Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Trafficking to the Plasma Membrane of the Seven-Transmembrane
Protein Encoded by Human Herpesvirus 6 U51 Gene Involves
a Cell-Specific Function Present in T Lymphocytes
LAURA MENOTTI,
1PRISCO MIRANDOLA,
1MASSIMO LOCATI,
2,3AND
GABRIELLA CAMPADELLI-FIUME
1*
Section on Microbiology and Virology, Department of Experimental Pathology, University of Bologna, Bologna,
1Institute of Pharmacological Research “M. Negri,” Milan,
2and Section on General Pathology, Department of
Biotechnology, University of Brescia, Brescia,
3Italy
Received 3 August 1998/Accepted 15 October 1998
The sequence of human herpesvirus 6 (HHV-6) U51 open reading frame predicts a protein of 301 amino acid
residues with seven transmembrane domains. To identify and characterize U51, we derived antipeptide
polyclonal antibodies and developed a transient expression assay. We ascertained that U51 was synthesized in
cord blood mononuclear cells infected with either variant A- or variant B-HHV-6 and was transported to the
surface of productively infected cells. When synthesized in transient expression systems, U51 intracellular
trafficking was regulated in a cell-type-dependent fashion. In human monolayer HEK-293 and 143tk
2
cells,
U51 accumulated predominantly in the endoplasmic reticulum and failed to be transported to the cell surface.
In contrast, in T-lymphocytic cell lines J-Jhan, Molt-3, and Jurkat, U51 was successfully transported to the
plasma membrane. We infer that transport of U51 to the cell surface requires a cell-specific function present
in activated T lymphocytes and T-cell lines.
Proteins with seven-transmembrane-domain structures are
divided into two superfamilies depending on their ability to
bind G proteins and are classified as G-protein-coupled
recep-tors and nonreceptor seven-transmembrane-domain proteins
(29). Beta- and gammaherpesviruses encode
seven-transmem-brane-domain proteins which function as chemokine receptors
and whose roles in the viral replicative cycle are diversified.
Thus, human cytomegalovirus (HCMV) US28 binds
b
-chemo-kines, and its signaling ability can be monitored as intracellular
Ca
21mobilization (20, 36). The protein encoded by human
herpesvirus 8 (HHV-8) open reading frame (ORF) 74
func-tions as a promiscuous chemokine receptor for
a
- and
b
-che-mokines and appears to be constitutively active in signal
trans-duction, raising the possibility that it is part of the transforming
potential of the virus (3, 9). The ECRF3 protein encoded by
herpesvirus saimiri behaves as an
a
-chemokine receptor with
specificity for interleukin 8, GRO-
a
, and NAP-2 (1, 37).
Hu-man and murine CMV encode two additional
G-protein-cou-pled receptors (UL33 and UL78) not yet characterized (4, 12,
34, 38). Recently, the viral and cellular seven-transmembrane
G-protecoupled receptors have been the focus of much
in-terest because some of them, including HCMV US28, act as
coreceptors for entry of human immunodeficiency virus into
cells (42) and, in the case of poxviruses, because they appear to
be effectors of viral immune response evasion strategies
(re-viewed in references 24, 43, and 44).
Notwithstanding extensive studies on the epidemiology of
HHV-6 infections and associated diseases (reviewed in
refer-ences 6 and 7), little is known of the functions of specific gene
products and of their role in the viral infectious cycle and
associated diseases. In vivo, HHV-6 appears to infect T
lym-phocytes, monocytes, astrocytes, oligodendrocytes (8, 10, 28,
30, 47), and possibly other not yet characterized cells in various
tissues (see references 6, 7, and 13). T lymphocytes and/or
monocytes are believed to be the site of latency (30). Because
of its ability to persist in the host after primary infection, the
virus must have evolved strategies to evade the immune
sys-tem. The molecular bases for this phenomenon are not yet
clear. In vitro, the virus grows in activated T lymphocytes and
monocytes, in T-cell lines (33), and, to a limited extent, in some
epithelial and endothelial cell lines (see references 6 and 46).
The sequence of the HHV-6 genome predicts two proteins
with seven transmembrane domains encoded by U51 and U12
ORFs (23, 31). U12 was recently shown to be expressed late in
the viral replicative cycle; to function as a receptor for the
b
-chemokines RANTES, MCP-1, and MIP-1
a
; and to respond
to chemokine binding with a signaling pathway evidenced as
mobilization of intracellular Ca
21(27).
The initial objective of this study was to identify the protein
encoded by HHV-6 U51 ORF. We ascertained that U51
pro-tein is expressed in productively infected cord blood
mononu-clear cells (CBMCs) and is transported to the infected cell
surface. In the course of these studies, a rather unusual
traf-ficking property of U51 emerged, as we noticed that U51 failed
to be transported to the cell surface in human monolayer cells,
where it accumulated predominantly in the endoplasmic
retic-ulum (ER), but reached the plasma membrane of
T-lympho-cytic cell lines. The results indicate that transport of U51 to the
cell surface requires a cell-specific function present in T
lym-phocytes.
MATERIALS AND METHODS
Cells and viruses.Human embryonic kidney 293 (HEK-293) cells, human 143tk2cells, and Vero green monkey kidney cells were grown in Dulbecco’s modified Eagle medium containing 5% fetal calf serum (Gibco Laboratories). Primary CBMCs were cultured as described elsewhere (17). HHV-6(A)U1102, HHV-6(B)Z29 (14, 32), and vaccinia virus recombinant expressing the T7 RNA polymerase (VacT7) (19) were described elsewhere. Infection of CBMCs with HHV-6 was monitored routinely by immunofluorescence with monoclonal anti-body (MAb) 2D10 to glycoprotein B (gB), as described elsewhere (18).
* Corresponding author. Mailing address: Dipartimento di
Patolo-gia Sperimentale, Sezione di MicrobioloPatolo-gia e ViroloPatolo-gia, Via San
Gia-como, 12, 40126 Bologna, Italy. Phone: 39 051 354733/34. Fax: 39 051
354747. E-mail: [email protected].
325
on November 9, 2019 by guest
http://jvi.asm.org/
Antibodies.Commercially available antibodies were antihemagglutinin (anti-HA) antibody (AntiXpress) (Invitrogen), anti-CC-chemokine receptor 5 (anti-CCR5) MAb LS100/2D7 (R&D Systems, Abingdon, United Kingdom), fluorescein isothiocyanate (FITC)-conjugated anti-mouse antibody (Jackson), FITC-conjugated anti-rabbit antibody (Dako), biotinylated anti-rabbit antibody
and avidin-biotin-peroxidase (Vector Laboratories), and FITC-conjugated goat F(ab9)2anti-mouse antibody (Becton Dickinson). MAb no. 30 to herpes simplex
virus (HSV) gD was described elsewhere (5). Rabbit polyclonal anticalnexin antibody was a gift of A. Helenius, University of Zurich.
cDNA synthesis and reverse transcription-PCR (RT-PCR).Total RNA was extracted with RNA-ZoldB (Tel-Test, Friendswood, Tex.) from 53106
unin-fected or HHV-6(B)Z29-inunin-fected Molt-3 cells maintained in the presence or absence of phosphonoacetic acid (PAA) (500mg/ml) from the time of infection, as described elsewhere (35). RNA was precipitated with isopropanol and resus-pended in 100ml of 100 mM sodium acetate–5 mM magnesium sulfate contain-ing 20 U of RNase inhibitor (Ambion, Inc., Austin, Tex.), and traces of DNA were removed by three cycles of digestion with 40 U of DNase H (Boehringer, Mannheim, Germany), each of 1 h at room temperature. RNA was purified by acid phenol-chloroform (1:1) extraction and ethanol precipitation and resus-pended in 50ml of diethylpyrocarbonate-water containing 10 U of RNase inhib-itor. One microgram of RNA was reverse transcribed with 10 U of avian myelo-blastosis virus reverse transcriptase (cDNA Cycle Kit; Invitrogen, Leek, The Netherlands) and hexamer random primers at 42°C for 1 h and extracted with phenol-chloroform. One-tenth of total cDNA was amplified with primers U51/5X (GAATCATTACCTCGAGTATTCAGGATGGAG) and U51/3 (TAA GAACGCGAGAAAACACT), able to amplify both HHV-6(A) and -6(B) DNA, with 0.15 U of thermostable DNA polymerase (Societa` Italiana Chimici, Rome, Italy)–20 pmol of each primer–3 mM MgCl2–200mM (each) dNTP.
Amplifica-tion was carried out with 35 cycles of 1 min at 94°C, 30 s at 55°C, and 1 min at 72°C, preceded by 5 min at 94°C and supplemented with 5 min at 72°C. To exclude the possibility that positive amplification might result from viral DNA contamination of RNA samples, 200 ng of total RNA was directly subjected to PCR amplification. Specificity of the primers U51/5X and U51/3 for viral se-quences was assessed by lack of amplification of uninfected CBMC DNA (10 ng) and the corresponding cDNA. Forb-actin amplification, primers were as de-scribed elsewhere (45), and the conditions differed from those above with respect to MgCl2(2 mM) and amount of primers (40 pmol). Thirty-five cycles were
performed at 95°C for 30 s, 55°C for 30 s, and 72°C for 30 s (increase of 1 s at each new cycle) preceded by 5 min at 94°C. U31 primers were described else-where (35). For U12 amplification, specific primers were GACAAGCGACGG GATCCACACTGTCATTGAGC and GAACAGACTGCATGATAGATG, and the PCR conditions were the same as those for U51.
Plasmids.U51 ORF (map coordinates 82,574 to 83,479 [AGMN GenBank locus]) (23) was cloned by PCR technology under the immediate-early CMV and T7 promoters in two versions. p51-HA contained the U51 ORF cloned in pcDNA3.1-His vector (Invitrogen) with the heterologous HA epitope and six-histidine tag at the N terminus. p51 contained the entire U51 ORF inserted in pcDNA3.1(2) Myc His vector, which is designed for insertion of Myc-His tag at the C terminus of the engineered proteins. In p51, the natural stop codon of the protein was maintained, thus preventing the addition of the Myc-His tag. For p51-HA cloning, U51 was amplified from 20 ng of HHV-6(A)U1102-infected J-Jhan cell DNA with 1 U of AmpliTaq Gold (Perkin-Elmer) in 3 mM MgCl2–
200mM (each) dNTP, with 10 cycles of 1 min at 94°C, 30 s at 50°C, and 1 min at 72°C, followed by 25 cycles of 1 min at 94°C, 30 s at 60°C, and 1 min (increase FIG. 1. (A) Hydrophobicity profile of U51 (Kyte-Doolittle) and localization
[image:2.612.56.292.75.337.2]of peptides 1 and 2, employed as antigens to derive immune sera 8 and 6, respectively. (B) U51 expression by in vitro transcription-translation (IVTT) (lane a). In vitro transcription-translation-produced U51 (arrowhead) was im-munoprecipitated with preimmune (PI) and immune (I) sera 6 (lanes b and c) and 8 (lanes d and e) and anti-HA (a-HA) antibody (lane f). aa, amino acid; TM, transmembrane; N-ter and C-ter, N and C termini, respectively. Number at left show molecular mass (in kilodaltons).
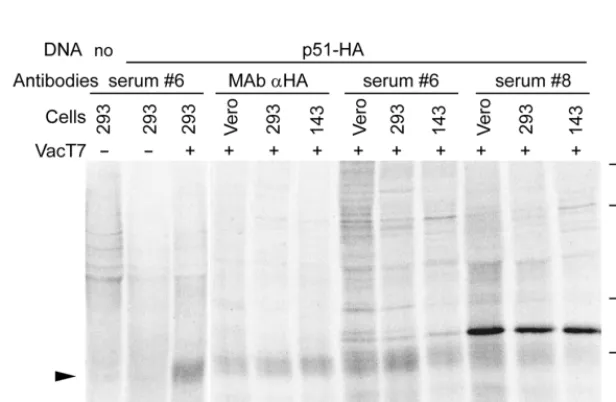
FIG. 2. Transient expression of U51 in human monolayer HEK-293, 143tk2, and Vero cells infected with VacT7 and transfected with p51-HA (p51-HA) or mock transfected (no). Cells were labeled with [35S]methionine from 4 h after transfection till harvesting at 18 h. U51 (black arrowhead) was immunoprecipitated by immune
sera 6 and 8 and by anti-HA-tag MAb. Numbers at right show molecular mass in kilodaltons.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.149.457.497.698.2]of 3 s at each new cycle) at 72°C. Primers U51/5B (GTTTATTCAGGATCCA GAAAGAAACGAAGTC) and U51/3X (CCATATTTGACCCTCGAGGAAT CAGCGCC) inserted a BamHI and an XhoI restriction site at 59and 39regions, respectively. The U51 ORF in p51-HA contained two substitutions at the 59end, which resulted in replacement of the first methionine (M) and of glutamate (E) with isoleucine (I) and glutamine (Q), respectively. For p51 cloning, primers 51N5 (GAATCATTACTTCGGCTAGCCAGGATGGAGAAAG) and 51H3ST (GAATCAGCGCCGAAGCTTTATTCTCTTATG) inserted NheI and HindIII restriction sites at the 59and 39ends of amplified fragments, respectively. HSV gD gene was cloned in pcDNA3.1 vector to yield pgD, by PCR amplification with primers TATCCTTAAGGGATCCTTTGTGTGGTGCG and TAAGGTCCCA AGCTTACCCCGCAGACC, which introduced BamHI and HindIII restriction sites at 59and 39ends of the amplification product, respectively. gD was amplified from 2 ng of HSV-1(F)-infected Vero cell DNA and 1 U of AmpliTaq Gold (Perkin-Elmer) in 2 mM MgCl2–200mM (each) dNTP, with 15 cycles of 1 min
at 94°C, 30 s at 55°C, and 1 min at 72°C, followed by 25 cycles of 1 min at 94°C, 30 s at 60°C, and 1 min (increase of 3 s at each new cycle) at 72°C. pCCR5 was described elsewhere (2).
Production of polyclonal antibodies.Two synthetic peptides spanning the region from residues 142 to 168 and from residues 218 to 235, second and third predicted extracellular domains, respectively, were coupled to polybranched polylysine carrier (Biopolymer Core Facility, Department of Microbiology and Immunology, University of Maryland, Baltimore, Md.). New Zealand White female rabbits were immunized with six subcutaneous injections of 400mg each, emulsified in Freund adjuvant (Difco Laboratories). When specified, the rabbit sera were cleared of antibodies binding specifically to uninfected cell proteins by preabsorption to acetone-fixed uninfected CBMCs for 2 h at 4°C followed by centrifugation at 43g for 15 min.
Synthesis of U51 by in vitro transcription-translation.The U51-HA fusion protein was synthesized by in vitro transcription-translation performed with the TNT T7 quick-coupled transcription-translation system (Promega). One micro-gram of p51-HA DNA was incubated with TNT Master Mix in the presence of 15 to 30mCi of [35S]methionine and [35S]cysteine (specific activity, 1,000 Ci/
mmol; the Radiochemical Centre, Amersham, England) in a final volume of 50
ml for 90 min at 30°C. For immunoprecipitations, aliquots of the reaction product were reacted with either anti-HA MAb or with the rabbit preimmune and immune sera, and the immunocomplexes were harvested on protein A-Sepha-rose beads, as detailed below. Aliquots of the reaction mixtures or the immu-nocomplexes were solubilized in solubilizing solution (2% sodium dodecyl sul-fate, 5% b-mercaptoethanol, 2.75% sucrose, 50 mM Tris HCl [pH 7], bromophenol blue), and separated on 10% polyacrylamide gels cross-linked with
N,N9-diallytartardiamide. Fixed gels were soaked in Amplify (the Radiochemical Centre), dried, and analyzed in a Bio-Rad molecular imager.
Immunoprecipitations.Uninfected or HHV-6(A)U1102- or HHV-6(B)Z29-infected CBMCs were labeled for 18 h with a [35S]methionine and [35S]cysteine
mixture, 40 to 50mCi of medium containing 1/10 the usual concentration of unlabeled methionine and cysteine per ml, and 1% fetal bovine serum. HEK-293, Vero, and 143tk2cells were infected with VacT7 (10 PFU/cell) and immediately thereafter transfected with p51-HA. Cells were incubated in medium containing 10 mM hydroxyurea and labeled with 40 to 50mCi of [35S]methionine and
[35S]cysteine per culture from 4 h after beginning of transfection till harvesting
at 18 h. Cells were solubilized in PBS* (phosphate-buffered saline [PBS], 1% sodium deoxycholate, 1% Nonidet P-40, 0.1 mg each of TLCK [Na -p-tosyl-L-lysine chloromethyl ketone] and TPCK [N-tosyl-L-phenylalanine chloromethyl ketone] per ml) (Sigma) and centrifuged at 55,0003g for 75 min. The
super-natants were incubated with anti-HA monoclonal or rabbit serum for 3 h on ice; immunocomplexes were harvested on protein A-Sepharose. Immunoprecipita-tions with rabbit sera were carried out by first reacting the lysates (100ml) with the corresponding preimmune sera and harvesting the immunocomplexes on protein A-Sepharose beads. The clear supernatants, devoid of proteins reacting aspecifically with preimmune sera, were then reacted with the immune sera.
Cell surface labeling by biotinylation.Uninfected or HHV-6(A)U1102- or HHV-6(B)Z29-infected CBMCs and 143tk2cells infected with VacT7 and transfected with p51-HA and pgD were labeled with [35S]methionine and
[35S]cysteine mixture. Immediately prior to harvesting, cells were washed with
PBS and incubated in a solution of 75mg of ImmunoPure Sulfo-NHS-LC-Biotin (Pierce) per ml in 50 mM NaHCO3–100 mM NaCl (pH 8.5) for 30 min at room
[image:3.612.56.289.69.684.2]temperature. Cells were rinsed with 50 mM NaCl in 100 mM Tris (pH 8), lysed with PBS*, and then subjected to immunoprecipitation with polyclonal serum no. 6 or anti-HA MAb, as detailed below. The immunoprecipitated proteins were separated by electrophoresis in a 10% polyacrylamide gel and transferred to a
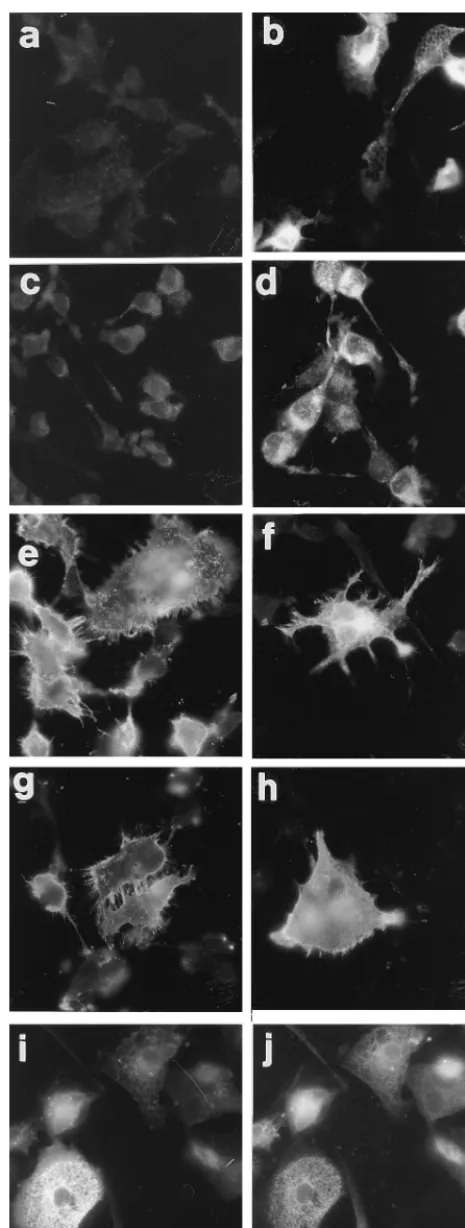
FIG. 3. Immunofluorescence localization of U51 (a to d and i) and, for comparison, of HSV gD (e and f), CCR5 (g and h), and calnexin (j) in transiently expressing 143tk2cells. Cells were preinfected with VacT7 (7 PFU/cell) and immediately thereafter transfected with p51-HA (a, b, i, and j), p51 (c and d), pgD (e and f), and pCCR5 (g and h). Cells were fixed at 14 h after transfection
and stained with anti-HA MAb (a, b, and i), immune serum 6 (c and d), MAb 30 to gD (e and f), MAb to CCR5 (g and h), and rabbit polyclonal serum to calnexin (j). Cells were paraformaldehyde fixed and permeabilized (b, d, f, h, i, and j) or not permeabilized (a, c, e, and g) with Triton X-100 prior to reaction with appropriate primary antibodies. (i and j) Double immunofluorescence of the same cells transfected with pU51-HA and stained with anti-HA antibody (i) or anticalnexin antibody (j).
on November 9, 2019 by guest
http://jvi.asm.org/
nitrocellulose sheet. The biotinylated proteins were detected by incubation with avidin-biotin-conjugated peroxidase (Vector Laboratories) for 30 min and dia-minobenzidine as substrate. The radiolabeled proteins were detected by autora-diography in a Bio-Rad molecular imager.
Immunofluorescence analysis.Monolayer cells were grown on glass coverslips, and suspension cells were allowed to deposit on Dynatech slides (PBI, Milan, Italy). Cells were fixed with 4% paraformaldehyde in PBS for 10 min at room temperature and permeabilized with 0.1% Triton X-100 in PBS, or with cold acetone for 10 min, as specified in the figure legends. The cells were blocked with 20% newborn calf serum for 30 min at 37°C and reacted with primary antibodies diluted in 20% newborn calf serum in PBS. Incubation was for 1 h at room temperature for monolayer cells and for 30 min at 37°C for suspension cells. For enhanced immunofluorescence, binding of primary antibody was detected with a biotinylated anti-rabbit secondary antibody (Vectastain Kit; Vector Laborato-ries), 30 min at 37°C, followed by incubation with Extravidin-tetramethyl rhoda-mine isothiocyanate (TRITC) (Sigma) diluted 1:100 in PBS for 30 min at 37°C. Fluorescence-activated flow cytometry.Transfected, VacT7-infected cells were harvested 14 h postinfection and resuspended in PBS at 53105/ml. When
required, cells in 2-ml aliquots were permeabilized by 3 min of incubation at 4°C with 2.5ml of phosphatidylcholine (20 mg/ml) in methanol and being washed with a large volume of 1% fetal bovine serum in PBS. Both intact and perme-abilized cells were then washed and resuspended in ice-cold fluorescence-acti-vated cell sorting (FACS) buffer (Hanks balanced salt solution, 1% fetal bovine serum, 0.02% NaN3) at 107/ml. Fifty microliters of cell suspensions was mixed
with 50ml of appropriate primary antibody diluted in FACS buffer and incubated for 30 min at 4°C. Cells were washed twice with 50ml of ice-cold FACS buffer, resuspended in 100ml of appropriate secondary antibody [FITC-conjugated goat F(a,b9)2anti-mouse], diluted 1:50 in FACS buffer, and incubated for 30 min at
4°C. After 20 min, 10ml of 25-mg/ml propidium iodide in PBS was added. At the end of incubation, cells were washed twice and resuspended in 500ml of ice-cold FACS buffer. Samples were analyzed in a FACStar fluorimeter (Becton Dickin-son, Irvine, Calif.).
RESULTS
Construction of U51 expression vectors.
The U51 ORF of
HHV-6(A)U1102 (map coordinates 82,574 to 83,479) predicts
a protein of 301 amino acid residues, 34.714 kDa in molecular
mass, with no predicted N-glycosylation site. Hydrophobicity
profiles, carried out with PHDhtm and Profile network
predic-tion of topology (PHDtopology) (40, 41), and studies of
well-characterized seven-transmembrane proteins predict that the
N terminus is located extracellularly and that the C terminus is
located intracellularly (Fig. 1A). The U51 ORF was cloned in
pcDNA3.1 vectors, under the control of the T7 and the
imme-diate-early CMV promoters, suitable for both in vitro
tran-scription-translation and eukaryotic expression systems.
p51-HA carried the U51 coding sequence fused at the N
ter-minus with a heterologous epitope derived from influenza virus
HA and a polyhistidine tract. p51 carried the U51 ORF with no
heterologous epitope and no polyhistidine tract. In vitro
tran-scription-translation reaction of p51-HA in the presence of
[
35S]methionine and [
35S]cysteine yielded a protein with an
apparent molecular mass of 28 kDa immunoprecipitated with
the MAb to HA (Fig. 1B, lanes a and f). The observed Mr
is
slightly lower than that expected for the protein encoded by the
U51 ORF. Addition of microsomes to the reaction did not
result in a decrease in the electrophoretic mobility of U51
protein, suggesting that it is not subjected to extensive
post-translational processing, at least in the in vitro system.
Production of antipeptide antibodies.
Peptides 1 and 2,
spanning residues 142 to 168 and 218 to 235, respectively, are
predicted to be located in the second and third extracellular
domains, respectively (Fig. 1A). They were coupled to
poly-branched polylysine carrier and served as immunogens to
de-rive polyclonal sera 8 and 6, respectively. Both sera
immuno-precipitated U51 synthesized in vitro in the presence of
[
35S]methionine (Fig. 1B, lanes c and e) and were even more
effective than anti-HA MAb (lane f), particularly serum 6.
Preimmune sera (lanes b and d) did not immunoprecipitate the
U51 protein, demonstrating the specificity of the immune sera.
Transient expression of U51 gene.
To establish a eukaryotic
expression system, HEK-293, 143tk
2
, and Vero cells were
transfected with p51-HA DNA and labeled with [
35S]methi-onine and [
35S]cysteine from 4 h after transfection till
harvest-ing at 18 h. Figure 2 shows that, in cells infected immediately
prior to transfection with a recombinant vaccinia virus carrying
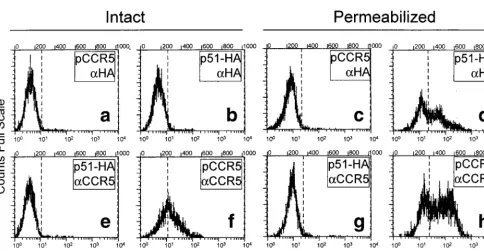
FIG. 4. Profile of fluorescence-activated flow cytometry of HEK-293 cells transiently expressing U51 protein (p51-HA) and stained with anti-HA antibody (aHA) (b and d) or transiently expressing CCR5 (pCCR5) and stained with anti-CCR5 antibody (aCCR5) (f and h). The cutoff values were defined in cells transfected with pCCR5 and stained with anti-HA (a and c) or in cells transfected with p51-HA and stained with the heterologous anti-CCR5 antibody (e and g). Cutoff values were defined separately for permeabilized cells (c and g) and intact cells (a and e). (a, b, e, and f) Intact cells; (c, d, g, and h) permeabilized cells.on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.546.67.318.2]the T7 RNA polymerase (VacT7), U51 expression was readily
detected in all three cell lines by immunoprecipitation with
serum 6 and with anti-HA MAb (black arrowhead). Extent of
expression was higher at 18 than at 8 h after transfection (data
shown for 18 h). Serum 8, but not serum 6,
immunoprecipi-tated in addition a 33-kDa M protein visible in VacT7-infected
cells and not in uninfected cells, suggesting a possible
cross-reactivity of the serum with VacT7-encoded proteins. Cells
transfected with plasmid alone showed no expression of U51
protein in any of the cells tested (lane b for HEK-293 cells).
The reason for this is unclear, as, generally, pcDNA3 vectors
allow strong constitutive expression in the cell lines employed
in this study. Whether this reflects a specific instability of U51
mRNA is not known. In all subsequent transient expression
experiments, U51 synthesis was induced with VacT7 infection.
U51 is not expressed at the plasma membrane and
accumu-lates in the ER of transfected human monolayer cells.
The cell
surface localization of U51 protein in HEK-293 and 143tk
2
cells transfected with p51-HA, or p51 (no heterologous tag),
was investigated by immunofluorescence microscopy. As a
con-trol for cell surface localization, replicate cultures were
trans-fected with plasmid pgD, carrying the gene for the HSV
mem-brane protein gD gene, or plasmid pCCR5, carrying the gene
for CCR5, a seven-transmembrane protein, cloned in
[image:5.612.313.543.70.533.2]pcDNA3.1 and pcDNA3 vector, respectively. In all cultures,
expression was induced by preinfection with VacT7. As shown
in Fig. 3, in 143tk
2
cells U51 was located to the cytoplasm with
a diffuse reticulum-like pattern. Surprisingly, in
nonpermeabi-lized cells U51 was not detectable, suggesting either a very low
expression or its absence from the plasma membrane. This was
not the consequence of impaired transport of U51 due to the
heterologous HA epitope, since the untagged version of U51
(p51) also failed to be detected at the cell surface (Fig. 3c). As
expected, HSV gD and CCR5 were readily detected at the
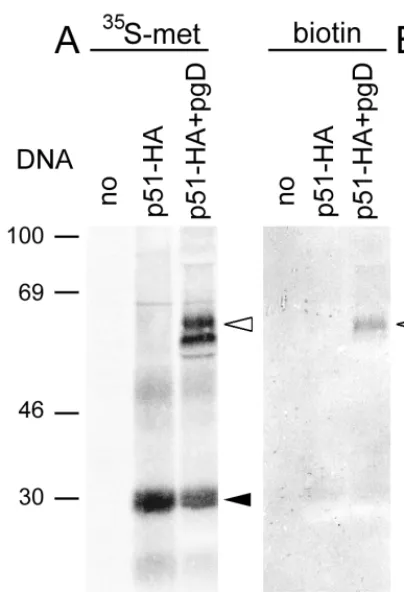
FIG. 5. Lack of cell surface expression of U51 in transfected 143tk2cellsdetected by biotinylation. VacT7-preinfected 143tk2cells were transfected with p51-HA (lanes b and e), cotransfected with p51-HA and pgD (lanes c and f), or mock transfected (lanes a and d). Cells were metabolically labeled with [35
S]me-thionine and [35S]cysteine and surface labeled with biotin immediately prior to
harvesting. U51 and gD were immunoprecipitated, separated by electrophoresis, and transferred to a nitrocellulose sheet. (A) Autoradiographic image. (B) Avi-din-peroxidase staining of biotinylated proteins. Note that U51 is detectable only as radiolabeled species (black arrowhead). gD is detectable both as radiolabeled and as biotinylated species (white arrowheads). Numbers at left show molecular mass in kilodaltons.
FIG. 6. Cell surface localization of U51 and, for comparison, of CCR5, in VacT7-preinfected T lymphocytes transfected with p51 or p51-HA. (a and b) J-Jhan cells transfected with p51 and stained with immune serum 6. (c and d) Molt-3 cells transfected with p51-HA and stained with anti-HA MAb. (e and f) Jurkat cells transfected with pCCR5 and stained with anti-CCR5 MAb. (g and h) Jurkat cells transfected with pCCR5 and stained with serum 6 and anti-HA MAb. Cells were fixed with 4% paraformaldehyde. (a, c, e, g, and h) Triton X-100-permeabilized cells. (b, d, and f) UnX-100-permeabilized cells.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.73.275.71.367.2]plasma membrane of transfected 143tk
2
cells (Fig. 3e to h).
The results with HEK-293 cells (data not shown) were
essen-tially similar to those obtained in 143tk
2
cells. A large portion
of U51 in transfected 143tk
2
cells accumulated in the ER, as
it colocalized with the ER-resident calnexin (25) in a double
immunofluorescence assay (Fig. 3i and j).
As availability of an expression system is a key prerequisite
to study the properties of U51 and its role in the HHV-6
infectious cycle, we checked for the presence of U51 at the
plasma membrane of transfected cells with higher-sensitivity
assays, specifically with fluorescence-activated flow cytometry
and with immunoprecipitation of biotinylated proteins.
By fluorescence-activated flow cytometry, expression of U51
in p51-HA-transfected HEK-293 and 143tk
2
cells was
ana-lyzed with anti-HA MAb prior to and after cell
permeabiliza-tion. Positive controls consisted of intact and permeabilized
pCCR5-transfected cells reacted with anti-CCR5 MAb. Even
by this assay, U51 could be detected in permeabilized cells
(Fig. 4d) but not in intact cells (Fig. 4b). CCR5 was readily
detected both in intact cells (Fig. 4f) and in permeabilized cells
(Fig. 4h). Figure 4 shows results with HEK-293 cells; results
with 143tk
2
cells were very similar.
In a further series of experiments, cells cotransfected with
p51 and pgD were metabolically labeled with a mixture of
[
35S]methionine and [
35S]cysteine, to detect the bulk of
pro-teins made in the cells, and with biotin immediately prior to
harvesting, to detect cell-surface-located proteins. U51 and gD
were then immunoprecipitated from the cell lysates, separated
by denaturing electrophoresis, transferred to nitrocellulose
sheets, and detected by autoradiography and avidin-peroxidase
staining. U51 was detectable only as radioactive protein and
not as biotinylated species (Fig. 5, compare lane b, panel A,
with lane e, panel B) while the control membrane protein HSV
gD was detectable both as radiolabeled and as biotinylated
species (Fig. 5, compare lane c, panel A, with lane f, panel B).
The observation that U51 could not be detected at the plasma
membrane of transfected monolayer cells even by sensitive
assays like biotinylation and fluorescence-activated flow
cytom-etry argues against a low-level cell surface expression and is
rather consistent with the lack of cell-surface-located U51 in
these cells. Accumulation of a large portion in the ER indicates
a block in transport in early compartments of the exocytic
pathway.
[image:6.612.94.254.72.213.2]U51 is expressed at the cell surface of transfected
T-lym-phocytic lines.
We investigated the possibility that expression
of U51 at the cell surface requires a cell-specific function
present in T lymphocytes. J-Jhan, Molt-3, and Jurkat cells,
preinfected with VacT7, were transfected with the two
plas-mids carrying the untagged or tagged version of U51 gene and
analyzed by fluorescence microscopy with immune serum 6 or
anti-HA-tag MAb. In contrast with the results obtained with
human monolayer cells, in all three T-lymphocytic lines U51
could be detected also in nonpermeabilized cells at the cell
surface, with a characteristic peripheral distribution pattern
almost indistinguishable from that of CCR5 transfected in
replicate cultures (typical examples are shown in Fig. 6b, d, and
f). The immunofluorescence staining was specific, as
p51-HA-or p51-transfected cells did not stain with preimmune serum 6,
anti-CCR5 MAb, or secondary antibodies alone, and
con-versely, cells transfected with pCCR5 did not stain with serum
6, nor with anti-HA MAb (Fig. 6g and h). The overall efficiency
of transfection and extent of expression were rather low, a
characteristic observed in transfections of T-lymphocytic lines.
The results indicate that in T-cell lines U51 is transported to
the cell surface and accumulates in this compartment at a level
detectable by immunofluorescence. A comparison of the
de-gree of U51 expression in transfected monolayer cells with that
FIG. 7. Immunofluorescence staining of U51 in HHV-6(B)Z29-infectedCB-MCs. Cells fixed with acetone were reacted with immune serum 6, followed by biotinylated anti-mouse antibodies and Extravidin coupled to TRITC.
FIG. 8. Synthesis of U51 in CBMCs infected with HHV-6(A)U1102 (A) or HHV-6(B)Z29 (B and C). (A and B) Detection of U51 by metabolic labeling with [35S]methionine and [35S]cysteine (35S-met). (C) Detection of cell surface expression by labeling with biotin prior to harvesting (biotin). Immunoprecipitations were
performed on lysates of infected (1) or uninfected (2) CBMCs with immune serum 6 (Immune) or with preimmune serum (Pre). Note that U51 (black arrowheads) was detectable as radiolabeled protein in lanes d and h and as biotinylated species in lane l. Numbers at left show molecular mass in kilodaltons.
on November 9, 2019 by guest
http://jvi.asm.org/
in T-lymphocytic lines revealed a brighter staining in the
monolayer cells, indicative of an overall higher level of
expres-sion (compare permeabilized cells in Fig. 3b and d with those
in Fig. 6a and c). This rules out the possibility that lack of
detection of U51 at the plasma membrane of transfected
monolayer cells was simply the consequence of an overall low
level of expression.
U51 protein is transported to the cell surface in
HHV-6-infected CBMCs.
As a preliminary assay to ascertain if U51 is
made in HHV-6-infected CBMCs, indirect
immunofluores-cence of HHV-6(A)U1102- and HHV-6(B)Z29-infected
CB-MCs or T-cell lines was performed with serum 6 by standard
methods. As only very low levels of specific fluorescence were
detected, an enhanced immunofluorescence assay was
devel-oped, in which binding of primary antibody is revealed by
biotinylated secondary antibody followed by Extravidin
cou-pled to TRITC. By this assay, U51 could be detected in
HHV-6(A)- and HHV-6(B)-infected CBMCs and localized mainly to
round cytoplasmic structures (Fig. 7). However, cell surface
expression could not be analyzed due to the overall low level of
expression. To address this latter question, we employed the
cell surface biotinylation-immunoprecipitation assay used
pre-viously for transfected monolayer cells. HHV-6(A)U1102- and
HHV-6(B)Z29-infected CBMCs were labeled with [
35S]methi-onine and [
35S]cysteine for 12 h and with biotin immediately
prior to harvesting. A 28-kDa protein could be
immunopre-cipitated specifically from HHV-6(A)U1102-infected and
HHV-6(B)Z29-infected CBMCs by immune serum 6 (Fig. 8A
and B, lanes d and h). The protein was not precipitated by the
preimmune serum (lanes b and f) and was absent from
unin-fected cells (lanes c and g), accounting for the specificity of the
precipitated protein. The apparent Mr
was slightly lower than
that of the in vitro transcription-translation U51 product, as
expected, given that the in vitro-made protein carried the HA
heterologous epitope and the polyhistidine stretch. Panel C
shows that U51 was detectable also as a biotinylated species
(lane l), demonstrating that in HHV-6-infected CBMCs it is
expressed at the cell surface [results shown for
HHV-6(B)Z29].
HHV-6(A)- and HHV-6(B)-infected CBMCs express U51
mRNA as an early gene.
As this work reports the first
identi-fication of U51 in HHV-6-infected CBMCs, it was of interest
to determine the temporal regulation of U51 gene
transcrip-tion. We checked whether U51 mRNA is expressed as an early
or a late gene by RT-PCR of RNA extracted from Molt-3 cells
infected with HHV-6(B)Z29 and maintained in the presence
or absence of PAA (a specific inhibitor of herpesvirus DNA
replication) for 24 h. U51 temporal regulation of expression
was compared to that of U31, a
b
-early gene (35), and that of
U12, a late
g
-gene (27). The results in Fig. 9 show that U51
mRNA was expressed in both PAA-treated and untreated
cells, demonstrating that it is regulated either as an
immediate-early gene or as an immediate-early gene. U31 and U12 were expressed as
early or late genes, respectively, as described elsewhere (27,
35).
b
-Actin gene expression was not affected by PAA
expo-sure, as expected. All specific amplification products were
de-tectable when the infected-cell DNA was used as template and
were not detectable when the RNA before RT was used as
template, ruling out contamination of the RNA preparations
with DNA.
DISCUSSION
U51 ORF of HHV-6 predicts a protein of 301 amino acid
residues with seven transmembrane domains (23). There are at
least three examples of sevtransmembrane proteins
en-coded by different members of the Herpesviridae family. They
contribute to the lifestyle of the specific viruses with varied
mechanisms: HHV-8 ORF74 contributes to the oncogenic
po-tential of the virus (3, 9), and HCMV US28 acts as coreceptor
for human immunodeficiency virus (42). Herpesvirus saimiri
encodes a chemokine receptor homolog, ECRF3 (37), that is
functional in signaling (1).
Here we report on the first identification of the protein
encoded by HHV-6 U51 gene and on a peculiar requirement
for a cell-specific function present in T lymphocytes for
expres-sion at the cell surface. This property was not observed
previ-ously for seven-transmembrane proteins encoded by other
her-pesviruses and, to our knowledge, by other viruses.
Both variant A and variant B HHV-6 express U51 in
acti-vated CBMCs and in T-lymphocytic lines. The temporal
reg-ulation of U51 mRNA expression differs from that of U12, as
U51 is expressed as an immediate-early or an early gene,
whereas U12 is expressed as a late gene (27). Thus, the two
seven-transmembrane proteins encoded by HHV-6 appear to
be subjected to different temporal regulations of expression. In
CBMCs, the U51 protein accumulates in fairly low amounts
and localizes mainly to cytoplasmic round structures and, to a
limited extent, to the plasma membrane.
[image:7.612.54.291.69.253.2]When transiently expressed in human monolayer HEK-293
and 143tk
2
cells, the U51 protein could not be detected at the
plasma membrane, despite the fact that it was made in
rela-tively high amounts. The protein accumulated predominantly
in the ER, as it colocalized with the ER-resident protein
cal-nexin (25), indicative of a block in early compartments of the
exocytic pathway. Lack of cell surface expression could be due
either to failure of the protein to reach the plasma membrane
or to transport to the plasma membrane followed by a very
rapid recycling to other compartments. Recycling is expected
to result in a low-level steady-state expression rather than in an
absence of protein. The sensitivity of the assays employed
argues for absence from the plasma membrane. Retention in
FIG. 9. Comparative analysis of U51, U31, U12, andb-actin temporalreg-ulation of transcription by RT-PCR in HHV-6(B)Z29-infected Molt-3 cells. In all panels, amplification of the indicated specific fragment from RNA (lanes 3 and 4), cDNA (lanes 5 to 7), and cellular DNA (lanes 1) is shown. Lanes 2, negative control of reaction lacking template (Mock). Lanes 5, amplification from cDNA retrotranscribed from the viral inoculum (i) shows that the inoculum was free of RNA. Lanes 4 and 7, nucleic acids from PAA-treated cells (1). Lanes 3 and 6, nucleic acids from cells not PAA treated (2). MW, molecular weight markers. Arrows point to the specific amplification products: U51, 515 bp; U31, 831 bp; U12, 442 and 365 bp for the unspliced and spliced forms, respectively;
b-actin, 654 bp.
on November 9, 2019 by guest
http://jvi.asm.org/
ER also argues for a defect in transport along the exocytic
pathway. In contrast with the monolayer cells, in transfected
T-cell lines U51 reached the cell surface. The results provide
evidence that intracellular trafficking of U51 is subject to a
cell-type-specific modulation and that expression at the plasma
membrane requires a function provided specifically by
acti-vated primary T lymphocytes and by T-lymphocytic lines.
Among herpesvirus-encoded proteins, gH represents a
well-known example of a membrane protein whose intracellular
trafficking to the plasma membrane requires a cooperation
with another protein. gH is conserved among all the known
members of the Herpesviridae family and plays a role in virion
infectivity and cell-to-cell spread of the viruses (16, 21, 22). gH
accumulates intracellularly in cells transfected with the single
gene but is readily transported to the plasma membrane in
infected cells or in cells cotransfected with gH and gL genes.
Heterodimer formation with gL allows gH to assume the
proper folding in the ER and to exit this compartment (26, 39).
The notable difference between gH and HHV-6 U51 is that in
the case of U51 a cellular function, rather than a viral protein,
cooperates in its intracellular trafficking. The cellular protein
which accomplishes this function for U51 protein, as well as its
mode and site of action, remains to be determined. An
inter-esting model is provided by the HCMV gB, whose
accumula-tion at the cell surface, rather than its transport to this
com-partment, appears to be regulated in a cell-type-dependent
manner. Thus, in human fibroblasts, HCMV gB is readily
de-tected at the cell surface, whereas in the human astrocytoma
cell line U373 it accumulates intracellularly. In this case, the
differential steady-state plasma membrane expression was
shown to be dependent on the state of phosphorylation of a
serine residue located in the cytoplasmic tail of the
glycopro-tein (15). Also the trafficking of HHV-6 gB appears to be
peculiar, in that in HHV-6-infected lymphocytes HHV-6 gB is
present in intracellular vesicles and vacuoles and in
intracellu-lar and extracelluintracellu-lar virions but is absent from the plasma
membrane (11). As an expression system was not developed in
that case, influence of the cell type in HHV-6 gB trafficking was
not investigated.
Current data indicate that the peculiar trafficking properties
of U51 may result in modulation of surface expression in cells
from different lineages. As HHV-6 infection in the human host
does not appear to be restricted to T lymphocytes and
mono-cytes but extends to a number of cell types as yet unidentified
(for a review, see reference 7), it is conceivable that even in
vivo the trafficking properties of U51 may result in differential
plasma membrane exposures in different cell types. Thus, in
productively infected T lymphocytes, and possibly monocytes,
U51 may be present at the plasma membrane, whereas in other
cell types it may be absent from the plasma membranes. It is
tempting to speculate that differential displays at the cell
sur-face may represent a way by which U51 activity and function
are regulated in vivo in a cell-type-dependent manner.
ACKNOWLEDGMENTS
We thank T. Baechi, University of Zurich, for confocal microscopy
analysis of transfected cells; Elisabetta Romagnoli and Giada
Frasca-roli for assistance in viral and cell cultivation and immunofluorescence
detection of U51; and A. Helenius, University of Zurich, for the gift of
anticalnexin rabbit antibody.
The work was supported by grants from the AIDS Project from
Istituto Superiore di Sanita` contract no. 40A.0.22 to Department of
Experimental Pathology-Section on Microbiology and Virology and
contract no. 30A.0.72 to Institute of Pharmacological Research “Mario
Negri;” BIOMED2 BMH4 CT95 1016 grant from UE; Target Project
in Biotechnology; MURST (40%), University of Bologna (60%); and
pluriannual plan. M.L. was a recipient of a fellowship from the Italian
Federation for Cancer Research.
REFERENCES
1. Ahuja, S. K., and P. M. Murphy. 1993. Molecular piracy of mammalian interleukin-8 receptor type B by herpesvirus saimiri. J. Biol. Chem. 268: 20691–20694.
2. Alkhatib, G., M. Locati, P. E. Kennedy, P. M. Murphy, and E. A. Berger. 1997. HIV-1 coreceptor activity of CCR5 and its inhibition by chemokines: independence from G protein signaling and importance of coreceptor down-modulation. Virology 234:340–348.
3. Arvanitakis, L., E. Geras Raaka, A. Varma, M. C. Gershengorn, and E. Cesarman.1997. Human herpesvirus KSHV encodes a constitutively active G-protein-coupled receptor linked to cell proliferation. Nature 385:347–350. 4. Bankier, A. T., S. Beck, R. Bohni, C. M. Brown, R. Cerny, M. S. Chee, C. A. D. Hutchison, T. Kouzarides, J. A. Martignetti, E. Preddie, et al.1991. The DNA sequence of the human cytomegalovirus genome. DNA Seq. 2:1–12.
5. Brandimarti, R., T. Huang, B. Roizman, and G. Campadelli Fiume. 1994. Mapping of herpes simplex virus 1 genes with mutations which overcome host restrictions to infection. Proc. Natl. Acad. Sci. USA 91:5406–5410. 6. Braun, D. K., G. Dominguez, and P. E. Pellett. 1997. Human herpesvirus 6.
Clin. Microbiol. Rev. 10:521–567.
7. Campadelli-Fiume, G., P. Mirandola, and L. Menotti. Human herpesvirus 6: an emerging pathogen. Emerg. Infect. Dis., in press.
8. Carrigan, D. R., W. R. Drobyski, S. K. Russler, M. A. Tapper, K. K. Knox, and R. C. Ash.1991. Interstitial pneumonitis associated with human herpes-virus-6 infection after marrow transplantation. Lancet 338:147–149. 9. Cesarman, E., R. G. Nador, F. Bai, R. A. Bohenzky, J. J. Russo, P. S. Moore,
Y. Chang, and D. M. Knowles.1996. Kaposi’s sarcoma-associated herpesvi-rus contains G protein-coupled receptor and cyclin D homologs which are expressed in Kaposi’s sarcoma and malignant lymphoma. J. Virol. 70:8218– 8223.
10. Challoner, P. B., K. T. Smith, J. D. Parker, D. L. MacLeod, S. N. Coulter, T. M. Rose, E. R. Schultz, J. L. Bennett, R. L. Garber, M. Chang, et al.1995. Plaque-associated expression of human herpesvirus 6 in multiple sclerosis. Proc. Natl. Acad. Sci. USA 92:7440–7444.
11. Cirone, M., G. Campadelli Fiume, L. Foa`-Tomasi, M. R. Torrisi, and A. Faggioni.1994. Human herpesvirus 6 envelope glycoproteins B and H-L complex are undetectable on the plasma membrane of infected lymphocytes. AIDS Res. Hum. Retroviruses 10:175–179.
12. Davis Poynter, N. J., D. M. Lynch, H. Vally, G. R. Shellam, W. D. Rawlinson, B. G. Barrell, and H. E. Farrell.1997. Identification and characterization of a G protein-coupled receptor homolog encoded by murine cytomegalovirus. J. Virol. 71:1521–1529.
13. Di Luca, D., P. Mirandola, T. Ravaioli, B. Bigoni, and E. Cassai. 1996. Distribution of HHV-6 variants in human tissues. Infect. Agents Dis. 5:203– 214.
14. Downing, R. G., N. Sewankambo, D. Serwadda, R. Honess, D. Crawford, R. Jarrett, and B. E. Griffin.1987. Isolation of human lymphotropic herpesvi-ruses from Uganda. Lancet ii:390.
15. Fish, K. N., C. Soderberg-Naucler, and J. A. Nelson. 1998. Steady-state plasma membrane expression of human cytomegalovirus gB is determined by the phosphorylation state of Ser900. J. Virol. 72:6657–6664.
16. Foa`-Tomasi, L., E. Avitabile, A. Boscaro, R. Brandimarti, R. Gualandri, R. Manservigi, F. Dall’Olio, F. Serafini Cessi, and G. C. Fiume.1991. Herpes simplex virus (HSV) glycoprotein H is partially processed in a cell line that expresses the glycoprotein and fully processed in cells infected with deletion or ts mutants in the known HSV glycoproteins. Virology 180:474–482. 17. Foa`-Tomasi, L., A. Boscaro, S. di Gaeta, and G. Campadelli Fiume. 1991.
Monoclonal antibodies to gp100 inhibit penetration of human herpesvirus 6 and polykaryocyte formation in susceptible cells. J. Virol. 65:4124–4129. 18. Foa`-Tomasi, L., S. Guerrini, T. Huang, and G. Campadelli-Fiume. 1992.
Characterization of human herpesvirus-6(U1102) and (GS) gp112 and iden-tification of the Z29-specified homolog. Virology 191:511–516.
19. Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss. 1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that syn-thesizes bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. USA 83:8122–8126.
20. Gao, J. L., and P. M. Murphy. 1994. Human cytomegalovirus open reading frame US28 encodes a functional beta chemokine receptor. J. Biol. Chem. 269:28539–28542.
21. Gompels, U., and A. Minson. 1986. The properties and sequence of glyco-protein H of herpes simplex virus type 1. Virology 153:230–247. 22. Gompels, U. A., and A. C. Minson. 1989. Antigenic properties and cellular
localization of herpes simplex virus glycoprotein H synthesized in a mam-malian cell expression system. J. Virol. 63:4744–4755.
23. Gompels, U. A., J. Nicholas, G. Lawrence, M. Jones, B. J. Thomson, M. E. Martin, S. Efstathiou, M. Craxton, and H. A. Macaulay.1995. The DNA sequence of human herpesvirus-6: structure, coding content, and genome evolution. Virology 209:29–51.
24. Graham, K. A., A. S. Lalani, J. L. Macen, T. L. Ness, M. Barry, L. Y. Liu, A.
on November 9, 2019 by guest
http://jvi.asm.org/
Lucas, I. Clark Lewis, R. W. Moyer, and G. McFadden.1997. The T1/35kDa family of poxvirus-secreted proteins bind chemokines and modulate leuko-cyte influx into virus-infected tissues. Virology 229:12–24.
25. Hammond, C., I. Braakman, and A. Helenius. 1994. Role of N-linked oli-gosaccharide recognition, glucose trimming, and calnexin in glycoprotein folding and quality control. Proc. Natl. Acad. Sci. USA 91:913–917. 26. Hutchinson, L., H. Browne, V. Wargent, N. Davis Poynter, S. Primorac, K.
Goldsmith, A. C. Minson, and D. C. Johnson.1992. A novel herpes simplex virus glycoprotein, gL, forms a complex with glycoprotein H (gH) and affects normal folding and surface expression of gH. J. Virol. 66:2240–2250. 27. Isegawa, Y., Z. Ping, K. Nakano, N. Sugimoto, and K. Yamanishi. 1998.
Human herpesvirus 6 open reading frame U12 encodes a functional beta-chemokine receptor. J. Virol. 72:6104–6112.
28. Kempf, W., V. Adams, N. Wey, R. Moos, M. Schmid, E. Avitabile, and G. Campadelli Fiume.1997. CD681cells of monocyte/macrophage lineage in the environment of AIDS-associated and classic-sporadic Kaposi sarcoma are singly or doubly infected with human herpesviruses 7 and 6B. Proc. Natl. Acad. Sci. USA 94:7600–7605.
29. Kolakowski, L. F., Jr. 1994. GCRDb: a G-protein-coupled receptor data-base. Receptors Channels 2:1–7.
30. Kondo, K., T. Kondo, T. Okuno, M. Takahashi, and K. Yamanishi. 1991. Latent human herpesvirus 6 infection of human monocytes/macrophages. J. Gen. Virol. 72:1401–1408.
31. Lindquester, G. J., N. Inoue, R. D. Allen, J. W. Castelli, F. R. Stamey, T. R. Dambaugh, J. J. O’Brian, R. M. Danovich, N. Frenkel, and P. E. Pellett. 1996. Restriction endonuclease mapping and molecular cloning of the hu-man herpesvirus 6 variant B strain Z29 genome. Arch. Virol. 141:367–379. 32. Lopez, C., P. Pellett, J. Stewart, C. Goldsmith, K. Sanderlin, J. Black, D. Warfield, and P. Feorino.1988. Characteristics of human herpesvirus-6. J. Infect. Dis. 157:1271–1273.
33. Lusso, P., M. S. Malnati, A. Garzino Demo, R. W. Crowley, E. O. Long, and R. C. Gallo.1993. Infection of natural killer cells by human herpesvirus 6. Nature 362:458–462.
34. Margulies, B. J., H. Browne, and W. Gibson. 1996. Identification of the human cytomegalovirus G protein-coupled receptor homologue encoded by UL33 in infected cells and enveloped virus particles. Virology 225:111–125. 35. Mirandola, P., P. Menegazzi, S. Merighi, T. Ravaioli, E. Cassai, and D. Di
Luca.1998. Temporal mapping of transcripts in herpesvirus 6 variants. J. Vi-rol. 72:3837–3844.
36. Neote, K., D. DiGregorio, J. Y. Mak, R. Horuk, and T. J. Schall. 1993. Molecular cloning, functional expression, and signaling characteristics of a C-C chemokine receptor. Cell 72:415–425.
37. Nicholas, J., K. R. Cameron, and R. W. Honess. 1992. Herpesvirus saimiri encodes homologues of G protein-coupled receptors and cyclins. Nature 355:362–365.
38. Rawlinson, W. D., H. E. Farrell, and B. G. Barrell. 1996. Analysis of the complete DNA sequence of murine cytomegalovirus. J. Virol. 70:8833–8849. 39. Roberts, S. R., M. Ponce de Leon, G. H. Cohen, and R. J. Eisenberg. 1991. Analysis of the intracellular maturation of the herpes simplex virus type 1 glycoprotein gH in infected and transfected cells. Virology 184:609–624. 40. Rost, B., R. Casadio, P. Fariselli, and C. Sander. 1995. Transmembrane
helices predicted at 95% accuracy. Protein Sci. 4:521–533.
41. Rost, B., P. Fariselli, and R. Casadio. 1996. Topology prediction for helical transmembrane proteins at 86% accuracy. Protein Sci. 5:1704–1718. 42. Rucker, J., A. L. Edinger, M. Sharron, M. Samson, B. Lee, J. F. Berson, Y.
Yi, B. Margulies, R. G. Collman, B. J. Doranz, M. Parmentier, and R. W. Doms.1997. Utilization of chemokine receptors, orphan receptors, and her-pesvirus-encoded receptors by diverse human and simian immunodeficiency viruses. J. Virol. 71:8999–9007.
43. Smith, C. A., T. D. Smith, P. J. Smolak, D. Friend, H. Hagen, M. Gerhart, L. Park, D. J. Pickup, D. Torrance, K. Mohler, K. Schooley, and R. G. Goodwin.1997. Poxvirus genomes encode a secreted, soluble protein that preferentially inhibits beta chemokine activity yet lacks sequence homology to known chemokine receptors. Virology 236:316–327.
44. Smith, G. L., J. A. Symons, A. Khanna, A. Vanderplasschen, and A. Alcami. 1997. Vaccinia virus immune evasion. Immunol. Rev. 159:137–154. 45. Walther, W., U. Stein, and C. Eder. 1994. RNA analysis using miniprep RNA
in reverse transcription PCR. BioTechniques 17:674–675.
46. Wu, C. A., and J. D. Shanley. 1998. Chronic infection of human umbilical vein endothelial cells by human herpesvirus-6. J. Gen. Virol. 79:1247–1256. 47. Yamanishi, K., T. Okuno, K. Shiraki, M. Takahashi, T. Kondo, Y. Asano, and T. Kurata.1988. Identification of human herpesvirus-6 as a causal agent for exanthem subitum. Lancet ii:1065–1067.
on November 9, 2019 by guest
http://jvi.asm.org/