm that A157L can assemble into capsids that containexposedLepitopesontheirsurface,weexpressedthis construct in Xenopus oocytes and fractionated the oocyte extracts on astepgradientof10 to 60%sucrose; 14fractions werecollected from thegradientandanalyzedby immunopre-cipitation followed by SDS-PAGE, autoradiography, and Phosporlmager quantitation. Similar experiments were also performed with oocytes programmedtoexpress HBV-CL and DHBV-C. The gradient profile of A157L (Fig. 2B) clearly showsacapsid peak (fractions7 to 12) that closely mirrors the profile produced by HBV-CL and DHBV-C (Fig. 2 and references 20 and 21), indicating that A157L indeed retains assembly competence. Immunoprecipitation under native con-ditions with anti-L verified that theepitopewasexposedonthe particle surface;coreparticles lackingthe tagwerenot
precip-itated under these conditions(datanot shown). Similar gradi-entprofilesofWHV-C and GSHV-C(datanotshown) confirm that they also assemble efficiently into capsids in Xenopus oocytes.
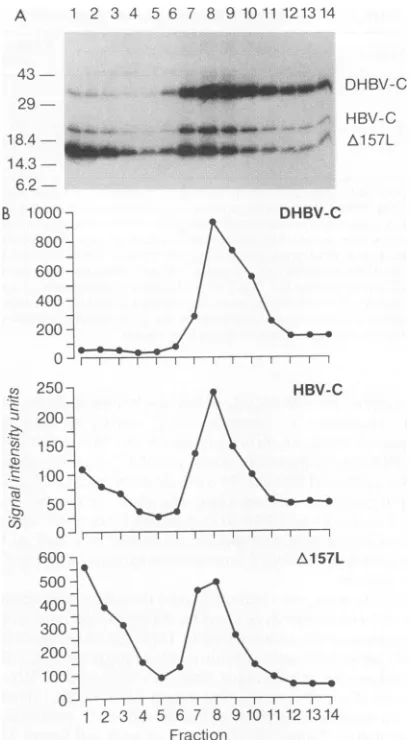
Mammalian hepadnavirus cores form mixed capsids. We nextdeterminedwhether hybrid capsid particles composed of Cpolypeptidesfrom differenthepadnaviruses could be assem-bled in Xenopus oocytes. mRNA encoding A157L was coin-jected into oocytes along with equal quantities of either WHV-C, GSHV-C, or DHBV-C transcripts. 3S-labeled ly-sates werefractionatedon 10 to60%sucrosegradients into 14 fractions. Fractions 8 to 12, representing assembled capsids, were combined and immunoprecipitated with either anti-L MAb 18/7, an anti-HBV-C antibody, or an anti-DHBV-C
DHBV-C
*_X_~~~.i HBV-C
[image:4.612.336.542.70.440.2]_,; 'r Al57L
FIG 250. A HBV-C
200-/\
s:150-100- 1
600- Ae 57L
300~
200
O0
1 2 3 4 5 6 7 8 1t 11121314 Fraction
FIG. 2. Assemblyofataggedandtruncated HBVcoreproteinin
Xenopus oocytes. (A) Lysatesfrom 35S-labeledXenopus oocytes
in-jected with mRNA encoding HBV-C, DHBV-C, or 1v157L were
combined and resolved on step gradients of 10 to 60% sucrose.
Fourteenfractionswerecollected from the top ofeachgradientand analyzed by immunoprecipitation under nondenaturing conditions withpolyclonal anti-HBV-C and anti-DHBV-C antibodies followedby SDS-PAGEona13.5% gel.Thegradientfractionsareshown in order from the top fraction at the left(fraction 1)tothe bottom fractionat
theright (fraction 14).The positionsof the DHBV-C, HBV-C,and A157L speciesareindicatedattheright.Sizes in kilodaltonsareshown atthe left.(B)Phosphorlmager quantitationof35Ssignalintensityin eachfraction for the indicatedcorepolypeptideband in thegelshown inpanelA.
antibody. The labeledproteinswere analyzed by SDS-PAGE (Fig. 3A). Immunoprecipitation with anti-L clearly demon-strated that A157L core protein forms capsids with both WHV-C and GSHV-C(lanes 1 and4).The ratio of WHVto HBV and GSHV to HBV chains in the anti-L precipitates (lanes1and4)is similar tothat in theanti-HBV-Cprecipitates (lanes 2 and 5) and to that in the total lysate fraction (not shown). Inaddition, theabsoluteamountofWHV orGSHV core chains precipitated by anti-L is comparable to that precipitated by the broadly cross-reactive polyclonal anti-HBV-C antibodies. Together, these observations indicate that most orall of the animalhepadnavirusCpolypeptidesinthese fractionsare incorporated into mixedcapsids.
-E S J : -- ...P-4-~~~~~'
IL. ...;...i
B 1000 DHBV-C
800 600 400 200
0 -F--.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.125.240.72.236.2]Al57L co-injection with: WHy-C GSHV-C DHBV-C
r i I_ i_
aL aH aD aL aH aD aL aH aD
Al57Lco-injectionwith:
WHV-C GSHV-C DHBV-C
1 1l [
aL aH aD aL aH aD aL aH aD
...s>sF DHBV-C
29-184
3B
_om o DHBV-C
a- WHV/GSHV-C
to-& Al57L 14.3
-1 2 3 4 5 6 7 8 9
B WHV-C
I
aL aH aD
GSHV-C
I
aL aH aD
DHBV-C
I aL aH aD
43-18.4- __It
14.3
-_ _
_ _!'
__
*--WHV/GSHV-C
A157L
[image:5.612.55.296.70.364.2]1 2 3 4 5 6 7 8 9
FIG. 4. Compositionof dimer intermediates assembled in Xenopus oocytescoinjected withmRNAsencodingcoreproteins fromdifferent hepadnaviruses. Fractions 1 to 5 from the experiment of Fig. 3A, containing core dimerintermediates, were analyzed by immunopre-cipitation exactlyasdescribed for thecapsidfractions in the legend to Fig.3.Designations are as for Fig. 3.
-*-WHV/GSHV-C
40640 -A157L
14.3-1 2 3 4 5 6 7 8 9
FIG. 3. Composition of capsid particles assembled in Xenopus oocytescoinjected with mRNAsencoding core proteins from different hepadnaviruses. (A) Oocytes were coinjected with mRNAs encoding A157Land either WHV-C,GSHV-C,or DHBV-C.35S-labeledlysates wereresolved on step gradients of 10 to60%sucrose. Fractions 8 to 12, containing assembled capsids, were combined and immunoprecipi-tated undernondenaturingconditions with either an anti-LMAb (aL) or ananti-HBV-C(aoH) oranti-DHBV-C (aD) polyclonal antibody. The labeledprecipitatewasanalyzed by SDS-PAGE. The positions of the DHBV-C,WHV-C, GSHV-C, andA157Lspecies are indicated at theright.Sizes are indicated in kilodaltons at the left.(B) Analysis of mixed extracts. Oocytes were injected with a single transcript. 3S-labeled lysates werecombinedinthe indicated pairings and subjected
to sucrose gradient fractionation and immunoprecipitation as de-scribed forpanelA.Sizes are indicated in kilodaltons at the left.
Analysis of coinjections ofA157L with DHBV-C transcripts yielded very different results. Anti-L and anti-HBV-C, neither of which recognizes DHBV-C, immunoprecipitated only A157L (Fig. 3A, lanes 7 and 8); similarly, anti-DHBV-C antibody immunoprecipitated only DHBV-C (lane 9). This result shows thatunlike the WHV and GSHV core proteins, DHBV-C and HBV-Ccannotcoassemble;when coexpressed, theyyieldseparate homo-oligomeric capsid populations. This inability of mammalian and avian hepadnavirus cores to inter-actis notentirely unexpected given thesignificantdivergence intheir coreprotein sequences and their lack of immunologic cross-reactivity.
Toruleouttheformalpossibility that the results supporting mixedcapsid formationare duetononspecific aggregations of different homocapsids immunoprecipitated by anti-L, we per-formed a mixing experiment control (Fig. 3B). Oocytes were injected with either HBV-C (A157L), WHV-C, or GSHV-C mRNA,sothat onlyhomo-oligomeric capsids can form. Pairs oflysates containing one taggedand oneuntagged C polypep-tide species were then admixed and subjected to sucrose
gradient fractionation; capsid fractions were then immunopre-cipitated as described earlier. In contrast to the coinjection experiments, under these conditions L antibody was able to immunoprecipitate very little WHV-C and GSHV-C (lanes 1 and 4) compared with anti-HBV-C immunoprecipitation (lanes 2 and 5). Not surprisingly, the mixing experiment involvingDHBVcapsids and HBV capsids likewise showed no interaction (lanes 7 to 9). These results indicate that only a small percentage of the signal indicating mixed capsid forma-tion in thecoinjection experiment is attributable to nonspecific interactions ofhomo-oligomeric capsids.
Hepadnavirus core dimerization is cis preferential. In au-thenticcapsid assembly, dimers are the initial intermediates in assembly.We next set out todetermine whether heterodimers formduring mixedcapsidassembly in oocytes. To do this, we examined gradient-purified dimer intermediates fractionated from thecoinjected oocytes described above. Immunoprecipi-tation with anti-L revealed that A157L dimerized primarily with itself and formed few heterodimers with either WHV-C or GSHV-C(Fig.4, lanes1and4 versuslanes2and5).There are two possibleexplanations for this result: (i) monomer-mono-meraffinitydifferencesarising from sequence divergence favor homodimer formationoverheterodimer formation in trans, or (ii) the dimerization process operates preferentially incis;i.e., it favors theassociation ofmonomerstranslated from thesame mRNA.
Todistinguish between thesetwopossibilities,wecoinjected oocyteswith SP6 transcripts encodingHBV-C and HBV-CL; here, thetwoproteinsareidentical except for the presence of the L epitope tag. Both proteins assemble into capsidswith comparable efficiencies (data not shown). Coinjected lysates
were fractionated on 3 to 25% sucrose gradients into 14 fractions. Each fraction was then immunoprecipitated with anti-L or anti-HBV-C and analyzed by SDS-PAGE. Under thesesedimentationconditions,dimer intermediates sediment in fractions 3 to 10 and capsids pellet in fraction 14 (20). Coprecipitation of both chains with anti-L in fraction 14 confirms that mixed capsids form as expected (Fig. 5A). But
precipitationof the dimerfractions with anti-L(Fig. 5A,lanes 3 to 10) clearly demonstrated that HBV-CL forms dimers primarily with itself rather than with HBV-C, despite the presence ofHBV-Cin thesefractions, asrevealedby precipi-tation with anti-HBV-C (Fig. SB). Thus, the mixed capsids A
43-
29-18.4 d
_W -DHBV-C
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.318.551.74.212.2]a-L
29-A
41 ----HBV-CL
4_ -B--HBV-C
18.4
-14.3
-a-HBV-C
29-B
--HBV-CL
--BHV-C
18.4
-
[image:6.612.65.300.73.242.2]14.3--1 2 3 4 5 6 7 8 9 10 11 12 13 14
FIG. 5. Composition of dimer intermediates formed in Xenopus
oocytes coexpressing tagged and untagged HBV-C. Oocytes were
coinjected with mRNAs encoding HBV-C and the L-epitope tagged HBV-CL. 35S-labeled lysateswereresolvedon3 to25%sucrosestep
gradients. Each of the 14 fractions was immunoprecipitated under
nondenaturing conditions with eitheran anti-L MAb (a-L; A) anti-HBV-C polyclonal antibody (ct-HBV-C; B) and analyzed by
SDS-PAGE. Note that dimerintermediates sediment infractions 3 to 10,
andcapsidspelletat the bottom (fraction 14). Sizesare indicated in
kilodaltonsatthe left.
assembled inthecoinjectedoocytes(Fig. 5, lanes 14) mustbe composed predominantly of homodimers of tagged and
un-tagged C chains.
Using a Phosphorlmager to quantitate the amount of HBV-C and HBV-CL produced, we estimate that
approxi-mately 35% of dimers found in the coinjection experiment should have been heterodimers, assuming free and random association of HBV-C and HBV-CLmonomers leading to a
binomial distribution ofdimerproducts. Infact, ifwe correct
forantibodycross-reactivityandimmunoprecipitation efficien-cies, our results show that heterodimers account for at most
7% (and probably less) of the dimer pool. These findings indicate that C proteins display a marked cis preference for
dimerization in the oocyte. Possible explanations for this will beconsideredbelow.
DISCUSSION
Sequence requirements forcore-coreinteractions. We have
used the two-hybrid systemin apermissive human hepatoma
cell line (HepG2) to characterize core protein interactions
involved in HBV capsid assembly. We have shown that the full-length core protein fused downstream of the GAL4 or
VP16 domainis able to associate with itself; this association, which normally occurs in the cytoplasm, must have stably persistedinthenucleus,where the hostRNApolymerase II is located.
Corefusions withatruncationof the C-terminal 41 aaalso
oligomerized with full-length cores, aspredicted from in vivo
capsid assembly experiments. Adeletion that extended 68aa
upstreamfromtheCterminus failedtointeract; this result also
agreeswith findings from earlier assembly studies (2, 3). The effectofN-terminal deletionsoncore-core association hasnot
beenpreviously reported. We engineered corefusions lacking
the first 28 and 37 residues from core; neither deletion
constructoligomerized with the full-lengthcorefusion.
Unfor-tunately,oneofthe major limitations in usingthe two-hybrid
assay is that only fusions that yield a positive CAT signal are informative. Fusions that fail to associate with core in this assay mayindeed lack a domain critical for core interaction, or they mayhave simply folded into an improper conformation or be unstable. The fact that the full-length and the Arg tail-deficient core fusions were able to associate suggests that the GAL4 and VP16 domains are discrete structures that did not interfere with the folding of the C portion of the fusion. We favor the view that the assembly-defective deletions may have globally disrupted the secondary and tertiary structure of the core polypeptide. Although the core polypeptide structure has not been elucidated by crystallography, Agros and Fuller have proposed a structure based on modeling with the vp3 capsid protein of a picornavirus (1). Their sequence alignment anal-ysis suggests that the core protein will adopt aneight-stranded antiparallel ,B-barrel structure, with most of the Arg-rich tail located distally. Both of the N-terminal deletions and the 68-aa C-terminaldeletion would be predicted to remove at least two of the 3 strands and probably disrupt the secondary and tertiary structure important forintersubunit association. The structure of C protein may preclude the identification of a discrete contiguous domain that mediates core association; discontinuous residues critical to forming a contact surface during dimerization may be brought together only during the properfolding of the chain.
Assembly of mixed capsids in the Xenopus oocyte. Coexpres-sion ofA157L, a truncated and tagged HBV-C, with WHV-C or GSHV-C led to the assembly of mixed capsid particles. Coexpression of A157L and DHBV-C, which differs consider-ably in sequence and size from the mammalian cores, did not lead to the formation of mixed capsids. Assembly of HBV-C into capsids in the oocyte proceeds without detectable assem-blyintermediates other than dimers (20, 21); our coexpression experiments with the animal viral cores also failed to reveal any higher-molecular-weight assembly intermediates in hetero-oligomeric capsid formation. A number of explanations for this are possible. Most likely, such intermediates are extremely transient as a result of the efficient and highly cooperative assembly of capsids from dimer building blocks (15, 20); alternatively, they may be unstable in cell extracts or may dissociate duringcentrifugation.
Our results with the oocyte assembly system raise the intriguing issue of whether mixed nucleocapsids can form duringhepadnavirus replication. Such a situation could occur in vivo for WHV and GSHV, since both can replicate in woodchuck hosts (14a). Hybrid nucleocapsids might affect several processes in the hepadnavirus life cycle. Cores are intimately involved in RNAencapsidation, DNA synthesis, and viral budding. How a mixed nucleocapsid would affect the specificity or efficiency of any one of these process remains to be seen. There is little precedent for mixed nucleocapsid formationin other viruses with icosahedral capsids. In retro-viruses, a recent report has demonstrated that mixed core particles between MuLV and HIV type 1 can be made, but only when artificial chimeric Gag precursors containing both HIV and MuLVdeterminants are coexpressed with wild-type MuLV Gag proteins (7). This finding suggests that homo-dimerizationof the MuLV Gag proteins is the driving force for assembly, with the heterologous HIV sequences being incor-poratedas aresult. As yetthere have been no reports of mixed capsidformation between wild-type Gag proteins.
A cis preference in dimer formation. During the mixed capsid assembly reaction in Xenopus oocytes, the coexpressed core proteins formed primarily homodimers. Even when the coexpressed C polypeptides are identical except for the L epitope tag, homodimers still form preferentially. These results
on November 9, 2019 by guest
http://jvi.asm.org/
indicate thatcore dimerization in theoocytedisplays a strong cis preference. Thissuggests that core monomers are not free todiffuse and associatewith other monomers and is consistent with the notion that dimerization occurs very rapidly after monomerproduction.In thisview, a monomer is mostlikelyto associate with the nearestmonomeravailable inthe region of itssynthesis, i.e., those translated from thesamepolysome.In the extreme case, dimers mayform cotranslationally between adjacent, nascent corepolypeptides still tethered tothe elon-gating ribosome. This may account for the absence of detect-able monomers in the oocyte assembly system (20). Once formed, dimers are free to diffuse and associate with other dimers.
We donot know ifthiscis preference operates in mamma-lian cells, but this seems likely to be so. Our results in the mammalian two-hybrid system certainly do not exclude this possibility. Since GAL4 binds to DNAas a dimer(12), it is likely that the GAL4-core fusion proteins active in this assay are themselves dimeric and that all C-C interactions being detected in thissystemthusinvolve homodimers of Cproteins.
ACKNOWLEDGMENTS
WethankSophieRoy and Jon Pollack for manyhelpfuldiscussions duringthe courseof thiswork and IanTaylor forcriticalreading ofthe manuscript.
Thiswork was supportedbygrantsfrom the National Institutes of Health.
REFERENCES
1. Argos,P.,and S. D. Fuller. 1988. Amodelforthe hepatitis B virus
coreprotein: predictionofantigenic sitesandrelationshiptoRNA viruscapsidproteins. EMBOJ.7:819-824.
2. Beames, B., and R. Lanford. 1993.Carboxy-terminal truncations of the HBV coreprotein affect capsid formationand theapparent sizeofencapsidated HBV RNA.Virology 194:597-607. 3. Birnbaum,F., and M. Nassal.1990.HepatitisBvirus nucleocapsid
assembly:primarystructure requirements in the coreprotein. J. Virol. 64:3319-3330.
4. Borisova, G., B. Arya, A. Dislers,0.Barschukova, V. Tsibinogin, D.Skrastina, M.Eldarov,P.Pumpens, K.Skryabin, and E. Grens. 1991. Hybridhepatitis Bvirusnucleocapsidbearingan immuno-dominantregion from hepatitisB virussurface antigen.J.Virol. 67:3696-3701.
5. Clarke, B. E., S. E. Newton, A. R. Carrol, M. J. Francis, G. Appleyard, A. D. Syred, P. E. Highfield, D. J. Rowlands, and F. Brown. 1987.Improvedimmunogenicity ofapeptide epitope after fusiontohepatitisBcoreprotein. Nature(London) 330:381-384. 6. Cohen,B., andJ.Richmond. 1982. Electronmicroscopy of hepa-titis B core antigen synthesized in E. coli. Nature (London) 296:677-678.
7. Deminie, C. A., and M. Emerman.1993.Incorporationof human immunodeficiencyvirus type 1Gagproteinsinto murineleukemia virusvirions.J.Virol. 67:6499-6506.
8. Fearon, E. R, T. Finkel, M. L. Gillison, S. P. Kennedy, J. F.
Casella, G. F. Tomaselli, J. S. Morrow, and C. V. Dang. 1992. Karyoplasmic interactionselection strategy: a general strategyto detect protein-protein interactionsinmammalian cells. Proc. Natl. Acad.Sci. USA89:7958-7962.
9. Fields, S., and0. Song. 1989. A novelgeneticsystem to detect protein-proteininteractions.Nature(London)340:245-246. 10. Hirsch, R.,R Colgrove,and D.Ganem. 1988.Replicationof duck
hepatitis B virus in two differential human hepatoma cell lines aftertransfection with cloned viral DNA.Virology167:136-142. 11. Luban,J., K. B. Alin, K. L. Bossolt, T. Humaran, and S. P. Goff.
1992. Genetic assay formultimerizationof retroviralGag polypro-teins.J.Virol.66:5157-5160.
12. Marmorstein, R., M.Carey, M. Ptashne,andS.C. Harrison.1992. DNArecognitionbyGAL4: structureofaprotein-DNAcomplex. Nature(London) 356:408-414.
13. Miyonahara, A., T. Imamura, M. Araki, K. Sugawara, N. Ohtomo, and K. Matsubara. 1986. Expression ofhepatitis B virus core antigen geneinSaccharomycescerevisiae:synthesis oftwo polypep-tides translated fromdifferent initiation codons.J. Virol. 59:176-180.
14. Nassal, M.1992. Thearginine-richdomain of thehepatitisBvirus coreprotein isrequired forpregenomeencapsidationand produc-tive viralpositive-strand DNAsynthesis butnotfor virus assembly. J.Virol.66:4107-4116.
14a.Seeger, C., P. L. Marion, D.Ganem,and H. E.Varmus. 1987. In vitro recombinants ofground squirrel andwoodchuck hepatitis viral DNAsproduce infectious virusinsquirrels.J.Virol. 61:3241-3247.
14b.Seifer, M., and D. N. Standring. 1993. Recombinant human hepatitis Bvirusreversetranscriptase isactive in the absence of the nucleocapsid or the viral replication origin, DRI. J. Virol. 67:4513-4520.
15. Seifer,M., S. Zhou, and D. N.Standring. 1993.Amicromolar pool ofantigenicallydistinct precursorsis requiredtoinitiate cooper-ativeassemblyofhepatitisBviruscapsidsinXenopusoocytes. J. Virol. 67:4513-4520.
16. Stahl, S. J., and K. Murray. 1989. Immunogenicity ofpeptide fusionsto hepatitisBvirus coreantigen. Proc. Natl. Acad. Sci. USA86:6283-6287.
17. Standring, D. N. 1991. The molecularbiologyof thehepatitis B viruscoreprotein,p.145-169. In. A. McLachlan(ed.),Molecular biologyof thehepatitisB virus.CRC, BocaRaton,Fla. 18. Standring,D. N., J.-H.Ou,and W.J. Rutter. 1986.Assemblyof
viral particlesin Xenopusoocytes: pre-surface antigens regulate secretion of thehepatitisB viralsurfaceenvelope particle.Proc. Natl. Acad. Sci. USA 83:9338-9342.
19. Zhou, S., and D. N. Standring. 1991. Production ofhepatitisB virusnucleocapsidlikecoreparticlesinXenopusoocytes:assembly
occursmainlyinthecytoplasmand doesnotrequirethenucleus. J.Virol.65:5457-5464.
20. Zhou, S., and D. N. Standring. 1992. Hepatitis Bvirus capsid particlesareassembled fromcore-protein dimerprecursors.Proc. Natl. Acad.Sci.USA 89:10046-10050.
21. Zhou,S.,S.Q.Yang, and D. N.Standring.1992.Characterization ofhepatitisBviruscapsid particle assemblyinXenopusoocytes.J. Virol.66:3086-3092.